
Abstract
Stem cells are subdivided into two main categories: embryonic and adult stem cells. In principle, pluripotent embryonic stem cells might differentiate in any cell types of the organism, whereas the potential of adult stem cells would be more restricted. Although adult stem cells from bone marrow have been initially the most extensively studied, those derived from human adipose tissue have been lately more widely investigated, because of several advantages. First, they can be easily obtained in large amounts from subcutaneous adipose tissue, with minimal pain and morbidity for the patients during harvesting. In addition, they feature low immunogenicity and can differentiate not only in cells of mesodermal lineage (adipocytes, osteoblasts, chondrocytes and muscle cells), but also in cells of other germ layers, such as neural or epithelial cells. As their multilineage differentiation capabilities are increasingly highlighted, their possible use in cell-based regenerative medicine is now broadly explored. In fact, starting from in vitro observations, many studies have already entered the preclinical and clinical phases. In this review, because of our main scientific interest, adipogenic, osteogenic, chondrogenic, and neurogenic differentiation abilities of adipose-derived mesenchymal stem cells, as well as their possible therapeutic applications, are chiefly focused. In addition, their ability to differentiate toward muscle, epithelial, pancreatic, and hepatic cells is briefly reported.
Introduction
S
Although adult stem cells were formerly believed to be capable of differentiating into cell lineages typical of the resident tissue, successive evidence indicate that they feature much higher degree of plasticity [1]. Studies on adult stem cells began more than 50 years ago [2], when at least two populations of stem cells were discovered in the bone marrow (BM). One population, named hematopoietic stem cells (HSCs), forms all blood cells. A second population, called bone marrow stromal cells (BMSCs) was discovered a few years later [3]. According to their mesenchymal stem cell (MSC) nature, these cells show self-renewal ability and can differentiate into adipocytes, osteoblasts, and chondrocytes, other than several other cellular lines [4 –6].
Subsequently, the existence of MSCs was verified in most adult tissues (adipose tissue, dermis, periosteum, dental tissue, salivary glands, endometrium, limb bud, menstrual and peripheral blood, synovial fluid) as well as in fetal tissues (amniotic fluid, placenta, umbilical cord blood, Wharton's jelly) [7,8].
Regardless of different procedures for isolation, characterization, and expansion, MSCs are defined by a set of minimal criteria by the international society for cellular therapy: cells must be plastic adherent when maintained in standard culture conditions; they must express surface molecules such as CD105, CD73, and CD90, and lack the expression of CD45, CD34, CD14, CD11b, CD79α or CD19, and HLA-DR; finally, they must be capable of differentiating in vitro into adipocytes, osteoblasts, and chondroblasts [9].
Within differentiated tissues, MSCs are believed to reside near or within blood vessels, in two vascular locations, the subendothelial region and the tunica adventitia, thus identifiable as MSC niches [10].
Adipose-Derived MSCs
Among the many sources of MSCs, the adipose tissue appears to be of great interest because it is available in large quantities, and can be harvested with little invasive methods such as liposuction.
Schematic representation of procedures adopted in our laboratory to isolate adipose-derived stem cells (ASCs) is reported in Fig. 1. Briefly, after collagenase digestion, lipoaspirate is centrifuged to obtain a pellet consisting of stromal vascular fraction (SVF). To expand this cell population, pellets are resuspended, seeded, and cultured in Dulbecco's modified Eagle's medium (DMEM) containing fetal bovine serum (FBS), penicillin/streptomycin, glutamine, and mesenchymal stem cell growth supplement. After 2–4 passages, adherent cells show a quite homogeneous fibroblast-like morphology, typical of ASCs. Their stem nature is routinely verified by the expression of MSC markers, using immunocytochemical and/or flow cytometry techniques.
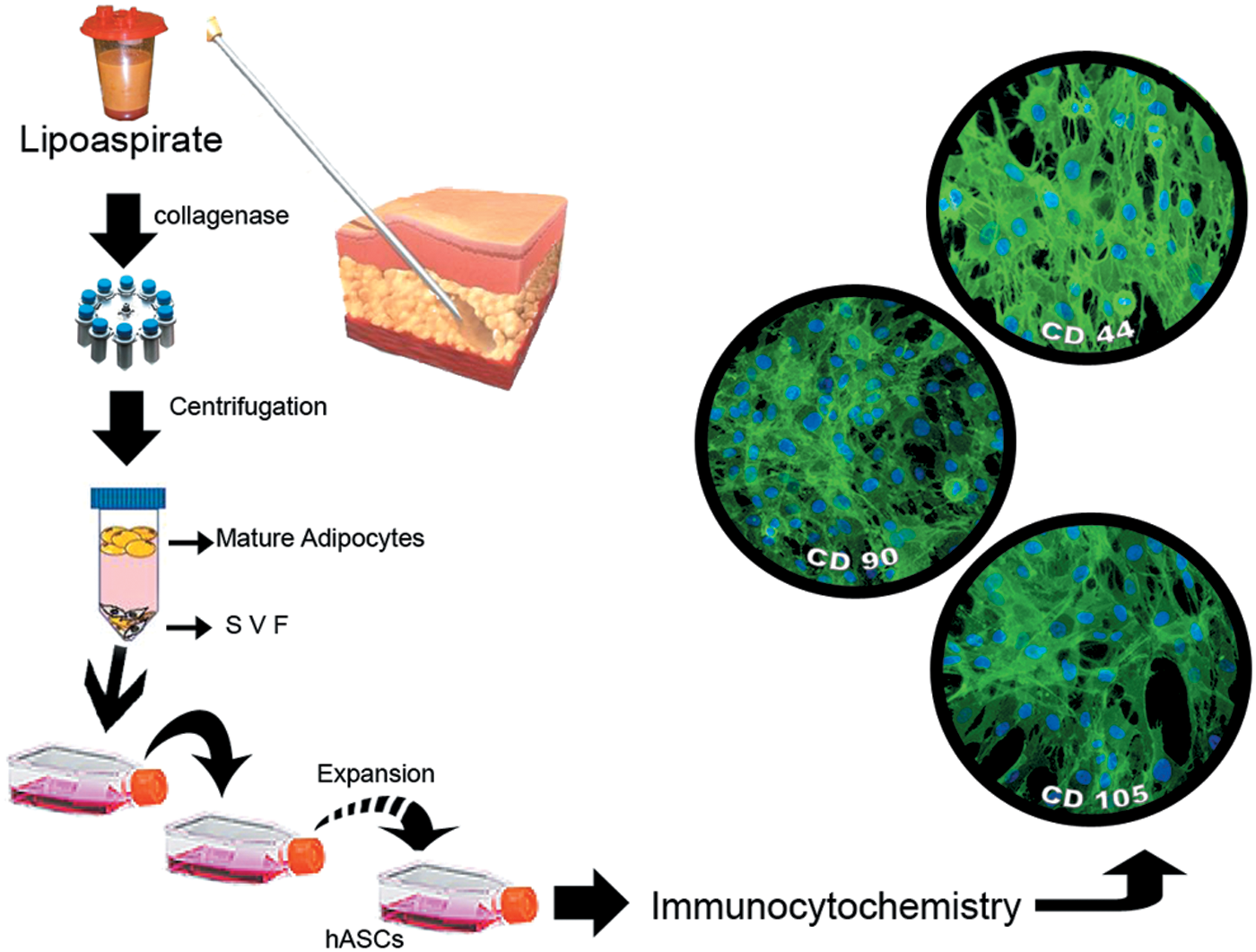
Schematic representation of human adipose tissue processing to isolate hASCs. Adipose tissue obtained by liposuction, after collagenase digestion, is centrifuged. Subsequently, buoyant mature adipocytes and supernatant are removed and the SVF at the bottom is cultured using appropriate culture medium. After expansion, the stem nature of adherent fibroblast-like cells is revealed by their immunopositive reactions to specific markers. Microphotographs in the right show their immunopositivity to CD44, CD90, CD105, as shown by green fluorescence. Blue dots denote DAPI staining of nuclei. hASCs, human adipose-derived mesenchymal stem cells; SVF, stromal vascular fraction. Color images available online at
The use of animal-derived reagents for in vitro cell culture techniques should be avoided in clinical trials because of safety reasons; xeno- and serum-free reagents have been developed for isolation and cell expansion of ASCs [11]. Many studies have demonstrated that both ASCs and BMSCs feature low immunogenicity, express similar immunophenotype, and can differentiate into adipocytes, osteoblasts, and chondrocytes [12,13]. As effective as BMSCs for research and clinical applications, ASCs offer some relevant advantages. The yield of MSCs from adipose tissue is much higher, the age of donors is almost irrelevant, they feature higher proliferation rate [14,15], and can be cultured for more numerous passages. MSCs from human adipose tissue were first isolated in 1976 [16], but they were well identified and characterized by Zuk et al. in 2001 [17].
As already mentioned, in addition to expression of MSC typical cell surface markers, the ability to differentiate into mesodermal lineage cells is also one of the determining properties of ASCs. Differentiation into adipocytes, osteocytes, and chondrocytes can be confirmed by immunocytochemical, histochemical, and PCR analysis, by the presence of oil droplet, mineralized matrix formation, and expression of type II collagen, respectively. In addition, several studies demonstrate that ASCs can transdifferentiate in many other cells of endodermal and ectodermal lineages [7,18]. Some examples of human ASCs (hASCs), differentiated in our laboratory, are illustrated in Fig. 2.
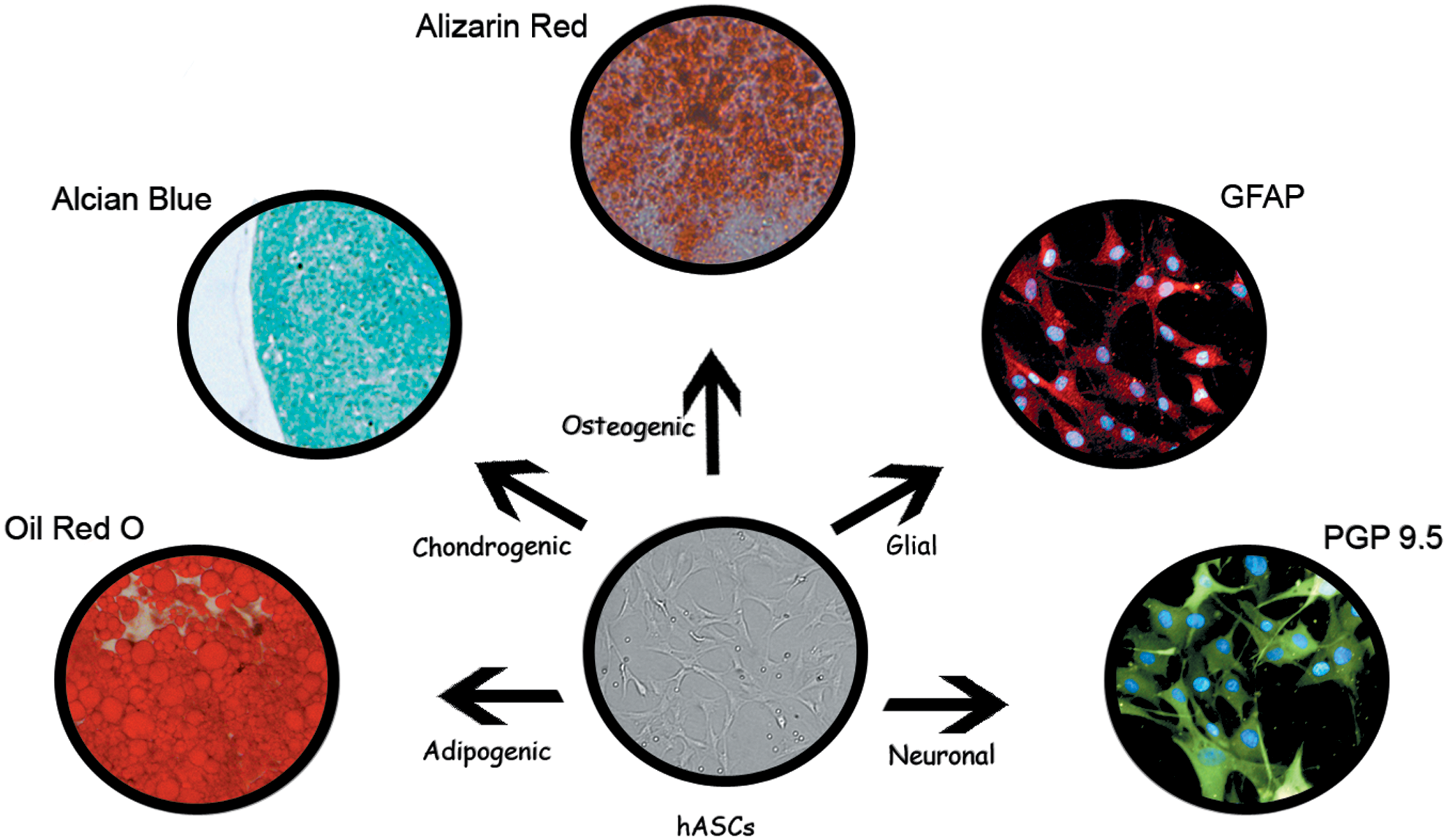
hASCs are normally cultured in various induction media for their differentiation. The adipogenic, chondrogenic, and osteogenic differentiation can be verified by Oil Red O, Alcian Blue, and Alizarin Red staining, respectively. Immunopositivity for GFAP (red cytoplasmic fluorescence) and PGP 9.5 (green cytoplasmic fluorescence) is indicative for glial or neuronal differentiation; blue dots denote DAPI staining of nuclei. GFAP, glial fibrillary acidic protein; PGP 9.5, protein gene product 9.5. Color images available online at
Adipogenic Differentiation
Differentiation of ASCs into adipocytes can be induced in vitro by culture media opportunely supplemented with various molecules able to activate adipogenic transcription factors (Table 1). For this purpose, ASCs are commonly cultured for 3–4 weeks in growth medium supplemented with dexamethasone, indomethacin, insulin, and isobutyl methyl xanthine [17,19]. Other induction factors have been reported, such as 3,5,3-triiodothyronine, transferrin, calcium pantothenate, and biotin [20]. Increased adipogenesis was observed when dexamethasone and isobutyl methyl xanthine were replaced by dehydroepiandrosterone and histamine in the differentiation cocktail [21].
ASCs, adipose-derived mesenchymal stem cells; DMEM, Dulbecco's modified Eagle's medium; FBS, fetal bovine serum; TGF, transforming growth factor.
Moreover, adipogenic differentiation may be efficiently enhanced by the addition of linoleic, palmitic, and oleic fatty acids [22]. Some other supplements can also be used, such as agonists of peroxisome proliferator-activated receptor γ (PPARγ) or 5% rabbit serum. Better results have been observed by alternating every 3 days the induction medium with the maintenance medium, until mature adipocytes are obtained [15]. During adipogenic differentiation, ASCs would acquire more resistance to apoptosis [23]. In fact, the expression of apoptotic markers (p53, bax, PDCD4, and PTEN) are significantly downregulated in a time-dependent manner as differentiation progresses.
Adipose cells so differentiated are commonly identified by cytoplasmic accumulation of lipid droplets (usually revealed by Oil Red O histochemical stain) as well as the expression of adipocyte-specific markers, such as PPARγ, adipocyte protein 2 (ap2), fatty acid-binding protein 4 (FABP4), acetyl-CoA carboxylase (ACC), and lipoprotein lipase (LPL). In addition, lipolytic response and adipokine (adiponectin, leptin) secretion have been observed upon adrenergic stimulation [24,25].
Recent in vitro and in vivo studies report a significant increased adipogenic differentiation for hASCs in the presence of basic fibroblast growth factor (bFGF). In particular, when cultured in the presence of bFGF, the number of ASCs increased by more than 1,000-fold. Moreover, larger cross-sectional areas of newly formed adipose tissue were observed by implanting into the back of nude mice a collagen/gelatin scaffold containing ASCs and bFGF [20,26]. According to the authors, these effects result not only by differentiation of implanted progenitor cells, but also by novel induction of local adipogenesis.
Potential therapeutic applications
Therapeutic applications of ASCs have been particularly explored in the last decade in plastic and reconstructive surgery to improve the engraftment and survival of transplanted autologous adipose tissue, often in cases of breast reconstruction and skin rejuvenation. In fact, early techniques of adipose tissue transplantation frequently showed significant limitations, because of reduced survival time and significant reabsorption of the transplanted tissue. ASCs and their paracrine activities can improve survival of the transplanted fat by secreting a large number of cytokine and growth factors, which enhance vessel density and tissue regeneration [27].
Various strategies have been developed to obtain a fat graft as rich as possible of ASCs. One method consists of two separated surgical procedures. The fat tissue is first harvested by liposuction and then treated to expand ASCs. Two weeks later, cultured ASCs are combined with fresh lipoaspirate before transplantation [28]. An alternative strategy, called cell-assisted lipotransfer, was developed [29]. By this method, in a single surgical sitting, a part of lipoaspirate is used to obtain fresh SVF, which is added to the other portion of untreated lipoaspirate before reinjection. Positive results have been reported by the authors, as for the survival time and the residual volume of fat tissue transplanted.
Other than useful for therapeutic applications, data on mechanisms regulating adipogenesis can be exploited in studies about obesity, by testing lipolytic or antiadipogenic molecules during differentiation of ASCs into adipocytes. For example, activation of A2b adenosine receptor is able to inhibit differentiation of mouse and human preadipocytes into adipocytes [30]. The addition of Citrus bergamia extract in the induction medium reduces the accumulation of intracellular lipids and promote lipolysis, by activating lipolytic genes and repressing the expression of adipogenic ones [31]. Treatment with Caffeic Acid Phenethyl Ester (Cape), isolated from propolis, not only prevents adipogenesis, but also regulates the insulin resistance of differentiated adipocytes extensively exposed to high glucose [32].
Osteogenic Differentiation
The osteogenic induction of ASCs is usually obtained by supplementing culture medium with ascorbic acid, β-glyceralphosphate, dexamethasone, and 1,25 dihydroxy vitamin D(3) [24,33] (Table 1). During differentiation, cultured cells mineralize their extracellular matrix, as indicated by the presence of increasing calcium phosphate deposits, revealed by Alizarin Red and von Kossa histochemical stains. Consistently, matrix mineralization is paralleled by increased alkaline phosphatase activity and osteogenic gene expression, such as Smads, type II bone morphogenetic protein receptor (BMPR-II), osteonectin, osteocalcin, and runt-related transcription factor 2 (Runx2) [34,35].
Although some studies report that the osteogenic capacity of ASCs is lower than that of BMSCs, [36], some comparative studies indicate that ASCs may feature a greater osteogenic potential [15,37]. Indeed, hASC-derived osteocytes were characterized by greater amounts of calcium stores within the cytoplasm and were able to form larger number and size of calcium stores in the matrix. In any case, the easier accessibility of ASCs represents a considerable advantage to stimulate further investigations about the clinical use of these cells in bone regeneration. More recently, osteogenic differentiation of hASCs has been tested in collagen/Mg-doped hydroxyapatite scaffolds, in the presence or absence of osteogenic-inducing factors in the culture medium. Although hASCs were able to differentiate into mature osteoblasts even in the basal growth medium, the presence of osteoinductive factors markedly improved the osteogenic process, as confirmed by the enhanced expression of osterix, osteopontin, osteocalcin, and osteonectin [38].
The existence of a converse relationship between adipogenesis and osteogenesis has been found during culturing MSCs, including ASCs, with different media supplements [39,40]. Signaling pathways involving Hedgehog [41,42], NEL-like protein 1 (NELL-1) [43], and β-catenin-dependent Wnt [44,45] are well known for their proosteogenic and antiadipogenic effects on MSCs differentiation. Similar effects are exerted by compounds, such as alendronate, which promotes osteogenesis and inhibits adipogenesis [46]. On the contrary, many agents stimulating adipogenesis, such as the thiazolidinedione PPARγ ligands, inhibit osteogenesis. In agreement, clinical studies show that the bone loss in osteoporosis is paralleled by increased marrow fat reserves [47]. However, there are various signaling cascades able to induce positive regulation of both adipogenesis and osteogenesis. Indeed, although bone morphogenetic proteins (BMPs) promote osteogenic differentiation, they can also promote proadipogenic effects [7]. Thus, the complex switch between adipocyte and osteoblast lineage deserves further investigations.
Potential therapeutic applications
In a preclinical study, ASC-seeded scaffolds were implanted in goats with induced osteochondral defects at the level of the knee [48]. After 4 months, significant improvements were observed in cartilage and subchondral bone regeneration of damaged regions. Successful administration of ASCs has been described in two case reports of patients with hip osteonecrosis [49]. In both cases, improvement of symptoms have been observed, likely associated to bone regeneration after percutaneous injections of a combination of autologous ASCs, hyaluronic acid, platelet-rich plasma (PRP), and calcium chloride. Many efforts are being made to optimize surgery protocols and healing time for treatments of craniofacial defects, in cases in bone nonunions from accidents and delayed fracture healing [36].
Chondrogenic Differentiation
For chondrogenic differentiation (Table 1), ASCs can be cultured in DMEM supplemented with insulin–transferrin–selenium (ITS), linoleic acid, pyruvate, ascorbate 2-phosphate, dexamethasone, and transforming growth factor-β III (TGF-βIII). In association with TGF-β, other growth factors, such as insulin-like growth factor-I (IGF-I) and BMP-2, seem to induce the differentiation of ASCs into chondrocytes [50]. Furthermore, the addition of BMP-6 to the differentiation cocktail strongly enhances ASCs chondrogenesis [51 –53]. In early phases of the induction, cultured ASCs evolve in prechondrocytes, which express type I and type II collagens, other than various adhesion molecules [54]. In the final steps, prechondrocytes differentiate into mature chondrocytes and express chondrogenic transcription factors like Sox9, L-Sox5, and Sox6 [55].
Besides growth factors, some hormones such as parathormone-related peptide (PTHrp) improve chondrogenesis [56]. In a coculture study, it has been reported that mature chondrocytes can provide cellular factors in the microenvironment, able to drive chondrogenic differentiation of ASCs [57].
Unlike other differentiation lineages, for chondrogenesis, ASCs are commonly maintained in a three-dimensional (3D) configuration. For example, cultures have been carried out as pellet or micromass [15,58], or within hydrogel scaffolds of agarose or alginate [59]. Although initial evidence indicated equal [60,61] or higher chondrogenic potential for BMSCs than ASCs [62], some authors reported opposite results [63]. These discrepancies might simply depend on supplements used, because former differentiation media were mainly designed for BMSCs.
In a comparative study between ASCs and BMSCs [15], it has been observed that ASCs exhibit a stronger Alcian Blue staining already at 7 days of cultures, so indicating the formation of higher amounts of glycosaminoglycans in the extracellular matrix. This trend was confirmed also after 14, 21, and 28 days, when Alcian Blue staining progressively increases. In a comparative study between differentiated ASCs in 3D clusters and monolayer cultures of mature chondrocytes, differentiated ASCs were able to produce collagen type I, II, and lubricin, whereas mature chondrocytes only express collagen type I [58]. Since the highest expression of collagen type II and lubricin was detected after 21 days of culture, it is concluded that ASCs at 21 days of chondrogenic differentiation may represent the best choice to obtain functional chondrocytes [64]. This finding could be relevant for future clinical approaches, in cases of cell-based therapies for degenerative pathologies of articular cartilage such as osteoarthritis.
Potential therapeutic applications
In clinical trials on patients affected by knee osteoarthritis, cartilage regeneration was induced after percutaneous injection of a combination of autologous ASCs, hyaluronic acid, PRP, and calcium chloride with addition of a very low dose of dexamethasone [49]. In a study of phase I/II clinical trial, different doses of ASCs were injected into osteoarthritic knee of three groups of patients. Best results were observed following intra-articular injection of the highest dose [65]. After 6 months, joint function and pain were ameliorated without side effects and cartilage defects were reduced by regeneration of hyaline-like articular cartilage.
Myogenic Differentiation
Cells of muscular lineage include cardiac, skeletal, and smooth muscle cells. Any of them may undergo degenerative diseases that significantly impair the quality of life of patients. Heart transplantation is currently the only treatment option in case of severe ischemic heart disease. Skeletal muscle dystrophies have no cure and the possibility to replace damaged muscles is not realistic at the present. Impairment of smooth muscle activity may have important effects on gastrointestinal peristaltic movements, blood pressure regulation, or airways ventilation. In this scenery, a stem cell-based therapy may represent a most valuable tool.
Cardiac muscle
Differentiation toward cardiomyocytes has been formerly induced, both in vitro and in vivo, for BMSCs cultured with 5-azacytidine (5-AZA) [66]. Later, other induction media have been successfully designed in the absence of 5-AZA, also for ASCs. According to the study of Planat-Benard [67], murine ASCs cultured in semisolid methylcellulose medium, also without 5-AZA, were able to transdifferentiate in electrophysiologically active beating cells, also expressing cardiac-specific markers, such as transcription factors GATA-4 and Nkx2.5.
In a comparative study, differentiation of ASCs into cardiomyocytes has been carried out adopting three protocols (standard 5-AZA protocol, straight Planat-Benard protocol, and Planat-Benard protocol +5-AZA). In all cases, positive results have been obtained for the expression of sarcomeric alpha-actinin protein and connexin-43. Overall, 5-AZA protocol seems the best choice, being the simplest, fastest, and lowest cost [68]. In particular, CD73+ ASCs show the best ability to differentiate toward cardiomyocytes [69].
Implantation of ASCs in animal models of myocardial infarction can improve cardiac function [70,71]. Longer-term improvements and cell engraftment could be achieved if ASCs are implanted through a carrier collagen sheet [72]. Main positive cardioprotective and angiogenetic effects would be induced by paracrine properties, whereas cellular differentiation of ASCs would contribute to a lesser extent [70].
Skeletal muscle
Several culture media have been tested to induce ASCs and BMSCs differentiation toward skeletal muscle cells: MSC growth medium (MSCGM), MSCGM +5-AZA, skeletal muscle myoblast cell growth medium (SkGM)-2 BulletKit, and other three culture media containing 5%, 30%, and 50% of human satellite cell-conditioned medium, respectively. The highest proliferation rate has been observed when ASCs and BMSCs are cultured with SkGM-2 BulletKit medium, whereas the strongest myogenic differentiation was induced by the 30% conditioned culture medium. Notably, for all media, ASCs showed the highest proliferation rate and the strongest expression of myogenic markers, such as skeletal muscle actin-α1, myosin heavy chain (MHC)1, MHC 8, and desmin [73].
In a coculture system of hASCs and murine C2C12 myoblasts, ASC potential myogenic differentiation was proven by their tendency to fusion and expression of human sarcomeric proteins [74].
Intravenously injected in SJL mice without immunosuppression, hASCs were able to engraft into the host muscle. They also expressed human muscle proteins, and improved the motor performance of these animals [75]. Still without immunosuppression, hASCs were locally and intravenously injected into golden retriever dog after inducing muscular dystrophy similar to human Duchenne muscular dystrophy. After 6 months from injections, ASCs were able to reach and engraft into the host dystrophic muscle, and expressed human dystrophin [76].
Smooth muscle
Although some authors describe cases of spontaneous differentiation of ASCs in a smooth muscle phenotype in basal growth media, better results can be obtained using induction supplements [77]. Using MCDB I31 medium containing 1% FBS and 100 U/mL heparin, alpha smooth muscle actin and smoothelin were expressed after 3 weeks of differentiation. At 6 weeks of culture, also MHC was present [78]. Their differentiation into functional smooth muscle cells certainly represents a useful source of autologous cells for reconstruction of damaged organs and tissues containing smooth muscle [79].
Periurethral injection of autologous adipose-derived regenerative cells was carried out in patients with persistent stress urinary incontinence after prostate surgery. After 1 year, symptoms and quality of life improved progressively in most patients, with no significant adverse events [80].
Neural Differentiation
Neural differentiation of ASCs has been reported in numerous studies, by using a variety of protocols. Examples of induction media, supplements utilized, and results obtained are summarized in Table 2.
BDNF, brain-derived neurotrophic factor; EGF, epidermal growth factor; bFGF, basic fibroblast growth factor; GAP-43, growth associated protein-43; GFAP, glial fibrillary acidic protein; MAP2, microtubule-associated protein 2; MSCs, mesenchymal stem cells; NeuN, neuronal-specific nuclei protein; NSE, neural-specific enolase; PGP 9.5, protein gene product 9.5; PMP-22, peripheral myelin protein 22; PSA-NCAM, polysialic acid–neural cell adhesion molecule; Trk-A, tropomyosin receptor kinase A.
In 2002, Safford et al. [81] were the first to induce ASCs into neural phenotype, by using a medium containing valproic acid, butylated hydroxyanisole, insulin, and hydrocortisone. The phenotype of ASCs changed toward neuronal morphology already after 1–3 h. The neuronal differentiation, indicated by the expression of intermediate filament M, nestin, and neuronal-specific nuclei protein (NeuN), was enhanced by pretreatment with epidermal growth factor (EGF) and bFGF. However, no functional tests were carried out, and cells died within 14 days of culture. In the same year, Zuk et al. [82] were able to differentiate ASCs toward neural-like cells. Again, a two-step procedure was adopted, a preinduction medium for 24 h and then a neurogenic medium for up to 9 h. At the end of the treatment, cells revealed the presence of nestin, neural-specific enolase (NSE), and NeuN.
In 2003, Ashjian et al. [83] reported the differentiation of ASCs into early neural progenitors, as indicated by the expression of NSE, Tropomyosin receptor kinase A (TrkA), and NeuN. However, these cells could not be considered as functional neurons since they were not capable of depolarization and repolarization in patch clamp tests.
Two protocols were adopted in another study to evaluate neural differentiation of ASCs and MSCs from BM, spleen, and thymus [84]. In the first protocol, by using a specific induction medium, three populations were recognized, regardless of the different origins of MSCs. The most numerous population (50%–70%) was morphologically similar to neurons or astrocytes and showed intense expression of Peripheral Myelin Protein 22 (PMP-22), Glial Fibrillary Acidic Protein (GFAP), NeuN, polysialic acid–neural cell adhesion molecule (PSA-NCAM), nestin, S-100, microtubule-associated protein 2 (MAP2), and galactocerebroside (Gal-C). The second population (about 15%) was similar to cells of the oligodendroglial lineage, and selectively expressed Gal-C. The last population (15%–20%) remained undifferentiated. However, cells gradually recovered their basal morphology and phenotype within 48–72 h of culture with basal medium.
In the second protocol, PKH-67-labeled MSCs were cocultured with human Schwann cells. After 7 days, almost one third of ASCs showed the phenotype of Schwann cells (PMP-22, S-100 protein), whereas significantly lower proportions were found for the other MSCs. Percentages of PKH-67-labeled MSCs from spleen, BM, and thymus increased only after pretreatment with the neural differentiation medium.
The neuronal differentiation of ASCs was also tested by inducing sphere formation in serum-free medium with bFGF and EGF [85]. After sphere dissociation, culture medium was supplemented with brain-derived neurotrophic factor (BDNF) and retinoic acid. As a consequence, about half of ASCs showed morphological, immunocytochemical, and electrophysiological evidence of initial neuronal differentiation. They expressed Nestin, MAP2, and NeuN. In functional tests, differentiated cells featured negative membrane potential (−60 mV), delayed rectifier potassium currents, and TTX-sensitive sodium currents. Such changes were stable for at least 7 days after removal of differentiation medium.
By addition of PRP to neural induction medium, after 2 weeks of culture, differentiated ASCs displayed higher level of NSE, MAP2, growth-associated protein-43 (GAP-43), neural cell adhesion molecule (NCAM), and synapsin 1 [86]. Also estradiol (E2) addition to a neural induction medium may have positive effects, as revealed by significantly higher expression of nestin and MAP2 [87]. Differentiation of ASCs toward the neuronal lineage was also induced using conditioned media obtained from cultured rat olfactory ensheathing cells (OECs) or human B104 neuroblastoma cells [88]. In both cases, ASCs acquire morphological features of neuronal-like cells and, in a time-dependent manner, they express nestin, protein gene product 9.5 (PGP 9.5), and MAP2. Overall, OEC conditioning medium has proven more efficient.
Potential therapeutic applications
Neural differentiation of ASCs has been positively utilized in regenerative therapies of lesions in the peripheral nervous system. In rats with sciatic injuries, previously differentiated ASCs were implanted through silastic conduits that bridged a 13 mm nerve gap [89]. After 4 months, a significantly improved functional recovery was obtained, as attested by anatomical and functional tests. When hASCs were implanted in the proximal and distal stumps of a sectioned rat sciatic nerve, fewer cells were lost in the lumbar ganglia, less prominent amputation neuroma was observed, and lower level of inflammatory activity was induced [90].
Results obtained in in vivo studies with animal models have shown that ASCs transplanted into the brain may be useful in the regeneration of the central nervous system, for example, in cases of neurodegenerative diseases or stroke. In a mouse model of Alzheimer's disease, intravenous or intracerebral injection of ASCs significantly improved learning and memory and restored neuropathology, including amyloid beta deposition [91]
Intraventricular injection of ASCs in a rat model of intracerebral hemorrhage induced their differentiation into neuron-like and astrocyte-like cells around the hematoma, upregulation of vascular endothelial growth factor (VEGF) expression, and improvement of neural function [92]. In conclusion, although the mechanisms involved are largely unknown, ASCs might be used to replenish lost cells, promote neuronal regeneration, protect neuronal survival, and play a role in recovering neurological function [93].
Epithelial Differentiation
Among epithelial differentiations of ASC, those concerning skin and hairs are probably the most investigated.
Skin
Autologous skin grafting is the main reconstructive procedure for cutaneous injuries, although often challenged by donor-related issues and limited results from aesthetic or functional point of view. In spite of advances in tissue engineering technology, the use of skin substitute often is not satisfactory because of microbial contamination and unsatisfactory mechanical and cosmetic outcomes [94].
In these cases, stem cell therapy may represent a better tool. Transdifferentiation of hASCs in keratinocytes has been reported in a coculture study [95] of hASCs with keratinocytes using CnT-57, a serum-free medium containing bovine pituitary extract, FGF-1, and endothelial growth factor. Differentiated hASCs were characterized by the increased expression of keratin 5, involucrin, and filaggrin. Differentiation was more efficiently achieved when ASCs were cultured in a fibrin-based matrix containing molecules normally involved in wound healing. In this case, a significantly higher number of positive cells was found for cytokeratin (CK) markers such as CK14, CK5, and CK19. Besides, this fibrin-based matrix might also be used as vehicle for cell transplantation [96].
In a preclinical study, intradermal injections of allogeneic ASCs, autologous ASCs, and autologous BMSCs were compared on cutaneous wound repair in rabbits. Best results were observed for autologous ASC-treated wounds, which showed a more advanced re-epithelialization and a perfectly differentiated epidermis after 21 days [97]. The treatment with cryopreserved ASCs shortened the time needed for complete healing of wounds in nude mice. In fact, whereas 21.2 days were needed for spontaneous healing of mouse skin, 17.4 days were necessary when treated with the vehicle alone and only 14.6 days when ASCs were added. Moreover, blood perfusion of the healed wound was significantly improved in ASC-treated mice compared with the control group [98].
Hair
Several disorders, including genetic, hormonal, traumatic, and iatrogenic events, may cause hair loss. Although drug therapies, hair transplantation, or dietary supplementation are commonly used, they often result in very few, if any, positive effects. Therefore, cell-based alternative therapies deserve to be explored [99,100]. Hair cycle progression is largely determined by cellular interactions and signaling pathways in the follicle environment [101]. Since adipocytes and/or their secretomes seem to play an important role, many investigations have been carried out to test the effects of ASCs and their paracrine activities [102]. In fact, ASCs would provide a number of regenerative growth factors [VEGF, hepatocyte growth factor (HGF), IGF, platelet-derived growth factor (PDGF)] that might contribute to hair morphogenesis and regeneration [100,103].
Beneficial effects of ASC-conditioned media (ASC-CM) on hair growth have been described by Won et al. [104], both in vitro and in vivo studies. Encouraging results have been observed in clinical studies by intradermal injections of ASC-CM [99]. Application of ASC-CM is found effective to treat female pattern hair loss [100]. After 12 weeks of therapy, both hair density and thickness significantly increased, without side effects. Various culture strategies have been tested to obtain ASC-CM as effective as possible. For example, enhanced hair growth might be obtained by using ASC-CM from hypoxic cultures. The greater efficiency would be due to higher content of several growth factors in the CM [105]. Also, low-dose ultraviolet B preconditioning might improve results, by increasing concentration of growth factors and generation of reactive oxygen species in ASC-CM [106].
Pancreatic Differentiation
Biliary tree and pancreatic ducts contain progenitor cells able to differentiate to mature liver and pancreatic cells, but they often are not sufficient to repair damages, so other interventions are needed. Although allogeneic pancreas or islet transplantation remains the gold standard for severe cases of diabetes, the shortage of donors induces to consider alternative strategies [107].
A valuable alternative option for β-cell replacement might then be represented by implantation of opportunely differentiated stem cells capable to recognize glucose and secrete insulin [108,109]. ASCs have been successfully differentiated in endocrine pancreatic-like cells when cultured in serum-free medium supplemented with nicotinamide, activin, extendin, HGF, and pentagastrin. Differentiated ASCs expressed mRNAs for glucagon, insulin, and somatostatin, as well as the transcription factors isl-1, Ipf-1, Ngn-3, and Pax-6, critical for β-cell development [110].
Potential therapeutic applications
In a preclinical study, aggregates of ASCs differentiated into pancreatic islet-like cells were encapsulated in calcium alginate and transplanted into streptozotocin-induced diabetic mice. By this procedure, mice restored normoglycemia within 2 weeks [111]. In patients with type 1 diabetes, cotransplantation of ASC-derived insulin-secreting islets and HSCs reduced exogenous insulin requirement, increased c-peptide levels, and prevented ketoacidosis [112]. Similarly, coinfusion of ASC-derived insulin-producing cells and HSCs was able to normalize glycemia [113].
Hepatic Differentiation
Although liver transplantation is currently the best option for patients with liver cirrhosis, therapeutic possibilities are often impaired by organ shortage and graft failure or rejection. Therefore, stem cell therapy might represent a promising alternative.
hASCs can be successfully transdifferentiated into hepatocyte by using differentiation culture media in addition to various supplements such as EGF, bFGF, HGF, nicotinamide, oncostatin M (OSM), dexamethasone, and ITS [114,115]. The addition of liver extracts would promote the hepatic differentiation of ASCs to an even greater extent than chemicals [116]. Like BMSCs, also differentiated ASCs express biochemical characteristics of hepatocytes. In fact, they show albumin, α-fetoprotein (α-FP), and urea production [117]. Although ASCs and BMSCs display similar hepatogenic differentiation in vitro, longer survival periods and higher proliferation capacity were observed for ASCs. In particular, CD105+ ASCs exhibit a higher hepatic differentiation capability than other ASCs [118].
Better hepatocyte-like morphology and expression of specific markers were observed for hASCs cultured in serum-free differentiation medium. These differentiated cells were able to store glycogen and uptake low-density lipoprotein (LDL). Analogously to human liver cells, albumin production and LDL uptake were enhanced by trichostatin A [119].
Potential therapeutic applications
Studies in animal models demonstrate that hASCs can significantly restore liver function. In rat models, differentiated hepatocyte-like cells secreting albumin- and α-FP were detectable after 1 week from hASCs direct injections in damaged livers. Total bilirubin, prothrombin time, and albumin levels confirmed appreciable recovery of liver function. Also liver fibrosis and inflammatory activity decreased significantly [120]. In another investigation, transplanted ASCs could be found in hepatic histological sections up to 12 weeks after surgery. At the same time, blood levels of albumin, total protein, glutamic oxaloacetic transaminase and lactate dehydrogenase were significantly increased [121].
The intravenous administration of undifferentiated ASCs in carbon tetrachloride-injured mice has revealed that, 1 month after the transplantation, human albumin-positive ASCs could be identified in damaged livers [119]. In a recent study, hASCs were transplanted into spleens of rats with induced acute liver failure (ALF). Although most transplanted cells remained in the spleen and only a few cells migrated to the liver, ALF was attenuated [122]. Noticeable improvements in survival of ALF rats was observed after intravenous injection of ASC-CM and ASC lysates, containing high levels of HGF and VEGF. Consistently, intravenous injection of ASCs or ASC secretomes in partially hepatectomized mice improved liver regeneration and albumin expression [123]. Altogether, this cell-free therapy, based on the paracrine properties of stem cells, can provide novel treatments for liver diseases, eluding limitations of cell-based strategies.
Adipose-Derived MSCs as Carrier for Gene Delivery
Based on their immunomodulatory and immunosuppressive effects upon transplantation, their homing ability to damaged tissue, tumor, and metastases, ASCs seem ideally suited as vehicles for gene delivery [124]. In fact, the combination of stem cell therapy and gene therapy has recently been shown to improve a number of therapeutic effects, such as bone formation or wound healing [125]. Encouraging results have been obtained in cancer therapy for their ability to selectively target tumor cells [126]. In vivo experiments in nude mice showed that hASCs transduced with yeast cytosine deaminase can significantly inhibit the growth of human colon adenocarcinoma cells HT-29 [127].
In a preclinical study, coinjections of human bone metastatic prostate cancer cells and transduced ASCs in nude mice treated with 5-fluorocytosine induced a complete tumor regression in a dose-dependent manner or did not even allow the formation of the tumor. Moreover, it was demonstrated that intravenous administration of the therapeutic cells significantly inhibited prostate tumor growth [128]. However, since genetic modification using lentiviruses or retroviruses raises safety concerns for the potential risk of tumor formation, development of safer and more efficient vectors represents a great challenge. Adenoviral vectors may be a safer alternative, as well as nonviral vectors, characterized by lower immunogenicity, increased biocompatibility, and reduced cytotoxicity [129].
Conclusions
Scientific investigations in humans and in animal models, by in vivo or in vitro methodologies, have shown the enormous potential of ASCs in cell-based regenerative medicine. In this field, they offer numerous advantages. First of all, ASCs can be easily obtained in large amount from subcutaneous adipose tissue and easily accessible by liposuction. For their multilineage differentiation ability, ASCs may be used in a variety of human acute and chronic diseases, such diabetes, cardiovascular disorders, neurodegenerative diseases, osteoarthritis, and many others. Since paracrine activity of ASCs is likely responsible for at least some of their beneficial effects, the use of cell-free conditioned medium, which mimic effects of cell transplant, might be successfully employed. In this case, therapeutic applications could be definitely easier.
For their low immunogenicity and putative immunosuppressive effects, ASCs might be utilized for allogeneic transplantation. This represents a most valuable tool since, when appropriately cryopreserved and stored, as it occurs for hematopoietic stem cells, they would be immediately ready for use, avoiding the necessary steps of fat harvesting and processing to obtain autologous ASCs.
Although a large number of clinical trials using ASCs have been already successfully performed in the last decade, in many cases their clinical application is restricted at present for safety reasons. Indeed, more work is needed to verify and standardize the procedures for obtaining, expanding, storing, and using these cells. Hopefully, the worldwide research will be able, in the next few years, to approach the gateway from bench to bedside for more numerous therapeutic applications. In this context, the use of ASCs may represent one of the best choices.
Footnotes
Author Disclosure Statement
No competing financial interests exist.