
Abstract
Billions of dollars are annually invested in pharmaceutical industry and cosmetic sector with intent to develop new drugs and treatment strategies for alopecia. Because the hair looks an important characteristic of humans—an effective appendage in perception, expression of beauty, and preservation of self-esteem—the global market for hair loss treatment products is exponentially increasing. However, current methods to treat hair loss endure yet multiple challenges, such as unfavorable outcomes, nonpermanent and patient-dependent results, as well as unpredictable impacts, which limit their application. Over recent years, remarkable advances in the fields of regenerative medicine and hair tissue engineering have raised new hopes for introducing novel cell-based approaches to treat hair loss. Through cell-based approaches, it is possible to produce hair-like structures in the laboratory setting or manipulate cells in their native niche (in vivo lineage reprogramming) to reconstruct the hair follicle. However, challenging issues still exist with the functionality of cultured human hair cells, the proper selection of nonhair cell sources in cases of shortage of donor hair, and the development of defined culture conditions. Moreover, in the case of in vivo lineage reprogramming, selecting appropriate induction factors and their efficient delivery to guide resident cells into a hair fate—with the aim of reconstructing functional hair—still needs further explorations. In this study, we highlight recent advances and current challenges in hair loss treatment using cell-based approaches and provide novel insights for crucial steps, which must be taken into account to develop reproducible, safe, and efficient cell-based treatment.
Introduction
M
Currently, significant advances in regenerative medicine and tissue engineering have opened new doors to address unmet medical needs, including hair loss treatment. Several research groups have developed strategies to generate hair-like structures in the laboratory setting [23 –29]. Although different groups have succeeded to generate hair-like structure in vitro with embryo and neonate rodent cells, the results have not still been implemented in humans because of multiple challenges [30]. Poor functionality of the cultured human hair cells, proper selection of nonhair cell sources in case of shortage of donor hair, and development of defined culture conditions are challenges hindering application of currently developed in vitro cell-based approaches in clinical applications.
To overcome these challenges, scientists have expressed interest in the conversion of resident cells to the desired cell fate in their native niche, defined as in vivo lineage reprogramming, with the aim to reconstruct functional hair in situ. However, selection of proper induction factors and their efficient delivery to guide resident cells into the hair fate remain to be explored. In this review, we highlight recent advances and critical challenges in treating hair loss according to cell-based approaches and provide novel insight to developing robust cell-based technologies for in vitro and in situ hair loss treatment.
Human Hair Follicles Cells and the Hair Cycle
Hair is considered a complicated mini-organ within the skin, consisting of broad cell population diversity, ranging from epithelial to mesenchymal and neural crest cells. Communication between these cells results in generation, maintenance, and renewal of hair during development, cell cycle, and wound repair [31 –35].
Multipotent epithelial cells are located in a specialized region called the bulge. In humans, the bulge region is considered a hair follicle (HF) stem cell niche located between the sebaceous gland and arrector pili muscle. Bulge epithelial stem cells can support not only growth of HFs and sebaceous glands but also provide all skin lineages to reconstitute new epidermis during wound repair [31,33,36]. During the continual HF cycle, descendants of bulge epithelial stem cells contribute to the outer root and inner root sheath, as well as hair shaft (HS) [32].
Melanocyte stem cells are also located in the bulge niche. They are originated from neural crest cells and have the capacity to differentiate, produce melanin, and contact adjacent keratinocyte cells to distribute pigment granules along the skin and hair [22,37]. Melanocyte stem cells cyclically proliferate and differentiate during hair cycle.
In addition to the above-mentioned cells, other important hair niche components include cells with mesenchymal identities such as dermal papilla (DP) and dermal sheath cells (DSCs). These cells are located in the base of hair and surround the outer root sheath (ORS), respectively. DP cells (DPCs) regulate hair formation during embryogenesis and cell cycle during postnatal life. DPCs and DSCs are originated from mesoderm. Moreover, the neural crest origination of DPCs has been confirmed [38,39]. Each HF is associated with a sebaceous gland that has its own stem cells, where its main cells, sebocytes, produce lipid-rich products [40]. Other components of hair niche, including blood vessels, nerves, and adipocyte cells, surround the hair within the dermis to support hair growth and regeneration [41].
Although several studies have shown the importance of niche components and signals such as Wnt, bone morphogenetic proteins (BMPs), and sonic hedgehog (Shh) in regulating mouse HF behavior [42], the nature of human HF cells and their interaction with niche have not yet been fully understood.
Normally, mature hair has a regenerative cycling system that consists of three phases: anagen, catagen, and telogen. Anagen phase lasts 1–6 years on the human scalp with rapidly proliferating epithelial cells that finally differentiate into trichocytes and receive melanin granules from melanocyte cells, leading to the generation of a pigmented long HS. Following these active events, as the consequence of an undetermined hair cycle clock, epithelial and melanocyte cells undergo apoptotic processes, where a new HS replaces the old one in the next anagen phase. Catagen phase is the regression and distractive phase of the hair cycle that lasts only 4–6 weeks. Telogen phase follows this preprogrammed suicide where the remaining hair cells relax for 2–3 months on the scalp. Next, bulge epithelial stem cells undergo subsequent proliferation and differentiation in a new anagen phase. Therefore, the older HS is shed and replaced by a new HS [19,43 –45]. However, in some cases, a new HS is not generated, leading to hair loss.
Therapeutic Strategies to Treat Hair Loss
Hair loss can occur if any factor, causing abnormalities in HF cycling, decreases the stem cell activity or failures happen in regrowth of hair fibers from existing HFs [19,46]. Common events promoting hair loss include heredity factors, diseases, hormonal changes, chemotherapy agents, physical and emotional stress, poor nutrition, excessive hair styling, and aging [13]. Generally, hair loss can be categorized as scarring (permanent, cicatricial, nonreversible) and nonscarring (temporary, noncicatricial, reversible) alopecia. In scarring alopecia, HF stem cells are damaged, while nonscarring hair loss is associated with progenitor cell destruction [47]. Currently, different methods and strategies are available to treat hair loss, such as cell-derived conditioned media [3 –5], platelet-rich plasma [6 –10], plant extracts [11 –13], phototherapy [14], surgical treatments [15,16], and medications [17,18]. However, the majority of these methods suffer from multiple challenges.
Conditioned media produced using keratinocyte stem/progenitor [4], dermal stem/progenitor [3], adipose stem cells [5], and platelet-rich plasma [6,7] are enriched with growth factors; thus they are proposed as alternative methods for hair loss treatment. Although simple, these methods are mainly used as adjuvant treatments, while they are not effective enough to be used alone [8 –10]. Additional randomized, controlled double-blind studies are required to confirm the role of these methods in direct stimulation of hair growth [6,50].
Plant extracts are another possible method to treat hair loss. Rho et al. have explored 45 oriental plant extracts for hair loss treatment and suggested that Asiasari radix had a stimulatory effect on hair growth in C57BL/6 and C3H mice. This extract promoted the expression of vascular endothelial growth factor in the cultured human DPCs that strongly played a role in control of hair growth, particularly during the anagen phase [11]. The majority of studies that investigated the effects of plants and their extracts on hair growth promotion have been performed on animals. More academic clues and defined mechanism of actions about herbal extracts are still needed before their widespread use for clinical applications [12,13].
Phototherapy is another suggested option for hair loss treatment. However, controversial results regarding its efficacy and serious concerns about the risk of cutaneous malignancies with plus ultraviolet-A light make phototherapy a less-favorable treatment option [14].
HF transplantation is a surgical treatment that involves taking plugs of normal occipital hair and transplanting them into bald areas [15,16,22]. However, this method has the following limitations: invasive and painful procedure, high cost, shortage of donor hair, and redistribution of only the remaining hairs with no new hair generation [16,49].
Pharmaceutical industries invest billions of dollars annually to introduce new drugs for hair loss treatment [50]. Analysis of patent trends during the last decade shows that numerous companies mainly focus on discovering new “antihair loss” agents to stimulate HF regeneration and neogeneration. Despite significant advances, most currently available drugs have unsatisfactory outcomes [19 –21].
Recent progresses in stem cell biology and regenerative medicine have opened new roots to cope with challenges in the conventional hair treatment methods. To date, many research groups have attempted to reconstruct HFs by culturing HF epithelial and DPCs in vitro and transplanting them in animals [22,51].
In Vitro Cell-Based Approaches for Hair Reconstruction and Current Challenges
Basically, human HF stem cells (HFSCs) and DPCs are two important cell types affected by hair loss. In scarring alopecia, including cutaneous lupus erythematosus and lichen planopilaris, inflammatory cell infiltration around bulge causes an irreversible loss of HFSCs [19,21]. In nonscarring alopecia, such as alopecia areata and androgenic alopecia (AGA), hair progenitor cells become damaged, however, HFSCs are preserved. Preservation of HFSCs shows the potential reversibility of this condition [47]. Moreover, in AGA, as the most common type of hair loss in men and women, decrease in DPC number at the base of hair leads to a gradual decrease in the hair size and subsequent alopecia [22,52,53]. Thus, maintenance of HFSCs and DPCs along with their well-orchestrated interactions is necessary for normal hair regeneration as well as hair bioengineering and reconstruction in vitro [30].
In vitro hair reconstruction involves four keys steps: (i) selection of proper starting cell sources; (ii) isolation of desired cell sources and the development of effective protocols that generate adequate numbers of functional HFSCs and DPCs; (iii) assembling organized HF-like structures by coculturing HFSCs and DPCs in conditions enabling proper epithelial–mesenchymal interaction and compartmentalization; and (iv) transplantation of bioengineered HF-like structures in patients or animals to generate fully organized hair structures (Fig. 1) [30]. We intend to describe these key steps in detail and highlight existing challenges.
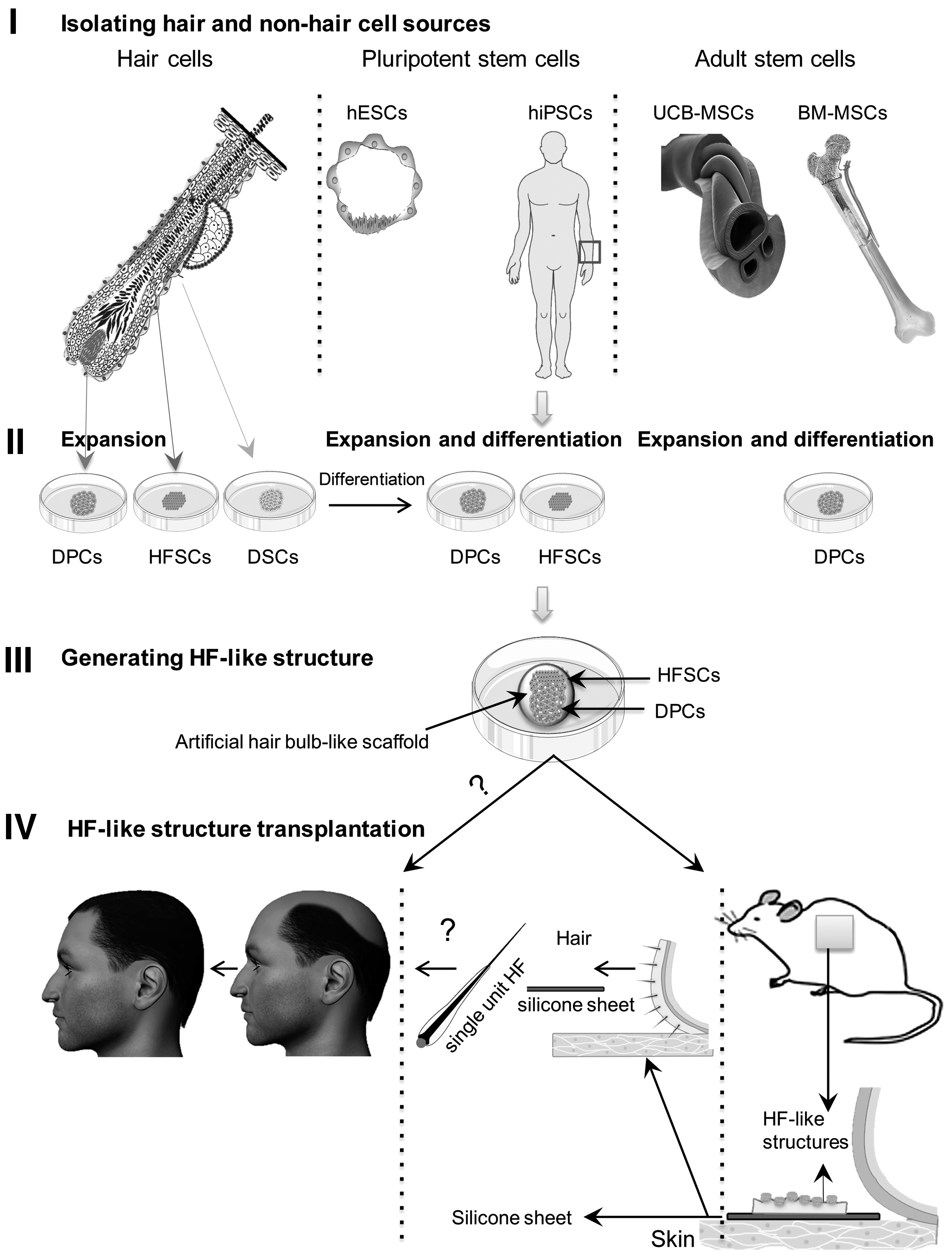
Hair and nonhair (pluripotent and adult stem cells) cell sources used for hair reconstruction in vitro.
Selection of proper starting cell sources for in vitro hair generation
Bioengineering of HFs requires two types of hair cells: competent HFSCs and inductive DPCs. These cells can be derived from hair and nonhair sources [30]. Therefore, the correct choice of proper, available sources to treat hair loss is the first consideration.
Hair cell sources
Isolation of HFSCs and DPCs from hair tissue is the best option to produce the HF structure in vitro. HFSCs are located between the insertion of the arrector pili muscle and sebaceous gland duct in the ORS [31,54]. Proposed markers to identify HFSCs include K15, K19, PHLDA1, FST, FZD1, and CD200, as well as a cocktail of CD200+ CD24− CD34− CD71− CD146− Connexin 43− MHC-Ia−, which are more applicable to the clinical setting and can be used to sort live cells [31].
In addition to HFSCs, cells that have a mesenchymal identity such as DPCs are another essential component, which is considered for HF reconstruction. These cells are located in the base of hair. DPCs regulate hair formation during embryogenesis and cell cycle within postnatal life [55]. Mouse DPCs can be successfully isolated and identified using CD133 and integrin alpha 9, as two cell surface markers, by fluorescence-activated cell sorting (FACS). In contrast, human DPCs are separated by manual microdissection [30,56 –58].
Although isolation of HFSCs and DPCs from patient hair can be considered a suitable strategy for hair cell therapy, other nonhair sources should be considered for scarring alopecia (permanent hair loss) and nonscarring alopecia cases, who have a shortage of donor hair.
Nonhair cell sources
Different nonhair cell sources can be used to generate HFSCs and DPCs, including human pluripotent stem cells (hPSCs) and adult stem/progenitor cells [30,59 –62]. hPSCs comprised human embryonic stem cells (hESCs) and human induced pluripotent stem cells (hiPSCs). These cells are interesting sources to develop cell-based therapy approaches, due to their tremendous ability to generate all body cell lineages as well as the availability of robust protocols for their large-scale expansion and integrated differentiation. In the case of hESCs, ethical and immunological challenges should be overcome before their use as starting cell material. One critical approach to overcome immunological challenges of hESCs could indeed be developing national/international large HLA-matched hESC cell banks—a potentially complicated process [63].
Generation of iPSCs overcomes these ethical and immunological concerns. However, the safety concerns about hiPSC clinical applications could generally be arisen for the cells modified by genetic transcription factors and some viral delivery systems in the reprogramming process, causing integration of viral genetic materials into the host genome after transfection. This integration can increase the risk of harmful endogenous genetic and epigenetic modifications such as insertional mutagenic lesions in the host cell genome that can cause tumor formation after transplantation of differentiated cells.
To date, new potential approaches have been suggested to decrease the unwanted outcome of cell reprogramming such as the use of nonintegrating viruses, transposon-based and cell reprogramming factor-based plasmid systems, and recombinant reprogramming factor proteins, nanoparticles, small molecules, mRNA encoding reprogramming factors, and miRNAs [64]. As a case, Igawa et al. have used a piggyBac transposon to successfully generate functional keratinocyte cells from transgene-free iPSCs, which is closer to clinical application than transgene-residual iPSCs [65]. Although these proposed methods are more suitable for safe cell reprogramming from a medical point of view, their efficacies are still lower than viral delivery [66].
Adult stem/progenitor cells can offer an alternative source to overcome the ethical, immunological, and tumorigenic concerns with hPSCs. These cells can be differentiated or transdifferentiated into desired cells.
For instance, investigations demonstrated conversion of somatic human epidermal keratinocytes into their precursor cells in vitro, using only basic fibroblast growth factor (bFGF) rather than external gene interventions. In this study, terminally differentiated cells were changed into keratinocyte precursor cells at three levels: protein expression, morphology, and function. Coordination of Wnt and BMP along with FGF has important roles in skin and hair embryonic specification [67]. Moreover, few studies have reported that adipose-derived stem cells and circulating monocytes have a transdifferentiation potential to convert into keratinocyte-like cells (KLC) [68,69]. Although these studies are noteworthy, it has been shown that generated epithelial cells have interfollicular identity and are not folliculogenic.
In terms of DPCs, DSCs can provide a powerful source of trichogenic mesenchymal cells for induction of new hair because DP and DS have the same embryological origin. Paus et al. reported that DSCs, obtained from a male human scalp, formed new hair in the arm of a female recipient without any immunological rejection. In addition, transition of cells between DP and DS during the normal hair cycle could occur [57,70,71]. In line with these evidences, cotransplantation of passage-1 DSCs and passage-60 DPCs (that did not have the capability to induce new hair) led to HF induction. These results indicated that certain trichogenic genes were stimulated to express in DSCs when cocultured with DPCs [72].
In addition, culturing and preconditioning human bone marrow-derived mesenchymal stem cells (hBM-MSCs) and human umbilical cord blood-derived MSCs (hUCB-MSCs) in DP forming medium (DPFM) caused formation of cell-aggregated spheroidal DP-like structures (DPLTs). These DPLTs showed DPC-specific markers (MRPS, HSP70, laminin, type IV collagen, and versican) in vitro. Transplantation of DPLTs plus ORS cells into the inner dermis of the scalp of athymic nude mice resulted in the generation of new hair on their heads [73]. On the other hand, coculturing human MSCs (hMSCs) with DPCs caused hMSC differentiation into DPCs. Increased DP marker expressions such as versican, CD133, stem cell factor, endothelin-1 (ET-1), and bFGF during the coculture along with the production of more aggregates in comparison with hMSCs alone, confirmed this ability of hMSCs. Transplanting hMSCs in BALB/c-nu/nu mice showed that established cells were nontumorigenic and they were potentially able to differentiate into DPs [74].
Skin-derived precursor cells (SKPs) are introduced as another possible cell source to produce DPCs. These cells are a multipotent subpopulation in the juvenile and adult rodent skin dermis that can differentiate into neurons, glia, smooth muscle cells, and adipocytes in vitro [75]. Transplanting SKPs into the back skin of nude mice has been shown to induce hair morphogenesis, homing back to the DP and DS, and serial reconstitution of HFs [76]. SKPs can be isolated from human dermis scalp [75] and neonatal human foreskin tissue. Scalp and neonatal foreskin SKPs are a readily accessible source compared to hBM-MSCs and hUCB-MSCs. They grow in suspension as spheres in the presence of the mitogen FGF-2 and epidermal growth factor (EGF), and express embryonic TFs. It is proposed that this source may provide promising starting cells to produce DPCs in future [77].
Generation of adequate numbers of homogeneous and functional clinical-grade HFSCs and DPCs
To produce adequate numbers of clinical-grade target cell type for hair cell therapy, selected tissue/stem and progenitor cell sources should be isolated, expanded and/or differentiated in xeno-free and fully defined culture conditions. The culture system should be robust enough for expansion of stem/progenitor cells, while maintaining their genetic stability and differentiation potential. Besides, an integrated differentiation culture system should be developed to produce mature and functional target cells [63].
The required number of cells for each patient cell therapy may vary due to the extent and type of hair loss. It has been implicated that thousands of HFs should be generated for a single patient to achieve an acceptable cosmetic appearance [46]. Zheng et al. reported that 2,500 epidermal and 5,000 dermal cells would be sufficient to regenerate a single HF in a mouse hair reconstitution assay [78]. Thus, HFSCs and DPCs isolated from a patient should be expanded before transplantation as an autologous strategy. On the other hand, if nonhair sources were used as starting cells, they should first be expanded. After reaching an adequate number of cells, they could differentiate into HFSCs and DPCs to achieve clinically relevant cell numbers. Essentially, all of these processes should be conducted under good manufacturing practice (GMP) conditions with defined culture systems in an affordable manner for clinical applications.
Scalable expansions of HFSCs and DPCs
HFSCs, which are isolated according to the CD200hi CD24lo CD34lo CD71lo CD146lo markers [31], should be cultured in a system that supports their robust, scalable expansion to obtain sufficient quantity of cells with the ability to respond to trichogenic dermal signals. The conventional method for culturing epithelial cells with fetal bovine serum (FBS) and murine 3T3 feeder layers has been approved by FDA, as a xenogeneic product [79]. However, risk of HFSC contamination is considerable due to utilizing feeder cells and their xenogeneic components such as pathogens, subsequently leading to promote the chance of infection or immune reactions after transplantation of the product. These issues can be solved by culturing HFSCs on GMP-grade human feeder layer cell lines [63]. Moreover, instead of FBS, researchers have proposed to use autologous human serum, defined FBS, human platelet lysates [80], or serum-free medium with defined growth factors [81].
Another hair starting cell source is DPCs. The final goal in culturing DPCs is the production of adequate numbers of these cells, while maintaining their hair inductive properties. DPCs can be isolated by time-consuming microdissection processes before propagation in culture [30]. To date, different approaches have been suggested to maintain the capacity of these cells to induce hair regeneration. Coculturing DPCs with keratinocyte cells or keratinocyte-conditioned medium and the use of different signaling regulators such as 6-bromoindirubin-3-oxime (BIO), Wnt-3a, recombinant BMP2, BMP6, bFGF, glycogen synthase kinase-3β inhibitor, and vitamin D3 (an analogue of TGFβ) can help to maintain their inductive properties [30,39,82].
Self-assembling culture conditions and three-dimensional (3D) dermal spheres can also help to preserve or restore the inductive properties of DPCs in vitro [83 –86]. Biomaterials, used to maintain cell motility such as poly-EVAL (ethylene-co-vinyl alcohol) membranes [87] and fibronectin-coated EVAL [85], can promote DP self-assembly into many compact spheroidal microtissues and induce new HFs [87]. DPC self-aggregation can facilitate their culture in a scalable suspension bioreactor and produce adequate numbers of DPCs for clinical applications [46]. However, most of these protocols are still under optimization and generally use undefined and animal-derived culture media, which make them inappropriate for clinical applications.
Integrated expansion and differentiation of hPSCs into HFSCs and DPCs
When using hPSCs as nonhair cell sources, clinical-grade hESC and hiPSC lines should be available or generated under GMP conditions. Next, a robust culture system should be developed for scalable expansion of hPSCs, followed by their integrated differentiation to HFSCs and DPCs under xeno-free or defined culture conditions. Fortunately, different culture systems and media have been used for scalable production of hPSCs in xeno-free or defined culture systems [63].
However, current protocols for differentiation of hPSCs into epithelial cells and DPCs mainly depend on the application of mouse feeder cells and the use of a xenogenic culture system, which are not compatible to clinical-grade cell production [38,59,88 –91]. To develop a defined differentiation protocol for generation of HFSCs from hPSCs, Ji et al. have generated embryoid bodies (EB) from hESCs and guided them into keratinocyte cells in a defined keratinocyte serum-free medium on a gelatin matrix [89]. In another study, Metallo et al. differentiated hESC colonies with retinoic acid (RA) and BMP. Subsequently, the differentiated cells were cultured in the defined keratinocyte serum-free medium [92]. RA and BMP were used to promote and block, respectively, ectodermal and neural fates. Using a similar approach, normal and recessive dystrophic epidermolysis bullosa-induced pluripotent stem cells were differentiated into keratinocyte cells by RA and BMP in defined keratinocyte serum-free medium [90].
In another study that aimed to minimize animal component during differentiation of hESCs into keratinocyte cells, Kidwai et al. introduced an autogenic feeder-free, serum-free (AFF) system. In this AFF system, hESCs were seeded on ECM extracted from hESC-derived fibroblasts in an animal component-free TeSR2 medium. For keratinocyte differentiation, TeSR2 was replaced by a defined keratinocyte serum-free medium supplemented with 1 mM of RA [91]. These findings have paved the way for establishment of animal-free and defined differentiation protocols to produce safe target cells for potential clinical applications. However, keratinocytes generated with these culture systems have interfollicular identity, but not folliculogenic.
In an attempt to produce epithelial cells with folliculogenic characteristics, Veraitch et al. differentiated hiPSCs into ectodermal precursor cells (EPCs). In this study, dermal fibroblasts derived from the facial or scalp dermis were reprogrammed into hiPSCs using three or four reprogramming factors: POU5F1, SOX2, KLF4, and MYC. Generated EB from hiPSCs were exposed to RA and BMP4, then transferred to collagen I-coated plates in a defined keratinocyte serum-free medium. Transplantation of generated EPCs with inductive mice dermal cells into immunodeficient mice led to HF formation [60].
To generate patient-specific cells, Yang et al. differentiated hiPSCs into epithelial hair stem cells. First, human fibroblasts were reprogrammed into hiPSCs by a retrovirus solution, and then cultured on mouse embryo fibroblast (MEF) feeder and hESC culture medium. The generated EB from hiPSCs were seeded onto 3T3 fibroblasts previously treated with mitomycin C in a differentiation medium. In this sequential differentiation protocol, RA was used to induce hiPSCs into ectodermal-like cells (stage 1); BMP4 and EGF were used to differentiate ectodermal-like cells into epithelial stem cells (stage 2); and finally, EGF was used to expand mature keratinocyte lineages (stage 3). Epithelial hair stem cells or CD200+/ITGA6+/SSEA3− cells derived from iPSCs were sorted and combined with freshly isolated neonatal mice dermal cells to check their ability to produce hair in the nude mice. Human iPSC-derived CD200+/ITGA6+ cells had an ability to produce all HF lineages in skin reconstitution assays [59]. The generated epithelial cells in these two studies had an ability to form HF in vivo. The main drawbacks of these works included using a viral delivery system in the reprogramming process, application of mouse feeder cells, and media containing xenogeneic animal proteins, which made these protocols unsuitable for clinical application.
In the case of DPCs, Gnedeva et al. successfully produced DPCs by using neutralization and DP media from ESCs. Transplantation of these cells with mouse epidermal cells, isolated from newborn animals, has shown capability of the differentiated cells to induce hair [38]. However, further studies are needed to determine the accurate method and defined conditions for production of DPCs from hPSCs. Generally, the development of scalable and defined integrated culture systems to produce HFSCs and DPCs from hPSCs is still in progress.
Integrated expansion and differentiation of adult stem/progenitor cells into HFSCs and DPCs
Adult stem/progenitor cells are also promising sources for the generation of clinical-grade HFSCs and DPCs. These cells should first be expanded, then differentiated/transdifferentiated into target cells in xeno-free/defined culture conditions, for clinical applications. Transdifferentiation of adipose-derived stem cells (ADSCs) into KLC has been performed by two approaches: using cocultures of ADCs with KLCs or keratinocyte-conditioned media. Although ADSCs were successfully transdifferentiated into KLCs and could form a stratified epidermis, the final cells did not have folliculogenic characteristics. In addition, the expansion and differentiation media contained xenogeneic animal protein components [68].
In another study, circulating monocytes transdifferentiated into KLCs in the culture medium composed of 49% Dulbecco's modified Eagle's medium and 49% defined Keratinocyte serum-free medium with its growth supplement plus 2% FBS. However, the final cells had interfollicular identity and were not folliculogenic [69].
In the case of DPCs, hBM-MSCs and hUCB-MSCs in xenogeneic DPFM have the capacity to produce DPLTs. The produced DPLTs have an ability to induce new HFs in athymic mice [73,74]. Recently, different serum-free media based on a cocktail of growth factors have been introduced for MSC expansion for therapeutic use [93,94]. Thus, developing a xeno-free or defined media for differentiation of MSCs into DPCs can largely facilitate process of clinical applications.
Purification of HFSCs and DPCs generated from nonhair sources
While nonhair sources, particularly hPSCs, are used as starting material to produce HFSCs and DPCs, the presence of unwanted cells (ie, hPSCs) in the final product (due to different differentiation efficacies) may cause serious problems such as teratoma formation after transplantation. Thus, separation of HFSCs and DPCs from undesired cell populations generated in the differentiation process is a critical step that should be considered to ensure the product safety and quality. Passaging [90] as well as utilizing positive and negative specific cell surface markers against starting (ESCs or iPSCs) and final cells can minimize contamination of undifferentiated cells [59]. It can also increase purity of the final cells. In addition, GMP-grade selective markers should be used for separation of HFSCs (CD200+ CD24− CD34− CD71− CD146− Connexin 43− MHC-Ia−) by FACS or MACS technology. However, specific cell surface markers for DPCs should be explored.
After successful production of pure HFSC and DPC populations, it is crucial to properly mix these cells and optimize their compartmentalization to generate new hair with correct architecture and density [30]. We intend to discuss conditions where both cells have maximum interaction and can provide hair structure after transplantation in animal models.
Coculture of DPCs and HFSCs and transplanting hair-like structures for final maturation
Good hair bioengineering consists of producing sufficient numbers of inductive DPCs together with highly competent HFSCs and then establishing their proper combination in an appropriate scaffold. Providing a native-like 3D environment that properly mimics the developmental process of hair organogenesis can enhance and maximize the interactions between DPCs and HFSCs [26]. In addition, designing complex scaffolds that can mimic HF bulb structure with spatiotemporal delivery of growth factors to the cell niche may facilitate the reconstruction of a human hair-like structure in vitro. For example, layered and mixed sandwich organotypic culture systems combined with serum-free low calcium conditions have led to spheroid epithelial cell aggregate formation from human ORS keratinocyte (ORSK). In both structures, collagen-I was mixed with human dermal fibroblasts and used as a pseudodermis. In the layered system, pseudodermis was first covered by Matrigel™ that contained human DPCs, followed by a layer of an ORSK cell suspension. In the sandwich system, top of the pseudodermis construct was covered by a mixture of Matrigel, human DPC, and human ORSK [23]. Later, Havlickova et al. developed a pragmatic human folliculoid microsphere (HFM) approach to assess epithelial–mesenchymal interactions in the HFM. In this method, human HF keratinocytes and DPCs encapsulated in a collagen-I and Matrigel basement membrane matrix were entered into the Miglyol-FC-40 mixture and formed HFM. Evaluation of HFM showed an effective interaction between the HF keratinocytes and DPCs, and formed spheroid cell aggregates. In addition, HF keratinocytes exhibited HF-type keratinization, considerable proliferation, and a low level of apoptosis in this system. DPCs had minimal proliferation and apoptosis along with a strong expression of versican [24].
Another significant progress involved in this field is construction of “protohairs” in vitro and their transplantation into nude mice to allow further maturation of HFs. In this study, hanging drop-derived aggregates consisted of embryonic mouse follicular dermal and epidermal cells. These aggregates were maintained for 5–7 days in a cell culture and successfully developed hair-like structures called “protohairs.” The “protohairs” were intradermally transplanted into nude mice to develop mature HFs. Implanted “protohairs” that were anchored to the mice skin fully developed into HFs, which were maintained for at least 6 months [25]. Folliculoid microtissues were also constructed from DPCs and keratinocyte cells. In this method, rat keratinocytes and DP cells seeded on poly-EVAL formed aggregates, similar to spheroidal microtissues. These spheroidal microtissues consisted of a DPC core surrounded by a keratinocyte shell. Spheroidal microtissues formed new hairs after transplantation into the hypodermis of nude mice [27].
In another attempt, microfollicles were prepared in tissue culture from human follicular keratinocytes, melanocytes, and DPCs encapsulated in extracellular matrices. In this method, the emerging fibers from these microfollicles were very similar to vellus (unpigmented) hair [28]. Since efficient treating of hair loss requires a large numbers of HFs with a cosmetically acceptable appearance, Lee et al. have developed a simple procedure in which newborn mouse cells were seeded into a clinical-grade matrix (Integra Bilayer Wound Matrix) to produce large numbers of follicle with a cosmetically acceptable appearance. Transplanting this construct in full-thickness skin wounds of athymic mice showed that newborn mice cells could differentiate properly and form a reconstituted skin with different skin components, including hairs, sebaceous glands, dermis, and subcutaneous adipose tissues. These HFs had the ability to cycle and regenerate continuously for at least 1 year. Although this method was a very promising approach to provide tissue implants with flexible size and shape for treating alopecia, it is required to produce a large number of multipotent skin stem cells as starting material [95].
HF neogenesis using the “organ germ method” is another approach to produce hair-like structures in vitro. Nakao et al. have produced bioengineered pelage and vibrissae follicle germ. Bioengineered pelage follicle germ was generated from epithelial and mesenchymal cells isolated from mouse embryo back skin in collagen gel. In addition, bioengineered vibrissa follicle was reconstituted from a mixture of cultured DPCs and adult epithelial vibrissae-derived bulge region in collagen gel. Both bioengineered structures could produce fully functional hair by transplanting into the subrenal capsule. The bioengineered HF showed correct structure, connected with the surrounding host tissues, hair cycle repetition, and piloerection ability [26,29]. To develop regeneration therapy for hair loss, mature bioengineered HFs generated by ectopic transplantation were intracutaneously transplanted on the back skins of nude mice, similar to follicular unit hair transplantation surgery. The bioengineered HFs properly connected with nerves and arrector pili muscle of host skin with a piloerection ability. These results provided a novel therapeutic approach for alopecia using HF regenerative therapy [51]. Following these studies with mouse cells, researchers successfully reconstituted a human bioengineered HF germ. Epithelial cells and intact DP derived from the bulge area and scalps of AGA patients, respectively, generated human hair germ. In addition, human hair germ could generate pigmented HS when transplanted into the back skins of nude mice [29].
In another study, using human cultured cells, Leiros et al. showed that human HF cells produced hair-like structures in vivo. In this method, a porcine acellular dermal matrix seeded with adult cultured HFSCs and human DPCs induced embryonic hair bud-like structures containing human cells when grafted in nude mice [96]. These noteworthy results along with the previous studies generated new hope on future hair regenerative therapy and human hair reconstruction in vitro. However, current technology is still far from successful generation of a fully organized human hair structure in vitro.
It is still necessary to transplant these bioengineered structures in mice or rat host environments to complete hair structure formation. Notably, species-specific differences between mice, rats, and humans in HF biology necessitate additional studies to translate these results to human applications [40,97]. Moreover, majority of the studies have shown that human HFSCs or DPCs should be cotransplanted with inductive mouse neonatal cells to produce hair in an animal model. No report is available to discuss the successful, functional mature HF construction from dissociated adult cultured HFSCs and DPCs in vitro [20,30,83,98,99]. To cope with these two critical challenges and other limitations in vitro, such as high numbers of required cells, lack of GMP grade and cost-effective expansion and differentiation culture media, reagents, enzymes, or scaffolds, potential undesired modifications in cultured cells, a time-consuming and labor-intensive process, as well as difficulties during transplantation of follicle-like structures, scientists are getting motivated to develop technologies that convert resident cells to the desired fate in their native niche. Conversion of resident cells in their niche or in vivo lineage reprogramming has recently gained increasing attention.
Conversion of Resident Cells to Hair Follicle Fate in Their Niche
Recently, expressing specific cocktails of TFs successfully converted exocrine cells into insulin-producing β cells [100], induced cardiomyocyte-like cells from cardiac fibroblasts [101], and converted resident astrocytes into mature neurons in mice [102]. In conjunction with a number of similar studies, they could be considered the basis for developing innovative hair loss treatment strategies for alopecia patients (Fig. 2A). Clues from wound repair in skin have shown that the reprogramming process could happen in the wound environment where cells in the wound center acquire embryonic-like competency and lineage plasticity to induce new hairs. To date, a number of studies showed strong evidence for reprogramming in wound healing. For instance, overexpression of Wnt ligand in the epidermis has resulted in remarkable hair neogenesis during wound healing [103 –105].
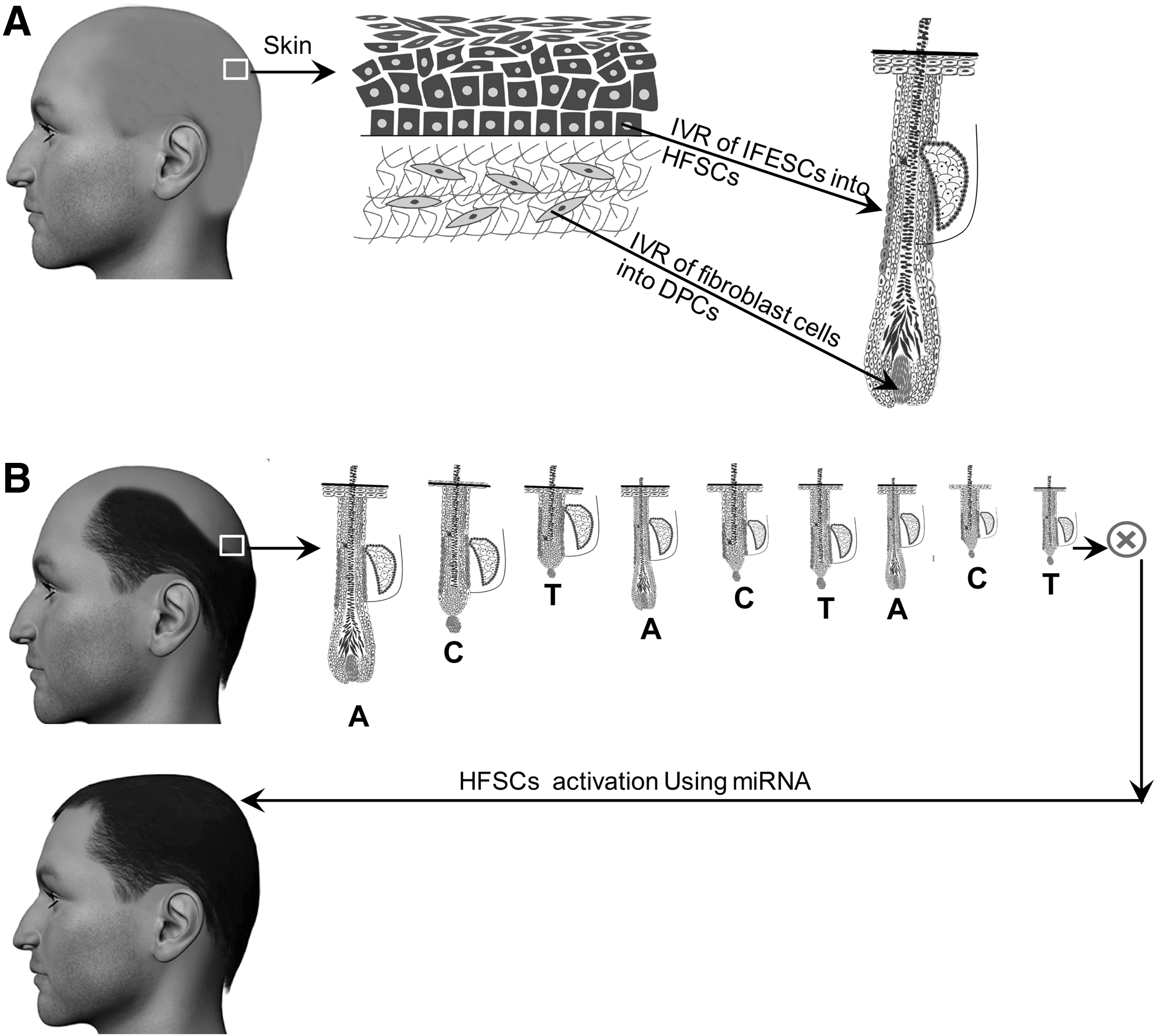
Conversion of resident cells to hair follicle fate in their niche may lead to treat hair loss.
Although majority of these studies highlighted the role of an inductive wound microenvironment in adopting a new cell fate, identification of developmental and wound healing-related TFs and converting skin epithelial stem cells or MSCs to hair epithelial cells or fibroblasts to DPCs, are promising concepts.
Generally, common embryological origin of HFSCs and interfollicular skin stem cells encourage development of this approach, particularly for refractory and scarring alopecia [106,107]. Modulating signaling pathways involved in hair morphogenesis such as Wnt, BMP, Shh, and TNF family member ectodysplasin-A may convert resident keratinocytes to follicular keratinocyte cells [30]. For instance, β-catenin activation in epithelial skin stem cells may induce new hair growth, independent from DPC signals [108]. This process may subsequently trigger dermal-like condensation from dermal fibroblasts and instruct them to form DPC-like structures [109]. Reynolds and Jahoda have shown that adult cultured rat pelage follicle DPCs had an ability to induce follicles by transdifferentiation of adult footpad epidermis. Normally, footpad comprised afollicular epidermis expressing different sets of keratins [110]. In addition, Silva-Vargas et al. showed that sebaceous glands and interfollicular epidermis have remarkable ability to acquire certain characteristics of bulge stem cells by β-catenin activation. Stimulated new HFs induced DPCs and provides a niche for melanocytes [111].
In addition to TFs, microRNAs (miRNAs) are other innovative approaches that can be considered in lineage reprogramming in vivo for future work. miRNAs are noncoding small RNAs regulating gene expression in a posttranscriptional manner [112].
A pioneering study by Fuchs has shown that miRNAs differentially express in skin and hair [113]. Later studies have focused on individual miRNAs to determine their roles in development, cell cycle, proliferation, differentiation, and apoptosis in skin and hair. For instance, miRNA-203 suppresses expression of master transcription factor p63 [114], miRNA-31 is expressed differentially during hair cycle [115], miRNA-125b is responsible for stem cell commitment in HFs [116], miRNA-205 is a vital factor to maintain skin stem cell expansion [117], and miRNA-34a/c is suppressed by p63 to maintain cell cycle progression [118]. Therefore, specific miRNAs may have the ability to stimulate stem cells, forcing transition from the hair resting stage (telogen) to the active stage (anagen), especially in AGA (Fig. 2B). In addition, miRNAs are involved in hair loss, as suggested by miRNA expression changes in DPCs of AGA patients [119]. Accumulation of 5α-dihydrotestosterone in DPCs in AGA as well as increased levels of reactive oxygen species in balding tissue can most likely promote changes in miRNA expression patterns and subsequently trigger hair loss [120 –122].
Developing strategies for inhibiting or overexpressing miRNA in the bald scalp may enable dermatologists to restore or compensate miss-functional signaling, which can consequently lead to reestablishment of the normal hair cycle. These techniques may induce changes in HFSC or DPC activities or their microenvironments to regenerate or neogenerate hair. Although miRNA has shown great clinical promise, safe and efficient methods for its efficient delivery remain challenging.
Generally, both viral- and nonviral-based methods are used to deliver miRNA into desired cells. Viral delivery is not well suited for clinical application due to potential toxicity and immunogenicity, among other concerns. Alternatively, nonviral delivery systems such as naked oligonucleotides, chemically modified oligonucleotides, lipid-based systems, dendrimers, synthetic and natural polymers, as well as exosomes are more adoptable strategies for therapeutic applications. Among these methods, Miravirsin, a locked nucleic acid-modified miRNA, has been applied to treat hepatitis C virus infection in a phase II clinical trial. Together, chemically modified liposomes, dendrimers, and especially nanoparticle delivery systems have gained attention for safe, efficient delivery of miRNAs [123]. Since the HF is considered an accessible organ, topical and local use of miRNA is possible for hair disorders to achieve enhanced delivery using lower doses of miRNA, which could avoid any undesirable side effects potentially associated with systemic delivery [124]. To date, several studies have been submitted in the formal clinical trial website (
Conclusion
Recent advances in human hair biology dramatically improved our understanding on hair regeneration and neogeneration mechanisms. Thus, they have raised new hopes and paved the way for developing regenerative hair loss therapies when conventional treatments are not effective.
This can be achieved by in vitro/vivo reconstruction of the HF. To date, isolation, maintenance, and expansion of hair and nonhair cell sources have witnessed a series of tremendous advances in the development of culture conditions and production of starting cell materials for human hair reconstruction in vitro. However, the development of xeno-free/defined expansion and differentiation culture systems for scalable and clinical grade production of hair and nonhair cell sources remains challenging. The next emerging and challenging field is the development of innovative bioengineering strategies for mimicking/reconstruction of the hair niche in vitro using the proper cell combinations and their integration with well-designed scaffolds for clinical-grade hair reconstruction, followed by subsequent large-scale production of relevant clinically applicable numbers of hair.
To overcome these challenges, in vivo induction of hair resident cells in their native niche, to stimulate the regeneration process, is a promising cell-free approach. However, the main challenges are determining efficacy and stability of therapeutic modifications for modulation of damaged hair with existing defects in the respective niche, as well as defining proper induction factors and delivery methods. More research can overcome these issues and bridge the gap between current hair regenerative therapy technologies and widespread clinical applications.
Footnotes
Acknowledgments
This study was funded by a grant provided from Royan Institute. We express our appreciation to all members of the Skin Program at Royan Institute for their helpful deliberations and consultation during this work. We would like to thank Dr. Sara Pahlavan, Dr. Vahid Ezatizadeh, and Lida Habbibi Rezaie for their critical comments.
Author Disclosure Statement
No competing financial interests exist.