
Abstract
POU5F1 (OCT4) is a crucial transcription factor for induction and maintenance of cellular pluripotency, as well as survival of germ cells in mammals. However, the homologues of POU5F1 in teleost fish, including zebrafish and medaka, now named Pou5f3, exhibit considerable differences in expression pattern and pluripotency-maintaining activity. To what extent the POU5F1 homologues are conserved in vertebrates has been unclear. In this study, we report that the POU5F1 homologue from the Nile tilapia (Oreochromis niloticus), OnPou5f3, displays an expression pattern and biological activity somewhat different from those in zebrafish or medaka. The expression of Onpou5f3 at both mRNA and protein levels was abundant in early development embryos until blastula stages, barely detectable as proceeding, and then displayed a transiently strong expression domain in the brain region during neurula stages similar to zebrafish but not medaka. Afterward, OnPou5f3 appeared as germline-restricted (including primordial germ cells and female and male gonad germ cells) expression just like medaka. Notably, OnPou5f3 depletion through morpholino oligos caused blastula blockage or lethality and failure of survival and proliferation of blastula cell-derived cells. These findings indicate that equivalent POU5F1-like expression and activity of Pou5f3 might be conserved accompanying with species-specific expression pattern during evolution. Our study provides insight into the evolutionary conservation of the POU5F1 homologues across vertebrates.
Introduction
P
The POU5F1 homologues have also been identified and described in teleost fishes. In zebrafish, it was identified and originally named as Pou2, subsequently changed to Pou5f1 (or Oct4) and again, recently to Pou5f3 [11 –14]. Meanwhile, the POU5F1 homologue in medaka was identified and described as previously Oct4 [15,16]. Notably, there are salient differences between the two homologues in expression and function. Zebrafish pou5f3 transcripts are present in all blastomeres until the midblastula stage, restricted to the epiblast during gastrulation and present in the neural plate until midsomitogenesis, but not in PGCs [11,12,17]. In contrast, medaka pou5f3 shows not only pluripotency-restricted expression in early pluripotent embryo cells and ES cells but also in PGCs and adult gonadal germ cells [15,16]. Most importantly, zebrafish maternal and zygotic null pou5f3 mutants do not exhibit developmental blockage until the midgastrula stage, suggesting that the embryos are normal in cell division and proliferation during blastula stage [11,18], while medaka Pou5f3 depletion through morpholino oligos causes blastula lethality or blockage and abolishes ES cell derivation from midblastula embryo (MBE) [16]. Therefore, it is necessary to introduce other fish species to medaka and zebrafish to provide new insight into the evolutionary conservation of POU5F1 homologues in teleost fish.
As a worldwide farmed fish, the Nile tilapia (Oreochromis niloticus) features fast growth, easy breeding, and strong resistance to environment hardness. Moreover, it is also an excellent fish model besides zebrafish and medaka for early embryo development study. For example, embryos can be available every day throughout the year owing to the availability of monosex offspring and short spawning cycle (14 days) [19 –21]. In the present study, the POU5F1 homologue from Nile tilapia, named Onpou5f3, has been isolated and characterized. Through reverse transcription polymerase chain reaction (RT-PCR) and immunostaining assays using a specific antibody to OnPou5f3, the mRNA and protein expression of Onpou5f3 were detected in embryos during different development stages and gonads. Moreover, the biological activity of OnPou5f3 was investigated through morpholino-mediated knockdown.
Materials and Methods
Animals
Nile tilapias were kept in recirculating aerated water tanks at 26°C under an artificial photoperiod of 12 h light to 10 h darkness. All genetic females (XX) and males (XY) were obtained by artificial fertilization of eggs from control females (XX) with sperm from either sex reversed males (XX) or super males (YY), respectively. All animal experiments were carried out in accordance with the Guidelines for Care and Use of Laboratory Animals prescribed by the Regulation of Animal Experimentation of Chongqing, China.
cDNA cloning and sequence analyses
Total RNAs were extracted from the ovary and testis of adult Nile tilapias using RNAiso PLUS (Takara, Japan) as previously described [19]. One microgram of total RNA was reverse transcribed into cDNA using the PrimeScript II 1st Strand cDNA Synthesis Kit (Takara, Japan). By searching the GenBank and blast analysis, a predicted cDNA sequence with the full open reading frame (XM_003444407.3) was obtained. To verify the cDNA sequence validity, PCR primers for the cDNA were designed and then PCR products were purified, subcloned, and sequenced.
The multiple sequence alignment was generated using CLUSTALW and GeneDoc program (
Reverse transcription-PCR
Total RNAs were extracted from Nile tilapia embryos at the desired stage and adult dissected tissues, including brain, pituitary, gill, heart, spleen, liver, intestine, ovary, testis, kidney, muscle, and head kidney, and then reverse transcribed to cDNAs as mentioned above. The mRNA expression of POU5F3 was measured by RT-PCR with the validated primers (Supplementary Table S1; Supplementary Data are available online at
Antibody production and western blotting
A 16-aa peptide with the sequence “ENSENPQDMYKIERVF” was used to produce the rabbit anti-Pou5f3 antibody (αPou5f3) by a commercial antibody company (Qingdao, China). This peptide is located between the two POU subdomains of OnPou5f3 (Supplementary Fig. S1), which appears only once in the sequence of Nile tilapia genome. The reaction of αPou5f3 was verified by western blotting analysis using the crude total proteins of brain, ovary, and testis extracted from adult tilapias as described [20] and then the specificity of αPou5f3 was further detected through competition inhibition assay with the synthesized 16-aa peptide.
Immunofluorescence
Immunofluorescence (IF) was done as previously described [20,22]. Briefly, after the dissociated embryonic cells were fixed, perforated with 0.1% Triton X-100 (in PBST) and blocked with 5% fetal bovine serum, the cells were incubated with αPou5f3 or preserum at 1:1,000 dilution and detected by PE-conjugated anti-rabbit IgG (1:500) (Vector Laboratories), followed by nuclear staining with DAPI and mounting in the SlowFade Light Antifade Kit (Molecular Probes). Similarly, embryos at different developmental stages were sequentially fixed, dehydrated, rehydrated, perforated with cold Acetone, and blocked with 5% fetal bovine serum, respectively, and then embryos were treated as the above described. The fluorescent signals were observed under Nikon Ti-S fluorescence microscope or Leica AF6000 fluorescence microscopy.
Immunohistochemistry
Immunohistochemistry (IHC) was performed as described [20]. Briefly, samples from the Nile tilapias were dissected, fixed in Bouin's solution for 24 h, and subsequently dehydrated, embedded in paraffin, and serially sectioned at 5 μm thickness. Endogenous peroxidase activity was quenched using 3% H2O2 for 10 min at room temperature; nonspecific binding sites were blocked using 5% bovine serum albumin. The sections were incubated with 1:1,000 dilution of αPou5f3 or preserum and, subsequently, incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (Vector Laboratories). After being washed in 1 × PBS for several times, slides were developed with 3, 3′-diaminobenzidine and counterstained with hematoxylin.
Morpholino antisense oligos
Two morpholino antisense oligos (MO) were synthesized (Gene Tool, Oregon). One is MOoct (ATCCTCAGGACTTTCAGA
Cell culture
According to the protocols as previously described [23], the Nile tilapia blastula cells were dissociated and incubated in ESM2 with minor modifications, which is the Dulbecco's modified Eagle's medium with 25 mM HEPES, 100 U/mL penicillin and streptomycin, 15% fetal bovine serum, 2 mM
Alkaline phosphatase staining
The alkaline phosphatase (AP) activity of the in vitro cultured cells was detected as previously described [23]. Briefly, the cells were fixed with 4% formaldehyde, washed, and then stained with bromochloroindolyl phosphate/nitroblue tetrazolium (BCIP/NBT, Beyotime).
Results
cDNA sequence and analyses
The full open reading frame of Nile tilapia pou5f3 (Onpou5f3) is 1,383 bp in length and verified by sequencing. The cDNA encodes for a 460-aa putative protein with a calculated molecular mass of 50.9 kDa (Supplementary Fig. S1). Like the other vertebrate homolog, Onpou5f3 displays a five-exon genomic organization characteristic of the class V POU family genes. The putative protein OnPou5f3 consists of an N-terminus, a POU-specific domain (POUs), a short linker, a POU-homeodomain (POUh), and a C-terminus (Fig. 1A, B). Comparing with the POU5F1 homologues in other vertebrates, OnPou5f3 shares an identity higher than 60% to zebrafish and medaka, while relatively low identity to human (40%) and mouse (39%), and the lowest identity to Xenopus (34%). Meanwhile, the POU domains of the POU5F1 homologues are highly conserved among vertebrates, with identity higher than 90% to medaka and zebrafish, 60% to the others (Fig. 1B).
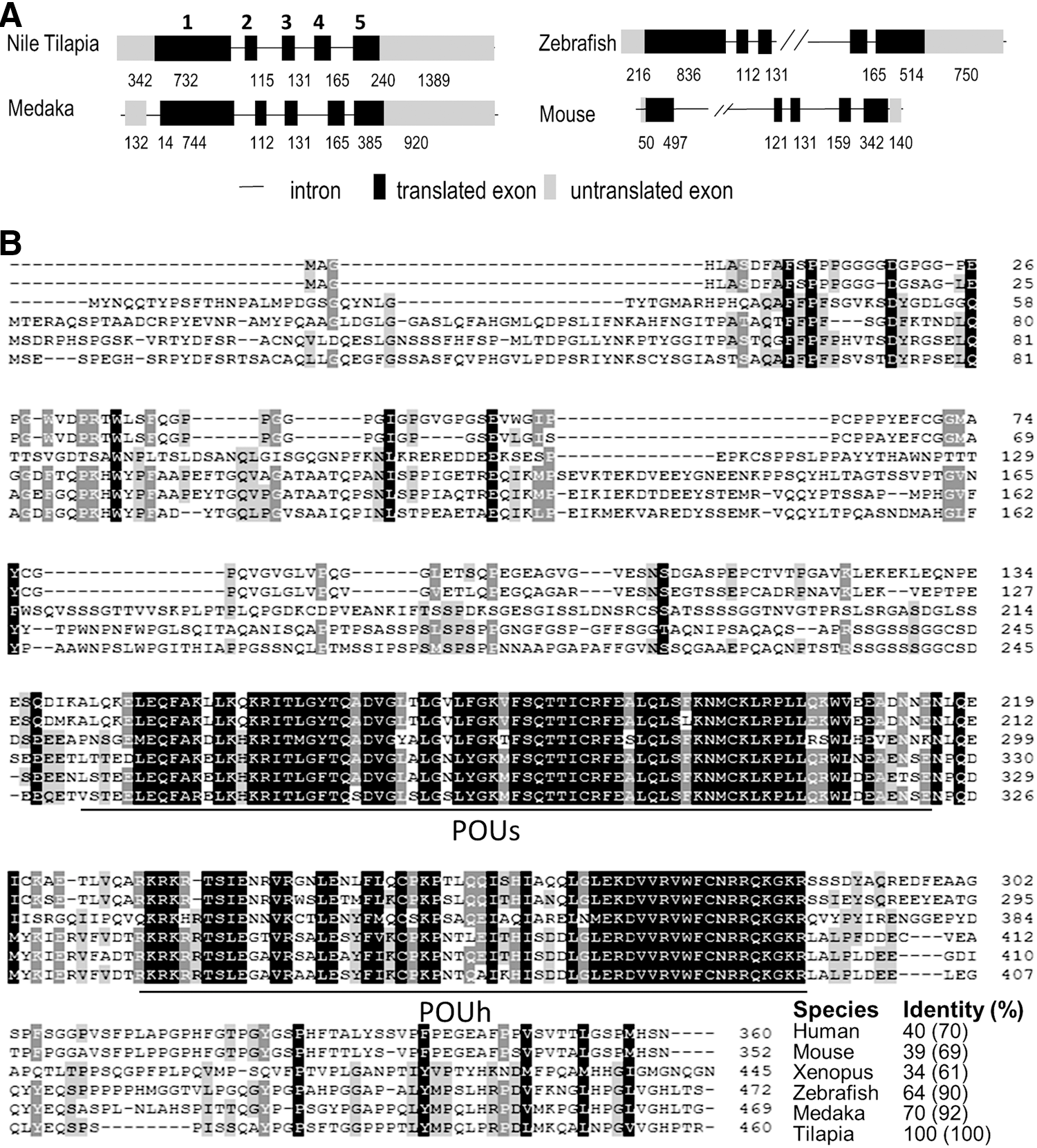
The Pou5f1 homologue genomic organization and amino acid sequence alignment.
In the phylogenetic tree, POU5F1 and POU5F3 formed two distinct clades. POU5F1 from eutherian mammals, Tammar wallaby, Platypus, and Axolotl was clustered into one clade, while OnPou5f3 together with the homologues of zebrafish and medaka was clustered into the other clade (Fig. 2A). A cross-species comparison of chromosomal location analysis indicates that the Nile tilapia pou5f3-containing region in LG 7 is syntenic to pou5f3-containing region in medaka chromosome 12 and zebrafish chromosome 21, respectively (Fig. 2B). This suggests that Onpou5f3 has good synteny to zebrafish and medaka. Collectively, Onpou5f3 is a true orthologue of zebrafish and medaka pou5f3.
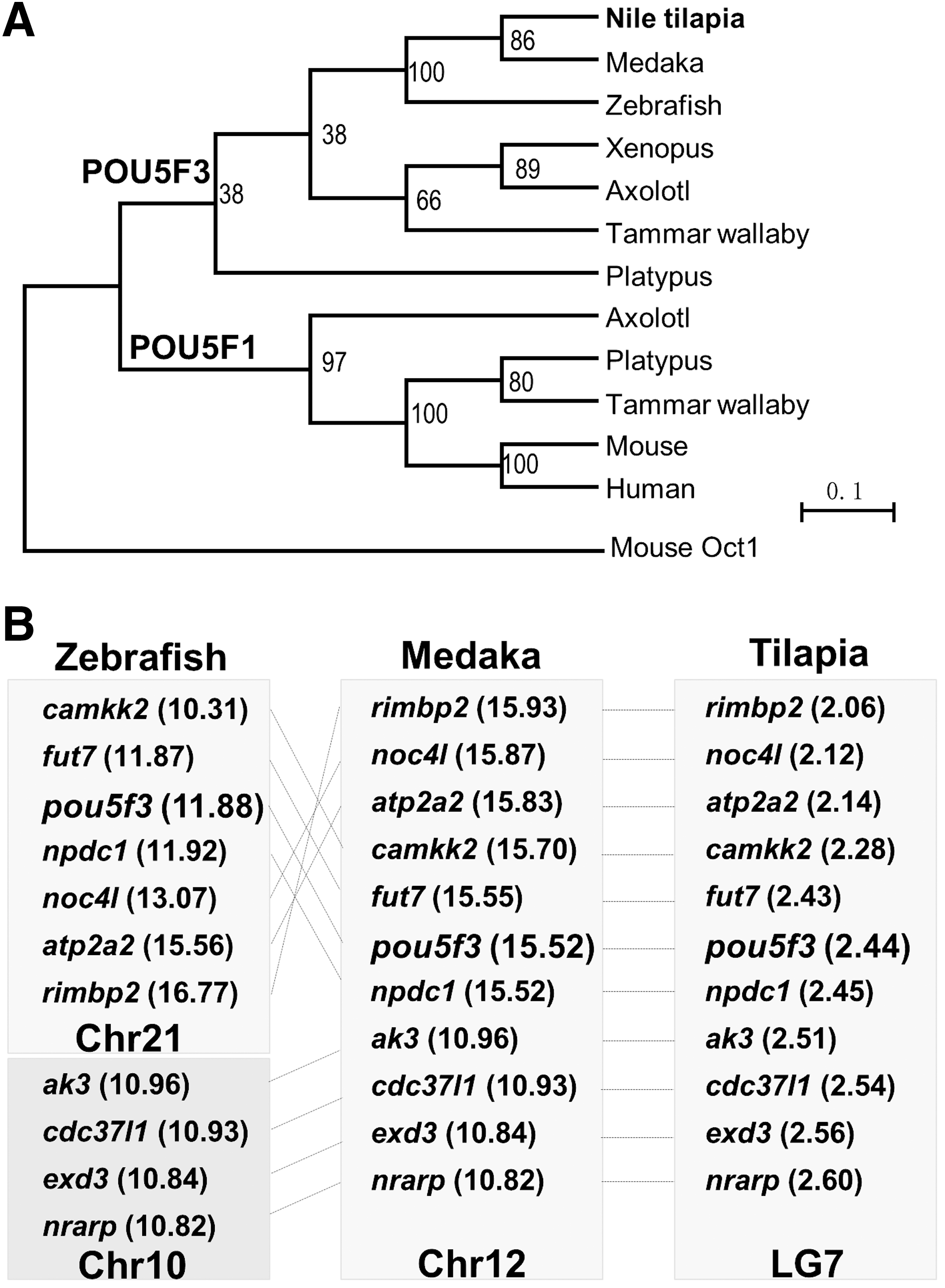
Phylogenetic relationship.
The reaction and specificity of the anti-Pou5f3 antibody
A rabbit polyclonal antibody (αPou5f3) was generated against a 16-aa peptide specific to OnPou5f3. This antibody detected one band in the testis and two bands in the ovary, but no band in the brain. One is ∼50 kDa, which is compatible with the predicted size 50.9 kDa for OnPou5f3, and the other is ∼75 kDa. Meanwhile, no obvious band was observed when the preserum of the rabbit was used (Fig. 3A).
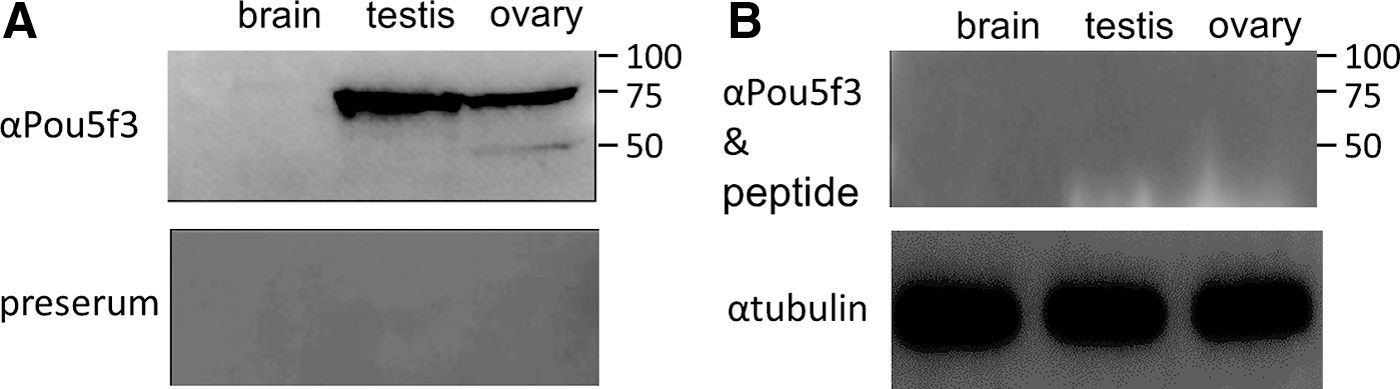
The reaction and specificity of the anti-OnPou5f3 antibody (αPou5f3).
To further verify the specificity, αPou5f3 was preabsorbed with the 16-aa peptide ENSENPQDMYKIERVF just before western blotting assay. As expected, both of the two bands at ∼75 and ∼50 kDa were vanished (Fig. 3B), suggesting that αPou5f3 has high specificity and there are two Pou5f3 isoforms or two Pou5f3-related proteins in the ovary of Nile tilapia.
Onpou5f3 expression in embryos
An RT-PCR analysis was undertaken to examine Onpou5f3 mRNA expression in embryos. The mRNA expression of Onpou5f3 was abundant until blastula stages (12 hpf, hours post fertilization), decreased at gastrulation stage onward (24 hpf), and became barely detectable except for transient increase at 52 hpf (Fig. 4A). The protein expression of OnPou5f3 in different stages of embryo development was detected by whole mount IF. Just like the homologues in mouse, zebrafish, and medaka, OnPou5f3 was detected in early developing embryo until blastula stages (8 hpf). At gastrulation onward (18, 45 hpf), the expression was diminished or barely detectable (Fig. 4B). At neurula stage, a second domain of OnPou5f3 expression evidently appeared (50 hpf), increased (52 hpf), and peaked (62 hpf) at the anterior part (the brain region) of the embryonic body (Fig. 4C), which was in agreement with the result by RT-PCR. OnPou5f3 expression in the brain region at neurula stage was further detected by IHC, where positive signals were observed in the cells of brain region (Fig. 4D). We did not detect any OnPou5f3 expression at 72 hpf (just before hatching). Meanwhile, no signals were observed at all different stages of embryo development using preserum (Fig. 4B-f).

OnPou5f3 expression in the different stages of embryo development.
Germline-specific expression in PGCs and adult gonads
After hatching, Onpou5f3 expression exhibited germline-specific pattern by RT-PCR analysis. That was high in the ovary and testis, but not detectable in all somatic tissues examined (Fig. 5A).
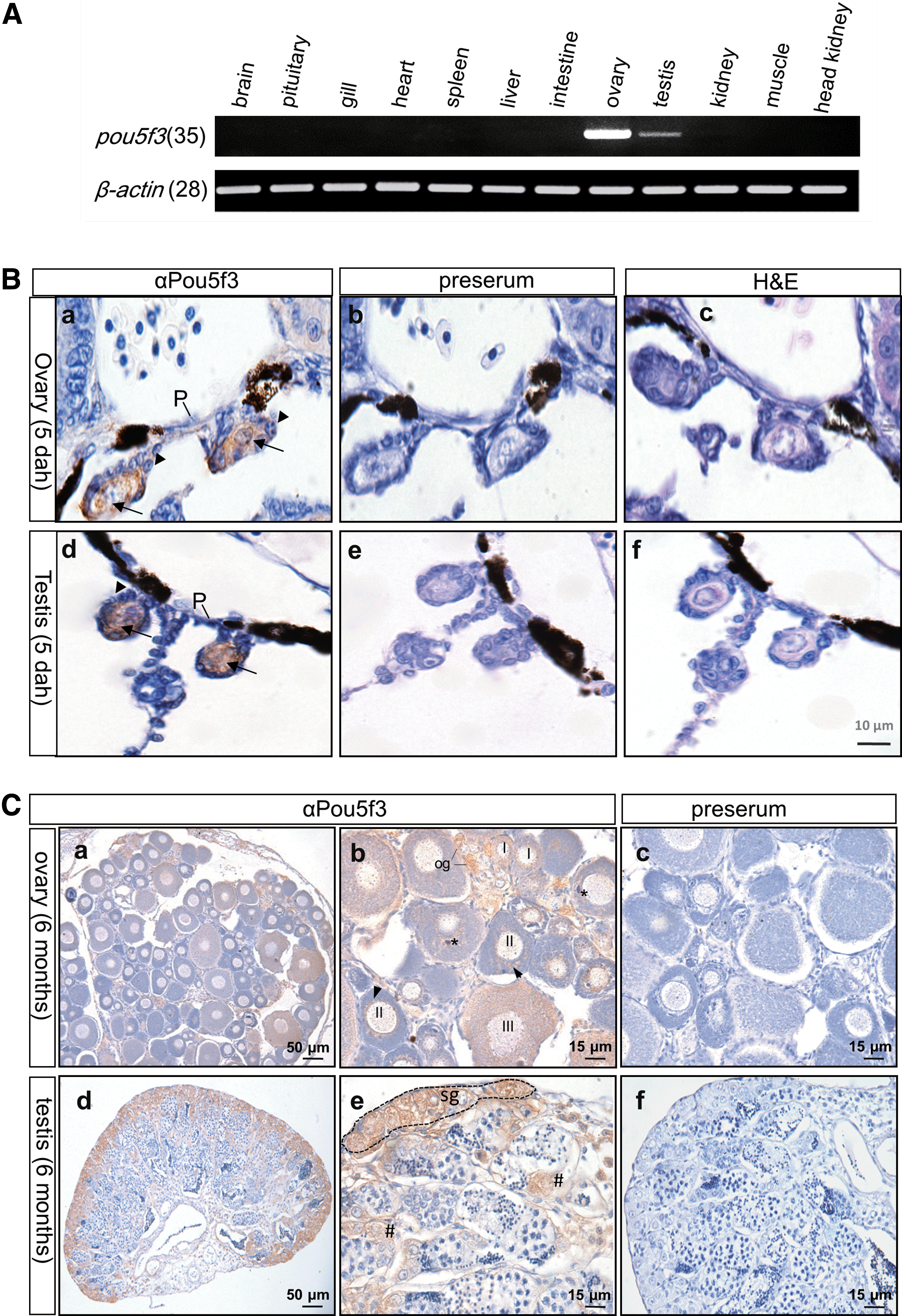
The germline-restricted expression of OnPou5f3.
The protein expression in germline cells was detected by IHC. Our results indicate that OnPou5f3 expression occurs at PGCs, but not somatic cells in 5 dah (days after hatching) female and male gonads (Fig. 5B).
In the adult ovary section, OnPou5f3 expression occurred in different stages of oocytes, where the signal was strong in oogonia and stage I oocytes. When oogenesis proceeded, OnPou5f3 protein expression reduced or diluted. In stage II oocytes, positive signals were predominantly around the nuclear and Balbiani body. In stage III oocytes, positive signals were observed evenly in the cytosol and nucleus (Fig. 5C).
In the Nile tilapia, a major part of undifferentiated spermatogonia is located at the most peripheral region close to the tunica albuginea, while a minor part of these cells spreads along the germinal compartment throughout the testis [24]. Our results indicate that OnPou5f3 expression is abundant in undifferentiated spermatogonia, which are mainly located at the most peripheral region, but barely detectable in primary spermatocytes and absent in spermatogenic male germ cells at more advanced stages (Fig. 5C).
A preserum did not produce any signal at both the ovary and testis sections (Fig. 5C-c, f), indicating the specific detection of OnPou5f3 protein expression by αPou5f3.
The biological activity of OnPou5f3
Both mouse POU5F1 and medaka Pou5f3 are essential for pluripotency in blastocyst/blastula formation and ES cell derivation, whereas zebrafish embryos completely devoid of maternal and zygotic Pou5f3 activity normally form blastula [11,16]. To analyze the pluripotency activity of OnPou5f3 in embryogenesis, a morpholino oligo that targets the Onpou5f3 mRNA (MOoct) was injected into the embryos at one-cell stage, while a morpholino with 5-base mismatch (MOoctm) was used as the control. The dissociated cells from the midblastula cells of embryos were detected for OnPou5f3 expression by IF. The cells from MOoctm-injected embryos had obvious OnPou5f3 staining in both nucleus and cytoplasm, suggesting no or less effect of MOoctm on the expression of OnPou5f3. The signals concentrated in the nucleus and displayed some punctuated staining in the cytoplasm (Fig. 6A-a1 to a3). Meanwhile, the cells from MOoct-injected embryos displayed barely detectable staining of OnPou5f3, suggesting that the morpholino oligo can specifically inhibit Onpou5f3 mRNA translation (Fig. 6A-b1 to b3).
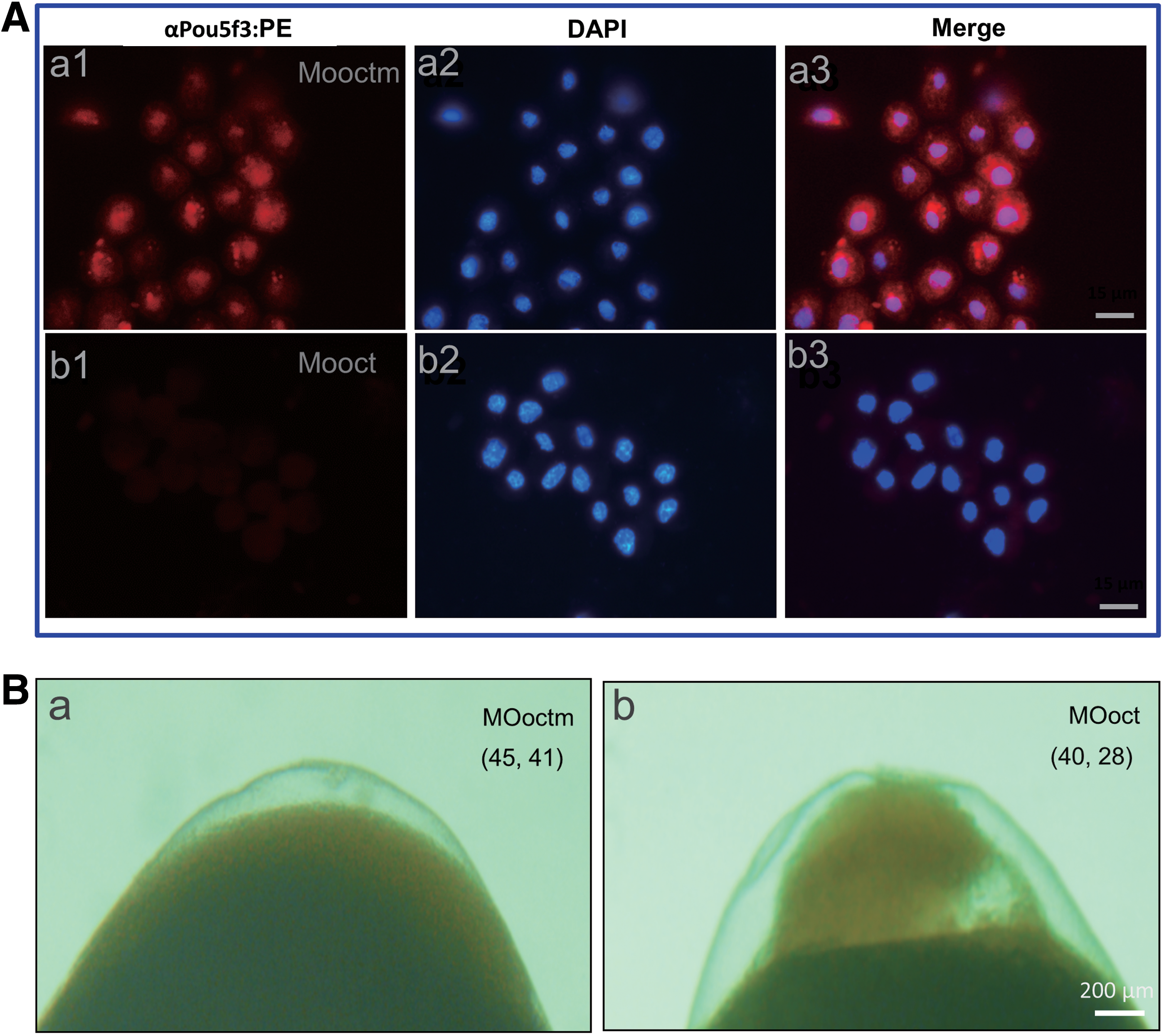
The effect of OnPou5f3 depletion on early embryo development.
Just like the noninjected control embryos, 91% of the MOoctm-injected embryos showed no abnormality and also developed to late blastula stage (n = 41). In contrast, 70% of the MOoct-injected embryos were blocked at the blastula stage as evidenced by the shrinkage of the blastoderm, deficiency of convergent extension, and enhanced apoptosis (n = 28) (Fig. 6B). Therefore, it is reasonable to deduce that OnPou5f3 is essential for blastula formation.
It has been documented that medaka Pou5f3 knockdown abolishes the ability for ES cell derivation [16]. In our preliminary study, stem-like cells derived from the MBEs were successfully developed and subcultured in ESM2 as previously described [16], which makes it possible to analyze the pluripotency-maintaining activity of OnPou5f3 directly in the Nile tilapia. Blastula cells were dissociated from the MOoctm- and MOoct-injected MBE and seeded in gelatin-coated 48-well plate containing ESM2, respectively. As expected, the cells dissociated from MOoctm-injected MBE attached and proliferated from 2 dpc (days post culture) (Fig. 7a–c). Moreover, most cells showed strong AP activity (Fig. 7d), which is extensively used as a valuable marker to monitor the pluripotency of ES cells [16]. In contrast, the cells dissociated from MOoct-injected MBE failed to attach and displayed no live cells at 2 dpc (Fig. 7f). Thus, OnPou5f3 is essential for the survival and proliferation of blastula cell-derived cells.
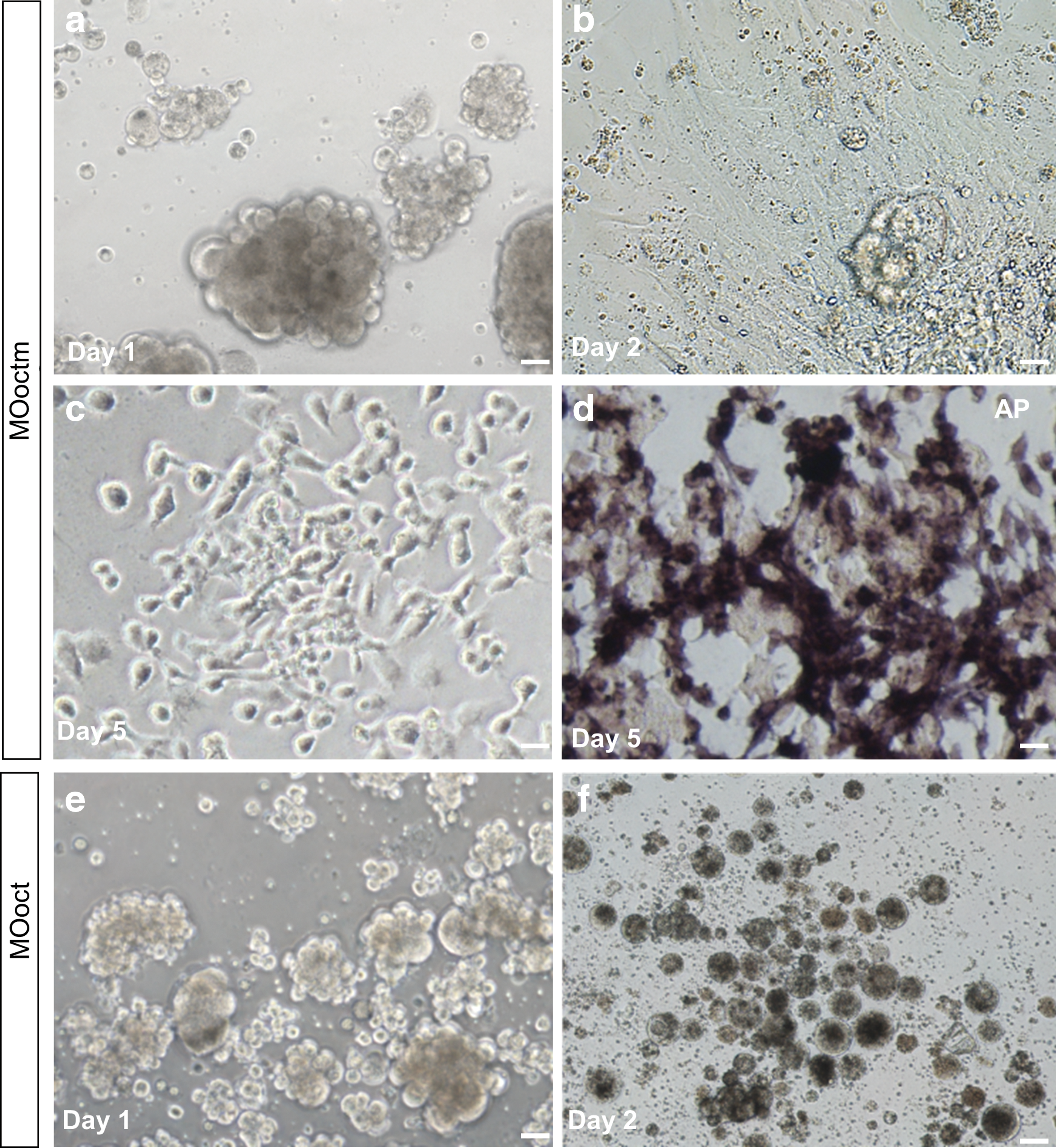
OnPou5f3 depletion on the survival and proliferation of blastula cell-derived cells. Embryos at the 1-cell stage were injected with MOoctm and MOoct at 10 ng, respectively; then the blastula cells were dissociated at the midblastula stage and seeded into 48-well plates.
Discussion
It has been unequivocally documented that the POU class V first appears in vertebrates; moreover, two homologues, namely POU5F1 and POU5F3, exist which presumably arose through gene duplication as early as the divergence of extant cartilaginous and bony fishes [13,14,25]. Through a complex and elusive evolution pattern, only one of the duplicated genes exists in some vertebrate lineages, that is, POU5F3 in fish, frogs, crocodilians, and birds, while POU5F1 in eutherian mammals and both POU5F1 and POU5F3 coexist in the others, including Marsupials and Monotremes [13,14]. However, to what extent the POU5F1 homologues, namely POU5F1 and POU5F3, are conserved during evolution remains unclear. Previous studies reveal that Pou5f3 in zebrafish and medaka shows considerable differences in expression and pluripotency-maintaining activity [11,15,16,26]. In this study, we report that Nile tilapia Pou5f3 exhibits expression and biological activity just like medaka but not zebrafish, whereas it displays a transiently evident expression domain in the brain region during the neurula stages similar to zebrafish but not medaka. Our study suggests that species-specific expression pattern of Pou5f3 exists besides equivalent OCT4-like expression and activity.
We reveal that OnPou5f3 is a true orthologue of POU5F3 by sequence and chromosome synteny. Just like the other POU5F1 homologues, Onpou5f3 exhibits a five-exon genomic structure. The putative protein OnPou5f3 shares high identity (more than 60%) to that of zebrafish and medaka. Moreover, it is clustered with both zebrafish and medaka Pou5f3 in the phylogenetic tree and shows good synteny to both of them through a cross-species comparison of chromosomal locations. Particularly, just like Pou5f3 in all species examined so far, Onpou5f3 is flanked by fut7 and npdc1 (Fig. 2B) [16]. It is previously described in medaka; multiple genes flanking Pou5f3 on chromosome 12 are homologous to those on human chromosome 9 and 12, but not homologous to those flanking POU5F1 on human chromosome 6. These data collectively suggest that OnPou5f3 is orthologous to zebrafish and medaka. In addition, Onpou5f3 appears as a single copy gene, which was verified through blasting NCBI database with the POU5F1/POU5F3 sequences from all the known vertebrates. What is the underlying evolutionary force, which might attribute to one or the other gene extinct in some vertebrate lineages, including eutherian mammals and fishes, is significantly worthwhile and challenging in the future study.
In our study, a rabbit antibody specific to OnPou5f3 (αPou5f3) was used to detect the protein expression of the gene. Four independent lines of evidence support that αPou5f3 has high specificity. First, the 16-aa peptide with the sequence “ENSENPQDMYKIERVF” used to produce the antibody is specific to OnPou5f3 and appears only once in the Nile tilapia genome. Second, by western blotting assay using crude protein extract from tissues, there are just one evident band in the ovary with ∼50 kDa compatible with the predicted size 50.9 kDa and another strong band with ∼75 kDa in both the ovary and testis, respectively, while no band in the brain (Fig. 3A). This result coincides with that by RT-PCR analysis (Fig. 4A). By the way, in medaka, a band with ∼75 kDa in ES cells and spermatogonia stem cells was also observed using the antibody specific to medaka Pou5f3 [16], suggesting that isoforms or posttranslational modifications might exist. Third, both the corresponding bands were completely vanished after incubating αPou5f3 with the synthesized 16-aa peptide (Fig. 3B), suggesting the high specificity of the antibody. Finally, positive signals were diminished through a morpholino oligo that targets the Onpou5f3 mRNA to interfere with the translation (Fig. 6A), further reflecting the specificity of the antibody from another perspective.
POU5F1 gene transcripts are deposited during oogenesis in the egg and its expression is closely associated with the cells at the pluripotency stages and during loss of pluripotency [2,4,25]. In zebrafish, pou5f3 (originally named pou2/spg or pou5f1/oct4) is maternally expressed, and the transcripts are present in all blastomeres until the midblastula stage and that the expression was restricted to the epiblast during gastrulation [12]. Afterward, it is expressed in the forming mid-hindbrain boundary and mediates the competence to respond to the Fgf8 inductive signaling in this region [18,27]. In medaka, Pou5f3 (originally as Oct4) expression occurs in all cells in early embryo development until blastula stages and afterward limited to a central group of cells and/or the posterior tip of the embryonic body, while no expression was detected in the brain region [15,16]. In the present study, our results show that the expression of Nile tilapia pou5f3 at mRNA and protein levels not only occurred during early embryo development until blastula stages but also transiently appeared in the brain region during neurula stages from 50 to 62 hpf and then faded just before hatching (Fig. 4). This is similar to the expression pattern in the brain region in zebrafish but not medaka. When development proceeded, the POU5F1 homologues in mouse, chick, and medaka are confined to PGCs and adult germline cells in the later stages of development, but not for the homologues in zebrafish or xenopus [11,16,28 –30]. In our study, OnPou5f3 also displayed a germline-restricted expression pattern, such as in PGCs in 5-dah female and male gonads, spermatogonia, oogonia, and oocytes, in different developmental stages (Fig. 5B, C). Confusingly, it has been reported that the expression of medaka pou5f3 was detected in the adult brain besides ovary and testis by RT-PCR [16]. In contrast, the expression of OnPou5f3 in the adult brain was not observed either by RT-PCR or by IHC (data not shown).
Mammalian POU5F1 is absolutely required for the pluripotency and self-renewal in ES cells and in the ICM from which ES cells are derived [2 –4,6]. Moreover, it is crucial for reprogramming differentiated cells to pluripotency [9,31]. In vivo, mouse POU5F1-deficient embryos fail to form the ICM and lead to unscheduled differentiation, suggesting its pluripotency-maintaining activity [5,32]. Similar to mammals, medaka Pou5f3 is crucial for blastula formation and ES cell derivation [16]. Nevertheless, zebrafish Pou5f3 is a lack of pluripotency-maintaining activity in heterologous system as evidenced by failure of replacement of other vertebrate homologues, but the reverse does work [25]. Furthermore, zebrafish maternal and zygotic null pou5f3 mutants could survive through gastrulation stages, suggesting that pou5f3 is dispensable for pluripotent cell division and proliferation until the midgastrula stage [11,12,18,27,33 –35]. In this study, our results reveal that Nile tilapia Pou5f3 is necessary for embryonic development in vivo, as well as survival and proliferation of blastula cell-derived cells in vitro. The protein expression of OnPou5f3 was depleted through morpholino-mediated translational interference (Fig. 6A). OnPou5f3 depletion can cause 70% of embryo blastula lethality or blockage, which might be due to the compromised streak expansion, deficiency of convergent extension, and enhanced apoptosis (Fig. 6B). Notably, the cells dissociated from OnPou5f3-depleted MBE were due to lack of adhesion and failed to survive, while the control cells adhered and proliferated (Fig. 7), suggesting that OnPou5f3 is indispensable for the survival and proliferation of blastula cell-derived cells in vitro. In contrast, combining the fact that some cells with ES-like characteristics, including strong AP activity, could be observed in the control and OnPou5f3 is an orthologue of Oct4, we speculate that OnPou5f3 might be necessary for pluripotency maintenance.
In conclusion, our data consolidate and extend the notion that equivalent OCT4-like expression and activity of Pou5f3 might be conserved accompanying with species-specific expression pattern and roles. As indicated in Fig. 8, except the expression in ES cells for zebrafish, brain for medaka and tail bud for Nile tilapia remain to be further investigated; both medaka and Nile tilapia Pou5f3 contain the combinational features for mouse (6/9) and zebrafish (4/9). This study is helpful for the understanding of the evolutionary conservation of POU5F1 homologues in vertebrates.

Comparison of the POU5F1 homologues among mouse, zebrafish, medaka, and Nile tilapia. +, presence; −, absence; -#, absence during brain regionalization, but presence in adult brain by RT-PCR [16]; ?, unknown.
Footnotes
Acknowledgments
We are grateful to Prof. Hong Yunhan at the National University of Singapore for helpful comments to this article. This work was supported by grant cstc2016jcyjA1844 from the Natural Science Foundation Project of CQ CSTC; grants XDJK2015A004 and XDJK2016E095 from Fundamental Research Funds for the Central Universities (Ministry of Education of China); and grants 31302170 and 31201979 from the National Natural Science Foundation of China.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.