
Abstract
Mesenchymal Stem Cells (MSCs) can be found in various body sites. Their main role is to differentiate into cartilage, bone, muscle, and fat cells to allow tissue maintenance and repair. During inflammation, MSCs exhibit important immunomodulatory properties that are not constitutive, but require activation, upon which they may exert immunosuppressive functions. MSCs are defined as “sensors of inflammation” since they modulate their ability of interfering with the immune system both in vitro and in vivo upon interaction with different factors. MSCs may influence immune responses through different mechanisms, such as direct cell-to-cell contact, release of soluble factors, and through the induction of anergy and apoptosis. Human MSCs are defined as plastic-adherent cells expressing specific surface molecules. Lack of MHC class II antigens makes them appealing as allogeneic tools for the therapy of both autoimmune diseases and cancer. MSC therapeutic potential could be highly enhanced by the expression of exogenous cytokines provided by transduction with viral vectors. In this review, we attempt to summarize the results of a great number of in vitro and in vivo studies aimed at improving the ability of MSCs as immunomodulators in the therapy of autoimmune, degenerative diseases and cancer. We will also compare results obtained with different vectors to deliver heterologous genes to these cells.
Introduction
M
In 2005, the new expression “multipotent mesenchymal stromal cells” (maintaining the acronym MSCs) was proposed to describe these fibroblast-like plastic-adherent cells. It was also proposed to use the term “MSCs” only for cells that meet specified stem cell criteria [4]. Caplan defined MSCs as “the sentinels and safeguards of injury.” He suggested that the main role of MSCs is protection from injuries and proposed to change their name into “Medicinal Signaling Cells” to keep the same acronym [5].
The discovery of a pool of multipotent cells present virtually in every tissue of adult animal raised great enthusiasm in the hope that they could be used in cell therapy, without limitations by those ethical concerns that have hindered the use of embryonic stem cells [6].
Among their properties, MSCs also exhibit immunomodulating ability. Their application in treating several pathologies has been largely investigated both in preclinical models and in clinical trials [
Since 2002, numerous attempts at improving MSC immunomodulatory potential by engineering them with various cytokines have been carried out [8 –10]. Several viral vectors have been used to deliver heterologous genes in vitro and in vivo to engineer MSCs (Table 1). Adenovirus (Adv) and retroviral vectors were used extensively to tweak MSC properties in experimental settings and most studies agree that such procedures are likely to ameliorate MSCs as cells in therapy [11] (Tables 2 and 3). Unfortunately, all these studies are at a preclinical stage; therefore, the major future goal will definitely be to test modified MSCs in clinics.
NOD-SCID, nonobese diabetic/severe combined immunodeficiency; ASNHL, autoimmune sensorineural hearing loss; EAE, experimental autoimmune encephalomyelitis; MSCs, mesenchymal stem cells; AT-MSCs, adipose tissue-derived MSCs; BM-MSCs, bone marrow-derived MSCs; WJ-MSCs, Wharton's Jelly derived MSCs; HGF, hepatocyte growth factor.
IL-12p40N220Q.
IFN-γ, interferon-γ; LV, lentiviral; NK, natural killer; TNF-α, tumor necrosis factor-α.
MSC Generation and Innate Properties
Sources
MSCs were initially isolated from the BM by a low-yield invasive process [12,13]. For this reason, other sources of cells were explored. Presently, MSCs may be derived from almost every tissue [14,15]: adipose tissue (AT) [16], lymphoid organs (thymus and spleen) [17], peripheral blood [18], endometrium and menstrual blood, scalp tissue, periodontal ligament, as well as placenta, amniotic fluid [19], umbilical cord blood (UCB) [20], and Wharton's Jelly (WJ) [21,22]. The therapeutic effect of hMSCs depends on many parameters such as tissue source of isolation, the methods of purification, and the animal model used to assess their efficacy; indeed, encouraging results in animal models do not always meet expectations in clinical trial, as discussed later [23]. Thus, even if MSCs derived from different sources may meet generally accepted criteria, they may differ in proliferation rate and differentiation potential [24].
Sources alternative to BM were also explored to attain higher proliferative potential from more accessible sites [15,19]. Currently, one of the main sources of MSCs is AT because it is readily available, the harvest procedure is less invasive than BM aspiration and the yield of MSCs is very high [16]; indeed, AT is 500 times richer in MSCs than BM [25]. AT-derived MSCs (AT-MSCs) proliferate rapidly in vitro and show low levels of senescence even after months of in vitro culture [26]. Nevertheless, isolation and expansion of AT-MSCs are somewhat less prone to automation in bioreactors.
Wharton's jelly-derived MSCs (WJ-MSCs) are also often used, since they are abundantly available after childbirth. Compared to the other sources, they are ontogenically primitive [21]. Most authors agree on the fact that they have higher proliferative potential compared to both AT and BM [15]. In contrast, Bortolotti et al. showed that hBM-MSCs grow faster in vitro and have higher proliferative potential than AT-MSCs even though they only survive to passage 12 of culture and then become senescent [23].
Definition
MSCs are generally characterized by expression of CD105 (SH2), CD73 (SH3 or SH4), CD90, CD166 (VCAM), CD44, CD 54/102 (ICAM1-2), CD 157, CD49, and Stro-1, and lack of CD11b, CD14, CD31, and CD45 (which identify hematopoietic or endothelial cells) [12]. Phenotypic markers alone are not sufficient to identify hMSCs unequivocally because the former can be expressed by other cell types as well; furthermore, during culture expansion, phenotypic marker expression can change, probably due to ageing of cells [27].
Different tissue origin, procedures of isolation, and culture conditions of MSCs in different laboratories produce heterogeneity of cell populations obtained [23]. This makes it hard to compare the biological properties and experimental outcomes in cell therapy trials. Therefore, the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy (ISCT) proposed a set of standards to define human MSCs unambiguously [28]. According to such criteria, MSCs are defined as a plastic-adherent population (when cultured in standard conditions), phenotypically expressing CD105, CD73, and CD90 for more than 95% of the population and lacking expression of CD45, CD34, CD11b or CD14, CD 19 or CD79a, and HLA class II (≤2% positive). Moreover, MSCs must be able to differentiate to osteocytes, adipocytes, or chondrocytes when cultured in appropriate differentiation media. These criteria apply only to human MSCs because cells derived from other species (such as rat and mouse) show different phenotypic markers [29].
All MSCs show similar phenotypes and functional features [15]. However, there are a number of differences that depend on their microenvironmental niche, original local function (stromal support of hematopoiesis in the BM vs. immune homeostasis by AT), and ontogenetic age (birth associated vs. adult) [15]. For instance, CD106 is significantly reduced on AT-MSCs compared to other MSCs [15]. Certain differences in potency may also exist; while AT-MSCs have multilineage differentiation potential in vitro and in vivo [30], there are conflicting reports regarding cord tissue-derived MSCs. Mattar and Karagianni reported that UCB-MSCs have significantly less adipogenic differentiation capacity [31]. Very recently, Reinisch also showed that only BM-MSCs are capable to form a functional hematopoietic niche [32].
All studies agree on the ability of all MSCs to carry out immunomodulation; in this respect, AT-MSCs proved to be more efficient than MSCs from other sources [24]. At present, comparative studies on MSCs do not consider the variables affecting their immunomodulatory efficacy, but only compare properties of cells in vitro, such as phenotypic markers, multipotency, and angiogenic ability ex vivo [23].
In conclusion, there is no consensus on the best source or culture method for MSC isolation because cells vary in their therapeutic efficacy depending on the model considered [6].
Properties
BM-MSCs support hematopoiesis by producing extracellular matrix components, such as cytokines and growth factors, in BM and most other tissues. Their main physiological role is to promptly mobilize and differentiate in response to signals of pathological or traumatic nature [33]. They actively respond to stress and injury, and can promote angiogenesis and regeneration.
Recently, pericytes were recognized as MSCs, that is, perivascular cells forming a unique niche, appointed to maintain the integrity of blood vessels. Pericytes can indeed be isolated from the BM and exhibit the same characteristics of MSCs [34,35]. They are positive for phenotypic markers CD10, CD13, CD44, CD73, CD90, and CD105 and negative for CD56, CD106, and CD133 [36]; in addition, they are able to differentiate into MSC lineages both in vitro and in vivo [14,36]. Perivascular location of MSCs is essential for their fast recruitment at the site of damage or inflammation.
MSCs downregulate activated immune cells and reduce damage to the site of inflammation; therefore, they may be considered one of the most important mechanisms of defense against excessive autoimmune reactions [5]. When stimulated by environmental signals of inflammation, MSCs release several molecules suppressing the immune response so that they can escape immune surveillance and easily migrate toward the site of injury [5]. Therefore, they can have many applications in immunotherapy, for example, in the treatment of degenerative and inflammatory pathologies.
Immunomodulatory properties have been widely demonstrated for all MSC populations, independent of their source of isolation, both in preclinical and clinical trials. However, AT-MSCs showed stronger effects compared to others models [24,37]. Since interaction of MSCs with cells of the immune system is very complex and often mutually dependent, many authors disagree on the mechanisms involved in immunomodulation; for this reason, the ISCT has also proposed standardized guidelines and protocols for their immunological characterization [38].
As mentioned above, MSCs may suppress or stimulate immunity depending on their surroundings. MSC immunomodulatory ability is not constitutive, but requires activation in a proinflammatory environment [6]. It has been shown that interleukin-1 (IL-1), interferon-γ (IFN-γ), and tumor necrosis factor-α (TNF-α) are involved in the priming of MSCs, which consequently interact with cells of both innate and adaptive immunity [6,7]. MSCs may alter cytokine secretion by dendritic cells (DCs), preventing their maturation and activation [39,40], and suppress T-cell and natural killer (NK) cell proliferation and their cytokine production [41] and B-cell functions [41,42].
MSCs may influence immune activity through different mechanisms, such as direct cell-to-cell contact, release of soluble factors, and through the induction of anergy and apoptosis [7]. Interaction with the immune cells takes place by expression of a large number of adhesion molecules, like VCAM-1, ICAM-1, LFA-3, HLA, and MHC-I, expressed on MSC surface [12,43]. Because they lack MHC-II and costimulatory molecules such as CD40, CD80, or CD86, MSCs are considered “immunoprivileged” cells and good candidates for allogeneic transplantation [44], with the caveat that, in inflammatory conditions, upregulation of MHC-II can increase their immunogenicity [45]. However, MSCs immunogenicity observed in certain studies may be due to the use of undefined serum or plasma for cell culture, rather than to MSCs themselves [46]. Currently, the question on MSC immunogenicity is being debated and under investigation.
MSCs release many soluble factors, including transforming growth factor-β (TGF-β), hepatocyte growth factor (HGF) [47], nitric oxide [48], indoleamine 2,3-dioxygenase [49], prostaglandin E2, interleukin-10 (IL-10), interleukin-1 receptor antagonist (IL-1Ra), interleukin 6 (IL-6), leukocyte inhibitory factor, HLA-G, hemeoxygenase, TNF-α, and many others. A pivotal role is played by IL-10 that supports the differentiation of regulatory T cells (T-regs). The ability of MSCs to induce T-regs has been observed both in vitro and in vivo [6,50 –52]. MSCs activate macrophages derived from circulating resident monocytes and initially polarize them to M1 profile [6,53]. Upon the influence of inflammatory cytokines released by M1 macrophages, MSCs start skewing macrophages toward the M2 subtype by a feedback mechanism [54].
Innate Immunomodulation Properties of MSCs and Their Exploitation in Clinic
The ability of MSCs to tightly interact with immune cells has been explored to treat many diseases where a pivotal role is performed by the immune system. In this study, we summarize the principal approaches.
Immunomodulation in the therapy of autoimmune and degenerative diseases
The first pioneering study on the effect of MSC administration in vivo was performed more than 10 years ago for the treatment of graft-versus-host disease (GVHD), a syndrome given by activation of donor T cells against recipient tissues after BM transplantation [55]. Severe acute GVHD is often refractory to immunosuppressants and has been reported to have an overall 2-year survival of 5%–30% [56]. In this regard, MSCs represent the main alternative to drugs. In the study mentioned above, haploidentical third-party MSCs were compassionately administered to a 9-year-old boy with steroid-resistant grade-IV GVHD, who completely healed after a year. The success of the therapy opened the way to clinical trials for treatment of GVHD with MSCs.
At present, phase II/III multicenter clinical trials have confirmed the efficacy of MSC treatment on adult and pediatric patients affected by steroid-resistant GVHD both in the USA and Europe. Most importantly, third-party-derived MSCs were shown to be safe as allogeneic cells [57,58]. As a result, the drug ProchymalTM (Osiris Therapeutics Inc., Columbia, MD), based on allogenic MSCs derived from the BM of adult donors, was approved by theHealth Canada to treat acute GVHD. Currently, Prochymal is being tested in two phase-III clinical trials on newly diagnosed patients (NCT00562497) and on steroidrefractory GVHD (NCT00366145). While the effect of MSC administration in acute GVHD has been demonstrated, their efficacy against chronic disease still needs further investigation; so far, chronic patients seem to be less responsive [59,60]. Therefore, efficacy ofMSC therapy for the treatment of chronic GVHD is still unclear.
After the extraordinary results obtained in acute GVHD, MSC therapy was also tested in the field of solid organ transplantation (SOT) as an alternative to immunosuppressive drugs, with the aim to prevent acute graft rejection and failure. Indeed, there are major concerns about currently used immunosuppressants; their side effects vary from acute and chronic nephrotoxicity, to hypertension, hepatotoxicity, and neurotoxicity, as well as an increased risk of gastrointestinal disorders and diabetes [61 –63]. Moreover, SOT is often followed by ischemia–reperfusion injury (IRI) that, in turn, causes systemic inflammation. This event can lead to delayed function and primary dysfunction of graft.
In the past, several studies performed in preclinical models showed the ability of MSCs to reduce IRI, preventing graft rejection, and promote immunotolerance in SOT [64 –68]. Since then, clinical trials have been carried out where MSCs proved to be successful alone or in association with low doses of immunosuppressive drugs in kidney transplantation [69,70]. Currently, a phase I/II clinical trial is testing MSCs for liver transplantation. However, preliminary results obtained in clinical trials are less satisfactory than expected on the basis of results on animal models.
MSCs have been used for the treatment of other autoimmune diseases, a category of pathologies where malfunctioning of the immune system is a determining factor. Administration of MSCs also appears to be a promising therapy in pancreatic islet transplantation for the treatment of type I diabetes, an autoimmune disease triggered by the immune response against autoantigens of β-cells. The effects of MSCs have been largely studied and clear evidence showed that MSCs can suppress autoimmunity against islets, support neoangiogenesis in damaged microvasculature of islets, and prevent acute rejection [67,71,72]. At present, more than 20 phase I/II clinical trials employing allogeneic MSCs or Prochymal for the treatment of diabetes are underway.
MSCs have been also tested in Crohn's disease, a chronic condition where the immune system attacks the gastrointestinal tract. A phase-I clinical trial has been performed by infusion of autologous MSCs in patients who did not respond to conventional treatment. Results showed an improvement of the disease with no significant adverse events either during BM harvesting or after MSC infusion; only mild allergic reactivity was observed in one patient probably due to dimethylsulfoxide used to cryopreserve cells. The administration of autologous BM-MSCs seems to be a safe and feasible therapy for the treatment of refractory Crohn's disease [73]. At the time of writing, Prochymal is being tested in Crohn's disease in a phase-III clinical trial (NCT01233960). Since Crohn's disease is often associated with the formation of perianal fistulae, autologous AT-MSCs have also been employed in their treatment with promising results and a phase-II clinical trial is being carried out [74].
Multiple Sclerosis (MS), a neurodegenerative pathology of the central nervous system (CNS) characterized by focal demyelination, neuronal damage, glial cell activation, and massive infiltration of immune cells, is thoroughly studied [75]. In the pathogenesis of MS, the inflammatory response is associated with an autoimmune reaction where both cellular and humoral components are activated against antigens of the CNS [76]. The neurodegenerative process, instead, is responsible for increasing disability in MS because it induces neuronal and axonal loss and leads to brain atrophy [77]. MSC transplantation has been proposed as a cell-based therapy for MS, in alternative to the one in use. In experimental models, MSCs seem to protect CNS from autoimmune demyelination; the observed therapeutic effect is thought to be mainly due to release of cytokines and chemokines, which suppress inflammation and enhance proliferation, migration, and differentiation of endogenous stem/progenitor cells [78].
Despite the promising results derived from experimental animal models of MS, the efficacy of MSC treatment in clinical trials is less evident. In a pilot study performed on 10 patients affected by MS, only 6 showed an improvement in the disease as a result of intrathecal injection of MSCs; in addition, 12 months after treatment, no improvement was evident by magnetic resonance [79]. Furthermore, in a phase-I and, more recently, in a phase-II clinical trial, intrathecal administration of MSCs induced modest improvement of only certain specific parameters related to vision impairment [80,81].
The most encouraging results have been obtained in the treatment of systemic lupus erythematosus (SLE), an autoimmune inflammatory disease with multiorgan involvement, including renal, cardiovascular, neural, musculoskeletal, and cutaneous systems. SLE is often refractory to conventional therapies based on corticosteroid administration [82]. Recently, SLE has been demonstrated to be a stem cell disorder caused by a defect in BM microenvironment [83]. Accordingly, MSCs derived from patients with SLE exhibited impaired proliferation, differentiation, and immune modulation capacities [84]. Moreover, the main features of SLE are alteration in cytokine homeostasis and defects in T-reg development [85]. In the light of such premises, allogeneic MSC transplantation seems the best approach for treatment of the disease. In a pilot study performed on SLE patients refractory to standard treatment, administration of allogeneic MSCs proved to ameliorate renal functions and restore T-reg cells, with no side effects observed [86].
Controversial results have been obtained by employing MSCs in preclinical models of rheumatoid arthritis, a systemic autoimmune disorder mediated by T cells and characterized by chronic inflammatory synovitis and progressive joint destruction. Alteration of autoreactive T cells and inflammatory cytokines such as TNF-α plays a pivotal role in the pathogenesis of the disease [87 –89]. Currently, no clinical trials have started.
In contrast, MSC therapy has proved clearly effective in the treatment of other inflammatory diseases, such as stroke [90,91], asthma [51], allergic rhinitis [92], lung injury [93], colitis, and sepsis. In this context, experiments in mice showed that MSC administration in murine models of experimental colitis and sepsis reduced the severity of the disease by inducing M2 polarization and the secretion of anti-inflammatory cytokines [94 –96]. MSC therapy has been also tested for the treatment of amyotrophic lateral sclerosis (ALS), a neurodegenerative disease that rapidly induces loss of motor neuron function. The pathophysiology of the disease is poorly understood, but it is believed that the immune response could also play a crucial role.
In a phase-I clinical trial performed with 10 patients, BM-MSCs were transplanted into the spinal cord; no side effects were observed [97]. In another phase I/II clinical trial, involving both ALS and MS patients, autologous MSCs were administered both intrathecally and intravenously to maximize homing to the CNS. In ALS patients, functional improvement was evident and only mild side effects were observed, probably due to residual dimethylsulfoxide. Moreover, an increase of CD4+, CD25+, Foxp3+, and T-regs and downregulation of activated lymphocytes and antigen-presenting cells were reported, whereas T-regs are normally reduced in patients with rapid disease progression [98,99]. The excellent outcomes and lack of safety concerns pave the way for other phase-II trials.
Immunomodulation in the therapy of cancer
MSCs are also able to adopt a proinflammatory phenotype [6,53]; they are able to recruit neutrophils through the production of chemokines and enhance their proinflammatory activity through the production of IL-6, IL-8, GM-CSF, and migration inhibitory factor [100,101]; in vivo murine MSCs strongly increase T-cell proliferation [48,102,103]. Since MSCs exhibit immunostimulatory properties, the application of MSCs in anticancer therapy has been explored. MSC homing ability toward the sites of injury is well known, but several studies demonstrated that they are also driven toward tumor sites and incorporated within the connective tissue stroma of tumors after intravenous injection [104]. Both in vitro and in vivo tests proved that MSCs home into glioma, both after intra-arterial and local intracranial delivery. This process is mediated by growth factors and chemokines [104,105].
MSCs injected in vivo have been shown to distribute mostly at the border between cancerous and normal parenchyma and also uniformly infiltrate into the tumor bed, while no MSCs were observed in healthy organs [8,104 –106]. At present, the mechanisms involved in MSC migration are unknown, but two possible alternatives have been hypothesized; homing of MSCs may be favored by the secretion of chemokines/cytokines by tumor tissues; alternatively, the interaction of cytokines or chemokines with their receptors might induce migration of MSCs toward the tumor microenvironment. Several studies have suggested that both cytokines, such as vascular endothelial cell growth factors (VEGF), TGFs, fibroblast and platelet-derived growth factors, monocyte chemoattractant, protein-1, and IL-8 [106,107], and adhesion molecules, such as β1- and β2-integrins and L-selectin, could be involved [108,109]. Furthermore, MSCs express several receptors like CXCR4, CX3CRl, and many others, which interact with their specific ligands [110]. CXCR4 and stromal cell-derived factor-1 (SDF-1) seem to play the main role, in the migration of MSCs toward the tumor site and in the pathology of glioma [111].
The extraordinary ability of MSCs to “sense tumor” has been exploited to deliver several anticancer gene products to tumor cells in vivo [8,112]; particularly 3 of the gene products have received the most attention: IL-2 [106], IFN-β [8,112], and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) [113].
Although MSCs look promising for cancer therapy, their antitumor effects are still controversial. Some authors showed that unmodified MSCs inhibit tumor growth and angiogenesis [114,115], while others report that MSCs support tumor development and metastasis [116]. Consequently, further preclinical studies are warranted before clinical trials can be started.
Enhancing MSC Immunomodulatory Properties by Exogenous Gene Transfer
The therapeutic potential of MSCs can be altered and routed by gene delivery. Indeed, delivery and expression of specific exogenous genes can yield MSCs releasing soluble factors, expressing specific receptors, conferring migratory ability or actions in response to specific stimuli, and so on.
The idea of using exogenous cytokines to, for instance, stimulate antitumor responses is not new. Direct administration of cytokines was attempted 20 years ago [117], whereas delivery of IL-2, IL-12, IFN-β, and yet other cytokines by viral vectors is more recent [118]. Both strategies have drawbacks; because of difficulties in reaching tumor sites and their short half-lives, free cytokines need to be inoculated at high doses and repeatedly, thus increasing their toxicity. In vivo injection of viral vectors, on the other hand, does not ensure specific targeting and is associated with safety concerns [118]. Thus, local cytokine delivery through engineered MSCs is a more promising, safe, and effective approach.
The main obstacle to the development of such MSC-based therapeutic approaches is the inherent resistance of MSCs to most transfection and transduction strategies. Transfection, which implies the use of plasmids, transposons, RNA, or other molecular constructs, can be achieved by treating cells with microprecipitates of nucleic acid–calcium complex, electroporation, liposomes, and so on. These methods have become routine and are successfully used on a wide variety of adherent and suspension culture cells [119]; furthermore, since molecular constructs do not integrate into the host cell genome, or are silenced/eliminated a few days after transfection, this approach presents a highly safe profile and is, therefore, preferable for clinical use. Unfortunately, the very poor performances shown with MSCs make these methods inapplicable for MSC-based therapeutic approaches.
Gene transfer in MSCs has been mostly carried out with vectors derived from Adv, adeno-associated viruses (AAV), and lentiviruses (LV). As detailed in Table 1, these vectors have pros and cons and provide a wide choice of approaches ranging from transient to permanent modification of cells, constitutive or inducible expression of the transgene (ie, the heterologous gene delivered by the vector), delivery of one or more transgenes, and so on. Detailed description of Adv, AAV, and LV vectors is available elsewhere [120]. In this study, we focus on same characteristics with particular reference to MSC transduction.
Adenoviral vectors
Adv vectors, (Fig. 1) transduce dividing and nondividing cells. Their DNA genome persists as an episome for a few days, ensuring transient high-level expression of the transgenes. For this feature, Adv vectors are considered safe, but gene modification is short lasting and is not transmitted to progeny cells. For these reasons, Adv vectors are used for vaccination purposes or transient cell manipulation [120 –122].
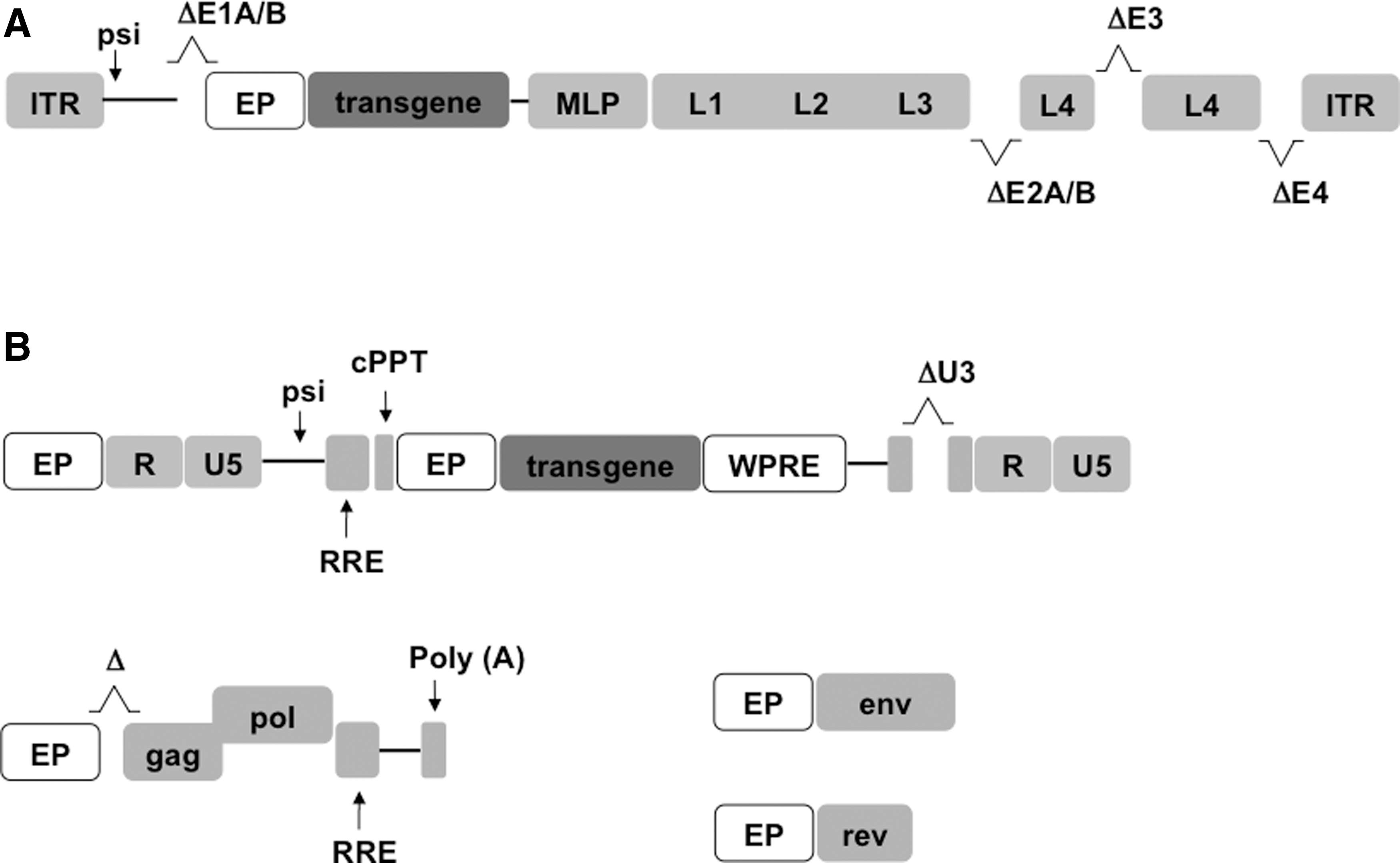
Schematic of adenoviral (Adv) and lentiviral (LV) vector genomes.
MSCs lack the Adv receptor and are therefore basically resistant to Adv vector transduction [123]. High vector/cell ratio is therefore necessary to transduce hBM-MSCs possibly causing various toxic effects and strong immune reactions in vivo [8]. Ongoing studies to improve efficiency of MSC transduction aim to generate recombinant vectors bearing other viral receptors [124 –127] or using cationic polymers, chelating agents, or small peptides to reduce electrostatic repulsions [128 –130].
Adeno-associated viral vectors
Parental AAV viruses infect a wide range of cells in vivo and insert their viral genome in a specific and harmless site of the cellular genome. AAV vectors transduce both dividing and nondividing cells, exhibit minimal immunogenicity and toxicity, but having a small genome, can transport minute amounts of heterologous genetic material, and owing to deletion of a substantial part of the viral genes, have lost their capacity to integrate into the host cell genome (Table 1). Despite these limitations, AAV vectors elicit long-term transgene expression in vivo and are used in a large number of clinical trials.
Early studies showed that AAV vectors can transduce MSCs without altering their multipotential ability but with low expression of transgenic product [131 –133]. Therefore, there is ample room for improving such vectors [134 –137].
Lentiviral vectors
LV vectors transduce both quiescent and dividing cells and integrate their vector genome into the host cell genome, thus ensuring long-lasting expression of the transgene [138]. For these and other features, shown in Table 1, they are the most widely used vectors in gene therapy. Transduction of MSCs with LV vectors has been extensively investigated [11,139]. In their simplest version (ie, single transgene controlled by a standard cytomegalovirus immediate early promoter) and using a low vector/cell ratio, LV vectors transduced over 25% human BM-MSCs and expressed the transgene for over 5 months in vitro [139]. Also, transduction did not alter MSC phenotype, in vivo homing, viability, proliferation of cells, or ability to differentiate into adipocytes and osteocytes [11,140,141].
Like AAV vectors, MSC transduction went up to 95% with LV vectors bearing different constitutive or inducible promoters [11,142], using synthetic polymers to reduce electronic repulsion [140] and/or, a second transgene to sort transduced cells [143,144]. High efficiency of transduction and the capacity to carry two or more transgenes make LV vectors the system of choice for MSC transduction as it permits, for instance, to deliver reporter genes to monitor localization, viability, and differentiation pathway of the engineered cells [9,145].
Alteration of MSC Immunomodulation Properties by Gene Transfer
MSCs have been transduced with the vectors described above and carrying transgenes capable of enhancing or counteracting immunosuppressive properties of naive MSCs. In the following sections, we summarize the main studies performed and the results obtained.
Enhancing MSC immunosuppression
These studies are based on the delivery of the following anti-inflammatory cytokines (also enlisted in Table 2).
Interleukin-1 receptor antagonist
IL-1α and IL1-β play a pivotal role in chronic inflammation and in the activation of innate immunity. IL-1α is released by a variety of cells undergoing necrosis, for example, after epithelial damage, myocardial infarction, stroke, tumor decay, or acute renal failure, and may be considered an “alarmin,” since it induces production of IL-1β, TNF-α, and IL-6 [146,147]. IL-1β is released by monocytes, macrophages, neutrophils, and DCs. It is a potent proinflammatory cytokine locally causing vasodilatation, granulocyte chemotaxis into damaged tissue, and expression of prostaglandins. IL-1β also brings about activation of the proinflammatory T helper (Th) 17 cells and the development of chronic inflammation [146]. IL-1β plays a pivotal role in the pathophysiology of various inflammatory conditions, such as rheumatoid arthritis, prosthetic aseptic loosening of arthroplasty, acute liver failure, hereditary autoinflammatory diseases, rheumatoid arthritis, osteoarthritis, type 2 diabetes mellitus, cardiovascular disease, and many others [146,148,149].
The IL-1Ra can be considered an antagonist of the proinflammatoy cytokines IL-1α and IL1-β, since it binds their receptor blocking IL-1α and IL-1β signaling [150]. IL-1Ra is expressed in all tissues and is present in serum at high levels. It is able to prevent uncontrolled activation by IL-1β both in vitro and in vivo, thereby reducing inflammation, but high concentrations are required [151]. In healthy conditions, there is a balance between the proinflammatory activities of IL-1α and IL-1β and the ability of IL-1Ra to control their production; alteration of this equilibrium can induce inflammatory diseases [150]. IL-1Ra could be valuable to treat inflammatory diseases; indeed, the infusion of exogenous IL-1Ra is being evaluated in clinical trials against GVHD and MS.
MSCs are also able to secrete IL-1Ra [152]. They have an improved anti-inflammatory effect compared to administration of exogenous IL-1Ra [153]. Moreover, combined therapy with IL-1Ra and MSC is more efficient than the use of the two separate treatments [154]. Thus, engineering of MSCs with IL-1Ra (MSCs/IL-1Ra) potentiates IL-1Ra release. BM-MSCs/IL-1Ra have been tested both in in vitro and in vivo models of several diseases (Table 2). hBM-MSCs transduced by Adv vectors expressing IL-1Ra in association with HGF (hBM-MSCs/HGF/IL-1Ra) or VEGF (hBM-MSCs/VEGF/IL-1Ra) have been studied in islet transplantation for the treatment of type I diabetes [155,156].
Because IL-1β plays a key role in the loss of islet viability after transplantation [157], systemic administration of MSCs or their local injection in transplanted islets partially reduces inflammation, promotes revascularization, and supports graft survival. However, they were not enough to completely prevent apoptotic cell death, and more than 70% of islets were lost in the first weeks after transplantation [158,159]. It was demonstrated that engineered cells were able to secrete high levels of IL-1Ra and HGF/VEGF and, by acting in synergy, cytokine-expressing MSCs prevented apoptosis of islet cells and improved islet engraftment, thus supporting long-lasting normoglycemia [155].
IL-1β hampers cartilage and bone regeneration [146]; together with TNF-α, it hinders MSCs differentiation to chondrocytes [160]. Accordingly, an in vitro study demonstrated that MSCs engineered with IL-1Ra by an inducible LV vector were able to support MSC chondrogenic differentiation, also in the presence of IL-1β [161]. The effect of MSC therapy combined to exogenous administration of IL-1Ra resulted effective also in the treatment of acute liver failure, a pathology characterized by severe liver cell damage [162]. MSCs/IL-1Ra have not yet been tested in this context.
Interleukin-10
IL-10, a pleiotropic cytokine, is considered to play a key role in quenching immune responses and is one of the most important soluble factors secreted by MSCs [163]. IL-10 has multiple immunoregulatory effects, such as downregulation of MHC-II and costimulatory molecule expression; inhibition of DC maturation and function; and secretion of proinflammatory cytokines and chemokines [164]. In vivo, IL-10 secreted by naive MSCs improved organ function and survival in a sepsis experimental model.
Since genetic delivery of IL-10 was shown to improve allograft survival in many animal models of heart or liver transplantation [165,166], MSCs genetically engineered with IL-10 were tested in a rat model of orthotopic liver transplantation. The injection of MSC/IL-10 significantly improved graft survival by increasing the expression of T-reg cell-associated cytokines (IL-10 and TGF-β1) and decreasing the expression of Th-17 cell-related cytokines (IL-17, IL-6, IFN-γ, TNF-α, and IL-23) [167]. Indeed, it was demonstrated that T-regs play an important role in inhibiting acute graft rejection and in the prevention of autoimmune diseases [168,169]. The ability of MSC/IL-10 to prevent liver transplant rejection was higher than the effect observed with naive MSCs.
In vitro studies were performed with hWJ-MSCs transduced with human IL-10 (hIL-10) or Epstein-Barr virus IL-10 (v-IL10) [9,170]. vIL-10 is similar to hIL-10, but only has the immunosuppressive domain of the cytokine [169 –173]. hWJ-MSC/vIL10 showed enhanced ability compared to naive MSCs to release IL-10 in solution and increased immunosuppressive properties. They were able to act both on innate immunity by decreasing TNF-α production by monocytes upon LPS stimulation and on adaptive immunity by reducing IFN-γ release by CD3-stimulated T lymphocytes [9].
Interleukin-4
IL-4, like IL-10, is a pleiotropic cytokine, involved both in innate and adaptive immunity. IL-4 is secreted by Th2 cells together with IL-10 and IL-13 to quench inflammation. It is also able to stimulate proliferation of B and T cells and foster differentiation of naive Th0 into Th2. Besides, IL-4 stimulates T-reg proliferation and chemotaxis [174].
Exogenous administration of IL-4 has been studied in several experimental models of autoimmunity where Th2 cells mediate disease remission, such as experimental autoimmune encephalomyelitis (EAE) as a model for MS [175 –177]. Exogenous administration of IL-4 was shown to ameliorate EAE in mice through a Th1 to Th2 shift, but repeated administration of high doses of IL-4 were required [178]. Subsequently, it has been demonstrated that injection of recombinant plasmid, Adv, and herpes simplex viral vectors encoding IL-4 in mice with EAE favored remission of the disease [175,179 –181]. Following these studies, AT-MSCs transduced with IL-4 by LV and transplanted at the onset of EAE were shown to be able to reduce the inflammatory immune response by shifting cytokine profile from Th1 to Th2 [182].
Pig BM-MSCs engineered with IL-4, injected into the inner ear of a guinea pig model of an autoimmune inner ear disease, were broadly distributed in the inner ear restoring hearing function [183].
Hepatocyte growth factor
HGF is a multifunctional cytokine involved in organogenesis, angiogenesis, cell proliferation, survival, and motility. Recently, it has been demonstrated to have regenerative properties in myelodegenerative diseases and to be able to improve acute GVHD, suggesting that HGF also has immunomodulatory effects [184,185]. It has also been shown to act additively with TGF-β to suppress T-cell proliferation in vitro [186].
hMSCs express varying amounts of HGF depending on their microenvironment [187]. They have been tested in several disease models after transduction with HGF. rBM-MSCs transduced with HGF were used in a rat model of liver transplantation and proved to significantly reduce graft rejection by decreasing the production of IL-2 and INF-γ and increasing IL-4 and IL-10 [188]. hAT-MSCs/HGF have also been employed for the treatment of radiation-induced lung injury, where again, they were able to reduce secretion of TNF-α, IFN-γ, IL-6, and TGF-β and increase the expression of IL-10, thus not only reducing inflammation but also inhibiting lung fibrosis [189]. hUC-MSCs/HGF were also used for the treatment of acute kidney injury in rats, where HGF delivery improved IRI through antiapoptotic and anti-inflammatory mechanisms, thus providing a novel therapeutic application for hUC-MSCs in acute kidney injury [190].
Interleukin-12p40
Interleukin-12p40 (IL-12p40) homodimer is an antagonist of two potent proinflammatory cytokines IL-12 and IL-23 [188 –190]. The latter are involved in inflammatory bowel disease, a chronic disorder of the digestive system that includes Crohn's disease and ulcerative colitis [191,192].
The effect of MSCs transduced by IL-12p40 has been investigated in dextran sulfate sodium salt-induced colitis, an animal model of bowel disease. Injection of Adv/IL-12p40 or MSC/IL-12p40 efficiently reduced colitis and tissue damage, thus increasing survival rate. IL-12p40 reduced both IFN-γ and IL-17 production brought about by IL-12 and IL-23, respectively. This effect was more pronounced after MSCs/IL-12p40 than Adv/IL-12p40 administration [193].
Interleukin-13, -27, -35
Interleukin-13 (IL-13) is a pleiotropic cytokine very similar to IL-4 both in structure and function [194]. As mentioned above, it is secreted by Th2 lymphocytes and contributes to the etiology of allergic reactions. Different from IL-4, T cells are generally unresponsive to IL-13 due to the absence of IL-13Ra1 expression on their surface [195]. Allogeneic MSCs transduced with IL-13 have been transplanted to modulate allograft-specific immunity in a murine model. It has been shown that MSCs/IL-13 injection in muscle and brain tissue prolonges allogeneic cell graft survival [196].
Interleukin-27 (IL-27) and interleukin-35 (IL-35) are new members of the IL-12 family. Because of their properties, both of them could be used in the therapy of autoimmune diseases. Recently, in vitro works have demonstrated that transduction of hAT-MSC and hWJ-MSC with IL-27 and IL-35 does not alter naive MSC properties [10,197].
Enhancing MSC immunostimulation
MSCs have also been engineered with immunostimulating/proinflammatory cytokines mainly for the treatment of different kinds of solid tumors. In the following section, we summarize the results obtained by using the cytokines shown in Table 3.
Interferon-β
Interferon-β (IFN-β) is a pleiotropic cytokine with a wide range of biological activities such as regulation of cell proliferation, innate and adaptive immunity, hematopoiesis, angiogenesis, and yet other functions. Most importantly, it can promote tumor regression [198,199].
In 2002, Studeny used IFN-β-transduced hBM-MSCs to treat mice bearing human melanoma. He demonstrated for the first time that MSCs were integrated into the tumor and released high levels of IFN-β in situ. Besides, hBM-MSCs/IFN-β inhibited melanoma growth both in vitro and in vivo [8]. This study can be considered a groundbreaker in the field of MSC-based tumor-targeted gene therapy and opened the way to many others studies where MSCs engineered with different proinflammatory cytokines have been used to suppress solid tumors in animal models.
Interleukin-2
Interleukin-2 (IL-2) is secreted by CD4+ and CD8+ T lymphocytes and NK cells and is essential for T-cell proliferation. It is considered a proinflammatory cytokine because it can foster expansion of NK cells, CD8+ T cells, and T-regs. Because of its immunostimulating properties, IL-2 has been tested in immunotherapy against several metastatic tumors. It was demonstrated that systemic administration of high doses of recombinant IL-2 in humans caused regression of metastatic melanoma, kidney cancer, and non-Hodgkin's lymphoma [200]; therefore, IL-2 was approved for the treatment of metastatic renal cell carcinoma and melanoma by FDA [201 –203].
To enhance the antitumorigenic effect of IL-2, rBM-MSCs engineered with IL-2 were tested in a rat model of glioma. Intracranial injection of such cells prolonged the survival of the recipients and reduced tumor mass to a greater extent than naive rBM-MSCs [106].
Interleukin-12
Interleukin-12 (IL-12) is a powerful activator of NK cells and macrophages. It also skews cellular immune responses toward a Th1 response and is produced by DCs and macrophages in response to activation [204,205]. IL-12 inhibits tumor growth and prolonges survival of different tumor bearing animals [206,207]. Exogenous IL-12 administrated in phase-I and II clinical trials for cancer therapy induced high levels of IFN-γ and increased susceptibility of tumor cells to cytotoxic T lymphocytes. For this reason, it is now considered the main antitumoral cytokine [208 –210].
BM-MSCs engineered with IL-12 were tested in experimental mouse models of melanoma, breast tumor, and hepatoma. Results showed that intravenous injection of BM-MSCs/IL-12 markedly reduced progression of metastases both in midway and late tumor stages, with no significant toxicity. The effect observed with BM-MSCs/IL-12 was 10-fold higher than the one produced by Adv/IL-12 administration; in addition, the latter failed to favor regression of end-stage tumors [211]. Again, BM-MSCs/IL-12 were able to migrate into the tumor site where they became part of the tumor architecture and released high levels of IL-12, thus stimulating the immune response inside the tumor.
In another study, hBM-MSCs engineered with mIL-12 were tested in a mouse xenograft model of renal cell carcinoma. Systemic administration of hBM-MSC/mIL-12 reduced renal cell carcinoma growth and significantly prolonged mouse survival. Intratumor IFN-γ levels were significantly higher after hBM-MSC/mIL-12 administration, thus suggesting that IFN-γ plays a key role in tumor regression. Furthermore, the observed antitumor effect of hBM-MSC/mIL-12 was mediated by NK cells [212]. In agreement with these results, it was also shown that mBM-MSCs/mIL-12 were also effective in the treatment of Ewing's sarcoma [213].
hUC-MSCs/IL-12 were evaluated in the SKOV3 experimental model of ovarian cancer. In vitro, the supernatant of IL-12-engineered hUC-MSCs was able to significantly inhibit proliferation of SKOV3 cells and trigger cellular apoptosis. In vivo, hUC-MSCs/IL-12 inhibited tumor growth in a nude mouse model [214].
More recently, a modified form of IL-12 (IL-12M) has been used to transduce rBM-MSCs. IL-12M was obtained by N-glycosylation of IL-12 at Asn220. This change reduces secretion of the p40 subunit and enhances antigen-specific T-cell responses, without altering IFN-γ secretion and IL-12 receptor binding affinity [215]. The effect of rBM-MSCs/IL-12M after intratumoral, intravenous, and subcutaneous administration was evaluated in two experimental models of melanoma and cervical cancer. Pronounced regression of tumors was observed, especially after following intratumoral injection, suggesting that this route of administration has great relevance in therapy. rBM-MSCs/IL-12M were more efficient than either Adv-IL [216]. In a different study, in vitro work suggested the ability of AT-MSCs transduced with LV/IL-12 to reduce lung adenocarcinoma ability of migration and invasion [217].
Taken together, the results from these studies suggest that IL-12-engineered MSCs are attracted by several tumor microenvironments, where they cause release of high levels of IFN-γ triggering a tumor-specific T-cell response. The antitumor effect is stronger than with IL-12 delivered by Adv vectors and the toxicity is significantly lower.
Interleukin-15
Interleukin-15 (IL-15) is functionally similar to IL-2, but does not stimulate T-reg proliferation. However, it does promote long-lasting antitumor T-cell immunity [217 –220]. Because IL-15 administration has shown antitumor effects in many animal models and in human clinical trials [221,222], the effect of hUCB-MSCs transduced with mIL-15 was investigated in an experimental model of pancreatic adenocarcinoma highly resistant to chemotherapy. The results showed that UCB-MSCs/mIL-15 systemically infused into syngeneic mice bearing Pan02 pancreatic tumors were able to migrate into the tumor site, where the secretion of IL-15 fostered activity by both NK and CD8+ T cells, thus inhibiting tumor growth. Interestingly, mice treated with UCB-MSCs/IL-15 became resistant to Pan02 pancreatic tumor rechallenge, inferring that MSCs/IL-15 were able to augment long-term tumor-specific T-cell immunity [223].
Interleukin-18
Interleukin-18 (IL-18), also known as IFN-γ-inducing factor, is similar to IL-1 both in structure and functionality; it is able to stimulate innate immunity, Th1- and Th2-mediated responses, and NK activity [224,225]. IL-18 is a potent antitumor cytokine, by inducing tumor cell apoptosis and inhibiting tumor angiogenesis [226].
IL-18 was shown to favor immune cell migration in gastric cancer and melanoma [227,228]. To test its effects on glioma, rBM-MSCs transduced with IL-18 have been tested in vitro and in vivo in a glioma bearing rat model. It was shown that rBM-MSCs/IL-18 inhibited glioma growth and prolonged the survival of glioma bearing rats. The study also showed that the antitumor effect of rBM-MSCs/IL-18 is carried out through the enhancement of T-cell cytotoxic activity [229]. Unfortunately, complete suppression of tumor growth could not be achieved; hence, to extend the efficacy of such treatment, rBM-MSCs were transduced with a bicistronic LV vector encoding IL-18 and IFN-β. In this case, inhibition of glioma growth was enhanced, demonstrating that IL-18 and IFN-β act synergistically [230].
Finally, the effect of hUC-MSCs/IL-18 was investigated in a model of breast cancer in vitro; engineered MSCs were able to suppress proliferation, migration, and invasion of breast cancer cells [231,232].
Interleukin-21
As an immunostimulating cytokine involved in T- and NK-cell activation, interleukin-21 (IL-21) is also able to induce strong cell-mediated immune responses in antitumor vaccine approaches [233]. It has been tested as a therapeutic agent in several animal models of cancer and in clinical trials, against tumors such as metastatic lymphoma, melanoma, ovarian cancer, and others [233,234].
More recently, IL-21-transduced hUC-MSCs (MSCs/IL21) were injected intratumorally in mice bearing SKOV3 ovarian cancer. Results showed that hUC-MSCs/IL-21 had potent antitumor effects mediated by IL-21 release and upregulated of NK activity through IFN-γ and TNF-α secretion [235]. MSCs/IL-21 have also been used in the treatment of B-cell lymphoma; MSCs/IL-21 intravenously injected were able to delay tumor incidence and improve survival through upregulation of NK and effector T cells [236].
Interleukin-28
Interleukin-28 (IL-28) belongs to type III IFN family, together with IFN-λ1, -λ2, and -λ3 (also known as IL-29), IL-28A, and IL-28B [237,238]. It has been shown that type III IFNs can exert antiproliferative activity and promote apoptosis in tumors such as lung and esophageal carcinoma [237,239]. Moreover, they proved to not only activate NKs and DCs but also to have antiangiogenetic effects [238,240,241].
Recently, hBM-MSCs transduced with IL-28A were tested for their antitumoral activity in mice bearing human esophageal carcinoma or colon carcinoma. While naive hBM-MSCs did not display any antiproliferative tumor effect, hBM-MSCs/IL-28A inhibited tumor growth and vascularization and stimulated NK activity to a greater extent than IL-28A alone [125].
Tumor necrosis factor-α
TNF-α has a pivotal role in the early immune response, it can activate both macrophages and neutrophils, and is involved in the maturation and activation of DCs. It is also a critical cytokine for promoting Th17 cell differentiation. TNF-α and CD40 ligand (CD40L) have been used to transduce MSCs and the immunomodulatory effects of engineered MSCs have been investigated in vitro and in vivo in animal models of human breast cancer [242].
In vitro results showed that MSCs/TNF-α/CD40L induced proinflammatory responses through maturation of DCs, increase of TNF-α, IL-6, and IL-12 levels, and reduction of TGF-β, IL-4, and IL-10 levels, suggesting that they were able to cause a shift toward Th1 profile [242]. These results have also been confirmed in vivo, where stimulation of Th1 cytokines and inhibition of the Th2 pattern reduced tumor progression to a greater extent than naive MSCs [242].
From all these studies, it is evident that MSCs engineered with immunostimulating cytokines are more powerful tools for cancer therapy than cytokines alone. All results underlined the ability of MSCs to migrate into tumor sites and stimulate release of proinflammatory cytokines, which, in turn, prompted a potent cytotoxic effect against cancer cells without affecting healthy tissues. It is worth emphasizing that the route of administration of engineered MSCs seemed more important than the type of cytokine transduced to obtain the best result; indeed, intratumoral injection increased their efficacy, while little difference was observed with different cytokines. In addition, MSCs transduced with combined cytokines greatly enhanced the results, most likely due to the synergistic effect of cytokines.
An interesting question arising from all these studies concerns the ability of immunoenhanced MSCs to keep releasing immunosuppressive factors, such as TGF-β, IDO, PEG-2, and many others. Could transduction modify the immunosuppressive profile of MSCs? Since it has been shown that transduction of MSCs with viral vectors did not affect MSC properties, we can speculate that their ability should not be modified. Nevertheless, the release of proinflammatory cytokines from MSCs could inhibit the production of anti-inflammatory ones in an autocrine manner, and the balance between the pro/anti-inflammatory profile of MSCs could be tipped in favor of the proinflammatory one, also in the presence of proinflammatory stimuli. This point was not investigated in the works examined, and the question is open and deserves further insight.
Stability and duration of proinflammatory cytokine expression depend on the method of transduction used. As said above, LV vectors guarantee long-lasting production compared to Adv and AAV vectors because they integrate their genome into the host's. Unfortunately, most of the work discussed in the present review does not address such an issue expressly.
Future Outlook
One of the main goals of regenerative and personalized medicine is the development of cellular therapies free of side effects and devoid of ethical concerns. MSCs were found to be suitable and promising candidates, as demonstrated by the remarkable results obtained in numerous clinical trials. Although the vectors alone are being used in various clinical settings and preclinical studies showed that MSC transduction is safe and can enhance therapeutic potential compared to naive MSCs, no clinical trials have yet been performed with engineered MSCs.
Large-scale application of engineered MSCs for cell therapy requires a high number of cells readily available; this obstacle could be overcome by implementation of small-scale bioreactors that have proved adequate to obtain large numbers of clinical-grade MSCs [243]. Another new promising source of MSCs is inducible pluripotent stem cells that can be reprogrammed into MSCs, making it possible to derive autologous MSCs from uninvasive biopsies (eg, skin biopsy) [244]. This derivation also solves troubles with the aged patient, who carries BM-MSCs with significantly reduced survival and differentiation potential; in contrast, using human iPSCs can generate indefinitely fresh MSCs. The only caveat seems their reduced adipogenic differentiation [245]. In addition, small-scale bioreactors have also proved effective at expanding pluripotent stem cells [246], making clinical trials with inducible pluripotent stem cell-derived MSCs accessible to small laboratories.
Viral vector manufacturing is quickly moving from plates to bioreactors, with advantages in terms of costs, standardization, and biosafety [247]. This shift will facilitate genetic engineering to move from bench to bedside. New generation, safer viral vectors have come of age as therapeutic tools, with the European Medicines Agency approving the first gene therapy using an AAV vector to treat lipoprotein lipase deficiency (Glybera®) in 2012. Furthermore, recent data from three clinical trials of gene therapy for hemophilia B are very encouraging [248]. These examples and many more undergoing gene therapy trials are increasing expectations of gene therapy at bedside.
However, with the first gene therapy now licensed and priced at around US$1 million per patient, cost and uncertain funding mechanisms present a potential barrier to patient access.
A combination of advances in gene transfer and manipulation (eg, gene editing) with advances in pluripotent stem cell reprogramming (eg, nonintegrative reprogramming and bench bioreactors), large-scale production, and cost/reimbursement optimization will likely increase access to MSC therapeutics in the coming decade [249 –253].
Footnotes
Acknowledgments
P.Q. is supported by Fondazione ARPA (
Author Disclosure Statement
The authors have no commercial, proprietary, or financial interest in the products or companies described in this article.