
Abstract
microRNAs are key regulators of gene expression that control stem cell fate by posttranscriptional downregulation of hundreds of target genes through seed pairing in their 3′ untranslated region. In fact, miRNAs tightly regulate fundamental stem cell processes, like self-renewal, proliferation, and differentiation; therefore, miRNA deregulation may contribute to the development of solid tumors and hematological malignancies. miR-382-5p has been found to be upregulated in patients with myeloid neoplasms, but its role in normal hematopoiesis is still unknown. In this study, we demonstrated that miR-382-5p overexpression in CD34+ hematopoietic stem/progenitor cells (HSPCs) leads to a significant decrease of megakaryocyte precursors coupled to increase of granulocyte ones. Furthermore, by means of a computational analysis using different prediction algorithms, we identified several putative mRNA targets of miR-382-5p that are downregulated upon miRNA overexpression (ie, FLI1, GATA2, MAF, MXD1, RUNX1, and SGK1). Among these, we validated MXD1 as real target of miR-382-5p by luciferase reporter assay. Finally, we showed that MXD1 knockdown mimics the effects of miR-382-5p overexpression on granulocyte and megakaryocyte differentiation of CD34+ cells. Overall, our results demonstrated that miR-382-5p expression favors the expansion of granulocyte lineage and impairs megakaryocyte commitment through MXD1 downregulation. Therefore, our data showed for the first time that the miR-382-5p/MXD1 axis plays a critical role in myelopoiesis by affecting the lineage choice of CD34+ HSPCs.
Introduction
M
In the last decade, growing evidence has unraveled the role of miRNAs as key regulators of stem cell fate by modulating essential processes like self-renewal, proliferation, and differentiation. In particular, the combinatorial expression of miRNAs can precisely characterize specific cell types and regulate different phases of differentiation [3,4]. Moreover, recent advancements in computational and molecular biology have highlighted the existence of regulatory networks where miRNAs modulate the expression of transcription factors and transcription factors regulate miRNA levels to promote or maintain some important cell functions [1]. On the other side, miRNA expression deregulation may contribute to the development of different human diseases, including cancer [5].
In this study, we focused our attention on miR-382-5p, which resulted to be deregulated in several solid tumors and hematologic neoplasms [6 –9]. In particular, miR-382-5p upregulation has been reported in Acute Promyelocytic Myeloid Leukemia (AML-M3) [10,11] and in Primary Myelofibrosis (PMF) patients [12]. Although an abnormal expression level of miR-382-5p has been described in these hematological malignancies, its role in normal hematopoiesis has not been described so far. Therefore, to characterize the function of miR-382-5p in myeloid development, we evaluated the effects of its overexpression on the commitment of human CD34+ hematopoietic stem/progenitor cells (HSPCs). Interestingly, we identified MAX dimerization protein 1 (MXD1) as a target of miR-382-5p and demonstrated that miR-382-5p favors granulocyte lineage at the expense of megakaryocyte one through MXD1 downregulation.
Materials and Methods
Human CD34+ cell purification
Umbilical cord blood (CB) samples were collected after normal deliveries, according to the institutional guidelines for discarded material (Clearance of Ethics Committee for Human experimentation of Modena: Secretary office Saverio Santachiara,
Electroporation of CD34+ cells
Human CD34+ cells were transfected using the 4D-Nucleofector System (Lonza Group Ltd.) as previously reported (12). Briefly, after 24 h of incubation in the cell-culture condition described above, CD34+ cells were nucleofected twice, once every 24 h, with mirVana miR-382-5p mimic or mirVana miRNA mimic Negative Control #1 (m-NegCTR) (Life Technologies). For each electroporation, 4 × 105 CD34+ cells were resuspended in 100 μL of P3 Primary Cell Solution (Lonza) containing 3 μg of mimic miRNA and pulsed with the program DS112. After each transfection, CD34+ cells were transferred into prewarmed fresh medium in 24-well plates (EuroClone) and maintained in the same culture conditions as described above.
For small interfering RNA (siRNA) transfection, each sample was electroporated thrice, once every 24 h, with 3 μg of a mix of three Silencer Select siRNA targeting human MXD1 (Supplementary Table S1; Supplementary Data are available online at
Cells were analyzed 24 and 48 h after the last nucleofection for both cell viability and miR-382-5p and/or MXD1 expression.
Viability measurement was assessed by trypan blue exclusion assay 24 h after the last nucleofection [14]. In a Neubauer chamber, at least 100 cells were microscopically analyzed in duplicate for viability. The mean percentage of living cells of the two analyses was calculated.
CD34+ cell-culture conditions and morphological and immunophenotypic analysis
CD34+ cells were plated (5 × 105/mL) in IMDM with the addition of 20% BIT 9500 serum substitute (bovine serum albumin, insulin, and transferrin; STEMCELL Technologies) 24 h after the last nucleofection, to set up a multilineage cell culture (SCF, 50 ng/mL; FLT3L, 50 ng/mL; TPO, 20 ng/mL; IL3, 10 ng/mL; IL6, 10 ng/mL; all cytokines from Miltenyi), megakaryocyte (MK) unilineage culture (TPO, 100 ng/mL), and granulocyte (GN) unilineage culture (SCF, 10 ng/mL; GCSF, 25 ng/mL) [13].
Differentiation of CD34+ cells was monitored by morphological analysis of May-Grunwald-Giemsa stained cytospins and by flow cytometry analysis of CD34, CD14, CD66b, CD15, Glycophorin A (GPA), CD41, and CD42b surface antigen expression at days 5, 7, 10, and 12 after the last nucleofection. Images were captured using an Ax10scopeA1 microscope equipped with AxioCam ERc 5S Digital Camera and Axion software 4.8 (all Carl Zeiss MicroImaging, Inc.). The images were then processed with Adobe Photoshop 11.0.2 software.
The following monoclonal antibodies (MoAbs) were used for flow cytometry analysis: phycoerythrin (PE)-conjugated mouse anti-human CD14 MoAb, fluorescein isothiocyanate (FITC)-conjugated mouse anti-human CD34 MoAb, FITC-conjugated mouse anti-human CD66b MoAb, FITC-conjugated mouse anti-human CD15 MoAb (all from Miltenyi Biotech), FITC-conjugated mouse anti- human CD41 MoAb, PE-conjugated mouse anti-human CD42b MoAb, and PE-conjugated mouse anti-human GPA MoAb (all from Dako). After staining, cells were analyzed using a BD FACSCanto II (BD Biosciences). At least 10,000 events were counted for each sample to ensure statistical relevance.
miR-382-5p expression during erythroid, megakaryocyte, granulocyte, and mono-macrophage differentiation
After immunomagnetic separation, CD34+ cells were seeded in 24-well plate at a density of 5 × 105/mL in IMDM (EuroClone SPA) containing 20% human serum (BioWhittaker), SCF 50 ng/mL, FLT3L 50 ng/mL, TPO 20 ng/mL, IL6 10 ng/mL, and IL3 10 ng/mL (all from Miltenyi Biotec). After a first phase of expansion, at 24 h of culture (day 0), cells were seeded (5 × 105/mL) in IMDM added with 20% BIT (STEMCELL Technologies) to set up erythrocyte (EPO 0.4 U/mL, SCF 10 ng/mL), megakaryocyte (TPO 100 ng/mL), granulocyte (GCSF 25 ng/mL, SCF 10 ng/mL), and monocyte (MCSF 100 ng/mL, SCF 20 ng/mL, IL6 20 ng/mL, and FLT3L 50 ng/mL) unilineage cultures [13]. CD34+ cell differentiation was monitored by morphological analysis of MGG-stained cytospins and by flow-cytometric analysis of differentiation marker expression. The medium was replaced every 2 days. miR-382-5p expression level was detected by quantitative reverse transcription polymerase chain reaction (qRT-PCR) at different time points (ie, days 1, 2, 3, 5, 7, 9, and 12) after seeding cells in erythrocyte, megakaryocyte, granulocyte, or mono-macrophage unilineage cultures. The day 0 sample was designated as the calibrator.
Methylcellulose and collagen clonogenic assays
Two different clonogenic assays were performed at 24 h after the last nucleofection. The methylcellulose assay was carried out by plating CD34+ cells in MethoCult™ H4434 Classic (STEMCELL Technologies, Inc.), as previously described [15]. In addition, MK colony-forming units (CFUs-MK) were assayed in collagen-based medium, using a commercial MK Assay Detection Kit (MegaCult-C; STEMCELL Technologies, Inc.) as previously reported [16].
RNA purification and qRT-PCR
Total cellular RNA, including small RNAs, was isolated from CD34+ cells using the miRNeasy Mini RNA Isolation Kit (QIAGEN) following the manufacturer's recommendations. The purity and integrity of RNA samples were determined using disposable RNA chips (Agilent RNA 6000 Nano LabChip Kit) and the Agilent 2100 Bioanalyzer (Agilent Technologies). NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies) was used to evaluate the RNA sample concentration, while 260/280 nm and 260/230 nm ratios were used to assess the purity of RNA.
cDNA was reverse transcribed from total RNA (100 ng per sample) using the High-Capacity cDNA Reverse Transcription Kit (Life Technologies), and real-time PCR was carried out in triplicate with TaqMan gene expression assays and Fast advanced master mix (all reagents from Life Technologies) using an AB 7900HT Fast Real-Time PCR System (Applied Biosystems) [17]. Gene expression relative quantification (RQ) was achieved using the comparative cycle threshold (CT) method using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as the housekeeping gene. The ss-NegCTR sample was considered as the calibrator, the RQ value was expressed as 2−ΔΔCT.
For miRNA cDNA synthesis, 5 ng of total RNA was reverse transcribed using the TaqMan microRNA Reverse Transcription Kit and specific primers included in TaqMan MicroRNA assays. qRT-PCR was performed using TaqMan MicroRNA assays (Life Technologies) as described above and U6 snRNA as a housekeeping control.
Luciferase reporter assays
Predicted target sites of miR-382-5p were cloned into the Pme1 and XbaI sites of pmirGLO Dual-Luciferase miRNA Target Expression Vector (Promega), following manufacturer's instructions. Every cloned plasmid contains the firefly luciferase gene upstream of a given miRNA binding site and the Renilla luciferase gene acting as a normalizer gene. Annealing oligonucleotides containing wild-type miRNA binding sites were designed based on TargetScan 7.0 sequences (Supplementary Table S2). Mutated miRNA binding sites were obtained from the wild-type miRNA binding sites by changing nucleotides 3–5 in the miRNA seed regions.
K562 cells were electroporated by means of the Amaxa 4D-Nucleofector™ System, according to the manufacturer's instructions. Briefly, K562 cells were cultured at a density of 3 × 105 cells/mL 2 days before nucleofection. Cells were then transiently conucleofected with either a miR-382-5p or m-NegCTR at a concentration of 3.6 μM and with either empty vector or miRNA binding site-containing construct at a concentration of 200 ng/sample. For each electroporation, 1 × 106 cells were resuspended in 100 μL of SF Cell line Solution (Lonza) and pulsed with the program FF120. Firefly and Renilla luciferase activities were measured 24 h after electroporation using the Dual-Luciferase Reporter Assay System (Promega), and luminescence was recorded on a GloMax®-Multi+ Detection System with Instinct™ Software (Promega), according to the manufacturer's protocol.
To analyze the data, three levels of normalization were performed as previously described: first, firefly luciferase activity was normalized to that of Renilla luciferase as the internal control for nucleofection efficiency; second, the data were normalized to the effect of the miRNA mimic on the empty vector control reporter; and finally, the luminescence signals were normalized to the effect of the m-NegCTR on the corresponding miRNA binding site-containing construct [12].
Western blot
MXD1 level in both miR-382-5p-overexpressing cells and MXD1-siRNA transfected CD34+ cells was detected by western blot analysis. Briefly, cells were harvested, washed twice with ice-cold PBS, and lyzed (5 × 105 cells/50 μL of lysis buffer) in 50 mM Tris (tris(hydroxymethyl)aminomethane)-Cl (pH 7.4), 150 mM NaCl, 10 mM KCl, 1 mM EDTA, 20 mM NaF, 0.25% Na deoxycholate, 5 mM dithiothreitol (DTT), 1% Nonidet P-40, and protease inhibitors (Complete, catalog #1697498; Roche). Total cell lysates (30 μg for each sample) were loaded onto 10% SDS-polyacrylamide gel and blotted on nitrocellulose membrane. The membranes were stained with Red Ponceau to control loading and transfer. Membranes were then preblocked in blocking solution, composed by 5% bovine serum albumin (BSA) in 0.1% TBST, for 1 h at room temperature (RT), and incubated with primary antibodies: (1) rabbit polyclonal anti-MXD1 antibody (1:500 dilution, anti-Mad1 (C19), catalog #sc-222; Santa Cruz Biotechnology, Inc.) at 4°C overnight; and (2) rabbit polyclonal anti-actin antibody (1:2,000 dilution; Thermo Fisher Scientific, Inc., catalog #PA1-16889) for 1 h at RT. After three washes with TBST, blots were incubated with HRP conjugated goat anti-rabbit secondary antibody (1:10,000 dilution, catalog #sc-2004; Santa Cruz Biotechnology, Inc.) for 1 h at RT and revealed by BM Chemiluminescence Blotting Substrate (POD) (Roche).
Statistical analysis
The statistics used for data analysis in overexpression/silencing experiments and luciferase reporter assays was based on two-tailed Student t-tests for averages comparison in paired samples. Data were analyzed using Microsoft Excel (Microsoft Office, 2008 release) and are reported as mean ± standard error of the mean (SEM). P < 0.05 was considered significant.
Results
miR-382-5p overexpression in CD34+ cells
To study the role of miR-382-5p in HSPC commitment, miR-382-5p was overexpressed in CD34+ cells. By means of qRT-PCR, we confirmed that the miRNA expression level was upregulated upon miR-382-5p mimic transfection at 24 and 48 h (RQ ± SEM, 1,677 ± 383, P < 0.05 and 625 ± 85 P < 0.01, respectively) after the last nucleofection (Fig. 1A). Trypan blue exclusion assay showed that cell viability was about 90% in both m-NegCTR and miR-382-5p samples (Supplementary Fig. S1A).
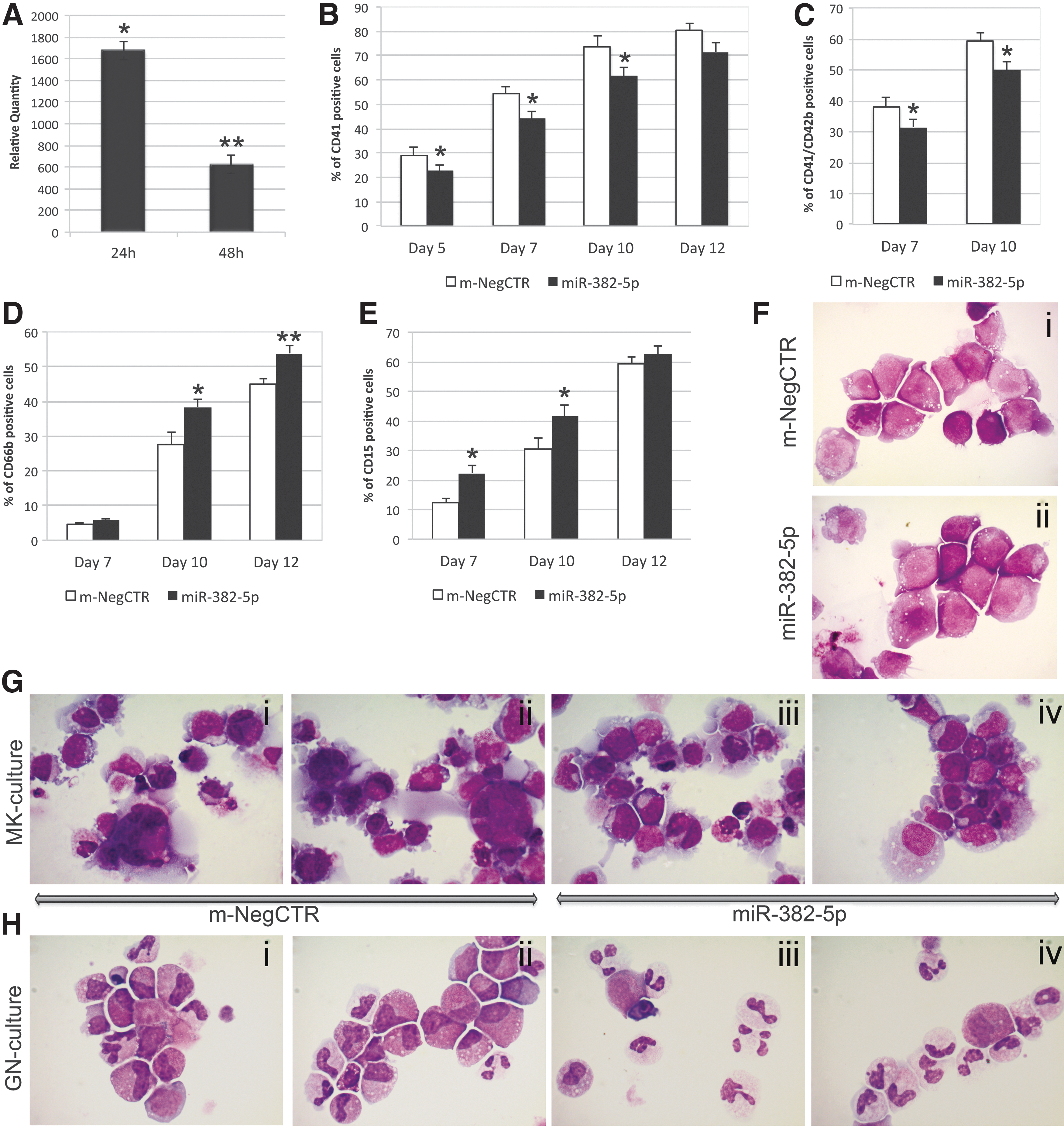
Effect of miR-382-5p overexpression on CD34+ HSPC differentiation.
Flow cytometry analysis of MK differentiation markers performed on TPO unilineage culture at days 5, 7, 10, and 12 showed that miR-382-5p overexpression induces a significant decrease in the CD41+ cell fraction compared with the m-NegCTR (Fig. 1B). In addition, the mature megakaryocytic CD41+/CD42b+ population was similarly reduced at days 7 and 10 (Fig. 1C). On the contrary, miR-382-5p overexpression in GCSF unilineage culture led to a significant increase in the percentage of CD66b+ and CD15+ cells at day 10 after the last nucleofection (Fig. 1D, E). According to flow cytometry results, the morphologic analysis of MGG-stained cytospins obtained from TPO unilineage culture showed a remarkable reduction in MK precursors at different stages of maturation in miR-382-5p-overexpressing cells compared with control (Fig. 1G i–iv). Moreover, morphological evaluation of GCSF-treated cells clearly displayed a considerable enrichment in granulocytes in miR-382-5p sample (Fig. 1H i–iv). Similarly, miR-382-5p upregulation in multilineage culture conditions further confirmed these results (Fig. 2). Flow cytometry analysis of mono-macrophage (CD14) and erythrocyte (GPA) markers did not highlight any significant modulation of expression between miR-382-5p CD34+ cells and the m-NegCTR sample (Supplementary Fig. S1B–D).
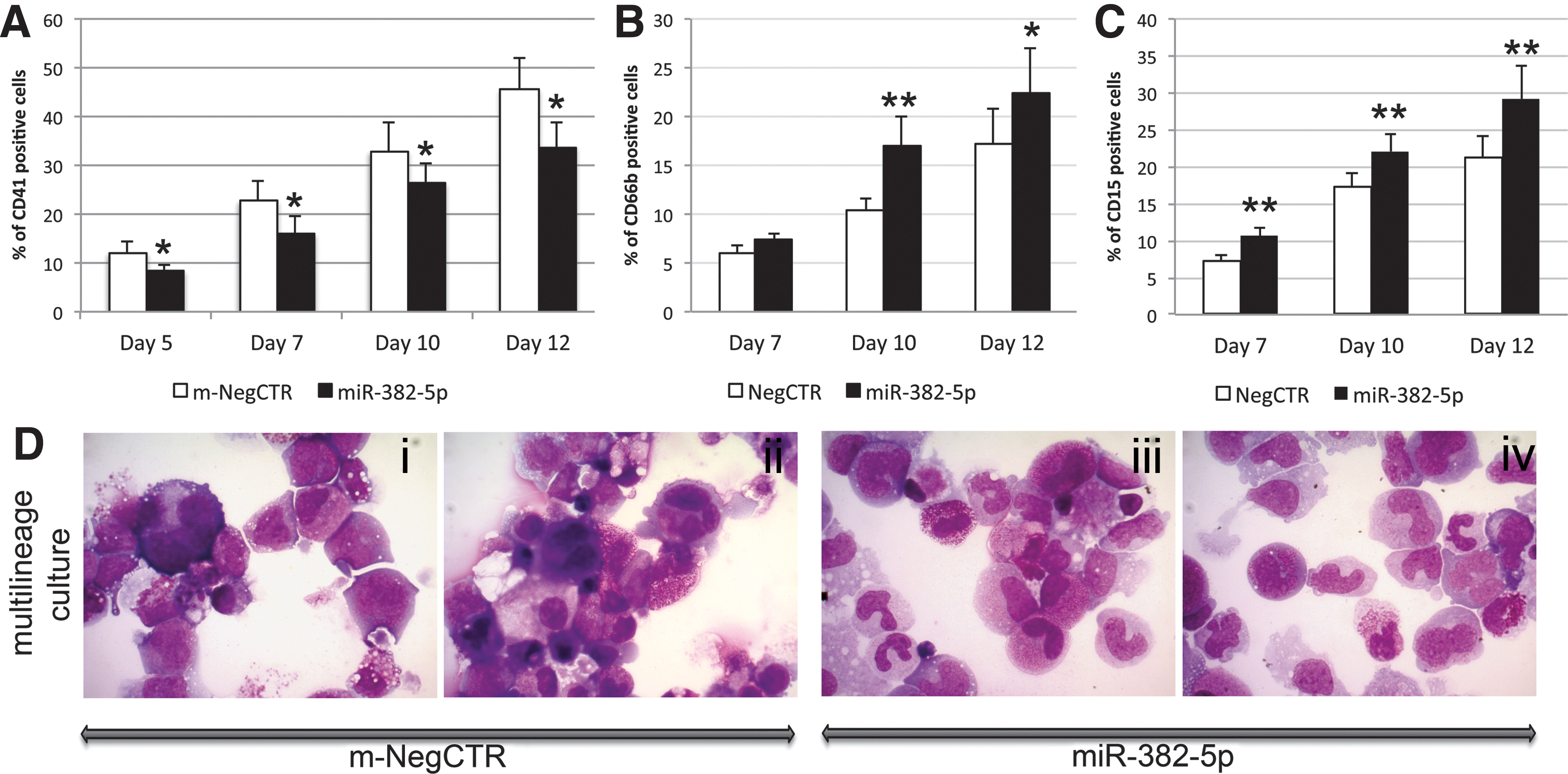
Effect of miR-382-5p overexpression on CD34+ HSPC differentiation in multilineage culture.
Consistently with previous results, methylcellulose assay highlighted a remarkable increase in the percentage of granulocyte colony-forming unit (CFU-G), whereas monocyte (CFU-M), erythroid (Burst-Forming Units [BFU-E] and CFU-E), and granulo-monocyte (CFU-GM) colonies were not significantly affected (Fig. 3A). Furthermore, the collagen-based assay that supports the growth of MK progenitors in vitro showed that miR-382-5p overexpression causes a significant decrease in the CFU-MK percentage coupled with a significant increase of non-MK colonies (Fig. 3B).
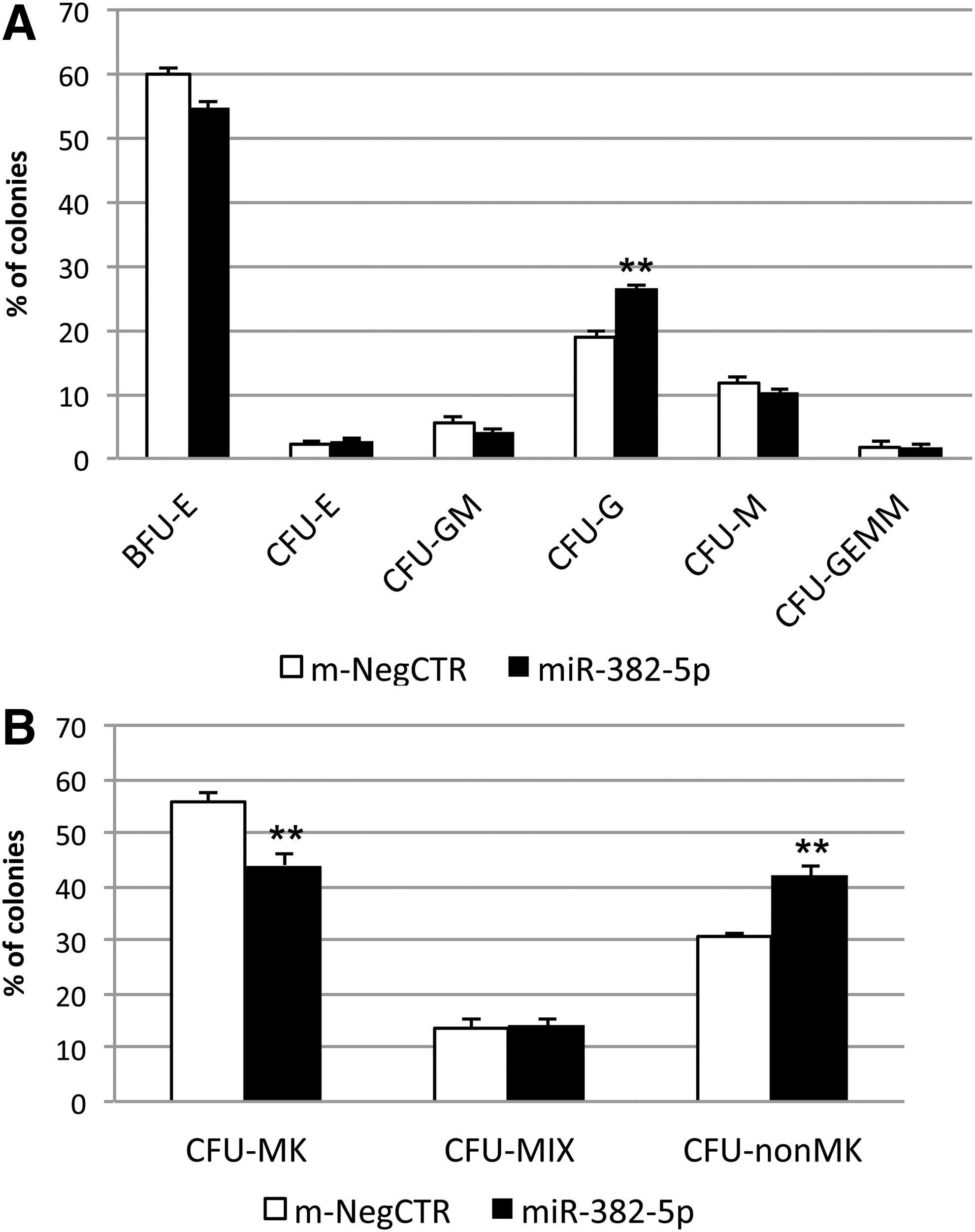
Methylcellulose and collagen-based clonogenic assays upon miR-382-5p overexpression in CD34+ HSPCs.
Since miR-382-5p interferes with lineage choice of CD34+ cells, we assessed its expression level in CD34+ and differentiated cells obtained from granulocyte, monocyte-macrophage, erythrocyte, and megakaryocyte unilineage culture conditions. As shown in Supplementary Fig. S2, miR-382-5p expression level was decreased during the initial phase of megakaryocyte and erythroid unilineage cultures, while its expression increased in the last phases of differentiation. On the contrary, miR-382-5p results upregulated during the granulocyte and monocyte-macrophage differentiation (Supplementary Fig. S2).
Overall, these results indicated that miR-382-5p overexpression in CD34+ HSPCs enhances granulocyte differentiation, while negatively interferes with the megakaryocyte one.
miR-382-5p target prediction and validation by luciferase reporter assays
To identify the molecular targets through which miR-382-5p modulates the MK and GN commitment in CD34+ HSPCs, several prediction algorithms (ie, TargetScan,
First, we assessed whether miR-382-5p overexpression in CD34+ HSPCs led to the downregulation of putative target expression by means of qRT-PCR. As shown in Fig. 4A, all mRNAs were downregulated in miR-382-5p-overexpressing cells at 24 h after the last nucleofection.

Identification of miR-382-5p targets.
Finally, to confirm whether the downregulated mRNAs are real targets of miR-382-5p, we validated every putative miRNA-mRNA interaction by means of luciferase reporter assay. To this end, the predicted miRNA binding sites were cloned into the 3′ UTR of the Dual Luciferase pmirGLO vector and transiently transfected in K562 cells. Luciferase activity was measured in the presence of miR-382-5p mimic or m-NegCTR and normalized using Renilla activity. As shown in Fig. 4B, miR-382-5p overexpression significantly inhibited the luciferase activity in the sample transfected with the wild-type miRNA binding site on MXD1 3′ UTR; conversely, the luciferase activity was not repressed by miR-382-5p overexpression in sample transduced with MXD1-mutated miRNA binding site. Likewise, miR-382-5p upregulation did not significantly reduce the luciferase activity in samples transduced with miRNA binding site of GATA2, FLI1, RUNX1, MAF, and SGK1 (Fig. 4B). According to luciferase reporter assay results, western blot analysis showed a marked reduction of MXD1 level upon miR-382-5p overexpression (Fig. 4C).
MXD1 silencing in CD34+ cells
To better characterize the role of miR-382-5p/MXD1 axis in myelopoiesis, we investigated whether MXD1 silencing could reproduce the effects mediated by miR-382-5p overexpression on GN and MK differentiation. Therefore, CD34+ cells were transfected with a mixture of three Silencer Select siRNAs targeting MXD1 mRNA (Supplementary Table S1) and with a nontargeting siRNA as a negative control (ss-NegCTR). As shown in Supplementary Fig. S3A, cell viability was >90% in all samples, without any significant differences between ss-NegCTR and MXD1-siRNA. The expression level of MXD1 in control samples and MXD1-siRNA cells was assessed by qRT-PCR and western blot analysis. As shown in Fig. 5A, we obtained a significant downregulation of MXD1 mRNA at 24 and 48 h after the last nucleofection (RQ ± SEM, 0.34 ± 0.03, P < 0.001 and 0.48 ± 0.10, P < 0.01, respectively). Accordingly, the MXD1 level is remarkably decreased at 24 h postnucleofection (Fig. 5B).
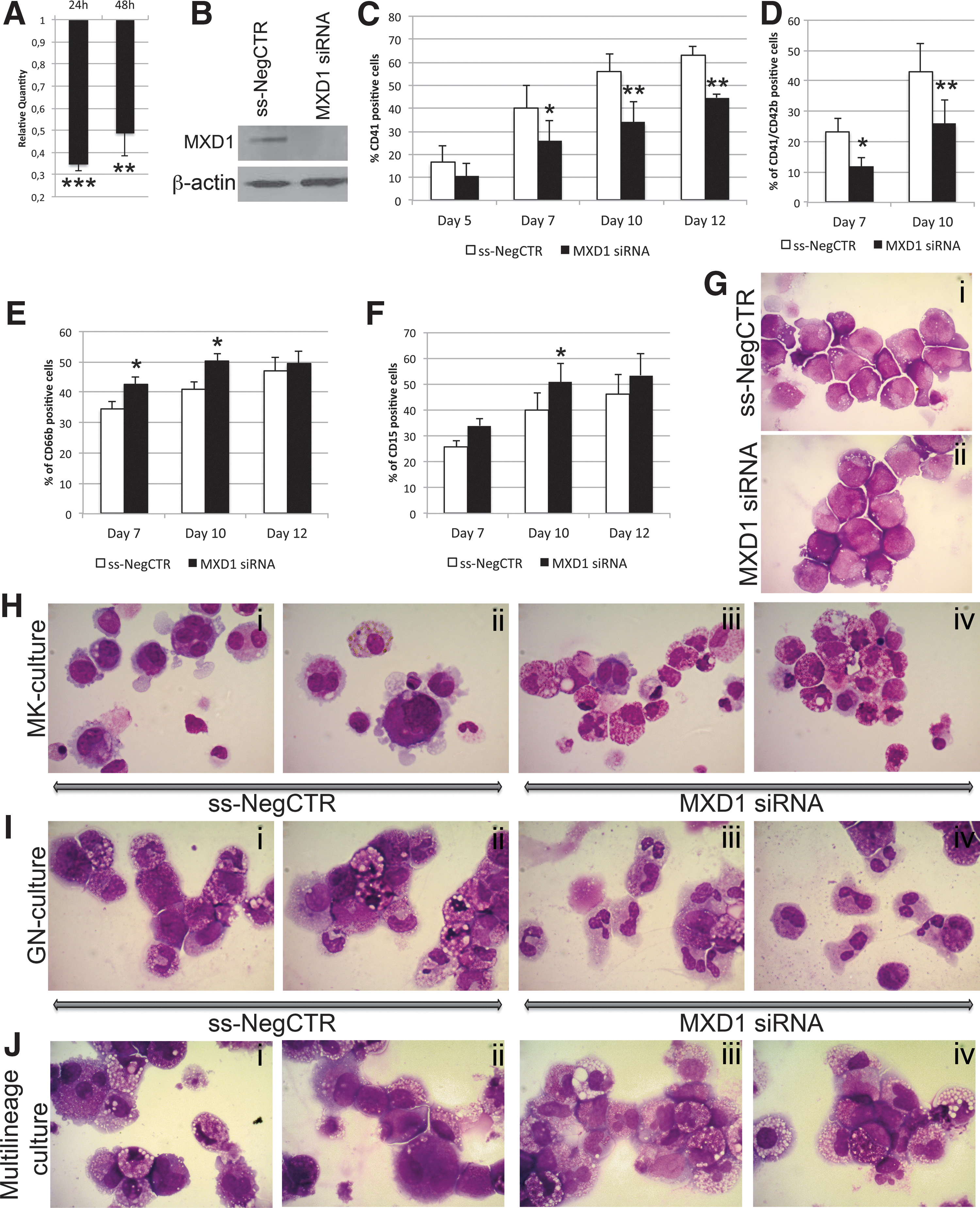
Effect of MXD1 silencing on CD34+ HSPC differentiation.
As observed for the miR-382-5p overexpression, MXD1 silencing led to a substantial decrease in the percentage of CD41+ and CD41+/CD42b+ cells at days 7 and 10 after the last nucleofection in MK unilineage culture (Fig. 5C, D). At the same time, MXD1 silencing induces a significant increase in the percentage of CD66b+ and CD15+ cells compared with the ss-NegCTR sample (Fig. 5E, F). Furthermore, the morphological analysis of MGG-stained cytospins showed a significant reduction in MK precursors (Fig. 5H i–iv) and a significant expansion of granulocytes in MXD1-siRNA cells (Fig. 5I i–iv). The same morphological differences could be appreciated also in MGG-stained cytospins obtained from multilineage culture (Fig. 5J i–iv).
Conversely, we did not observe any modulation of GPA and CD14 expression upon MXD1 knockdown (Supplementary Fig. S3B–D).
Finally, according to results obtained by means of clonogenic assay on miR-382-5p-overexpressing cells, we found a significant increase in the percentage of CFU-G, while CFU-M, BFU-E/CFU-E, and CFU-GM colonies were not significantly affected by MXD1 downregulation (Fig. 6A). In addition, the collagen-based assay confirmed that MXD1 silencing induces a decrease of MK progenitors as showed by a lower CFU-MK percentage in MXD1-siRNA sample compared with control (Fig. 6B).
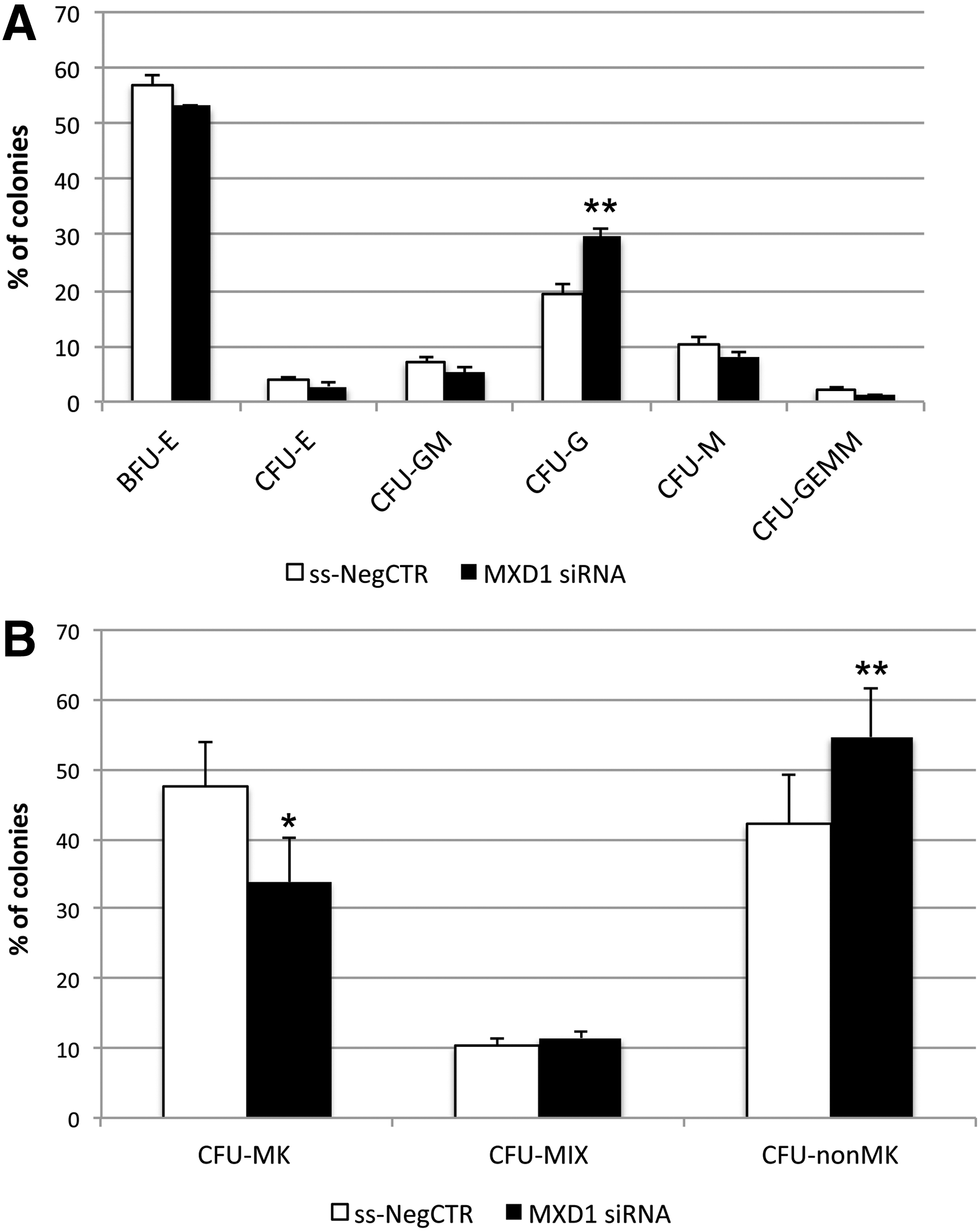
Methylcellulose and collagen-based clonogenic assays upon MXD1 silencing in CD34+ HSPCs.
Since the expression levels of the megakaryocyte transcription factors GATA2, FLI1, RUNX1, and MAF and of the kinase SGK1 were decreased upon miR-382-5p overexpression, although they have not been previously identified as functional targets, we still wondered whether MXD1 silencing could modulate their expression. By means of qRT-PCR, we showed that the mRNA levels of GATA2, RUNX1, and FLI1 were significantly downregulated in MXD1-silenced cells (Fig. 7). Conversely, MAF and SGK1 expression levels seem not to be affected by MXD1 knockdown.
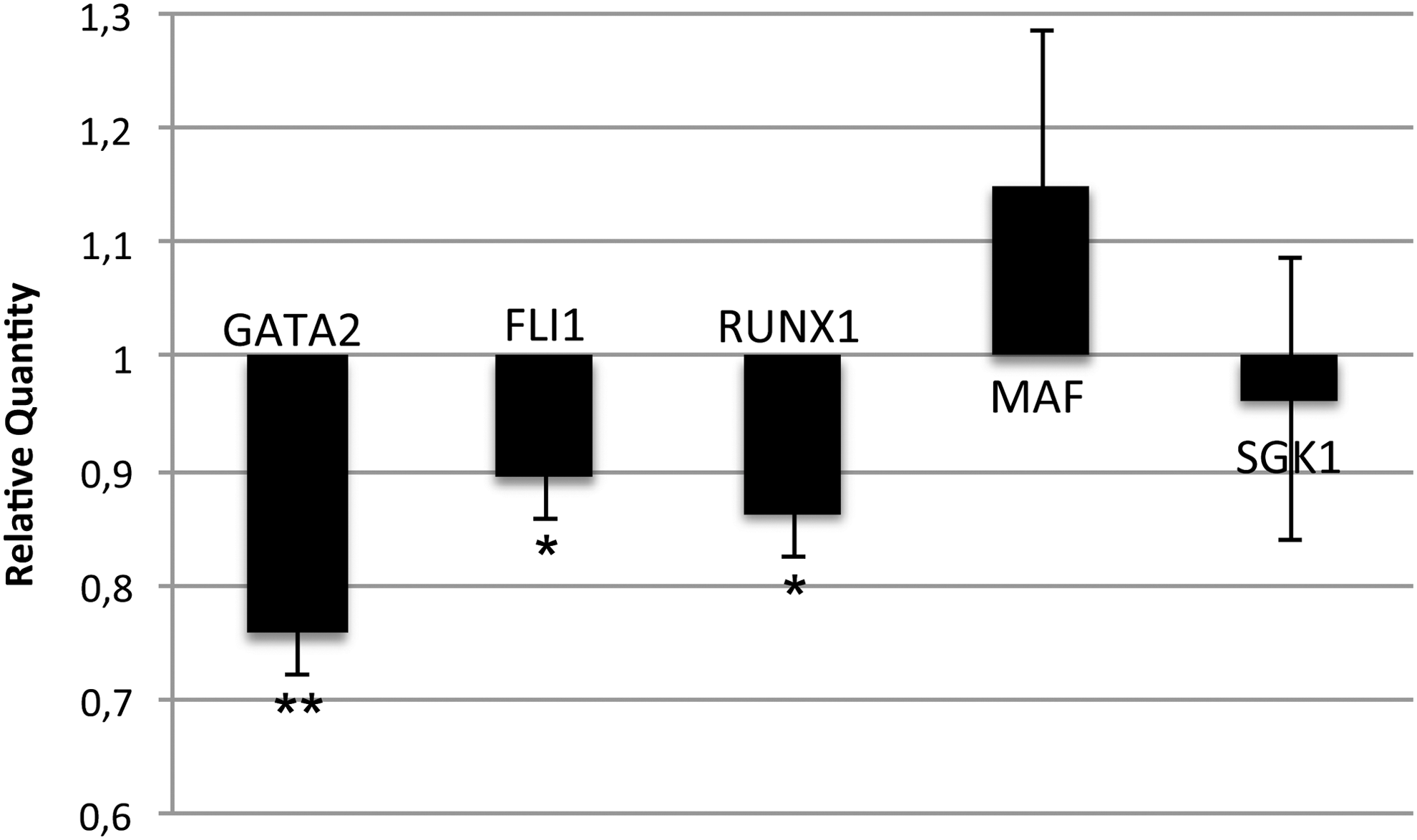
MXD1 downstream regulation of GATA2, FLI1, and RUNX1. The expression levels of GATA2, FLI1, RUNX1, MAF, and SGK1 were measured by qRT-PCR in MXD1 siRNA-transfected CD34+ cells at 24 h after the last nucleofection. Data are reported as Relative Quantity. Values are reported as mean ± SEM. The results come from five independent experiments. **P < 0.01 versus ss-NegCTR and *P < 0.05 versus ss-NegCTR.
Discussion
Growing evidence shows that miRNAs regulate stem cell fate decision by modulating fundamental processes like self-renewal, proliferation, and differentiation. For this reason, dysregulation of miRNA expression represents a common mechanism of tumorigenesis and cancer progression.
miR-382-5p overexpression has been described in some hematological malignancies, such as PMF [12] and Acute Myeloid Leukemia (AML) [10,11], even if its role in normal hematopoiesis was still unknown.
In this study, we showed for the first time that miR-382-5p plays a critical role in hematopoiesis by influencing the lineage choice of CD34+ cells. In particular, we demonstrated that miR-382-5p supports the expansion of granulocyte lineage, whereas negatively interferes with megakaryocyte commitment, as demonstrated by a significant increase of granulocyte precursors coupled to decrease of megakaryocyte ones in miR-382-5p-overexpressing cells (Figs. 1 –3).
Next, to identify the molecular targets by which miR-382-5p regulates HSPC fate, we found by a prediction analysis several putative targets known to be involved in MK and GN differentiation (ie, FLI1, GATA2, MAF, MXD1, RUNX1, and SGK1) (Table 1) [18 –22], whose mRNA expression was decreased in miR-382-5p-overexpressing cells (Fig. 4A). Therefore, by means of luciferase reporter assay, we were able to validate the MXD1-miR-382-5p interaction (Fig. 4B). Moreover, as shown in Fig. 4C, MXD1 level is strongly reduced in miR-382-5p-overexpressing cells compared to control.
MXD1, also known as MAD1, is a transcriptional repressor involved in the MYC/MAX/MAD pathway, one of the best known networks that control cell proliferation and differentiation [23]. In fact, MXD1 interacts with MAX antagonizing MYC/MAX dimer activity [23]. Queva et al. showed that MXD1 is rapidly induced upon cell differentiation, when MYC expression is downregulated and cells begin to exit the cell cycle acquiring a differentiated phenotype [24].
In this study, we demonstrated that MXD1 knockdown reproduces the effects of miR-382-5p overexpression on CD34+ HSPC differentiation leading to a significant decrease of megakaryocyte precursors coupled to increase of granulocyte ones (Figs. 5 and 6). According to our results, the increased expression of MXD1 has been previously reported also in megakaryocytes obtained in vitro from human CD34+ cells [21]. Therefore, the MXD1 expression in CD34+ cells seems to be essential for megakaryocyte differentiation; in fact, the MK transcription factors GATA2, FLI1, and RUNX1 were significantly decreased after MXD1 silencing (Fig. 7) [18], suggesting that the expression of these genes could be regulated by MXD1 in a direct or indirect way. For instance, the downregulation of GATA2, FLI1, and RUNX1 might be mediated by the MXD1-driven inhibition of a transcriptional repressor (ie, BCL6) [25]. Moreover, MXD1 can also heterodimerize with other proteins in addition to MAX (ie, MLX), even if the function of these complexes, as well as their targets, has not been defined so far [26,27].
Two independent studies reported that MXD1 induces erythroid differentiation in human [28] and mouse [29] erythroleukemia cells. In this study, we showed that MXD1 silencing did not impair erythroid differentiation in normal CD34+ cells, as demonstrated by the expression level of GPA (Supplementary Fig. S3C, D) and the percentage of red colonies (Fig. 6A), which did not change after MXD1 knockdown. A possible explanation for this different cellular behavior is that normal CD34+ cells might express some proteins that could replace MXD1 during erythroid differentiation.
Furthermore, our data demonstrated that MXD1 plays an important role in granulocyte differentiation; in fact, the MXD1 knockdown enhances the fraction of granulocyte precursors both in liquid and semisolid cultures (Figs. 5 and 6). In agreement with our data, Foley et al. demonstrated that MXD1 knockout in mice affects the granulocyte lineage by reducing cell cycle withdrawal during the late stage of granulocyte differentiation, resulting in increased frequency of granulocytic precursors in the bone marrow [20]. Moreover, another mechanism by which MXD1 inhibition could favor the expansion of granulocyte fraction is mediated by the reduction of MAX/MXD1 heterodimers arising in an increased availability of MAX protein, which in turn activates C/EBPalpha, a master regulator of granulocyte commitment [30].
Overall, these results demonstrated that miR-382-5p favors the expansion of granulocyte precursors through MXD1 downregulation. Noteworthy, two independent research groups reported a high level of miR-382-5p in Acute promyelocytic leukemia (APL) patients with t(15;17) [10,11]. This subtype of AML is characterized by PML-RARalpha fusion oncoprotein that is responsible for the differentiation block of leukemic promyelocytes. Of note, as demonstrated by Khan et al., PML-RARalpha can interact with MXD1 and inhibits its transcriptional repression activity [31]. This evidence suggests that the deregulation of miR-382-5p/MXD1 axis contributes to the development of APL in cooperation with PML-RARalpha fusion protein; therefore, miR-382-5p could be considered as a novel molecular target for APL treatment.
In summary, our results demonstrated that miR-382-5p supports the expansion of granulocyte lineage, whereas negatively interferes with megakaryocyte one through MXD1 downregulation. To our knowledge, this is the first study describing miR-382-5p/MXD1 axis as a crucial regulator of hematopoietic stem cell fate.
Footnotes
Acknowledgments
This work was supported by Associazione Italiana per la Ricerca sul Cancro (AIRC), project number #10005 “Special Program Molecular Clinical Oncology 5 × 1,000” to AGIMM (AIRC-Gruppo Italiano Malattie Mieloproliferative,
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.