
Abstract
Adipose-derived stem cells (ADSCs) are multipotent cells that have attracted much recent attention and emerged as therapeutic approaches in several medical fields. Although current knowledge of the biological impacts of ADSCs in cancer research is greatly improved, the underlying effects of ADSCs in tumor development remain controversial and cause the safety concerns in clinical utilization. Hence, we isolated primary ADSCs from the abdominal fat of mice and conducted interaction of ADSCs with Lewis lung carcinoma cells in culture and in mice to investigate the impacts of ADSCs on tumor development. Cytokine array and neutralizing antibody were further utilized to identify the key regulator and downstream signaling pathway. In this study, we demonstrated that ADSCs enhance the malignant characteristics of LLC1 cells, including cell growth ability and especially cancer stem cell property. ADSCs were then identified to promote tumor formation and growth in mice. We further determined that ADSC interaction with LLC1 cells stimulates increased secretion of interleukin-6 mainly from ADSCs, which then act in a paracrine manner on LLC1 cells to enhance their malignant characteristics. Interleukin-6 was also identified to regulate genes related to cell proliferation and cancer stem cell, as well as to activate JAK2/STAT3, a predominant interleukin-6-activated pathway, in LLC1 cells. Collectively, we demonstrated that ADSCs play a pro-malignant role in tumor development of Lewis lung carcinoma cells by particularly promoting cancer stem cell property through interleukin-6 paracrine circuit, which is important for safety considerations regarding the clinical application of ADSCs.
Introduction
S
The MSCs within adipose tissue, termed adipose-derived stem cells (ADSCs), are a promising stem cell source to treat various aging-related degenerative diseases. Compared to bone marrow, adipose tissue is easily harvested in large quantities with little donor-site morbidity and yields relatively high amount of stem cells. Hence, the advantages of ADSCs are easy and relatively cheap to obtain and can be generated in large amounts with less donor discomfort [8]. In addition, it was reported that ADSCs grow stably and maintain stemness in vitro and have a higher expansion rate than bone marrow-derived mesenchymal stromal cell (BMSC) [6]. ADSCs have been frequently used in several medical fields such as plastic [9], orthopedic [10], and cardiac surgery [11] or for reconstruction after oncological surgery [12], due to their convenient acquisition and regenerative potential. However, recent literatures exploring the potential role of ADSCs in the development of various cancers raise the safety concerns of clinical application [13].
It is well documented that cancer development is a multistep process that associates with many malignant characteristics, such as uncontrolled cell growth, abnormal angiogenesis, invasion of surrounding normal tissue, and spread through circulatory system. In 2014, Freese et al. systematically searched PubMed articles published between 2001 and 2014 to investigate the relationship between ADSCs and cancer. They summarized that ADSCs exhibit cancer-promoting properties on several cancer types such as endometrial, breast, ovarian, gastric, lung, and pancreatic cancers. Their cancer-promoting effects include promotion of cell proliferation, motility, and invasion and enhancement of tumor growth, angiogenesis, and metastasis [13]. In addition to above properties, ADSCs have been recently explored by us and other studies that they may enhance the properties of cancer stem cell (CSC) in breast and colon cancers [14,15]. However, what are the effects of ADSCs on cancer stemness and/or tumor initiation in lung cancer are still very poorly understood. Therefore, in this study, we conducted interaction of ADSCs with LLC1 cells in vitro and in vivo to identify the impacts of ADSCs on lung tumor development, especially on lung CSCs.
Materials and Methods
Cell lines and ADSC isolation
LLC1 (ATCC CRL-1642) Lewis lung carcinoma cells were infected with FUW-Luc-mCh-puro lentiviral particles and cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS) to stably express mCherry fluorescent protein. ADSCs were isolated as previously described [16]. ADSCs were cultured in αMEM supplemented with 10% FBS.
Animal studies
All animal studies were approved by the Institutional Animal Care and Use Committee of Taipei Medical University. Four- to six-week-old female C57BL/6 and green fluorescent protein (GFP) transgenic mice were purchased from National Laboratory Animal Center (Taipei, Taiwan). The mice were housed under pathogen-free conditions and fed autoclaved food and water. For tumor initiation experiments, 4 × 104 LLC1 cells were implanted with or without fivefolds of ADSCs through subcutaneous injection. For tumor growth experiments, 2 × 105 LLC1 cells were implanted with or without fivefolds of ADSCs through subcutaneous injection.
Flow cytometry
For cell surface marker analysis, cells were harvested, resuspended in 100 μL HBSS containing specific antibodies, and incubated at 4°C for 30 min. Samples were analyzed using FACSCanto II flow cytometer (BD Biosciences) and FCS Express software (De Novo). Antibodies were used at concentrations as recommended by the manufacturer. Antibody information is provided in Supplementary Table S1; Supplementary Data are available online at
Reverse transcription-polymerase chain reaction
Total RNA was extracted from cells using High Pure RNA Isolation Kit (Roche) according to the manufacturer's instructions. Reverse transcription-polymerase chain reaction (RT-PCR) was performed as previously described [17]. Primer sequences are listed in Supplementary Table S2. Relative quantitation was performed using ImageJ software (Version 1.46r).
Multilineage differentiation Assays
To evaluate the in vitro differentiation potential of cells, we conducted differentiation induction experiments of three major mesodermal lineages. The differentiation assays were performed as previously described [14].
Sphere formation assay
4 × 104 ADSCs, 4 × 104 LLC1 cells, or 4 × 104 LLC1 cells plus 2 × 105 ADSCs were incubated for 7 days in 4 mL of modified sphere medium (DMEM/F12 medium) supplemented with 1× B-27 supplement (Thermo Fisher Scientific), 20 ng/mL epidermal growth factor (PeproTech), 10 ng/mL fibroblast growth factor-basic (PeproTech), and 20 ng/mL human leukemia inhibitory factor (Sigma-Aldrich) in T25 flasks. Spheres (>50 μm diameter) in 10 random fields per flask were counted and photographed.
In vitro cell growth assay
1 × 105 LLC1 cells and 5 × 105 ADSCs were cocultured using transwell inserts with a 0.4-μm porous membrane (Corning). After coculture for 3 days, cancer cells were harvested and counted.
Bioluminescence imaging
Bioluminescence imaging (BLI) of animals was performed with an IVIS Imaging System 200 Series (PerkinElmer). For in vivo BLI, anesthetized mice were injected intraperitoneally with 75 mg/kg of
Cytokine array
Cells were cultured in complete media for 3 days. The levels of cytokines, growth factors, and chemokines in the culture media were assessed by RayBio® Mouse Cytokine Antibody Array 2 (RayBiotech) according to the manufacturer's instructions. Chemiluminescent data were collected using the Multifunction Gel Image system (TOP BIO CO.) and quantitated with ImageJ software (Version 1.46r) by measuring the intensities of detected spots.
Western blot analysis
The protein extraction and immunoblotting were performed as previously described [18]. Antibody information is provided in Supplementary Table S1.
Statistical analyses and replicates
The sizes of sample group in all data are at least n = 5, unless otherwise indicated. All data presented are representative of at least three independent experiments that yielded similar results. Statistical analyses were performed using GraphPad Prism 5 (GraphPad Software).
Results
Stromal cells from adipose tissue exhibit MSC characteristics
MSCs are recognized by their expression of specific surface markers and multipotency. Hence, we performed assays to determine whether stromal cells isolated from mice abdominal fat displayed MSC characteristics. Flow cytometry analysis showed that the isolated cells revealed MSC surface marker profiles, including negative for CD34 and CD45 (hematopoietic stem cell markers) and positive for CD90, CD105, and Sca-1 (MSC markers) (Fig. 1A). Reverse transcription-polymerase chain reaction (RT-PCR) and specific staining assays were used to evaluate the multilineage differentiation ability of isolated cells. The isolated cells were cultured under standard induction conditions and tested for in vitro differentiation into three mesenchymal lineages, including osteogenesis, chondrogenesis, and adipogenesis. Gene expression of osteogenic (OPN and RUNX2), chondrogenic (COL2A1 and ACAN), and adipogenic (PPARG and Leptin) markers was upregulated in cells after induction (Fig. 1B, left panel). We also found that lineage-specific stainings, including Alizarin Red S staining for osteogenic matrix, Alcian Blue staining for proteoglycan accumulation, and Oil Red O staining for lipid droplet, showed enhanced signals in cells upon induction (Fig. 1B, right panel). Above results suggest that these cells isolated from mouse abdominal adipose tissue reveal MSC properties and are known as ADSCs.
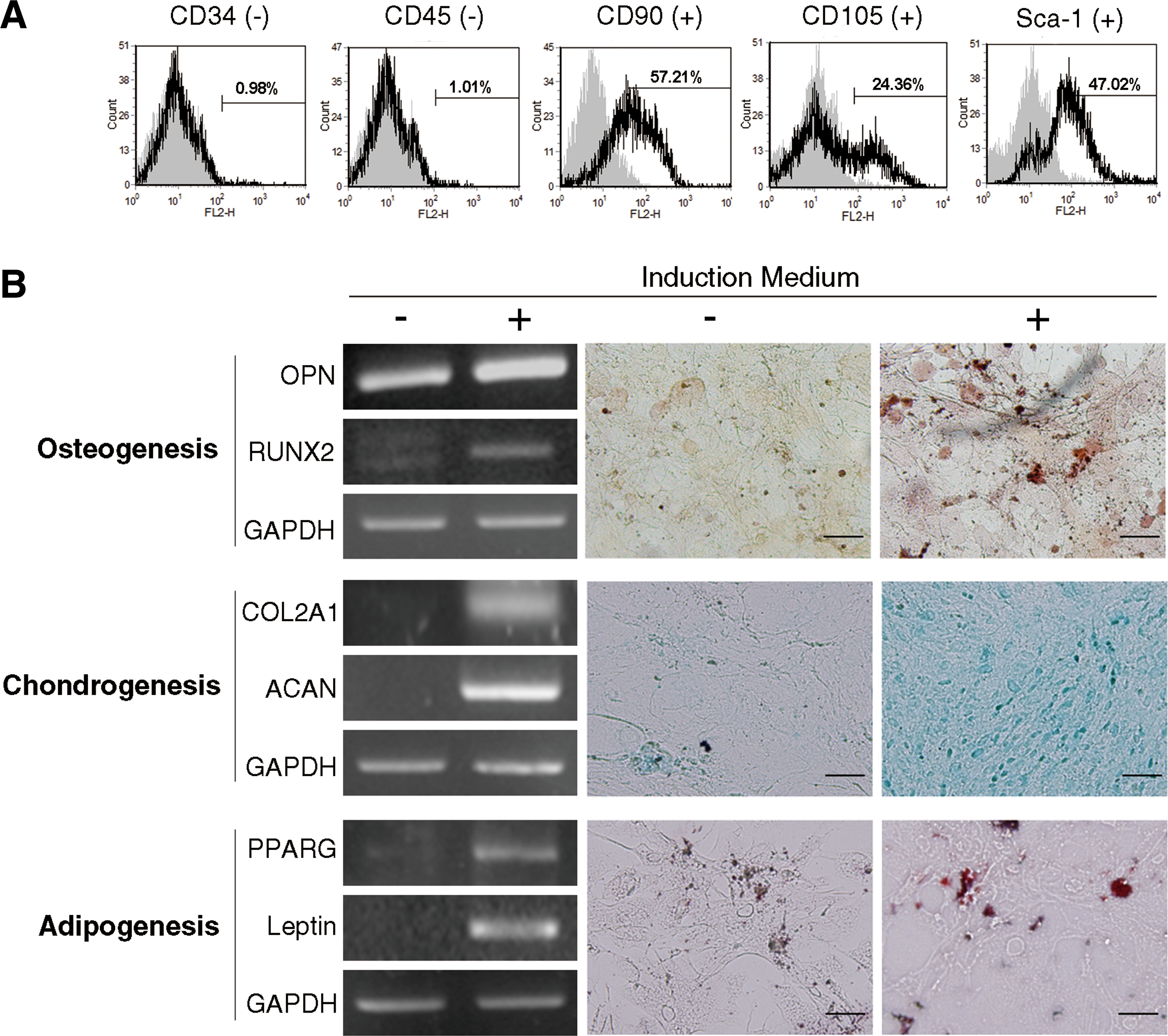
Characterization of adipose-derived stem cells.
ADSCs promote growth ability and CSC property of Lewis lung carcinoma cells
To investigate whether the growth of Lewis lung cancer cells was influenced by ADSCs, we cocultured LLC1 cells with ADSCs using transwell system. The cell growth of LLC1 cells was determined by cell counting. As shown in Fig. 2A, ADSCs could increase the number of LLC1 cells upon coculture. We then utilized RT-PCR to investigate whether ADSCs affected the expression of proliferation-related markers in lung carcinoma cells. The result showed that ADSCs increased the mRNA expression of MKI67 and PCNA in LLC1 cells (Fig. 2B). CSCs are a small population of tumor cells endowed with aberrantly regulated stem-like properties such as self-renewal and differentiation. They are considered to contribute to tumor initiation, heterogeneity, and recurrence. To identify the impacts of ADSCs on lung CSCs, we first conducted sphere formation assay. Tumor sphere formation is widely utilized to retrospectively identify CSCs based on their self-renewal ability at the single-cell level [19]. The results showed that the sphere-forming efficiency of LLC1 cells was significantly increased while cocultured with ADSCs (Fig. 2C). We transduced LLC1 cells with mCherry fluorescent protein and isolated ADSCs from GFP transgenic mice; therefore, fluorescent microscope could be utilized to monitor the interaction of ADSCs and LLC1 cells in sphere formation. Representative images showed that ADSCs survived and integrated into lung cancer spheres (Fig. 2D). RT-PCR was also utilized to evaluate the expression of CSC markers in LLC1 cells cocultured with or without ADSCs. As shown in Fig. 2E, ADSC coculture raised the gene expression of pluripotent (SOX2 and NANOG) and drug-resistant (ALDH1A1 and ABCG2) markers in LLC1 cells. These results suggest that ADSCs could promote the malignant characteristics of Lewis lung carcinoma cells in vitro, including growth ability and especially CSC property.
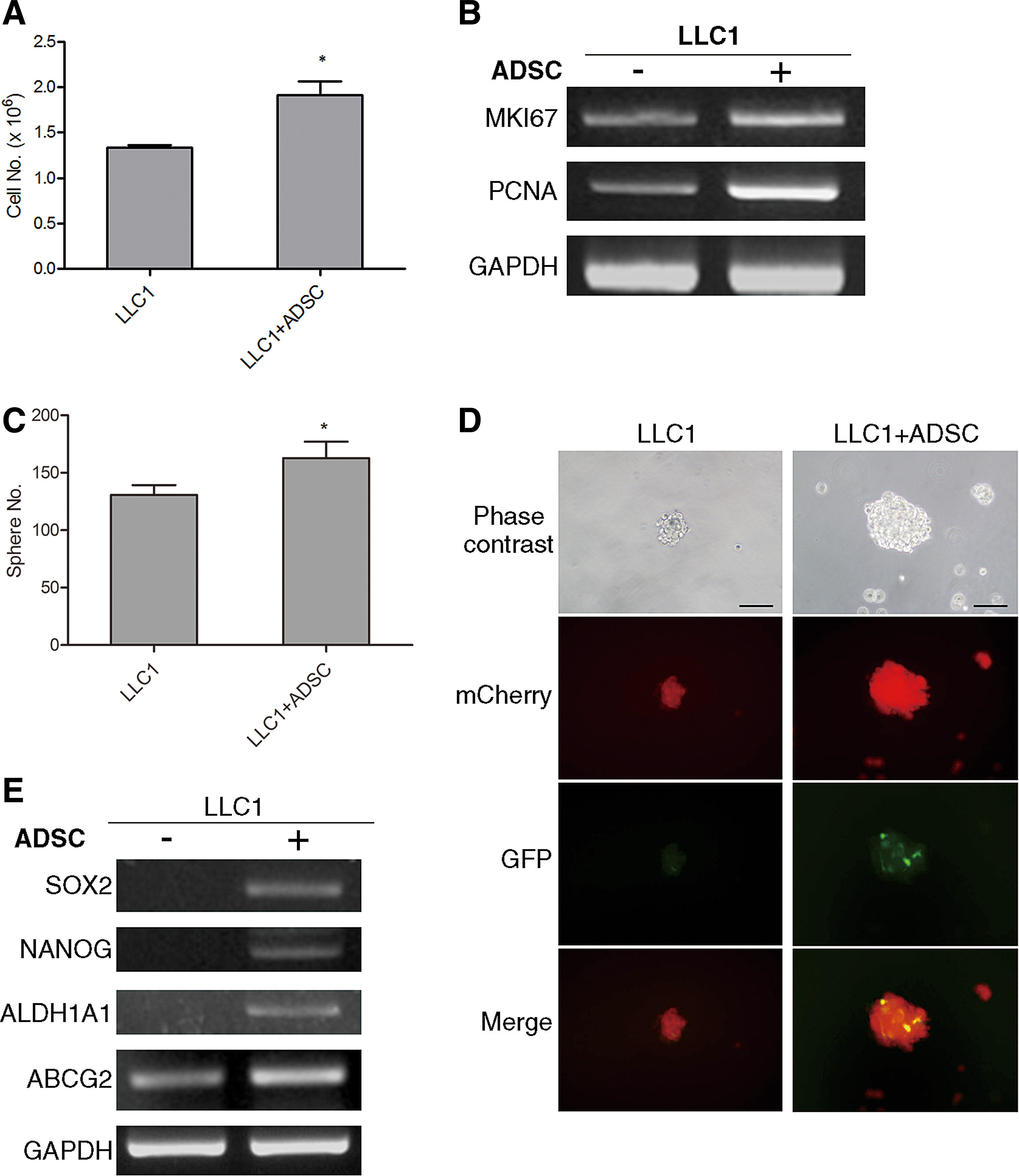
Enhanced growth ability and CSC property of LLC1 cells by ADSC stimulation.
ADSCs enhance the formation and growth rates of Lewis lung carcinoma tumor in mice
To in vivo validate the promoting effects of ADSCs on LLC1 cells, we conducted syngeneic tumor model and noninvasive molecular imaging. Representative images showed that subcutaneous injection of 2 × 105 LLC1 cells alone was able to induce tumor formation in C57BL/6 mice (Fig. 3A); however, no tumors were formed with only 4 × 104 cells (Fig. 3B). Quantitation of tumor volume shows that ADSCs significantly promote not only the growth of tumor (Fig. 3C) but also tumor formation in mice (Fig. 3D). Forty thousand LLC1 cells coinjected with ADSCs form tumors within 1 (60%) to 2 weeks (100%), while LLC1 cells alone formed no tumors in mice (Table 1). In fact, 2 × 105 of LLC1 cells (without ADSCs) were required to initiate tumor formation. Aforementioned result revealed that ADSCs induced de novo tumor formation by the number of LLC1 cells that originally could not form tumors, suggesting that ADSCs have the ability to induce not only accelerated tumor growth but also de novo tumor formation in vivo.

ADSCs initiate tumor formation and accelerate tumor growth of LLC1 cells in mice. Representative bioluminescence images of mice subcutaneously injected with
Bold values represent 100% tumor incidence.
ADSC, adipose-derived stem cell; w/o, without; w/, with.
Interaction of ADSCs and Lewis lung carcinoma cells stimulates secretion of interleukin-6
ADSCs are known as a rich source of cytokines and chemokines, which can communicate with other surrounding cells in a paracrine manner. To understand the crosstalk between ADSCs and Lewis lung carcinoma cells, we cocultured ADSCs and LLC1 cells and then utilized cytokine array to screen the expression of various cytokines and chemokines in their cultured medium. Figure 4A represents the location of various cytokine capture antibodies spotted onto the RayBio Mouse Cytokine Antibody Array Kit. We found that the signal of interleukin-6 (IL-6) spots was very weak in ADSCs alone and almost undetectable in LLC1 alone, whereas the signal expressed very strongly in ADSC+LLC1 coculture (Fig. 4B). The quantitative result showed that the high levels of keratinocyte chemoattractant (KC) and monocyte chemoattractant protein 1 (MCP-1) reflected the additive contributions of the two cell types when cultured on their own. Coculture of ADSCs and LLC1 cells increases the levels of macrophage inflammatory protein 2 (MIP-2), soluble tumor necrosis factor receptor I (sTNFRI), and IL-6. Notably, IL-6 had markedly raised its levels ∼30-fold higher than those produced by ADSC or LLC1 cultured alone (Fig. 4C). Thus, we then utilized RT-PCR analysis to identify the source of increased IL-6 production. The results revealed that ADSCs markedly raised IL-6 expression upon coculture with LLC1 cells (Fig. 4D). We further found that LLC1 cell slightly upregulated IL-6 expression and markedly increased the mRNA expression of IL-6 receptor (IL-6R) while coculture with ADSCs (Fig. 4E), suggesting that the promoting effects of ADSCs on LLC1 cells may act between IL-6 from ADSCs and IL-6R on LLC1 cells. These data demonstrate that the interaction of ADSCs and Lewis lung carcinoma cells stimulates increased production of IL-6 from both cells, but mainly from ADSCs.

Interaction of LLC1 cells and ADSCs induces IL-6 production.
IL-6 contributes to the tumor-promoting effects of ADSCs on Lewis lung carcinoma cells
Aforementioned results show that ADSCs enhanced the malignant characteristics of Lewis lung carcinoma cells, and the interaction of ADSCs with Lewis lung carcinoma cells stimulated IL-6 production, suggesting that IL-6 might correlate with the tumor-promoting effects of ADSCs. To identify the involvement of IL-6 in ADSC-mediated tumor promotion, we utilized neutralizing antibody to deplete IL-6 function. Corresponding to previous results, both cell growth and tumor sphere formation of LLC1 cells were enhanced by ADSCs; however, IL-6-neutralizing antibody significantly diminished the ADSC-induced enhancement of cell growth ability (Fig. 5A, top panel) and sphere-forming efficiency (Fig. 5A, bottom panel) in LLC1 cells. Semiquantitative RT-PCR analysis was then used to determine the impacts of IL-6 on the expression of proliferation and CSC markers in LLC1 cells. Gene expression of proliferation (MKI67 and PCNA) and CSC (SOX2, NANOG, ABCG2, and ALDH1A1) markers was upregulated in LLC1 cells upon coculture with ADSCs, whereas IL-6-neutralizing antibody also markedly abolished the ADSC-induced upregulation of these genes in LLC1 cells (Fig. 5B). Above results suggest that IL-6 plays a critical role that mediates the enhanced malignant characteristics of Lewis lung carcinoma cells by ADSCs.
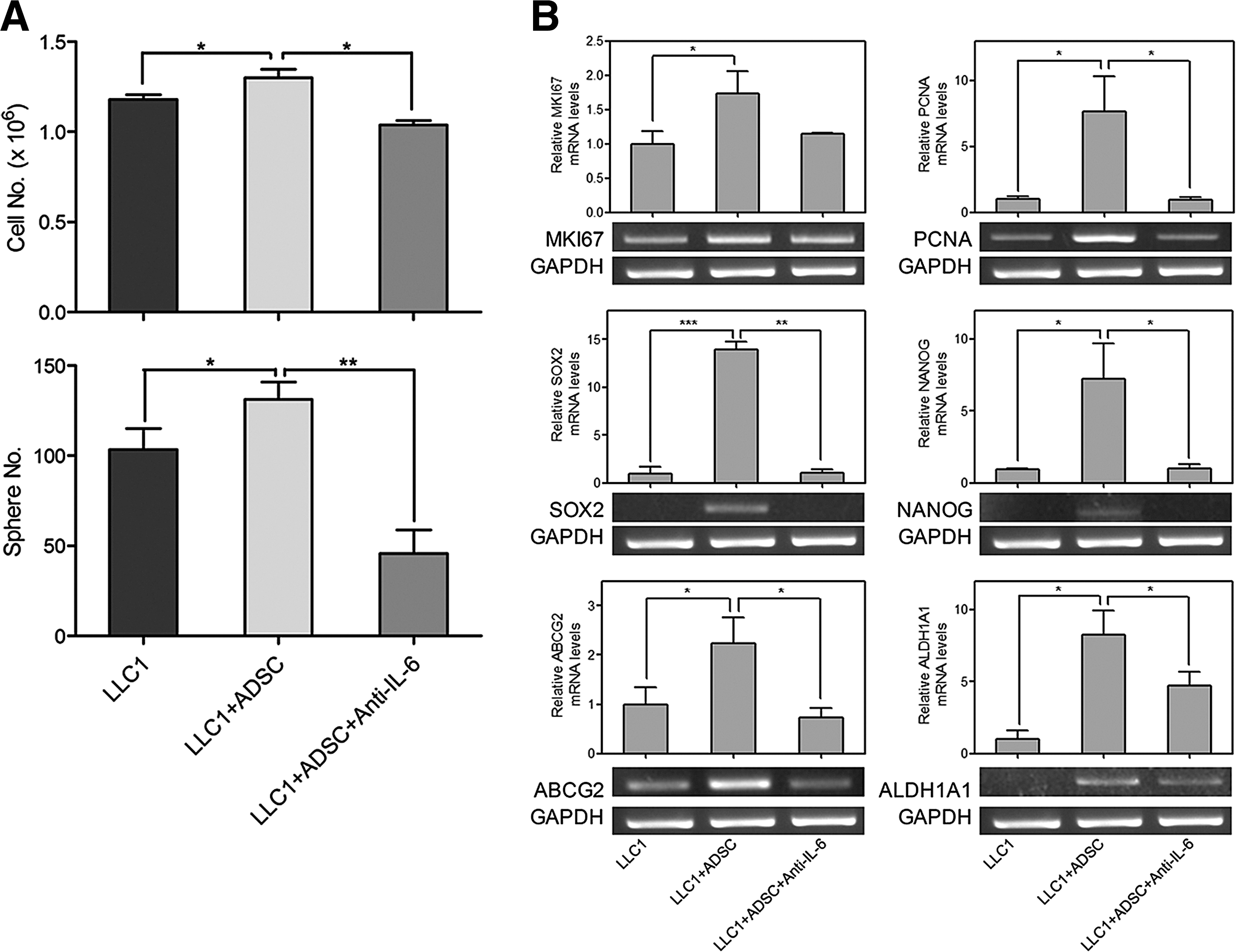
Involvement of IL-6 in ADSC-induced malignant properties of LLC1 cells.
ADSCs induce JAK2/STAT3 activation in Lewis lung carcinoma cells through IL-6
In the interaction of ADSCs and Lewis lung carcinoma cells, we found that ADSC-derived IL-6 played an important role in activating malignant characteristics of LLC1 cells. JAK2/STAT3 pathway has been reported as a predominantly downstream signaling activated by IL-6 [20]. Thus, we conducted western blot assay to evaluate the activation of JAK2 and STAT3 in LLC1 cells upon ADSC coculture. The results show that ADSC coculture slightly induced JAK2 phosphorylation, but dramatically increased the expression of phosphorylated STAT3 (Fig. 6A). Besides, we found that the activation of JAK2/STAT3 pathway in LLC1 cells by ADSCs was also blocked by IL-6-neutralizing antibody. The results suggest that upon interaction with Lewis lung carcinoma cells, ADSCs can induce IL-6 production and then activate the JAK2/STAT3 pathways in Lewis lung carcinoma cells.
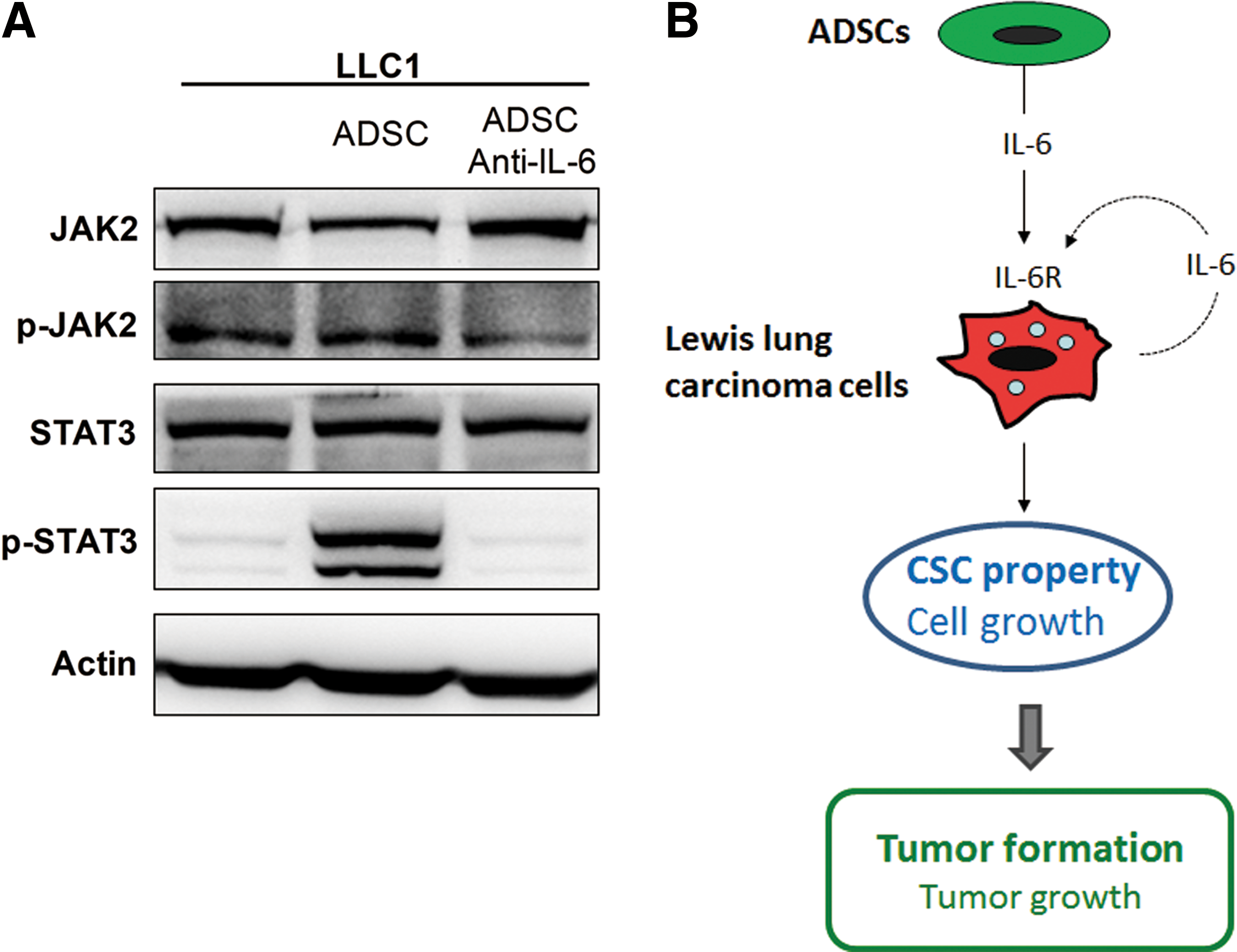
ADSCs activate IL-6-dependent pathway in LLC1 cells.
Discussion
In the past decades, ADSCs have been explored to possess a great potential in regenerative medicine [2,8]. However, accumulating evidence indicates that ADSCs reveal promoting effects on cancer development, raising the concerns of their safety in clinical application [13]. In lung cancer tumor models, ADSCs have been shown to participate in tumor stroma, including differentiation to cancer associated fibroblasts [21 –23] or myofibroblasts [24]. Moreover, they have been proved to promote tumor growth [22,23,25], enhance tumor angiogenesis [21], and induce the epithelial-to-mesenchymal transition of cancer cells [24]. However, little is known about the impacts of ADSCs on tumor initiation and/or CSC property. Previously, we reported that IL-6 mediates the promoting effects of ADSCs on tumor initiation and tumor growth in breast and colon cancer cells [14]. In the present study, we demonstrated that ADSCs enhance the malignant characteristics of Lewis lung carcinoma cells, especially tumor formation and CSC property, through an IL-6-dependent mechanism.
Several types of cells within tumor microenvironment contribute to the development of cancer. Among these cells, MSC plays a critical role during tumor development. Previously, we have demonstrated that BMSCs can migrate to and engraft into microscopic tumor lesions and participate in tumor stroma formation [26]. The knowledge of the biological impacts of MSCs on cancer has been greatly improved recently. However, the underlying effects of MSCs on tumor development remain controversial, especially when MSCs come from different sources. ADSCs have been shown to promote the tumor growth and metastasis of various cancers [22,27,28], whereas conflicting reports reveal the inhibiting impacts of ADSCs on tumor such as antiproliferation and pro-apoptosis [29 –31]. In this study, we showed that ADSCs can enhance the growth ability of Lewis lung carcinoma cells. Moreover, MSCs have been widely reported to promote CSCs in several cancer types, but most of these studies focused on BMSCs [32,33]. Recently, ADSCs have been shown to enhance CSC properties in breast and colon cancers [14,15,34]. In this study, we showed that ADSCs enhanced sphere formation, CSC marker expression, and in vivo tumor formation of Lewis lung carcinoma cells.
Our results reveal that ADSCs enhance the malignant properties of Lewis lung carcinoma cells through an IL-6 paracrine circuit. IL-6 is a pro-inflammatory cytokine with multifunctions that exhibit wide range of biological activities and has been related to many cancer types with a typical pro-tumorigenic effect. IL-6 is one of the major adipokines classically produced by the nonadipocyte members of the adipose tissue [35]. Various cancer types have been shown to exhibit increased production of IL-6, such as breast cancer [36], colorectal cancer [37], lung cancer [38], and cervical cancer [39]. Overexpression of both IL-6 and its receptors was also found in several cancers [40,41]. IL-6 has been demonstrated to reveal several pro-malignant effects on cancer cells, such as promotion of proliferation [42] and metastasis [43], as well as inhibition of apoptosis [44]. Recent reports indicated that IL-6 is implied in CSC development. In breast cancer, IL-6 upregulated the expression of stemness factors SOX2 and OCT4, as well as EMT factors Notch and Wnt through activation of the JAK2/STAT3 pathway. This results in increased breast cancer cell proliferation and self-renewal entry into EMT [45]. In addition to breast cancer, IL-6 and its downstream JAK2/STAT3 signaling have been proved to be critical to CSCs in various cancers, including lung cancer [46,47]. Moreover, IL-6R has been found to be present in CSCs and plays an important role in the proliferation of CSCs in lung cancer [48]. We found that IL-6 contributes to the ADSC-induced enhancement of growth ability and CSC property in lung cancer cells, suggesting a potential therapeutic target for lung cancer. Song et al. identified that IL-6 is elevated in subsets of human nonsmall cell lung cancers. They suggested that further study of IL-6 targeting with IL-6 neutralizing antibody should be considered in human clinical trials [49].
In conclusion, we demonstrated that ADSCs reveal promoting effects on the tumor development of lung cancer cells. Interaction of ADSCs and lung cancer cells stimulates increased production of IL-6, which then acted in a paracrine and slightly autocrine manner on cancer cells to enhance their malignant properties, including cell growth and especially CSC property. Moreover, ADSCs promoted the in vivo tumor development of Lewis lung carcinoma cells by enhancing tumor formation and growth capacities (Fig. 6B). IL-6 is involved in increased expression of genes related to cell proliferation and CSCs, as well as activation of JAK2/STAT3 in cancer cells. Our findings, for the first time, indicate that ADSCs promote CSC property and in vivo tumor formation in lung cancer.
Footnotes
Acknowledgments
The authors thank Core Facility Center, Office of Research and Development (Taipei Medical University, Taipei, Taiwan) for experimental instruments (IVIS Imaging System 200 Series and Becton Dickinson FACSCanto II). This work was supported by grants from the Ministry of Science and Technology (MOST 104-2221-E-038-016 and 104-2313-B-038-001) and the Council of Agriculture, Executive Yuan (104 AS-16.3.1-ST-a8 and 105 AS-13.3.1-ST-a2).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.