
Abstract
Cardiac adipose tissue-derived stem cells (cASCs) have the ability to differentiate into multiple cell lineages giving them a high potential for use in regenerative medicine. Cardiac fat tissue still raises many unsolved questions related to its formation and features. P2Y nucleotide receptors have already been described as regulators of differentiation of bone-marrow derived stem cells, but remain poorly investigated in cASCs. We defined, in this study, the P2Y4 nucleotide receptor as a negative regulator of cardiac fat formation and cASC adipogenic differentiation. Higher expression of P2Y4 receptor in cardiac fat tissue was observed compared to other adipose tissues. P2Y4-null mice displayed a higher mass of cardiac adipose tissue specifically. We therefore examined the role of P2Y4 receptor in cASC adipogenic differentiation. An inhibitory effect of uridine 5′-triphosphate (UTP), ligand of P2Y4, was observed on the maturation state of differentiated cASCs, and on the expression of adipogenesis-linked genes and adiponectin, a cardioprotective adipokine. Higher adiponectin secretion by P2Y4-null adipocytes could be linked with cardioprotection previously observed in the heart of P2Y4-null ischemic mice. We realized here left anterior descending artery ligation on simple and double-knockout mice for P2Y4 and adiponectin. No cardioprotective effect of P2Y4 loss was observed in the absence of adiponectin secretion. In addition, P2Y4 loss was correlated with higher expression of UCP-1 (uncoupling protein-1) and CD137, two markers of brown/beige cardiac adipocytes. Our data highlight the P2Y4 receptor as an inhibitor of cardiac fat formation and cASC adipogenic differentiation, and as a potential therapeutic target in the regulation of cardioprotective function of cardiac fat.
Introduction
I
Epicardial fat exhibits cardioprotective properties through local secretion of anti-inflammatory and antiatherogenic adipokines, such as adiponectin and adrenomedullin [2]. Cardioprotective effects of adiponectin are attributed to its ability to suppress cardiomyocyte apoptosis, oxidative stress, and inflammation [6,7]. Chronic coronary artery disease and obesity are thought to downregulate EAT cardioprotective adipokine secretion [8].
Recently, a new potential cardioprotective property of EAT was discovered. The expression of uncoupling protein-1 (UCP-1), the gold marker of brown fat, was found significantly higher in epicardial fat compared to other fat depots [9]. It has been hypothesized that EAT could act like brown fat to defend the myocardium against hypothermia and reactive oxygen species. Moreover, UCP-1 expressed in the heart of transgenic mice protects against ischemia–reperfusion damages to improve contractility and reduce oxidative stress [10].
There is a growing interest in the identification of the mechanisms that regulate the balance between protective and detrimental effects of EAT. The biology of this tissue remains unclear, partly because it is considered lacking in most common experimental models, like rodents, avians, or amphibians. However, mice have a fat depot covering the left ventricle of the heart named cardiac adipose tissue [11]. Little is known about its formation and its communication with the heart. It was recently demonstrated that this cardiac fat depot has the ability to express UCP-1 in response to cold stimulation as it is observed in human EAT [11].
Adipose tissue-derived stem cells (ASCs) are a new promising source of stem cells for cardiovascular repair. In humans, Bai et al. have demonstrated that transplantation of freshly isolated or cultured ASCs into ischemic hearts after acute myocardial infarction promotes cardiac function [12]. Cardiac adipose tissue-derived stem cells (cASCs) isolated from C57BL/6 mice have a higher potential of differentiation in cardiovascular cells in vitro than other fat depots and directly contribute to the regeneration of myocardium in a model of myocardial infarction [13].
Myocardial ischemia is associated with a release of extracellular nucleotides by the cardiac endothelial cells and cardiomyocytes [14]. The P2Y4 receptor is a uridine 5′-triphosphate (UTP) receptor coupled to the phosphoinositide/Ca2+ pathway [15]. Recombinant rat and mouse orthologs of human P2Y4 receptor are activated by UTP and ATP, as described for the ubiquitously expressed mouse P2Y2 receptor [16 –18]. The P2Y4 receptor is involved in postnatal cardiac development [19], exercise tolerance [20], and exercise-induced cardiac hypertrophy [20]. Interestingly, P2Y4 receptor was reported as a potential pivotal regulator in osteogenic and adipogenic differentiation of human mesenchymal stem cells [21].
Both P2Y2 and P2Y4 nucleotide receptors are involved in cardioprotection, but through very distinct mechanisms and effects. Activation of P2Y2 nucleotide receptor protects rat cardiomyocytes against hypoxic stress, and administration of UTP to rats reduces infarct size and enhances myocardial function [22]. We recently demonstrated that loss of mouse P2Y4 receptor is associated with a protection against infarction and reduction of cardiac inflammation, permeability, and fibrosis [23].
In this study, we identified high P2Y4 expression in cardiac adipose tissue and investigated its potential role in the formation and the cardioprotective function of cardiac fat.
Materials and Methods
Ethics statement
All mice used in this study were authorized by the Animal Care Use and Review committee of the Free University of Brussels.
Animals
P2Y4-null C57BL/6 mice (symbolized as P2Y4 0/− for male and P2Y4 −/− for female due to its location on X chromosome) were generated in our institute as described in Robaye et al. [24]. The adiponectin-null C57BL/6 mice (AdipoQ−/−) were obtained from Jackson Laboratory (L'Abresle, France). The double- null mice for P2Y4 and AdipoQ were generated in our institutional animal facility (Campus Erasme, ULB, Belgium).
ASC isolation and differentiation
Isolation
Cardiac or gonadal adipose-derived stem cells (cASCs or gASCs) were isolated from the stromal vascular fraction of cardiac or gonadal adipose tissue (CAT or GAT), respectively [25]. The adipose tissue was minced and incubated in collagenase A (2.5 g/L) at 37°C for 45 min. The resulting suspension was filtered through a nylon mesh (500 nm) and separated into two fractions (mature adipocytes and stromal vascular fraction) by centrifugation at 500 g for 5 min. The lysis of red blood cells was performed by osmotic shock and the ASCs were selected by adhesion to plastic.
Culture
ASCs were cultured for 3 days in a proliferation medium (DMEM:F-12, Gibco™; ThermoFisher Scientific, Waltham, MA), containing 3% newborn calf serum and 1% penicillin/streptomycin, and for 4 days in an adipogenic differentiation medium (the proliferation medium supplemented with 50 μM indomethacin, 1 μM dexamethasone, 0.5 μM isobutylmethylxanthine, and 5 μg/mL insulin). During differentiation, the cultures were stimulated with a physiological agonist of P2Y4/P2Y2, UTP, the P2Y4 specific agonist, MRS4062, and the P2Y2 specific agonist, MRS2768. The number of differentiated cells was measured by Oil Red O staining (Sigma-Aldrich, St Louis, MO). The percentage of differentiated adipocytes is referring to the ratio between the number of cells with lipid droplets (stained in red by Oil Red O) and the total number of cells in the field (nucleus stained in blue by Hoechst).
Lipid droplet size measurement
For quantitative analysis of adipocytes, the cultures were stained with Oil Red O after 4 days of adipogenic differentiation. The diameter of lipid droplets was measured using Axiovision 4.6.3 software (Carl Zeiss). About 50–60 cells for each condition, which correspond to 400–600 lipid droplets, were analyzed in two independent experiments. The percentage is referring to the ratio of lipid droplets with a defined diameter (four different categories) to the total number of lipid droplets measured. The four categories of size were arbitrarily selected as previously reported [26].
Hypoxic in vitro experiments
Cultures of cASCs were differentiated in the adipogenic medium for 4 days and incubated during the last 24 or 48 h in the differentiation medium in normoxic conditions, or at 1% of O2. For each culture experiment, a pool of cASCs isolated from 4 to 5 mice was used.
Adiponectin quantification in adipocyte supernatants
Supernatants of cASCs isolated from P2Y4 wild-type and null mice and differentiated for 4 days in the adipogenic medium were collected and total proteins were quantified using the Pierce™ BCA Protein Assay Kit (ThermoFisher Scientific). Adiponectin (Quantikine kit; R&D System, Abingdon, United Kingdom) level was measured by enzyme-linked immunosorbent assay (ELISA), according to the manufacturer's procedure.
UCP-1 staining
As described above, ASC cultures were differentiated in the adipogenic medium for 4 days and then incubated for 48 h in hypoxic conditions. Cultures were then fixed for 5 min in paraformaldehyde 4%. Cells were incubated with UCP-1 primary antibody over night at 4°C (Abcam, Cambridge, United Kingdom) at 1/500 dilution. An AlexaFluor®488-conjugated anti-rabbit IgG (1/400) was used as secondary antibody (Jackson Immuno Research Laboratory, West Grove, PA). Nuclei were stained using Hoechst (ThermoFisher Scientific). Pictures of immunostaining were acquired at room temperature in FluorSave Reagent (Calbiochem®; Merck, Darmastadt, Germany) using an Axio Observer Z1 microscope, a high-resolution charge-coupled device camera, and Axio Vision 4.6.3 software (Carl Zeiss). UCP-1 staining was quantified using Image J software.
Ischemia in vivo experiments: left anterior descending artery ligation
P2Y4 0/+/AdipoQ+/+, P2Y4 0/−/AdipoQ+/+, P2Y4 0/−/AdipoQ−/−, and P2Y4 +/+/AdiopoQ−/− mice aged 8 to 12 weeks were anesthetized with Avertin® (240 mg/kg; Sigma-Aldrich), intubated, and ventilated. A left side thoracotomy was performed, and the pericardium was incised. Myocardial infarction was then induced by the permanent ligation of the left anterior descending artery (LAD), proximal to its bifurcation from the main stem as previously described in [23]. Twenty-four hours after LAD ligation, the cardiac adipose tissue, the heart, and others fat tissues were collected, washed in cold phosphate-buffered saline, dried, weighted, and stored at −80°C for mRNA, or stained by 2,3,5-triphenyl-tetrazolium-chloride on 1-mm heart sections (TTC; Sigma-Aldrich).
Hematoxylin/eosin staining
Tissues were collected from P2Y4 +/+ and P2Y4 −/− mice aged 8 to 12 weeks and fixed in paraformaldehyde 4% and paraffin embedded (Histokinette®). Cardiac fat sections were cut at 4 μm thickness on precoated slices (VWR International, Heverlee, Belgium) and stained in hematoxylin/eosin. Pictures of hematoxylin/eosin staining were acquired using Zeiss AxioImager Z1 [Light Microscopy Facility (LiMiF), Erasme, Belgium]. The surface of adipocytes was measured using Axiovision software.
Quantitative reverse transcription polymerase chain reaction experiments
Total mRNA was extracted using TRIzol reagent (Life Technologies, Groningen, The Netherlands) and RNeasy kit column (Qiagen, Hilden, Germany) from cardiac ASCs, differentiated adipocytes, and adipose tissue from P2Y4 +/+ and P2Y4 −/− mice. mRNA was reverse transcribed using random hexamers and Superscript II Reverse Transcriptase (ThermoFischer Scientific). Reverse transcription polymerase chain reaction (RT-PCR) amplification mixtures contained 10 ng template cDNA. Reactions were run on a 7500 Fast Real-Time PCR System (Applied Biosystems, Foster City, CA). PCR data were normalized for each gene to RPL32 housekeeping gene.
Statistics
Data were expressed as mean ± standard error of the mean (SEM) for in vitro and in vivo studies. Endpoint comparisons with two groups were performed using an unpaired two-tailed Student's t-test and Mann–Whitney test for parametric and nonparametric data, respectively (Prism Software, version 5; GraphPad, San Diego, CA). For parallel repeated-measures studies, two-way ANOVA was used with Bonferroni post hoc evaluations to determine the significance for individual time point (Prism Software; GraphPad). For all studies, P value was considered *P < 0.05; **P < 0.01; ***P < 0.001.
Results
Increased mass of cardiac adipose tissue in P2Y4-null mice
Quantitative PCR experiments were performed to evaluate P2Y4 expression in CAT, GAT, and interscapular adipose tissue (IAT) (Fig. 1A). The GAT is typically characterized as white adipose tissue (WAT) and IAT as brown adipose tissue (BAT). P2Y4 expression was normalized to RPL32 expression and detected in all three adipose tissues, but more strongly in CAT (Fig. 1A).
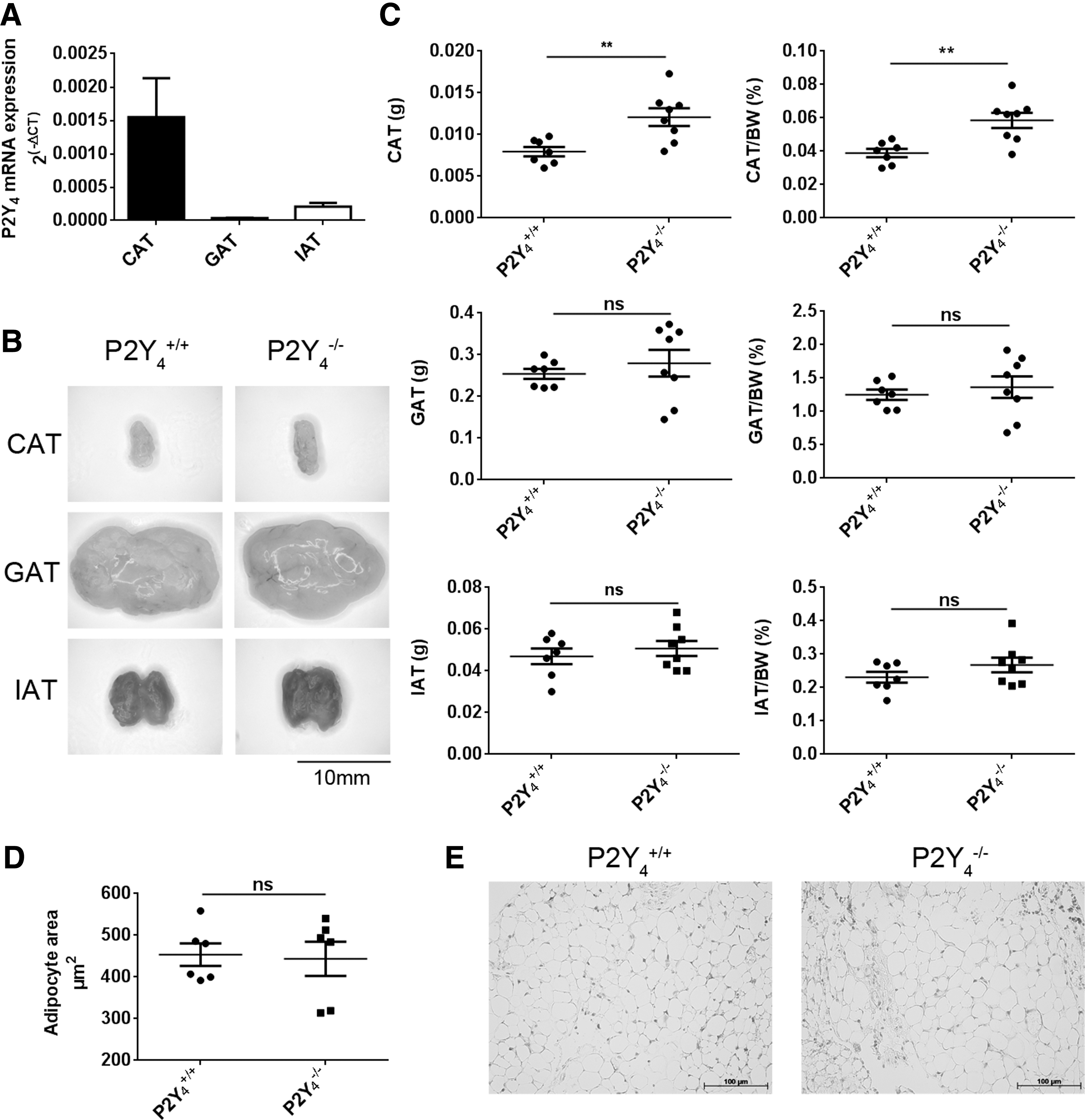
Increased cardiac fat mass in P2Y4 receptor-null mice.
We then identified a higher mass of CAT in P2Y4-null mice, whereas GAT and IAT weights were similar in P2Y4-null and wild-type mice (Fig. 1B, C). The increase in CAT weight normalized to body weight was 33% ± 6.8% in P2Y4-null mice compared to wild-type female mice (mean ± SEM; **P < 0.01). Similar data were obtained in male mice (data not shown). This increase of CAT mass in P2Y4-null mice was not associated with a hypertrophy of cardiac adipocytes (Fig. 1D, E). Our data highlight significant P2Y4 receptor expression in CAT among other adipose tissues and its potential involvement in CAT formation.
Effect of UTP on cardiac ASC adipogenic differentiation
The gain of CAT mass in P2Y4 mice led us to study the role of P2Y4 receptor in adipogenic differentiation. We used the classical model of adherent stromal vascular fraction as source of cASCs [25]. cASCs were isolated from cardiac fat of P2Y4-null and wild-type mice and plated in the proliferation medium for 3 days. cASCs were then cultured in the adipogenic differentiation medium containing a mixture of dexamethasone-IBMX-insulin-indomethacin. First, the expression of both P2Y4 and P2Y2 UTP receptors were followed during cASC differentiation into adipocytes until day 21 after plating (Fig. 2A). P2Y4 mRNA expression data displayed two peaks at day 3 and 14 after plating, whereas the P2Y2 receptor expression was relatively stable during differentiation (Fig. 2A).

P2Y4 receptor and its ligand UTP have inhibitory effects on adipogenesis.
We then further investigated a potential involvement of UTP in the early adipogenic differentiation process. We stimulated cASC cultures by addition of a daily dose of UTP—common physiological ligand of the P2Y4 and P2Y2 receptors—in the differentiation medium. In view of early expression of P2Y4 and to investigate its involvement in the early process of adipogenic differentiation, the effect of UTP stimulation was studied already 4 days after differentiation. We quantified, thus, the percentage of cells displaying lipid droplets after Oil Red O staining after 4 days of differentiation (Fig. 2B–E). We observed a concentration-dependent negative effect of UTP treatment on adipogenic differentiation (Fig. 2B).
Then, we decided to compare the effect of UTP to that of two specific receptor agonists: MRS4062, for P2Y4, and MRS2867, for P2Y2 (Fig. 2C). MRS4062 displays a negative effect comparable to that of UTP, whereas no effect was observed after MRS2867 treatment. These results suggest an involvement of P2Y4 receptor activation in the negative effect of UTP on adipogenic differentiation. To confirm the involvement of P2Y4 receptor, we isolated cASCs from P2Y4-deficient mice (Fig 2D). At a concentration of 100 μM UTP, the inhibition of adipogenic differentiation of wild-type cASCs was 42.5% ± 12.9% (mean ± SEM; **P < 0.01) (Fig. 2D, E). The negative effect of UTP on adipogenic differentiation was lost in P2Y4-null cASC cultures (Fig. 2D, E). Lipid droplets grow by the incorporation of triglycerides or by fusion of other small lipid droplets during adipogenesis [26,27]. A fully mature white adipocyte exhibits a large unilocular lipid droplet.
On day 7, we already observed the detachment of some lipid droplets from differentiated cells into culture supernatants (data not shown). We have here quantified lipid droplets interms of their diameter at early time (4 days) of adipogenic differentiation (Fig. 3A and Table 1). Four different categories of lipid droplet size have been defined as previously described [26]. We observed a positive effect of UTP on the percentage of small droplets (<2 μm) and a negative effect on the percentage of large droplets (>6 μm), which was lost in P2Y4-null adipocytes (Fig. 3A and Table 1). The negative effect of UTP on adipogenic differentiation was correlated with its negative action on the mRNA expression of peroxisome proliferator-activated receptor-γ (PPAR-γ), stearoyl-CoA desaturase-1 (SCD1), and thyroid hormone responsive (THRSP or Spot14), three genes involved in adipogenesis (Fig. 3B).
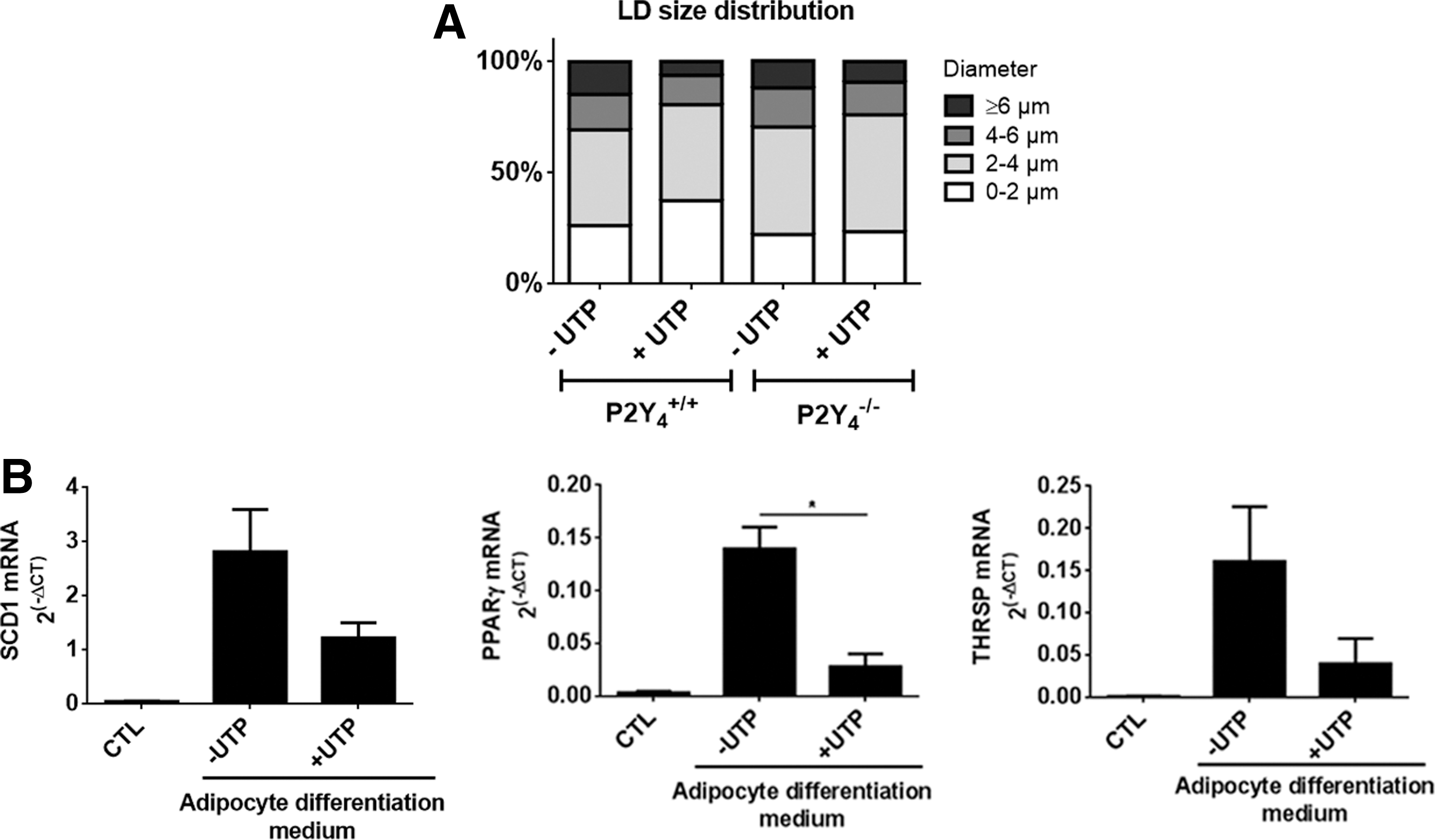
UTP regulates LD size and adipogenesis-related genes during differentiation of cASCs.
LD diameter was measured using Axio Vision software after Oil Red O staining of cASC cultures after 4 days of adipogenic differentiation. The percentage refers to the ratio of LD with a defined diameter (four categories) to the total number of LDs measured. Values represent mean ± SEM.
P < 0.05, compared to P2Y4 +/+ without UTP. For each condition, about 50–60 cells were analyzed corresponding to 400–600 LDs in two independent experiments.
cASCs, cardiac adipose tissue-derived stem cells; LD, lipid droplet; SEM, standard error of the mean; UTP, uridine 5′-triphosphate.
The staining and quantification of lipid droplets and the analysis of adipogenesis-related gene regulation in our differentiated ASC cultures support that UTP is a negative regulator of adipogenic differentiation of cardiac ASCs.
Evaluation of adiponectin role in the cardioprotective effect of P2Y4 loss
Our laboratory has previously demonstrated a protection against cardiac infarction in P2Y4-null mice in a model of myocardial ischemia based on ligation of the left anterior descending artery [23]. Adiponectin is an adipose tissue-secreted adipokine displaying cardioprotective properties [6 –8]. We investigated if adiponectin could play a role in the cardioprotective effect of P2Y4 loss. First, we evaluated the potential regulation of adiponectin expression by UTP in differentiated cASCs by quantitative polymerase chain reaction (qPCR) and ELISA (Fig. 4A).
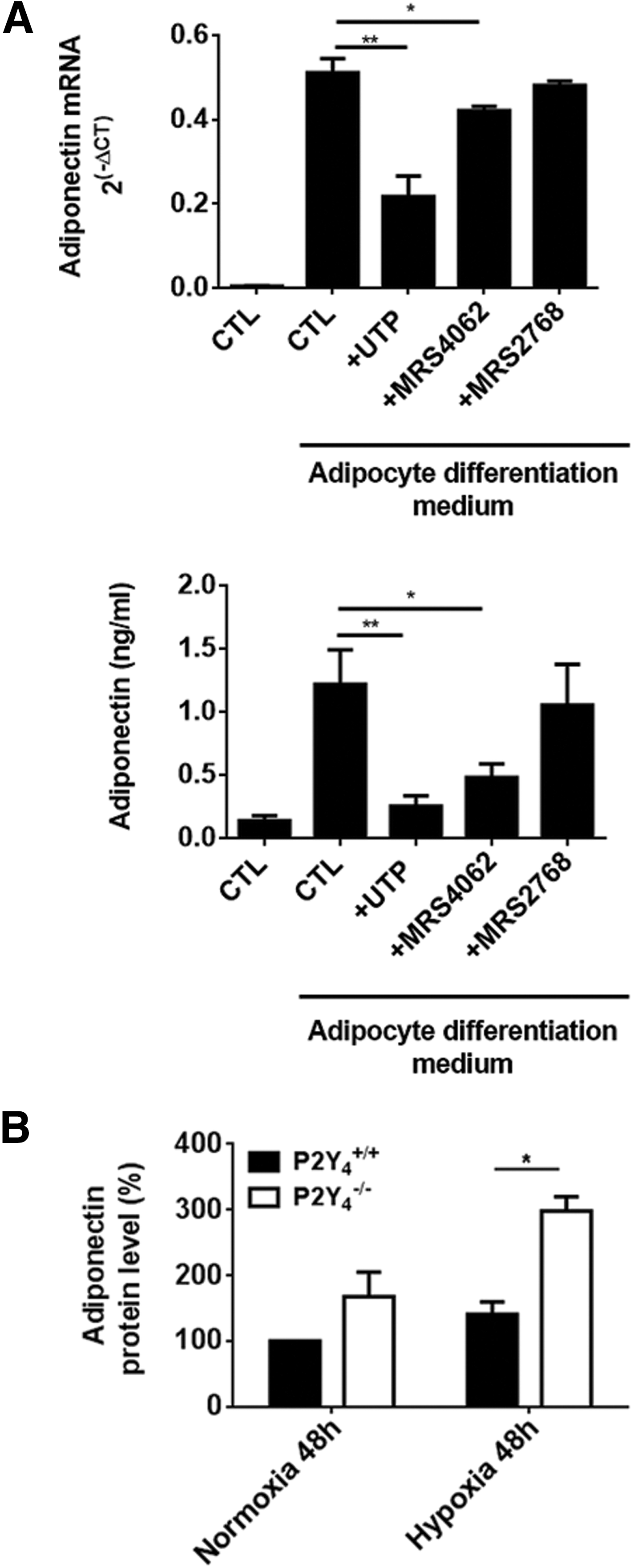
Increased adiponectin expression in P2Y4-null adipocytes differentiated from cASCs.
We observed a negative effect of daily stimulation by UTP on mRNA expression and secretion of adiponectin in wild-type cardiac adipocytes (Fig. 4A). The effect of P2Y4-specific agonist, MRS4062, was comparable to that of UTP, whereas no significant effect of P2Y2-specific agonist, MRS2768, was observed (Fig. 4A). P2Y4 appears, thus, as the receptor responsible for UTP-mediated negative regulation of adiponectin in differentiated cASCs, possibly through its involvement in adipocyte differentiation/maturation.
The role of adiponectin in cardioprotection has been mostly studied under ischemic conditions [6 –8]. We compared adiponectin secretion by our ASC-differentiated cultures in normoxic and hypoxic conditions. We evaluated in vitro adiponectin secretion by wild-type and P2Y4-null cASCs after 4 days of adipogenic differentiation with the last 48 h in normoxic (20% O2) or hypoxic (1% O2) conditions (Fig 4B). After 48 h in hypoxia, we observed higher concentrations of adiponectin in the supernatant of P2Y4-null cardiac adipocytes compared to wild-type cardiac adipocytes (Fig. 4B).
We then performed LAD ligation on P2Y4/AdipoQ simple and double-knockout mice. The percentage of irreversible infarcted area is defined by the ratio between infarcted tissue weight and left ventricular weight (IA/LV). We obtained the expected data using simple P2Y4- or AdipoQ-null mice: a cardioprotection in P2Y4-null mice [23] and increased infarct size in AdipoQ-null mice [7] compared to WT mice (Fig. 5). P2Y4/AdipoQ double-null mice displayed a higher infarct size than WT and P2Y4-null mice. There was no significant difference in infarct size between P2Y4/AdipoQ double-null mice and AdipoQ-null mice (Fig. 5). These results were also confirmed by a 2,3,5-triphenyl-tetrazolium-chloride staining of heart sections, which showed the unaffected part in red and the infarcted zone in white (Fig. 5). The cardioprotective effect related to P2Y4 loss appears to be dependent on the presence of adiponectin.
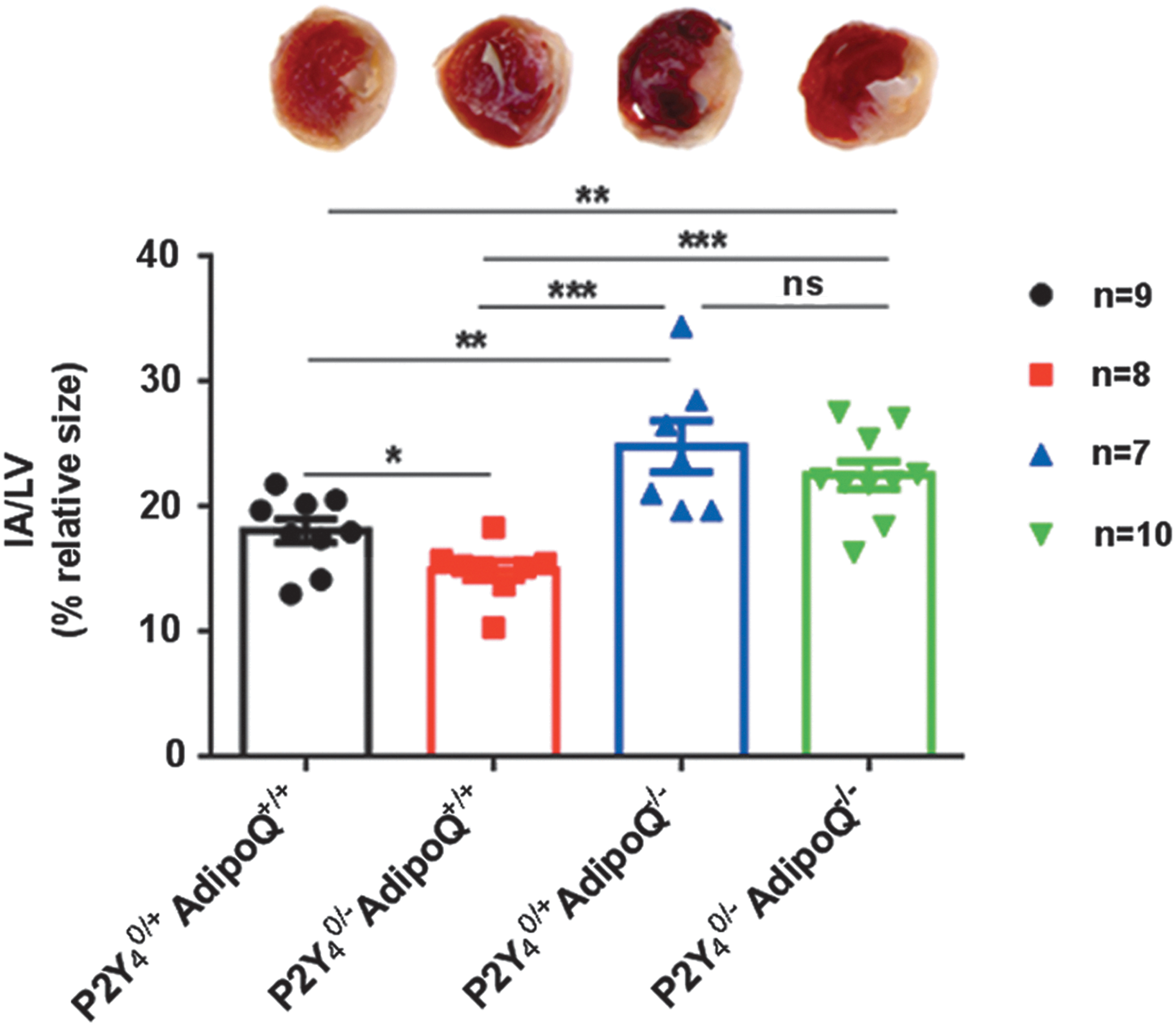
Infarct size in simple and double P2Y4-/AdipoQ-knockout mice after LAD. Infarct size was determined as the ratio IA/LV 24 h after LAD ligation. The infarcted area is illustrated in white on pictures of mice heart ventricular sections with triphenyl-tetrazolium-chloride. Measures were performed on four groups: 9 wild-type mice (P2Y4 0/+ AdipoQ+/+), 8 P2Y4-knockout mice (P2Y4 0/− AdipoQ+/+), 8 adiponectin-knockout mice (P2Y4 0/+ AdipoQ−/−), and 10 double-knockout mice for adiponectin and P2Y4 (P2Y4 0/− AdipoQ−/−). Values represent mean ± SEM: *P < 0.05, **P < 0.01 and ***P < 0.001. IA/LV, infarcted tissue weight and left ventricular weight; LAD, left anterior descending artery.
Increased level of beige adipocyte markers in P2Y4-null adipocytes in hypoxic conditions
Immunofluorescence staining of UCP-1, the gold marker of beige/brown fat, was performed to evaluate UCP-1 expression in P2Y4-null differentiated cardiac ASCs in hypoxic conditions (Fig. 6A). ASCs were placed for 4 days in the adipogenic differentiation medium and in hypoxic conditions (1% O2) during the last 48 h. We observed a higher number of P2Y4-null cardiac adipocytes expressing significant UCP-1 than wild-type cardiac adipocytes (Fig. 6A). A comparable experiment was realized with gASCs. No increase in UCP-1-positive cells was observed in P2Y4-null gonadal adipocytes compared to wild-type gonadal adipocytes (Fig 6A). The increase in UCP-1-positive P2Y4-null cells appears, thus, to be specific to cardiac adipocytes.
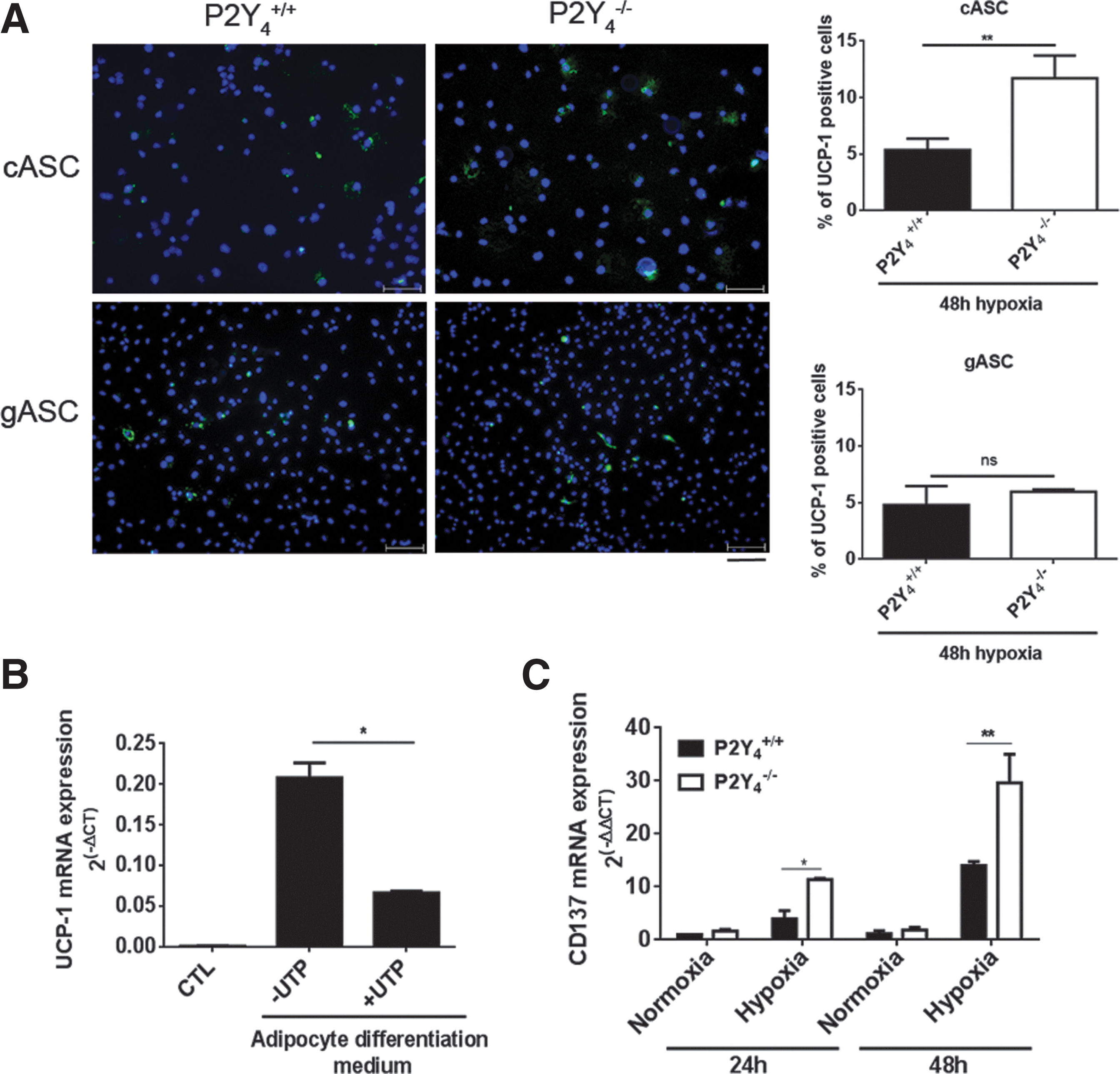
Increased level of beige adipocyte markers in P2Y4-null adipocytes in hypoxic conditions.
We also confirmed a negative effect of UTP on UCP-1 mRNA expression in cardiac differentiated ASCs (Fig. 6B). The mRNA expression of another beige adipocyte marker, CD137, was studied in wild-type and P2Y4-null differentiated cASCs in normal and hypoxic conditions (Fig. 6C). We observed an increase of CD137 in response to hypoxia at 24 and 48 h. Interestingly, CD137 mRNA level was higher in P2Y4-null adipocytes, respectively, after 24- and 48-h hypoxia (Fig. 6C). The identification of P2Y4 receptor as a potential actor involved in the regulation of white to beige cardiac adipocyte formation could present applications in the treatment of cardiovascular diseases.
Discussion
Human cardiac fat is composed of epicardial fat, located between myocardium and visceral pericardium, and pericardial fat, located outside the visceral pericardium [3]. Human EAT is considered an active endocrine organ [2,28], whereas secretion of adipokines by pericardial fat is still unclear [29]. The EAT displays a dichotomous role: under physiological conditions it exhibits biochemical and thermogenic cardioprotective properties [3], but in obesity, it is defined as a marker of visceral fat and cardiovascular risk [3,30]. In mice, the role of the EAT is less defined and its existence is even debated. The fat depot lining the left ventricle of the mice heart, down to the apex, is generally called the cardiac adipose tissue. However, recently, Yamaguchi et al. have demonstrated that rodents possess EAT in the atrioventricular groove derived from epicardium [31].
In this study, we have observed a gain of cardiac adipose tissue mass in mice deficient for the P2Y4 receptor. No obesity phenotype or increased mass of other adipose tissues were observed in P2Y4-null mice. The specificity of the increase in cardiac fat is consistent with higher P2Y4 expression in cardiac adipose tissue compared to other fat depots. We have thus investigated a potential role of P2Y4 in cardiac adipose tissue formation and function. P2Y subtypes such as P2Y4 and P2Y14 have already been described as potential pivotal regulators in early mesenchymal stem cell commitment [32]. They are modulated during osteogenic and adipogenic differentiation of human dental follicle cells and adipose stem cells derived from liposuction aspirate [32].
To investigate the impact of P2Y4 loss in cardiac adipose tissue formation, we have studied adipogenic differentiation of stem cells isolated from P2Y4-null and WT cardiac fats. We have observed a significant early expression of P2Y4 receptor in cardiac adipose tissue-derived stem cells (cASCs). We investigated the role of P2Y4 receptor in the first steps of adipogenic differentiation. We observed effects of P2Y4 loss and UTP on lipid droplet size in the first days of ASC differentiation. The lipid droplets are dynamic intracellular organelles involved in lipid metabolism and intracellular trafficking and signaling [27]. During adipogenesis, the lipid droplets grow in size by incorporating triglycerides or by fusion of small droplets [26].
The stimulation of cASCs during adipogenic differentiation with specific P2Y4 receptor agonist MRS4062 or UTP, inhibited the generation and maturation of adipocytes from cASCs, an effect lost in P2Y4-null mice. This effect was correlated with a negative action of UTP on adipogenesis-linked genes (PPAR-γ, SCD1, and THRSP). THRSP inhibition was previously associated with an altered morphology of lipid droplet [33]. Less is known on molecular mechanisms underlying the adipose lipid droplet formation, but some human and mice model deficient for droplet-associated proteins such as caveolin present abnormal droplet growth, which affects the integrity and function of adipose tissue (insulin sensitivity, inflammation, lipoatrophy, and dystrophy, etc.) [34 –36].
Cardiac fat constitutes a source of stem cells known to have a high proliferation activity and a prominent potential for differentiation into cardiovascular cells compared with those isolated from other adipose depots [13]. This cardiac adipose tissue has also a very interesting location, really close to the heart and able to interact with it by the secretion of adipokines. Transplantation of adipocyte cell sheets generated from adipose stem cells on the surface of the heart has a cardioprotective effect by reducing the infarct size and inflammation in ischemic mice [37]. This beneficial effect is attributed to cardioprotective factors secreted by induced adipocyte cell sheets, including adiponectin, hepatocyte growth factor and vascular endothelial growth factor [38].
Recently, our laboratory has demonstrated that cardioprotection against myocardial infarction in mice deficient for the P2Y4 receptor correlated with reduced endothelin-1 (ET-1) level and lower cardiac inflammation and permeability [23]. Adiponectin, an abundant circulating protein secreted from adipose tissue, is known to have suppressive effects on ET-1-induced hypertrophy in cultured cardiomyocytes [38] and to protect the heart against ischemic injury [7]. This study demonstrates an inhibitory effect of UTP on adiponectin mRNA and adiponectin secretion in adipocytes differentiated from cASCs. Moreover, higher secretion of adiponectin was observed in P2Y4-null adipocytes compared to WT adipocytes differentiated from cASCs.
To investigate if the cardioprotection previously observed in P2Y4-null mice [23] could be correlated at least partially to adiponectin regulation, we performed LAD ligation experiments with simple and double-knockout mice for both P2Y4 and adiponectin (AdipoQ) genes. As expected, we observed a cardioprotection in P2Y4-null mice and increased infarct size in AdipoQ-null mice compared to WT mice. Interestingly, there was no significant difference in infarct size between AdipoQ-null and P2Y4/AdipoQ double-knockout mice. These data suggest that the cardioprotective effect of P2Y4 loss could be due to an increased production of adiponectin.
The identification of P2Y4 as a specific and negative regulator of cardiac fat mass could have interesting applications. Several studies established a relationship between cardiac fat volume and cardiovascular disease risk [39,40]. In humans, epicardial fat thickness has been linked to potential coronary circulation dysfunction [40,41]. An increase in this fat volume is often associated with obesity, but could be potentially associated with lower cardiovascular risk in nonobese patients. Higher cardiac fat mass could have cardioprotective effects, but the nature and features of the adipocyte population, which is expanded, have to be determined, and the discrepancies between human and mouse epicardial fat have to be considered.
The effect of UTP and P2Y4 loss on adipogenic differentiation and lipid droplet size could explain the regulation of adiponectin secretion, but cannot be directly linked to the regulation of specific subsets of white and beige adipocytes. In humans, the expression of UCP-1 has been reported to be significantly higher in epicardial fat than in other fat depots, suggesting that EAT might function as brown fat to defend the myocardium and coronary artery against hypothermia and oxidative stress [42]. In mice, we have then focused on the potential involvement of P2Y4 in the regulation of markers of these so-called beige adipocytes. They express brown fat-specific genes such as UCP-1, but also beige-specific enriched markers such as CD137, TbX1, Tmem26, Cited1, CD40, and Shox2 [43] in response to β-adrenergic agonists or PPAR-γ [44].
The identification of the mechanisms and regulators involved in the WAT to BAT transdifferentiation has many therapeutic applications in the development of treatments for obesity and type 2 diabetes [45,46]. Interestingly, P2Y4 loss amplified hypoxia-mediated increase of UCP-1 and CD137 expression in cardiac adipocytes. More precisely, the number of UCP-1-positive cells is higher in P2Y4-null than WT adipocytes cultured in hypoxic conditions, and a negative effect of UTP was observed on UCP-1 expression in cardiac adipocytes. A beneficial effect of UCP-1 expressed in the heart of transgenic mice has already been demonstrated during ischemia–reperfusion and was associated with improved contractility and reduced oxidative stress [10].
The regulation of cardiac fat formation/function and beige markers by extracellular nucleotides is of major interest. This study defines P2Y4 as a regulator of cardiac adipogenesis and beige adipocyte generation. Our data suggest a therapeutic potential for P2Y4 antagonists to potentiate the cardioprotective function of cardiac fat and limit damage resulting from myocardial infarction.
Footnotes
Acknowledgments
We thank Hrag Esfahani (UCL, Brussels) for technical advice in ischemia experiments. This work was supported by the Fonds de la Recherche Scientifique Médicale of Belgium, Action de Recherche Concertée of the Communauté Française de Belgique, an Interuniversity Attraction Pole grant from the Politique Scientifique Fédérale (IAP-P6/30), Prime Minister's Office, Federal Service for Science, Technology and Culture, by grants of the Fonds de la Recherche Scientifique Médicale (F.R.S.M.), the Fund Lokumo, King Baudouin Foundation, the Fonds d'Encouragement à la Recherche (F.E.R.), the Fonds Emile DEFAY, the Walloon Region (Programme d'Excellence CIBLES), and the LifeSciHealth programme of the European Community (grant LSHB-2003-503337). D.C. is Senior Research Associate of the Fonds National de la Recherche Scientifique (F.N.R.S.). A.L. was supported by ULB, Belgium, and M.V., S.C., and M.H. were supported by the Fonds National de la Recherche Scientifique/FRIA, Belgium. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.