
Abstract
Direct conversion through transdifferentiation is a promising cell reprogramming approach that induces a cell lineage conversion among adult cells without passing through an intermediate pluripotent phase. However, there is a need to critically evaluate the efficacy and safety of direct conversion to establish its feasibility as a clinically viable cell reprogramming technique. This review article aims to delineate some critical constraints of direct conversion as a cellular reprogramming approach. We report the most important challenges of lineage reprogramming through direct conversion and divide them into two major sections. The first section explores the obstacles that limit the efficiency of the direct conversion process. In this study, we discuss challenges such as lack of understanding of molecular mechanism and transcriptional control of direct conversion, low proliferative capacity of converted cells, and senescence and apoptosis as critical barriers of direct conversion. The second section focuses on addressing concerns of safety of directly converted cells. We describe issues of transgene load and epigenetic memory retention along with the constraints of currently available validation tools to characterize reprogrammed cells. Each issue mentioned above is evaluated in view of their origin, implications, progress made toward their resolution and scope for development of new strategies to address the constraints of the present technique.
Introduction
T
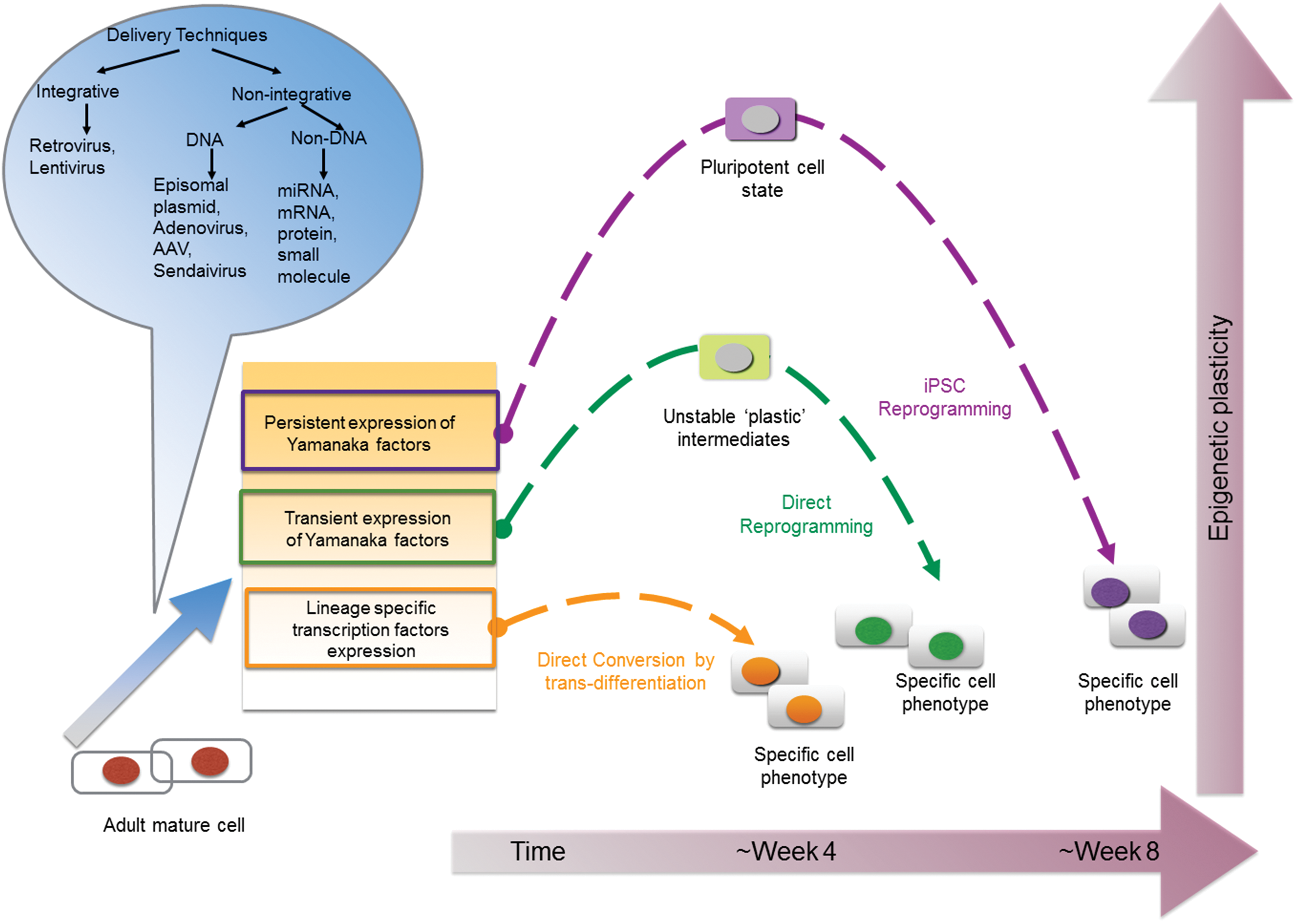
Schematic illustration of three autologous cell reprogramming techniques. An adult fully differentiated cell can be reprogrammed to a cell of target lineage by iPSC reprogramming, direct reprogramming and direct conversion through transdifferentiation. In iPSC reprogramming (purple arrows), Yamanaka factors (Oct4, Sox2, Klf4, and c-Myc) are exogenously expressed to dedifferentiate the cell to an early, pluripotent epigenetic state. The differentiation of pluripotent cells can then be directed toward the target lineage. In direct reprogramming, the Yamanaka factors are transiently expressed to induce an epigenetically “plastic”, multipotent state, following which growth factors and signaling cues direct the cell toward the target lineage. Direct conversion requires expression of lineage-specific transcription factors to induce a lineage switch (transdifferentiation) through unipotent precursors. The transcription factors required for all three techniques can be delivered through integrative and nonintegrating methods as illustrated in the blue callout. The axis on the right indicates the epigenetic state of the cells undergoing reprogramming and the axis below is an indicator of the time required to procure cells. iPSCs occupy the highest position on the epigenetic scale since the cells acquire pluripotency and have the potential to differentiate into all three germ layers. Also, it takes about 2 months to get to the target lineage using iPSC reprogramming. Direct reprogramming comes second on the epigenetic scale as it induces a plastic, but not a pluripotent state. This is followed by direct conversion, which induces least plasticity as the transdifferentiation mechanism involves a lineage switch through unipotent pathways in about 4 weeks. iPSC, induced pluripotent stem cell. Color images available online at
In the past few years, many groups, including ours, have reported that successful direct conversions in somatic cells is possible, for instance: human fibroblasts to cardiomyocytes as well as to neurons, rodent adult cells to oligodendrocytes, astrocytes and hepatocytes, human fibroblasts to Schwann cells, and pancreatic islet cells [4]. Nevertheless, the concerns regarding acquisition of effective, safe and clinically applicable reprogrammed cells from autologous cell source remain to be addressed.
Factors Limiting Efficiency of Direct Conversion
Understanding of molecular mechanisms and transcriptional regulation
As direct conversion is a relatively recent approach, scientists have yet to delineate the underlying molecular dynamics or mechanistic basis of transdifferentiation. Based on the reports of successful interlineage conversions achieved until date, one can conclude: (i) Within days after induction of transcription factors, the upregulation of target gene expression could be observed; (ii) The acquired cell identity would be stable after removal of exogenous factors; (iii) Cell division and cell proliferation would not be essential for direct conversion, and (iv) Reprogramming process would not transit through pluripotency [5]. While these are significant information to provide a basic roadmap of lineage transition, an in-depth understanding of the barriers to reprogramming could contribute to resolve the challenge of low conversion efficiencies.
A differentiated cell type only expresses tissue-specific genes in a temporal manner. In iPSC reprogramming technique, the nucleosome is displaced during cell division, thereby providing access for transcription factors to bind to the cis-regulatory regions [6]. Direct conversion is complicated by the fact that it can be achieved without cell proliferation or division. Hence, models accounting for direct conversion hypothesize that multiple transcription factors are involved to cooperatively remodel diverse areas of the genome to induce lineage conversion. To successfully achieve cell fate transition through direct conversion, the transcription factor has to find the appropriate binding site and sufficiently remodel the chromatin to facilitate activation of the target cell type's genomic program. However, epigenetically silenced regions of the genome are packed into nucleosomes, which are occluded by chromatin structures and repressors, rendering it inaccessible to transcription factors [7].
The most promising of these models, the “pioneer” transcription factor model, proposes that certain “pioneer” factors can access the relevant binding sites (which are inaccessible to the other factors). This displaces the nucleosomes, which initiates genome-wide transcriptional and epigenetic changes. These factors then recruit additional transcription factors and chromatin modifiers to execute the lineage conversion [8]. The “pioneer” transcription factor model is schematically illustrated in Fig. 2. The existence of a “master gene regulator” that drives lineage conversion was known as early as 1980 s, where Davis et al. demonstrated that MyoD is sufficient to induce fibroblasts to convert to cells of myogenic lineage [9]. However, it is only recently that a hierarchical mechanism was observed to regulate the initial stages of transdifferentiation. In this context, Wapinski et al. studied neuronal differentiation and reported that Ascl1 was found to assume a pioneering role by binding to neuronal target genes in fibroblasts and subsequently recruiting other transcription factors to facilitate neural lineage conversion [10]. It is noteworthy that Ascl1 could bind to closed chromatin and recruit secondary transcription factor (Brn2), which plays a key role in later stages of neuronal differentiation. Their results also suggest that ‘pioneer’ transcription factors help in initiating neural lineage commitment, whereas secondary transcription factors subsequently specify neuronal subtypes. Significant insight into the molecular level functions of Ascl1 was provided by Treutlein et al. who used single-cell transcriptome approach to dissect direct conversion pathways and characterize the series of intermediate states in the transdifferentiation process [11]. Their results reveal that transdifferentiating fibroblasts transit through two distinct phases, which the authors define as initiation and maturation. During initiation, the cells downregulate fibroblast-specific genes, cease proliferating, and transiently activate neural lineage-specific genes. These changes, orchestrated by Ascl1, take place in majority of cells. In contrast, only a subset of these cells progresses to the maturation phase that constitutes in stable establishment and maintenance of the acquired neural lineage. This indicates that the transition from initiation to maturation require additional factors (Brn2, Ascl1, and Myt1l). Interestingly, the results revealed that Ascl1 promotes transdifferentiation toward myogenic lineage and Brn2 and Myt1l are necessary to restrict the differentiation to neural lineage and stably acquire neural identity. This implies that although the expression of a single factor- Ascl1 is sufficient to induce direct lineage conversion, it is inefficient in restricting cells to the neuronal fate. Another study by Chanda et al. also demonstrated that overexpression of Ascl1 alone can induce transdifferentiation into neuron-like cells, but with lower efficiency than the Brn2, Ascl1 and Myt1l mixture [12]. Although there are other studies by Heinrich et al. indicating that the use of a single transcription factor, Sox2 or Neurog2, is sufficient to convert glia cells into functional neuron [13], as well as by Guo et al. who reported efficient transdifferentiation of both mouse and human astrocytes with single transcription factor, NeuroD1 [14], these results suggest that effective lineage conversion between different germ layers requires a synergy of both primary and secondary factors. Therefore, it is critical to identify the correct combination of pioneer and secondary transcription factors that can facilitate direct lineage conversion across germ layers, as well as to develop strategies that reset heterochromatic blocks, which potentially obstruct pioneer factor binding.
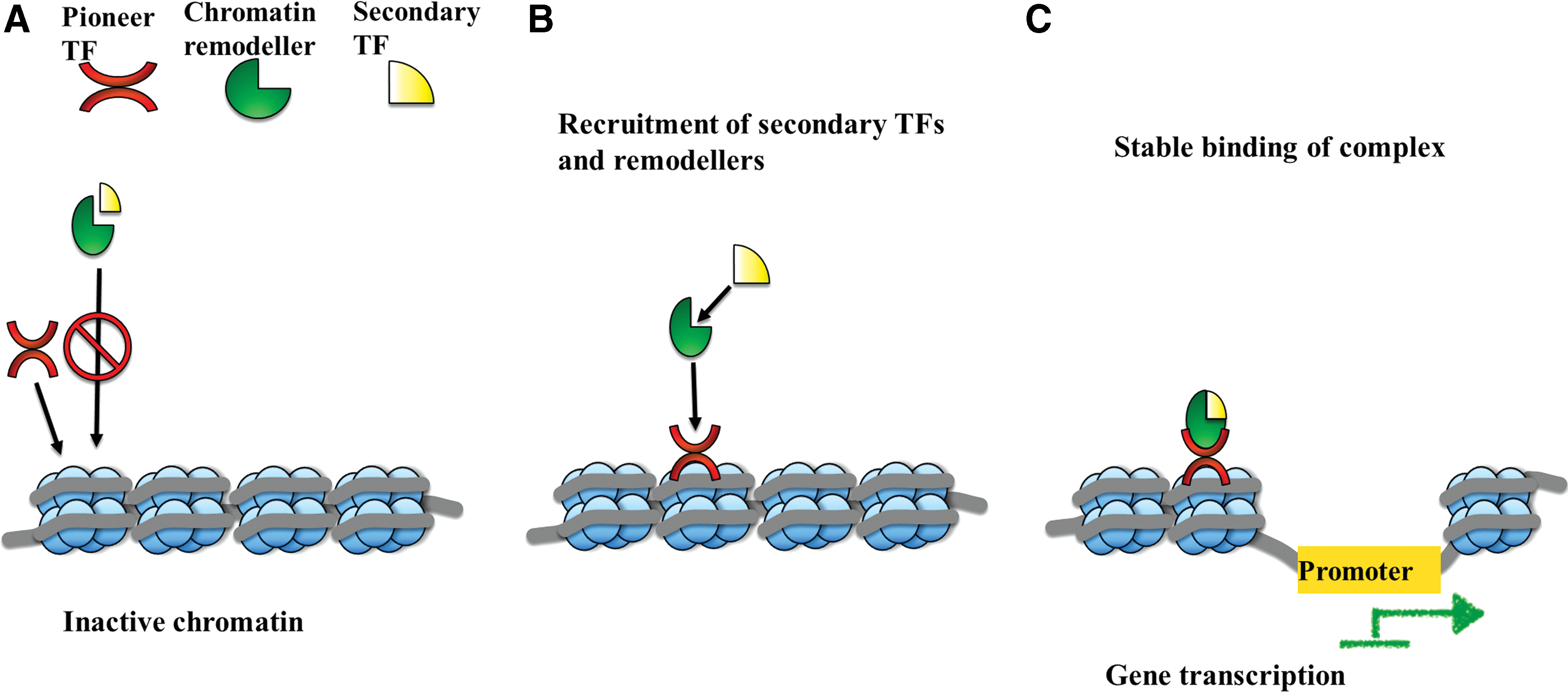
Illustration of pioneer transcription factor model of direct conversion.
Another crucial aspect of direct conversion is the identification of the optimal cell type that will efficiently respond to a specific combination of transcription factors. This poses a significant challenge as some cell types are not amenable to reprogramming, indicating that a certain degree of initial chromatin accessibility is required even for the “pioneer” transcription factors. In this regard, Gal Yam et al. have reported that cells, which have a “permissive enhancer state,” can better orchestrate promoter chromatin remodeling through binding of the pioneering regulatory factors [15]. Significant progress was made in the field of cardiac reprogramming by Zhou et al. who performed a shRNA-based loss-of-function screen to investigate the role of epigenetic factors in reprogramming to cardiac lineages. They identified polycomb ring finger oncogene Bmi1 to be acting as a critical epigenetic barrier during the early phase of reprogramming. Importantly, removing this early epigenetic barrier was found to be sufficient to generate functional cardiomyocytes with fewer transcription factors [16]. Overall, these studies indicate that two of the major strategic challenges to achieve efficient lineage conversion lie in identifying the most effective and robust transcription factors for a specific mature cell line, in addition to selecting the most permissible starting cell type.
Proliferation rate of directly converted cells
Cells undergoing transdifferentiation do not necessarily pass through an intermediate proliferative stage. This is advantageous as it avoids the possibility of erratic proliferation and acquisition of genomic aberrations. However, this may present a constraint on the quantity of cells as the amount of cells procured at the end of the reprogramming process is solely dependent on the initial number of cells used. Consequently, low proliferation rate of terminally differentiated cells presents a challenge of low scalability, which is a critical logistic parameter in a research setting.
A primed conversion approach, similar in definition to direct reprogramming, has been proposed to circumvent the issue of low proliferation. In this approach, the Yamanaka factors are transiently expressed (4 days) in the cells to achieve a plastic developmental state that functions as a springboard to propel subsequent differentiation to the target cell type [17]. The culture medium is generally conditioned to avoid induction of pluripotency by addition of STAT/JAK inhibitors or omission of leukemia growth factor. This primed conversion method proved to be particularly efficient in reprogramming cardiomyocytes. Efe et al. demonstrated that by combining transient expression of Oct4, Sox2, and Klf4 with cardiogenic developmental cues in early phases of reprogramming, a highly proliferating progenitor population was procured. These intermediates could be expanded and subsequently differentiated into mature phenotypes. Significantly, this direct reprogramming protocol was quicker, with a seven-fold increase in efficiency as compared with previous reports of direct conversion of fibroblasts to cardiomyocytes [18]. This approach involving restrained use of Yamanaka factors to destabilize somatic cells into a pre-iPSC phase before driving toward the target lineage also found success in reprogramming to neural lineages. Various reports of conversion of fibroblasts (human and rodent) to neural precursors (NPC) have emerged [19]. The common elements among these diverse protocols are transient expressions of a subset of Yamanaka factors in neuroinductive medium and coexpression of early neural transcription factors such as Brn4 and Foxg1. These conditions were found to be sufficient to propel the unstable intermediates into NPC lineage. Remarkably, direct reprogramming could be induced with the use of only one transcription factor, Oct4, in combination with lineage inductive culture conditions. Several studies have demonstrated that Oct4-mediated direct reprogramming can effectively convert fibroblasts into blood progenitor cells as well as neural progenitor cells, astrocytes into neural stem cells, peripheral blood cell into neural progenitor cells, and rat fibroblasts into oligodendrocytes [20].
Direct reprogramming through primed conversion appears to be an effective strategy to overcome the hurdle of low scalability in directly converted cells. In this approach, cells attain a short-term self-renewal capacity prior to differentiation, which aids in improving their proliferation rate and efficiency. However, recent studies have raised questions about the complete absence of pluripotency in this approach. This is due to lack of lineage tracing tools that could unambiguously validate the absence of pluripotency during direct reprogramming [21]. Further studies by Mara et al. adopted genetic lineage tracing to investigate the molecular trajectory of direct reprogramming by Yamanaka factors. Their results revealed a majority of NPCs and cardiomyocytes that were generated through the primed conversion method passes through a transient pluripotency stage [22]. Results by Mara et al. were also corroborated by Bar-Nur et al. They showed that, even with transient expression of combinations of Yamanaka factors, there is a reactivation of silent X chromosomes and presence of Oct4-positive pluripotent cells in the heterogeneous population of directly reprogrammed cells [21]. Although transient acquisition of pluripotency in this “primed conversion” approach may not affect in vitro applications of reprogrammed cells, it will be imperative to consider exclusion of residual pluripotent cells when performing in vivo studies.
Senescence as a reprogramming barrier
Since direct conversion does not involve dedifferentiation to an early developmental stage (pluripotency or multipotency), the cells undergoing lineage transition retain cellular age and are not biologically rejuvenated. This is in contrast to iPSC reprogramming, where cells acquire both potency and youth through the reprogramming process [23]. The retention of cellular age in lineage conversion through transdifferentiation may be of potential advantage in disease modeling studies as the cells manifest phenotypes of age-related diseases. For instance, Mertens et al. have recently demonstrated that in contrast to iPSC-derived neurons, directly converted neurons retain ageing-associated gene signatures [24]. However, cellular ageing or senescence has been shown to be an important barrier in achieving efficient cell reprogramming [25]. Senescence leads to a loss of replicative potential and upregulation of cell cycle-dependent kinase inhibitors. Activation of p53 pathway or culture-induced upregulation of p16Ink4a, p21CIp1, and p19Arf has been identified as the main cause of cellular senescence [26,27]. Sun et al. have recently demonstrated that manipulating the senescence process by overexpression of hTERT or inhibition of Ink4/Arf can greatly improve conversion efficiency. They successfully converted fibroblasts into neural cells within 1–2 weeks with approximately 70–80% of fibroblasts showing typical neuronal morphology and Tuj1 and Map2-positive staining within a week of culturing in neuronal induction medium. The converted neurons also demonstrated electrophysiological capability of generating action potentials and neurotransmitter receptor-mediated currents, suggesting fully functional neurons [28]. On the other hand, Yu et al. have shown that inhibiting let-7 microRNA leads to activation of a gene (HMGA2) that helps to overcome senescence-induced barriers, procuring self-renewing, rejuvenated neural stem cells from fibroblasts. HMAG2 is known to enhance self-renewal of NSCs through the repression of p16Ink4a and p19Arf [29]. These reports are significant advances made toward ablation of the molecular level barriers and bottlenecks in Direct Conversion, but caution needs to be exerted as INK4a/ARF are key regulators of chromosomal stability. Hence, it is imperative that reversible approaches are developed to transiently promote immortalization.
Another important hurdle of direct conversion, closely related to senescence is cell death during direct lineage transition. Gascon et al. used continuous single-cell live imaging to investigate direct conversion of fibroblasts and glia into neurons and identified the critical time points when the cells either get converted or apoptosed. Their results demonstrated that the peak of cell death occurred between 50–100 h postinduction, at a point, where morphological changes indicating fate conversion are evident. By exogenously expressing the antiapoptotic protein Bcl2, they were able to increase the number of surviving cells as well as the efficiency in induction of direct lineage conversion. Their study also revealed other critical reprogramming blocks such as oxidative stress and ferroptosis, which could be overcome by using ferroptosis inhibitors (Liproxstatin-1 or vitamin E) or by activation of antioxidant pathways through calcitriol [30]. Several groups have also reported increased conversion or reprogramming efficiency of differentiated somatic cells into targeted cells by inhibiting p53 activity that plays an important role in inducing apoptosis and senescence [31]. Liu et al. increased the conversion efficiency of fibroblasts to dopaminergic neurons by as high as 20-fold by blocking p53 activity using a dominant-negative p53. P53 pathway is generally activated upon the detection of stressful condition or dysfunctional telomeres, and is a critical tumor suppressor gene. One of the targets of p53 pathway is p21, a cyclin-dependent kinase inhibitor, which induces cell arrest at the G1 phase by negating the activity of cyclin-CDK2/4 complexes [32]. Jiang et al. also identified p53- mediated apoptosis pathway as a critical regulator of cell proliferation and survival in direct conversion. They could significantly increase derivation of dopaminergic neurons from human fibroblasts by blocking p53 through RNA interference. Moreover, they facilitated efficient reprogramming by arresting the cell in G1 phase by using three methods (serum withdrawal, the CDK2 inhibitor SU9516, and the mTOR inhibitor Torin1) which indicate that exiting cell cycle is a critical feature for efficient direct lineage conversion [33]. A schematic illustration of current strategies to overcome the problem of senescence and cell death in directly converted cells is provided in Fig. 3. In spite of these results, it is critical to note that senescence and apoptosis maintain genome integrity and they are cellular defense mechanisms against aberrant cell proliferation. p53 activity regulates key processes responsible for maintenance of cell health and is known to induce tumor regression in lymphomas and sarcomas [34]. Extensive manipulation of these processes could disrupt cellular homeostasis and hence, there is a need to strike a balance between attenuating senescence or apoptosis and maintaining the cell's intrinsic defense mechanisms.
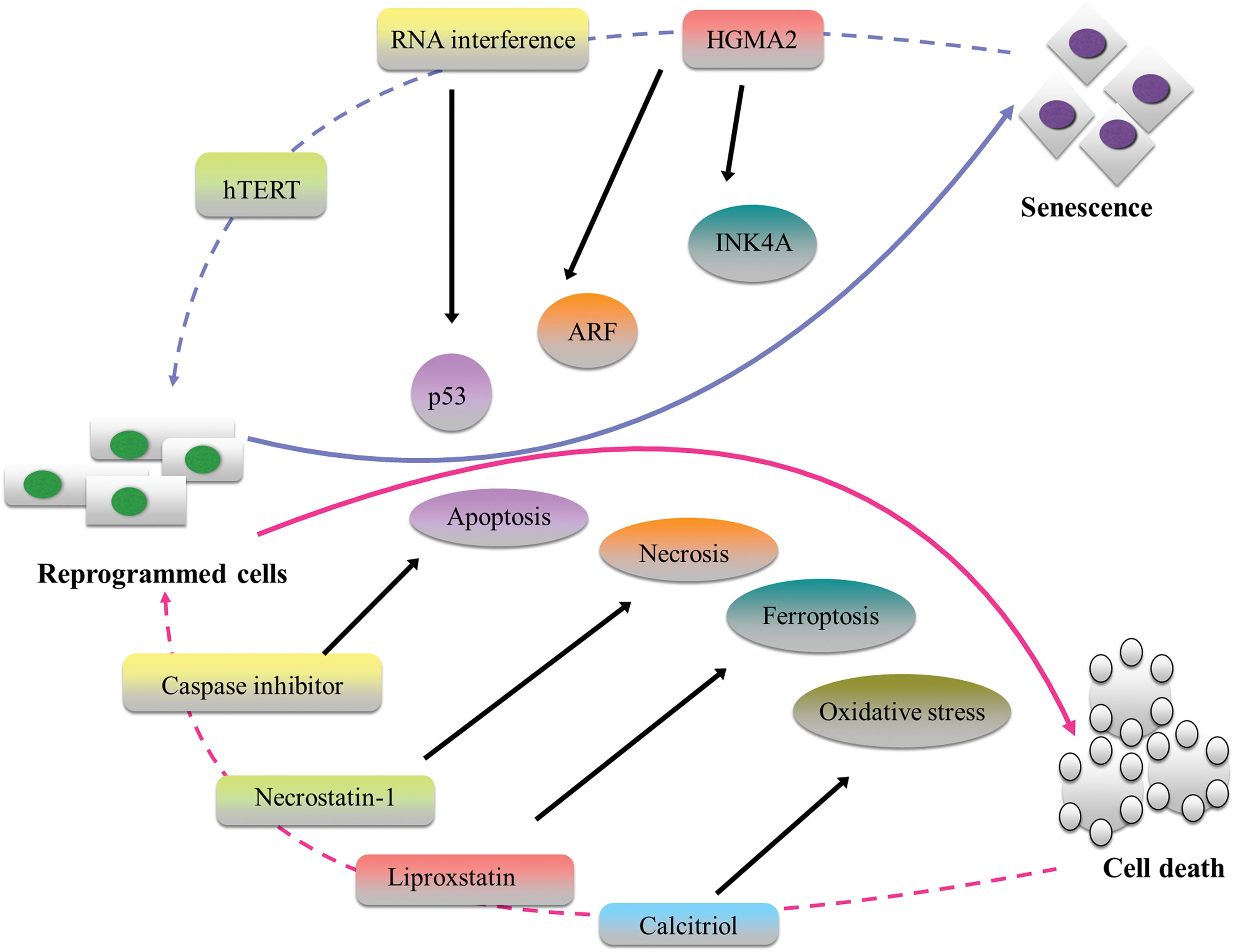
Schematic illustration of certain established strategies to overcome senescence and cell death during direct conversion. The directly converted cells have a risk of becoming senescent by activation of p53, ARF, and INK4a pathways. Highlighted are some of the effective strategies known to reverse senescence which include: overexpression of hTERT or inhibition of Ink4/Arf, activation of HMGA2, and RNA interference to block p53 activity. Cell death due to apoptosis, necrosis, ferroptosis, and oxidative stress, have also been shown to be countered by using Caspase inhibitor, Necrostatin-1, Liproxstatin, and Calcitriol, respectively. Color images available online at
Constraints on Safety of Direct Conversion
Transgene load
Transcription factor-based transdifferentiation requires ectopic expression of multiple genes to achieve the desired cell-type conversion. Retroviral vectors are a highly efficient and commonly used transgene delivery system that poses the threat of genome-wide integration, insertional mutagenesis, immunogenicity, and residual expression [35]. Various alternative routes for delivery of transcription factors have evolved to alleviate these problems. Adenoviral transduction through replication-incompetent vectors enables transient gene transfer without genome integration, although this technique compromises the transfection efficiency. Sendai virus is also an acceptable gene transfection approach as the virus replicates in the cytoplasm of the cell in the form of single-stranded RNA [36]. Adeno-associated virus (AAV) vectors have also emerged as a safe and efficient tool for exogenous gene delivery as they do not integrate into the genome and exhibit low immunogenicity. Significantly, an increasing number of phase I–III clinical trials using AAV vectors have procured promising results [37]. Episomal vectors without viral packaging or use of recombinant proteins, mRNA, miRNA is also a feasible nonintegrating transfection method. Development of novel systems that allow deletion after integration has further enhanced the feasibility of developing nonintegrating transfection protocols [38]. The piggyBac transposon uses inverted terminal repeat sequences to flank the transgene of interest. The transposase enzyme recognizes the sequence and performs a cut–paste function to move the sequence from the vector to the cell's genome. After achieving reprogramming, the transgene can be excised in a footprint-free approach [39]. Clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated (Cas) systems contribute to the intrinsic immune mechanism of archaea and bacteria to degrade foreign genomic material, and are effective at targeting eukaryotic DNA. In current applications, guide RNA (gRNA)-containing CRISPR and a complementary target sequence could be synthesized to work in a complex with Cas proteins to cleave nucleotides at targeted regions of the genome. Multiple gRNAs could also be used simultaneously to cleave the genome at multiple loci with high specificity [40]. While these nonintegrating techniques need application-specific protocol optimization to yield directly converted cells with sufficient efficiency, they are an important step toward procuring clinically relevant cell types for translational applications.
Significant advances in the field of cellular reprogramming have significantly revealed that cell fate transition cues are not limited to transcription factors. Increasingly, small molecules belonging to a class of chemical compounds with low molecular weight lower than 500 Da have also been utilized to induce somatic cell fate changes across germ layers with minimal genetic manipulation. These molecules can freely diffuse across cell membranes to modulate specific signaling pathways and gene expression. They elicit their mechanism by disruption of cell-specific programs and induction of alternative cell fate. For instance, Han et al. addressed concerns of genome integration by viral transduction in demonstrating direct conversion of mouse fibroblasts into neural stem cells through a combination of seven small molecules. They used small molecules such as VPA (histone deacetylase inhibitor), BIX01294 (G9a HMTase inhibitor), and RG108 (DNA methyltransferase inhibitor) that have been known to enhance reprogramming by modulating downstream signaling pathways (Notch, Shh, BMP, Wnt). Nevertheless, the induction efficiency was reported to be low (2%) and the cells were suspected to pass through a partial pluripotent stage [41]. In 2015, two groups provided a proof-of-principle study reporting that the direct conversion of neuronal cells could be achieved by a transgene-free, chemical-only approach without the introduction of ectopic genes [42]. Hu et al. began with a combination of three small molecule compounds (VPA, CHIR99021, and Repsox) known to induce neural progenitor state in cells. They supplemented these compounds with four more molecules (Forskolin, SP600125, GO6983, and Y-27632) that promote differentiation of neural progenitors into neurons. Inhibitors of HDACs, GSK3, and TGF-β mediate the activation of endogenous SOX2 expression and inhibitors of JNK, PKC, and ROCK facilitate neuron conversion. This final combination of chemical compounds yielded mature neurons in about 3 weeks without passing through an intermediate neural progenitor stage [42]. On the other hand, Li et al. used a more systematic approach by first identifying small molecules that enhance transcription factor based reprogramming. Then, they investigated whether exclusive use of selected small molecules (without transcription factors) could induce direct conversion. Individual compounds, such as Forskolin, ISX9, CHIR99021, and I-BET151, were identified to be capable of acting as promoters of Ascl1 (master gene regulator of neuronal differentiation). The use of these small molecules was sufficient to chemically reprogram fibroblasts to neurons with 90% Tuji-positive and 30% NeuN-positive cells [43]. Zhang et al. reported a mixture of nine small molecules (LDN193189, SB431542, TTNPB, Tzv, CHIR99021, VPA, DAPT, SAG, and Purmo) that were sufficient to directly convert human astrocytes into neurons. They provided transcriptional analysis of the reprogrammed cells to reveal that epigenetic silencing of glial genes and transcriptional activation of neural transcription factors, such as NGN2 and NEUROD1, regulate the molecular mechanism of this small-molecule-induced direct conversion. Their work is significant as the directly converted neurons could survive for more than 5 months in culture forming robust synapses and displaying synchronous burst activities. These chemically reprogrammed human neurons could also survive in vivo and integrate into the mouse's neural circuitry [44].
In the field of reprogramming toward the cardiac lineage, the importance of small molecules was demonstrated by Wang et al., who successfully generated functional cardiomyocytes from mouse fibroblast using single transcription factor (Oct 4) and a combination of small molecules: SB431542 (ALK4/5/7 inhibitor), CHIR99021 (GSK3 inhibitor), parnate (LSD1/KDM1 inhibitor, tranylcypromine) and forskolin (adenylyl cyclase activator) [45]. A year later in 2015, Fu et al., improved the technique by completely eliminating the use of Oct4 and demonstrating successful conversion with use of only pharmacological compounds. They optimized the protocol consisting of a mixture of compounds called CRFVPTZ (C, CHIR99021; R, RepSox; F, Forskolin; V, VPA; P, Parnate; T, TTNPB; and Z, DZnep) developed by Hou et al. to procure cardiomyocyte-like, spontaneously beating cells, proliferating cardiac precursors from mouse fibroblasts [46]. Significantly, Zhang et al. also derived proliferating cardiac precursors that could be propagated long term (>18 passages) using BMP4. Activin A, CHIR99021 (a GSK 3 inhibitor), and SU5402 (FGF, VEGF, and PDGF-signaling inhibitor) [47]. Since profibrotic signaling was found to attenuate successful fate conversion to cardiac lineages, Zhao et al. used small-molecule compounds to inhibit profibrotic signaling. This significantly enhanced the kinetics of cardiac reprogramming, with spontaneously contracting cardiomyocytes emerging in less than 2 weeks, as opposed to 4 weeks with transcription factor-based conversion [48].
The exact molecular mechanism of the small-molecule approach remains to be elucidated. Most combinations of small molecules contained CHIR99021 and Forskolin in the mixture, suggesting that glycogen synthase kinase-3 inhibition and cyclic AMP stimulation play a key role in lineage induction. Also, there appears to be a synergy between different small molecules which act on different aspects of the conversion process, where there is a destabilization of the starting cell's transcriptional network followed by acquisition of a new lineage identity [42]. Important mechanistic insight into the molecular functioning of small-molecule-induced reprogramming approach was provided by Zhang et al. who studied conversion of mouse fibroblasts to neural cell type. Their results indicate that chemically defined conditions induce an epigenetically activated cell state and activate lineage-specific transcription factors that act downstream of signaling pathways [49].
Although the scientific basis of this hypothesis remains to be investigated and the small-molecule approach needs to be better understood and optimized, it can be observed that small molecules play a critical role in induction of direct lineage transitions. Their optimal use in future may reduce the dependency on transcription factors, which requires risky genetic manipulation of cells.
Epigenetic memory retention
Direct conversion through transcription factors, chemicals, small molecules, or microRNA is a two-step process involving suppression of the starting cell type's gene regulatory network (GRN) and activation of the target cell type's GRN. This forced epigenetic reconfiguration is induced by intensive chromatin remodeling [35]. Epigenetic memory is defined as transcriptional remnants characteristic of the starting cell type that persist after the cellular reprogramming process. This possible retention of epigenetic memory could impact the integrity and functionality of directly converted cells [50]. For example, there have been two reports in 2008 and 2011 describing residual gene expression in macrophages transdifferentiated from fibroblasts and hepatocytes directly converted from neural cells. This raises concerns about the stability of the converted cells due to probable incomplete reprogramming [51]. Notably, both these studies did not record any significant impact of this retention of memory on acquisition of functional characteristics by the directly converted cells. In the first study, Feng et al. converted mouse fibroblasts into macrophages using transcription factors PU.1 and C/EBPα. The converted cells did not silence all of the fibroblast-specific genes, but could still display macrophage functions such as phagocytosis and inflammatory response [51]. In the second report by Marro et al., ectodermal cells (albumin-expressing hepatocytes) were directly converted into functional endodermal cells (neuronal cells). Single-cell analysis using Fluidigm dynamic real-time PCR revealed that lineage reprogramming had silenced most of the liver-specific genes, but there was residual hepatocyte gene expression observed. The authors conclude that these induced neuronal cells possess an epigenetic memory of the donor cell, but as they failed to elicit hepatic functional properties, this residual epigenetic signature may have had little functional significance [52]. This suggests that the presence of an immature cell type may not affect functionality as these residual epigenetic signals could possibly be silenced over time. Therefore, the impact of epigenetic memory on the functionality of the converted cell needs to be evaluated to enable objective assessment and validation of successful direct lineage conversion.
However, validation of epigenetic memory is a challenge as there is a lack of consensus criteria for evaluating degree of reprogramming as there are currently no universally accepted set of standards for defining a sufficiently ‘converted’ cell. To characterize directly converted cells and validate successful lineage transitions, it is necessary to outline the key characteristics that define the target cell type. General indicators include, but not limited to, gene expression, temporal protein expression of specific biomarkers, cytoskeletal organization and morphology, lineage-specific behavioral and functional attributes. Activation of reporter transgene and immunostaining for proteins are also commonly used to identify converted cells. Validation of direct conversion is typically through characterization of upregulation of target-specific genes and downregulation of starting cell type genes, but generally these studies analyze only a handful of cellular markers, and do not evaluate the genomic impact of the exogenous transcription factors [35]. In 2014, Cahan et al. developed a computational platform CellNet to address the aforementioned key issues by providing a metric of engineered cell identity through reconstruction of GRNs of various cell types and tissues [5]. CellNet is an algorithmic tool for assessing the quality of reprogrammed cells, which enables users to determine the extent to which the engineered cells have acquired the GRN that govern the target cell type. Furthermore, it identifies the regulatory nodes at which these engineered cells differ from the target cell type. It is noteworthy to mention that CellNet analysis is performed on bulk cell populations and is unable to distinguish target cell subtypes or evaluate the extent of reprogramming in a heterogeneous cell population. Thus, there still remains the need to perform single cell analysis to validate successful conversion.
In vivo direct lineage conversion
A significant breakthrough in in vivo direct reprogramming was made in 2008 when Zhou et al. introduced three factors (Pdx1, Ngn3, and Mafa) into the pancreas of adult mice and induced 20% of exocrine cells to convert into insulin-producing β-cells. However, a majority of these cells did not integrate into the host pancreas which limited their effective functionality [53]. Direct conversion bypasses acquisition of pluripotency and can be used for in vivo cell fate conversions without posing risks of tumorigenicity and immunoresponse. In the field of cardiac reprogramming, Qian et al. generated cardiomyocyte-like cells in vivo by using retroviral delivery of transcription factors (Gata4, Mef2c, and Tbx5). The induced cardiomyocytes could assemble sarcomeres, beat upon electrical stimulation, and attenuate cardiac dysfunction by decreasing infarct size up to 3 months postcoronary transplantation [54]. Although their results indicate no significant increase in reprogramming efficiency, the in vivo reprogrammed cells were phenotypically more mature and closely resembled the endogenous cardiomyocytes as compared with in vitro counterparts. The in vivo reprogramming approach has also been successful in converting resident glial cells into neuroblasts, which could eventually form differentiated neurons. Niu et al. demonstrated that a single transcription factor (Sox2) could efficiently convert endogenous brain astrocytes into proliferative neuroblasts. Upon addition of neurotrophic factors (BDNF) and small molecules (histone deacetylase inhibitor), these neuroblasts could differentiate into mature neurons and integrate into the existing neural network [55]. A critical observation made in this study was that only exogenous Sox2 expression was insufficient by itself to yield mature neurons. Therefore, a combinational strategy of using growth factors, small molecules, and signaling cues are necessary for enhanced neurogenesis in vivo. Another report by Guo et al. showed that overexpression of NeuroD1 could directly convert astrocytes into glutamatergic neurons, whereas oligodendrocyte progenitor cells (NG2 cells) were converted into glutamatergic and GABAergic neurons in vivo. Electrophysiological recordings demonstrated generation of spontaneous action potential, which could evoke synaptic responses in these converted neurons, suggesting their ability to integrate into the neural circuitry [56]. It is noteworthy that the reprogramming efficiency of single-factor NeuroD1-mediated lineage conversion was observed to be around 90%. This approach of targeting resident glial cells to replace neurons may be a viable strategy for cell replacement therapy in the central nervous system, as exogenous cell transplantation may be a highly invasive process with poor cell survivability.
Transdifferentiation within the in vivo parenchyma, where the targeted endogenous cells are capable of receiving microenvironmental cues, is certainly a very innovative idea. Interestingly, some of these in vivo conversions proved to be more easily facilitated than their in vitro counterparts. For instance, the induction of murine cardiomyocytes by transcription factors proved to be inefficient in vitro, but the same transcription factors when delivered into an injured murine heart could induce robust generation of cardiomyocyte-like cells [57]. Song et al. also showed successful induction of cardiomyocytes from endogenous cardiac fibroblasts by in vivo lineage reprogramming in an injured heart. In comparison to their in vitro counterparts, these converted cells were morphologically and functionally more similar to endogenous cardiomyocytes [58]. These results suggest that exposure to the in vivo niche provides the converting cells with the essential growth factors and the necessary mechanistic cues, thereby facilitating swift lineage conversion. However, some studies indicate the complexity in extending protocols designed for in vitro lineage conversions to in vivo reprogramming. For instance, while overexpression of Ascl1 alone is known to successfully induce neurogenesis in fibroblasts in vitro, it failed to do so in vivo, as reported by multiple studies [59]. Also, the in vivo approach could be restrictive to certain cell types and specific disease conditions. Moreover, there is a need to develop a safe and efficient method to only target cells of interest with stringent selectivity when delivering the reprogramming factors in vivo (See reviews- [60,61]).
Conclusion
The initial phase of research in direct lineage reprogramming focused on establishing the feasibility of transdifferentiation by lineage-specific factors. Today, cellular research is driven by the need to identify critical constraints of the technique and to enhance the existing methodology to scale up the generated cells for applications in regenerative medicine. In spite of the rapid progress in this field, several key challenges of direct conversion remain to be addressed, both in the context of research and clinical applications. Acquisition of the desired functional characteristics in directly converted cells is feasible, but imperfect, hence, requiring optimization of current protocols. In view of therapeutic purposes, it is essential that direct conversion is induced with least possible genetic manipulation or animal-derived cells/factors to minimize prospects of immune rejection upon cell transplantation. The demand for autologous cells also imposes a critical constraint on direct conversion, as human cells are essentially harder to reprogram. Some significant breakthroughs have been made in in vivo direct conversion technique that provides the targeted cells with the essential growth factors and the necessary mechanistic cues to facilitating swift lineage conversion in an endogenous microenvironment. Figure 4 and Table 1 provide a comprehensive assessment of direct conversion by evaluating it in view of critical parameters both from a research and clinical perspective.

Summarized evaluation of direct conversion. Direct conversion through transdifferentiation has an advantage of being an autologous cell reprogramming technique that bypasses pluripotency. The feasibility of nonintegrating, transgene-free direct conversion in a short duration and in vivo direct lineage conversion are its additional advantageous features. Current challenges of direct conversion include suboptimal efficiency, low proliferation, senescence, apoptosis, and possible retention of epigenetic memory. Color images available online at
DC, direct conversion.
The research avenue of stem cells and cellular reprogramming for applications in regenerative medicine presents a complex set of legal, moral, ethical, and conceptual issues that need to be addressed. The use of human embryonic stem cells proved to be controversial owing to the ambiguity in the ethical status of an embryo that is used for the acquisition of pluripotent cells. With the generation of iPSCs, involvement of human embryos in cell reprogramming was eliminated to a certain extent. However, the ethical questions raised about “natural potentiality” of iPSCs to develop into a human being given the correct environment still persist. Direct conversion does not induce pluripotency in the cells undergoing reprogramming, thereby, avoiding the ethical complications associated with it. However, there are other conceptual issues that still surround the use of directly converted cells for clinical applications. For instance, an important issue that arises from the use of human-derived cell source is the ownership of the cell/tissue once it is donated for research after acquiring informed consent from the donor. The legal ownership that the donor possesses over the claim to intellectual property, downstream research, financial, and nonfinancial benefits is ambiguous and controversial at present, although in vivo reprogramming can evade some of these ownership issues. There is an urgent need for collaborative effort between scientists, clinicians, and legislators to monitor and regulate research protocols and clinical studies globally. Overall, direct conversion is a rapid and safe cell reprogramming technique to generate patient-specific cells. Once the challenges such as those discussed above are overcome, it is possible that direct conversion proves to be a next-generation lineage reprogramming technique that enables generation of any cell type for cell therapy or tissue engineering-based therapeutic approaches in regenerative medicine.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.