
Abstract
Adult stem cells are undifferentiated cells with the capacity to proliferate and form mature tissue-specific cell types. Nestin is an intermediate filament protein used to identify cells with stem cell characteristics. Its expression has been observed in a population of cells in developing and adult cochleae. In vitro studies using rodent cochlear tissue have documented the potential of nestin-expressing cells to proliferate and form hair and supporting cells. In this study, nestin coupled to green fluorescent protein (GFP) transgenic mice were used to provide a more complete characterization of the spatial and temporal expression of nestin in the inner ear, from organogenesis to adulthood. During development, nestin is expressed in the spiral ganglion cell region and in multiple cell types in the organ of Corti, including nascent hair and supporting cells. In adulthood, its expression is reduced but persists in the spiral ganglion, in a cell population medial to and below the inner hair cells, and in Deiters' cells in the cochlear apex. Moreover, nestin-expressing cells can proliferate in restricted regions of the inner ear during development shown by coexpression with Ki67 and MCM2 and by 5-ethynyl-2′-deoxyuridine incorporation. Results suggest that nestin may label progenitor cells during inner ear development and may not be a stem cell marker in the mature organ of Corti; however, nestin-positive cells in the spiral ganglion exhibit some stem cell characteristics. Future studies are necessary to determine if these cells possess any latent stem cell-like qualities that may be targeted as a regenerative approach to treat neuronal forms of hearing loss.
Introduction
H
Since the first reports of avian auditory hair cell regeneration in 1988 [6,7], efforts have focused on understanding the molecular basis for hair cell regeneration to potentially inform the development of regenerative treatments for hearing loss in humans. One such effort has been the characterization of cells with stem cell characteristics in the organ of Corti. Identification of endogenous stem cells in the mammalian cochlea using cellular markers associated with multipotency has focused primarily on the molecules Lgr5 and nestin. Nestin is a type VI intermediate filament protein associated with pluripotency in embryonic and induced pluripotent stem cells, as well as multipotency in a number of adult tissues. Nestin is expressed throughout the central nervous system (CNS) during development [9]. In the adult CNS, however, its expression becomes restricted to proliferative regions of the dentate gyrus and subventricular zones [10 –15]. Following injury, upregulation of nestin is observed in the CNS [16 –18] and retina [19 –21], suggesting that nestin expression correlates with stem or progenitor cell-like qualities.
Several reports describe the expression of nestin in the murine cochlea [22 –26]. These studies provide insight into its distribution in neonatal, postnatal and adult mice; however, the dynamic expression pattern of nestin in the developing cochlea has not been systematically characterized. Differences in its spatial and temporal expression in the murine cochlea are reported, perhaps reflecting the experimental methods (nestin antibodies, nestin coupled to green fluorescent protein (GFP), βgal-nestin, or nestin-CreERT2/tdTomato-reporter mice) used to identify nestin-expressing cells. In addition, the spatial distribution and longevity of the nestin-expressing cell population in the adult cochlea remain unclear, as do their potential stem cell-like qualities [27 –30].
Understanding the dynamic expression of nestin during development and adulthood and identifying any regenerative capacity associated with its expression is of relevance to the development of novel therapies for hearing loss. Herein, we systematically examine nestin expression and its variation along the length of the cochlear duct using nestin-GFP transgenic mice. We show that nestin expression correlates with proliferating cells in limited regions of the developing inner ear. In early development, it is expressed in hair cell and supporting cell types, and is subsequently downregulated on cellular differentiation. Importantly, expression of this unique marker persists well into adulthood in discrete populations of Sox2-positive cells in gradients along the length of the organ of Corti.
Materials and Methods
Animal use
All experiments were performed with the approval of the Institutional Animal Care and Use Committee at the University of Wisconsin-Madison. Animals were handled and cared for in compliance with the Animal Welfare Act and National Institutes of Health Guidelines. Nestin-CreERT2/tdTomato-reporter mice and nestin-GFP transgenic mice were used to identify nestin-expressing cells and their progeny in developing and adult mouse cochleae. To lineage trace nestin-positive cells, nestin-CreERT2 mice and Rosa-tdTomato (Ai14) reporter mice were bred to create transgenic offspring [15,31]. CreERT2 is expressed under the control of a nestin promoter, which is inducible by means of tamoxifen (TMX). This allows it to enter the nucleus, recombine the LoxP sites, remove a stop codon, and express tdTomato in cells expressing Cre and their progeny henceforth. Nestin-GFP transgenic mice, in which enhanced GFP is expressed under the control of regulatory regions of the nestin gene [32], were used to evaluate the spatial and temporal expression of nestin in the developing and adult murine inner ear. Nestin-GFP transgenic mice were provided by Dr. Xinyu Zhao (University of Wisconsin, Madison, WI).
Genotyping
C57BL/6J transgenic mice were genotyped by polymerase chain reaction using genomic tail DNA. The following genotyping primers were used: GFP forward: 5′TGACCCTGAAGTTCATCTGCACCA3′ and GFP reverse: 5′GTGGCGGATCTTGAAGTTCACCT3′; Cre forward: 5′ATTTGCCTGCATTACCGGTC3′ and Cre reverse: 5′ATCAACGTTTTCTTTTCG3′; Trans forward: 5′GGCATTAAAGCAGCGTATCC3′ and Trans reverse: 5′CTGTTCCTGTACGGCATGG3′; Wild Type forward: 5′ACTTGCTCTCCCAAAGTCGCTCTGA3′ and Wild Type reverse: 5′TACTCCGAGGCGGATCACAAGC3′.
Timed pregnancies
To generate timed pregnancies, breeding pairs were placed together overnight. Pups of pregnant females were considered embryonic day (E) 0.5 the morning a vaginal plug was observed. Plugged females were housed separately and entered into the laboratory calendar for scheduled tamoxifen administration, 5-ethynyl-2′-deoxyuridine (EdU) administration, and/or tissue harvest. The gestation period for nestin-GFP C57BL/6J transgenic mice is ∼21.5 days, where E21.5 is also defined as P0 (the date of birth).
Proliferation analysis
EdU (A10044; Invitrogen) was administered to pregnant dams (200 mg/kg body weight) by intraperitoneal injection (ip) at E11.5, E12.5, E13.5, E14.5, E15.5, or E17.5. EdU incorporation was detected in embryonic pups using Click-iT Alexa Fluor 488 or 555 Imaging Kit (C10337 and C10338; Invitrogen).
Tamoxifen administration
The promoter of pregnant dams was activated with tamoxifen (250 mg/kg/ip body weight, 30 mg/mL in 10% EtOH mixed with sunflower oil) [15,26,33] at E11.5, E13.5, E14.5, E15.5, E16.5, or E19.5. A single dose of tamoxifen was administered to each pregnant dam. Nursing mothers were injected with tamoxifen (250 mg/kg/ip body weight) on P1 and P2 to activate Cre in nursing pups. For all samples, cochlear tissue was harvested within 1 week after tamoxifen injection. Previous analysis confirmed appropriate morphological appearance and expression of tdTomato in nestin-CreERT2/tdTomato-reporter mice, with no tdTomato observed within the cochlea in the absence of tamoxifen [26].
Auditory evoked response measures
To confirm auditory function in transgenic mice, evoked response measurements were performed on mice greater than 25 days of age using the Tucker-Davis Technologies RZ6 system. Mice were anesthetized using ketamine (100 mg/kg/ip) and xylazine (10 mg/kg/ip) and positioned in an audiometric sound attenuation chamber (Lafayette Instrument) on a warming pad. Three subcutaneous needle electrodes were used to measure evoked responses: the positive one at the vertex, the negative one at the mastoid, and the ground on the opposite mastoid. Click stimuli were presented using alternating polarity (rate: 21/s; period: 47.619 ms). Electroencephalographic activity for each stimulus was recorded for 10 ms. Auditory evoked response waveforms were recorded in 10 dB sound pressure level intervals down from the maximum amplitude of 90 dB. The threshold was defined as the lowest stimulus level at which response peaks for waves I–V were consistently present on visual inspection.
Tissue preparation
Observations of the distribution of nestin-GFP in each age category were made on three or more preparations, each from different animals. Embryonic pups were isolated from anesthetized dams under isoflurane at scheduled time points. Heads from embryonic pups at E11.5–E12.5 were removed over phosphate buffered saline (PBS) and immediately fixed in 4% paraformaldehyde, pH 7.4 (Electron Microscopy Services), at 4°C overnight. Cochleae from E13.5 and older pups were isolated from the head in PBS followed by fixation in 4% paraformaldehyde at 4°C overnight.
Postnatal mice were euthanized with carbon dioxide. Cochlear tissue was isolated from the head and fixed in 4% paraformaldehyde at 4°C overnight. Because neonatal mice are resistant to the effects of carbon dioxide euthanasia, heads of mouse pups up to 10 days of age were removed and their inner ear tissue harvested as described above. For P9 and older pups, decalcification of the bone was performed with a 4% solution of ethylenediaminetetraacetic acid (EDTA) for 1.5 days at 4°C following fixation. Samples were prepared as whole mounts or as cryosections.
Adult mice were anesthetized with isoflurane and transcardially perfused with saline, followed by 4% paraformaldehyde. Temporal bones containing the inner ear were removed and fixed in 4% paraformaldehyde overnight, after which they were moved to a 4% solution of EDTA for 3 days to decalcify the bone. Whole mount microdissections of the organ of Corti from the temporal bone were performed and processed for immunohistochemistry.
Cryosectioning
Samples were cryoprotected by incubating them successively overnight in 15% and 30% sucrose, after which they were incubated in a 50/50 solution containing Tissue-Tek O.C.T compound (23–730-571; Fisher) and 30% sucrose for 24 h. Samples were embedded in Tissue-Tek O.C.T compound and frozen for sectioning. Sections were cut at 20 μm thicknesses using a μicrom HM 505N cryostat or Leica CM 1950 cryostat. Following collection, sections were air-dried for 1 h and stored at −80°C. Heads from E11.5 and E12.5 pups were sectioned horizontally. For E13.5 and older pups, the inner ear was isolated from the head and oriented to produce midmodiolar sections through the cochlea.
Immunolabeling
Cochlear tissue sections were incubated at 4°C for 30 min in a solution containing 0.1% saponin (47036; Sigma-Aldrich) and 0.1% Tween 20 (P7949; Sigma-Aldrich) in PBS, followed by incubation for 1 h in a blocking solution consisting of 10% normal donkey or goat serum and 0.2% Triton X-100 in 1 × TBS (adapted from Hume et al. [34]). Tissue sections were then exposed to primary antibodies diluted in the blocking solution overnight. Primary antibodies used in this study include myosin VIIa (cat# 25-6790, 1:200; Proteus Biosciences), Sox2 (cat# sc-17320, 1:200; Santa Cruz), Ki67 (cat# PA1-38032, 1:500; Thermo Scientific), MCM2 (cat# 3619S, 1:500; Cell Signaling Technology), nestin (cat# AB5922, 1:200; Millipore), and GFP (cat# sc-101525, 1:200; Santa Cruz). For mouse monoclonal GFP antibody staining, tissue sections were incubated for 30 min in 3% mouse blocking reagent (cat# 715 007 003; Jackson) before the application of primary antibody to prevent crossover staining. Following overnight incubation to primary antibodies, samples were washed three times in PBS for 5 min and then exposed to fluorescent-labeled secondary antibodies and DAPI (1:1,000; Sigma-Aldrich) in the blocking solution overnight. Appropriate secondary antibodies were Alexa conjugated (Invitrogen or Jackson ImmunoResearch) and used at a 1:500 dilution. Phalloidin rhodamine (A22283; 1:1,000; Sigma-Aldrich) was used to label F-actin. The following day, samples were washed three times in PBS for 5 min and mounted with the Southern Biotech Fluoromount-G mounting medium (OB100-01; Fisher Scientific) for analysis.
Antigen retrieval was required to permit use of the nestin antibody, which disrupted the transgenic fluorescence. Antigen retrieval was performed by heating the samples to 90°C in citrate buffer (pH 6.0) for 20 min, after which the samples were allowed to cool to room temperature. Additional permeation in 1% Triton X-100 in 1× TBS was used for 1 h before incubation in the blocking solution. Because of the destructive nature of the antigen retrieval process, nestin-GFP fluorescence was no longer visible requiring the use of an antibody against GFP along with appropriate fluorescein isothiocyanate-conjugated secondary antibodies to localize nestin.
Tissue samples were imaged with a Nikon A1R-A1 confocal microscope using 10 × , 20 × , and 60 × objectives. Approximately 60 to 100 stacks were acquired per image ranging from 0.75 to 1.25 μm per Z-step. Sections were analyzed with Nikon Instruments Elements Software and exported into tiff-format. Images were adjusted for brightness, contrast, and sharpness using FIJI ImageJ software [35] and figure panels generated with Adobe Illustrator™ software.
Results
Cochlear expression of nestin-GFP during organogenesis and the early postnatal period
At embryonic days (E) 11.5 and 12.5, GFP was not detected in the auditory regions of the otic vesicle; however, as expected, GFP was observed in the neighboring neural tube. In the developing murine cochlea, GFP was first observed in the spiral ganglion region at E13.5 (Fig. 1A). By E14.5, GFP can be detected in the basal region of the developing sensory epithelium (Fig. 1B, C), at which time this cell population undergoes completion of terminal mitosis and begins terminal differentiation [36]. There was a base to apex gradient of GFP expression as development progressed, with GFP detected in all three turns of the cochlea at E15.5 (Supplementary Fig. S1; Supplementary Data are available online at
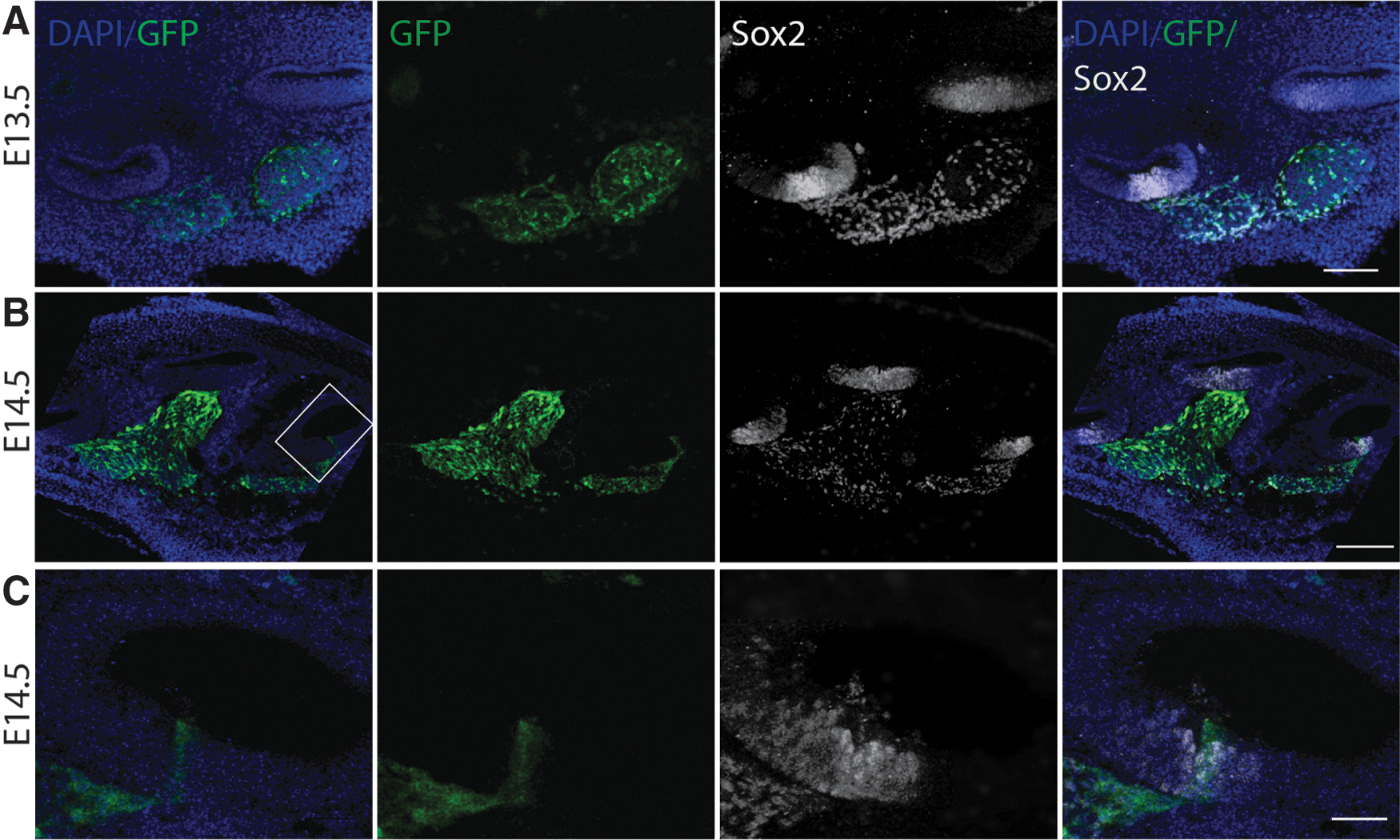
Cryostat section through the developing cochlea of an E13.5 and E14.5 embryo.
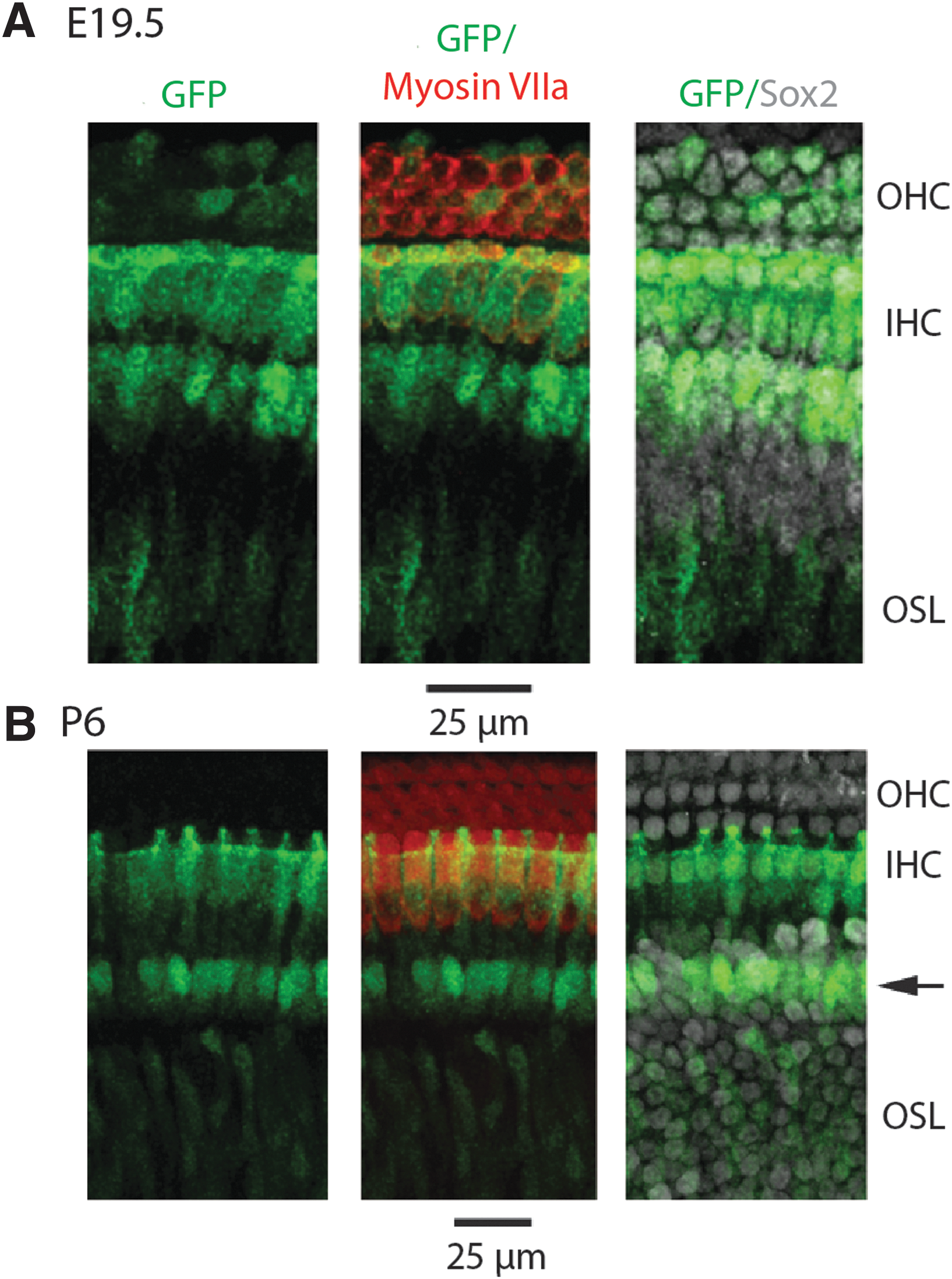
Using myosin VIIa as a marker for hair cells and Sox2 as a marker of supporting cells, we noted that all GFP-expressing cells expressed Sox2 beginning at E13.5 in the spiral ganglion region. By E15.5, GFP was detected in some Sox2-expressing cells in a basal to apical gradient in the prosensory region that gives rise to the organ of Corti [37,38]. It is at this time that myosin VIIa first appears in the basal region of the cochlea, concurrent with GFP expression in putative hair cell and supporting cell types (Supplementary Fig. S1). GFP expression progressed apically consistent with terminal differentiation. By E18.5, cell types of the inner ear are morphologically distinct and GFP could be detected throughout the sensory epithelia. Strong GFP expression was detected in the inner hair cells throughout the organ of Corti, with weaker expression in the outer hair cells. GFP was also observed in supporting cells, the osseous spiral lamina and in the spiral ganglion. By E19.5, there was widespread GFP expression throughout the cochlea (Fig. 2A), with differential expression overwhelmingly apparent from the apical to basal end of the cochlea in whole mount preparations at this developmental time (Fig. 2B). Inner hair cells begin to downregulate GFP basally, but in the middle and apical regions of the organ of Corti, nestin expression persists. In outer hair cells, there was variable GFP expression with decreased expression at the base of the cochlea. GFP was also observed in the osseous spiral lamina and was expressed in Sox2-expressing supporting cells.
Differential expression of nestin-GFP in the cochlea of an E19.5 embryo.
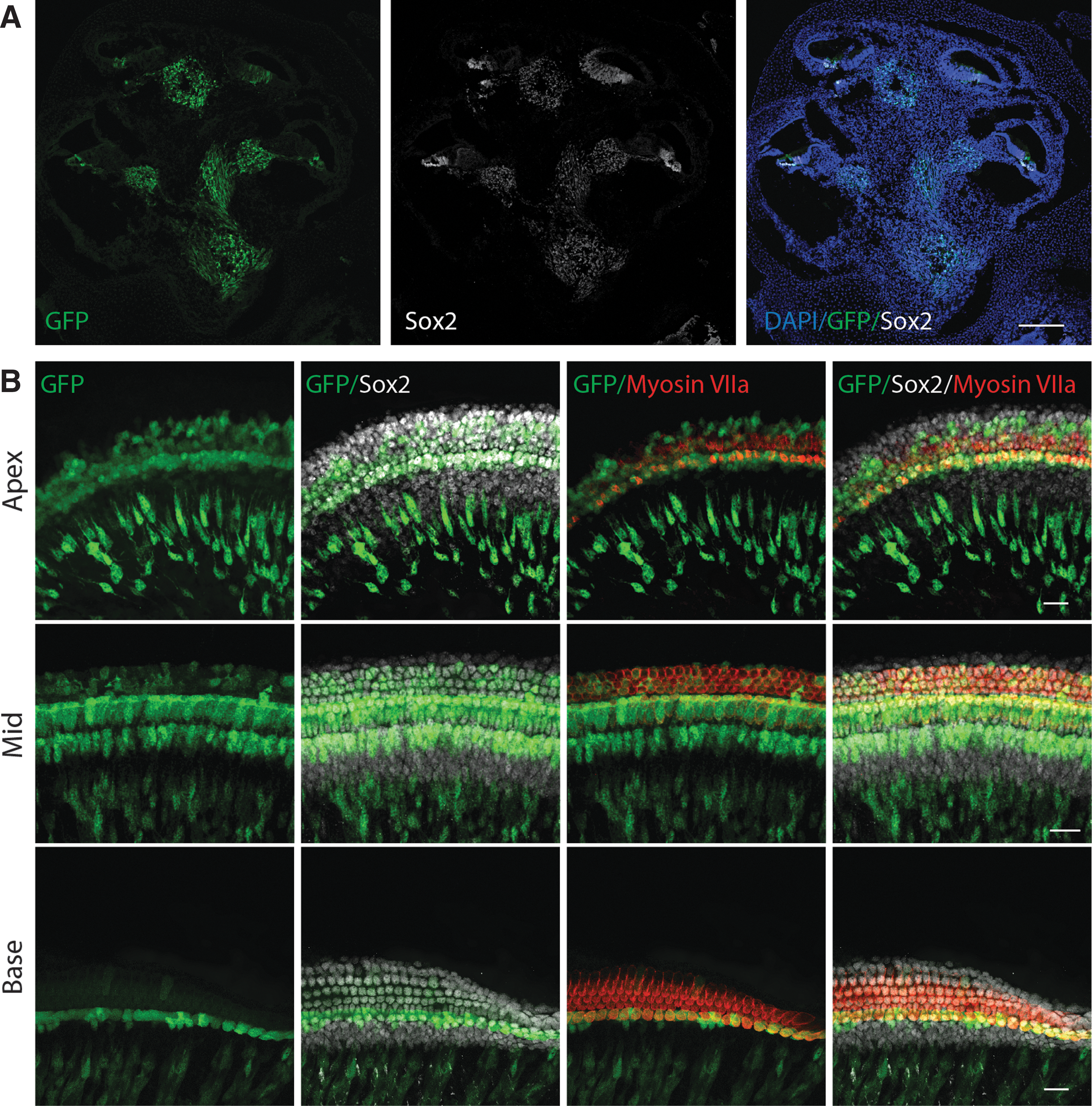
At postnatal day (P) 2, a differential gradient of GFP continues to be observed from base to apex (Fig. 3A). In the basal region of the cochlea, GFP is nearly undetectable in the inner and outer hair cells. In contrast, strong GFP was observed surrounding the medial facet of the inner hair cells in presumptive inner border and inner phalangeal cells. GFP expression continued to persist in cells throughout the osseous spiral lamina in all sections. In the midsection of the cochlea, there was still a hint of GFP in some inner and outer hair cells (Fig. 3B). As in the base, GFP was expressed in presumptive inner border and inner phalangeal cells and continued to be detected in the osseous spiral lamina. In the apical region of the cochlea, strong expression of GFP remained in the inner hair cells. In addition, GFP was present in Sox2-positive supporting cells and in the osseous spiral lamina.
Differential expression of nestin-GFP in whole mount preparations of the cochlea of a P2 pup.
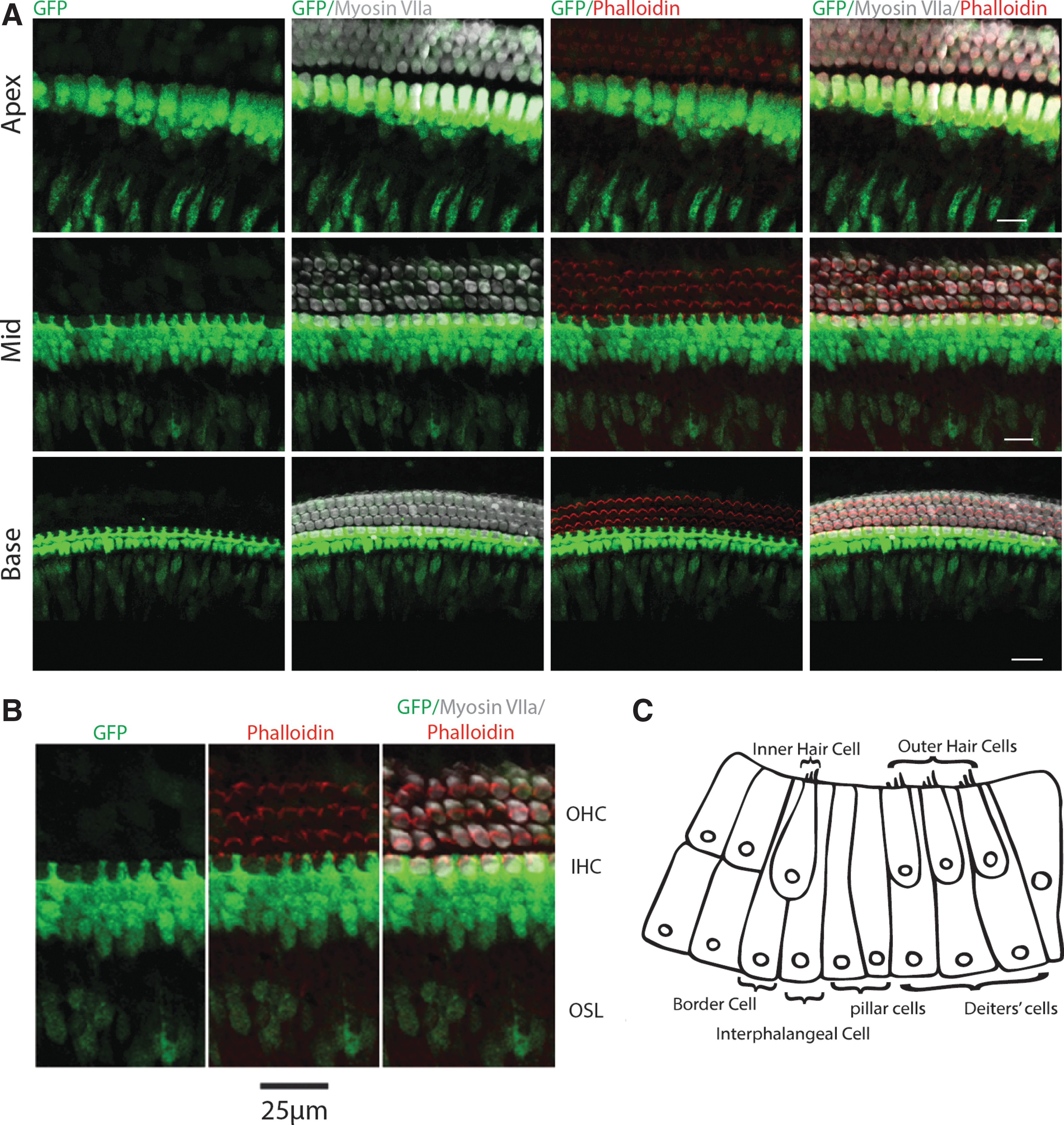
At P6, GFP persisted in presumptive border and inner phalangeal cells and within the osseous spiral lamina, but it was undetectable in inner and outer hair cells throughout the cochlea. Fewer GFP-positive cells were observed in the osseous spiral lamina in the basal end compared to the apical end of the cochlea. Of interest, a new expression pattern of GFP was detected in Deiters' cells in the basal region. This expression pattern slowly progresses in an apical manner, which reached the midregion around P12 and apex by P20. At P8, weaker GFP expression was observed in cells surrounding the medial facet of the inner hair cells (presumptive border and inner phalangeal cells) in the basal region of the cochlea compared to stronger expression observed in the middle and apical regions. In these cell types, GFP was subsequently downregulated starting at the base and progressing toward the apex over the course of 22 days, with no GFP observed in this cell type by P30.
In summary, the pattern of GFP expression alters dramatically during early cochlear development, with the most rapid changes occurring up to P6 (Fig. 4). The basal to apical gradient of GFP expression proceeded in a manner both chronologically and spatially similar to the gradient of terminal differentiation in the mammalian cochlea.
Whole mount high magnification of the expression of nestin-GFP in developing mouse cochleae.
Cochlear expression of nestin-GFP in the mature cochlea
By P30, the expression pattern of GFP remains fairly stable, with GFP observed in Deiters' cells and in a cell population just under and medial to the inner hair cells, in a similar cell population where we identified nestin-tdTomato in a previous study [26]. We were unable to determine if this cell population represents supporting cells at the medial aspect of the organ of Corti or peripherally located cells of the spiral ganglion region, hereafter referred to as the osseous spiral lamina [26]. At 6 weeks of age, GFP was observed in only these two regions. This pattern of expression persisted throughout the length of the organ of Corti, with increasing expression in Deiters' cells apically. By 9 weeks of age, this gradient of expression was more pronounced, with little GFP detected in Deiters' cells in the base and weak expression in the midregion of the cochlea (Fig. 5A). Quantitative analysis confirms that GFP expression exponentially declines with distance from the apex in the Dieters' cell population at P66 (Fig. 5B). Downregulation of GFP in Deiters' cells continued to gradually progress toward the apex throughout adulthood, with little GFP detected in the midregion by 19 weeks of age (similar to the base at 9 weeks). This expression pattern persisted at the last time point evaluated at 1 year of age (Fig. 6A). At this time, GFP expression persisted in a population of Deiters' cells at the apex (Fig. 6A, B). In contrast to the gradient changes observed in Deiters' cells, persistent expression of GFP just under and medial to the inner hair cell region was observed through 1 year of age, with more robust GFP observed in the base (Fig. 6A, C). Of importance, this cell population appears in the same region and cell type, which we observed tdTomato using nestin-CreERT2/tdTomato in our previous study [26].

Nestin-GFP expression in the P66 adult cochlea.
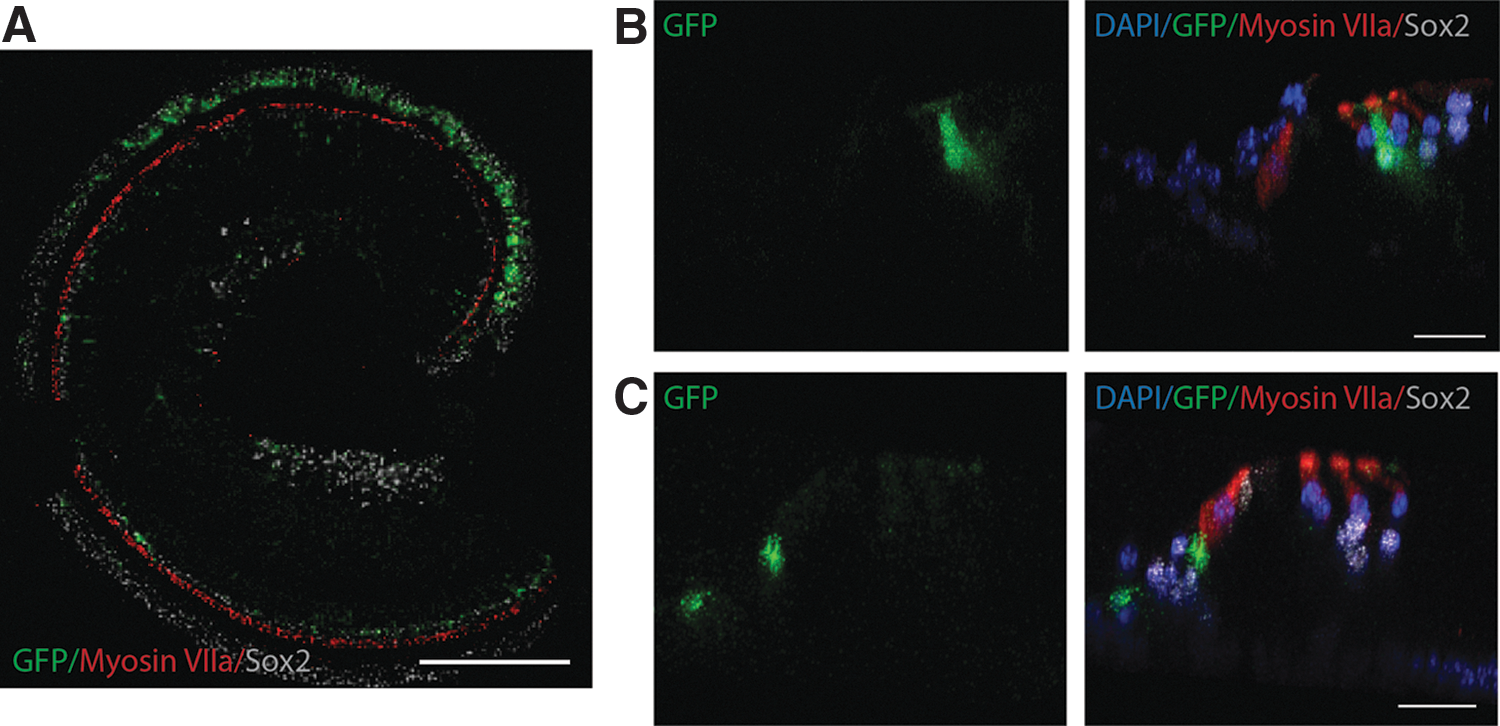
Nestin-GFP expression in the cochlea of a 1-year-old mouse.
GFP expression remained relatively consistent in the mature organ of Corti. Two differing expression gradients were observed, with decreasing GFP expression in the Deiters' cell population with distance from the apex and increasing GFP expression with distance from the apex in the cell population medial to and below the inner hair cells in the osseous spiral lamina. GFP expression was gradually downregulated over time in Deiters' cells in a basal to apical manner with continued expression in the apex at 1 year of age.
Vestibular expression of nestin-GFP
At E12.5, GFP was first detected in the vestibular regions of the otic vesicle. By E15.5, GFP was observed in myosin VIIa-expressing hair cells and Sox2-expressing supporting cells in the saccule, utricle, and cristae (data not shown). In addition, GFP was detected in the vestibular ganglia. This expression pattern persisted on E19.5 when cell types are more distinct, with GFP detected throughout the vestibular epithelia (Fig. 7A–C). At P0, weak GFP was detected in hair cells in the crista; however, strong expression persisted in supporting cells (Fig. 7D). Likewise, GFP remained in some hair and supporting cell types in the utricle and saccule (Fig. 7D–F). This expression pattern was no longer detected in the cristae at P12, when GFP was only observed in supporting cells (Fig. 7I). In contrast, a small fraction of hair cells and supporting cells coexpressed GFP in the utricle and saccule (Fig. 7G, H).

Nestin-GFP expression in the vestibular epithelia. GFP is expressed in hair cells and supporting cells in the E19.5 saccule
At 1 year of age, GFP was still identified in the vestibular sensory epithelia in the utricle, saccule, and cristae (Fig. 7J–L). In the utricle and saccule, some GFP-positive cells coexpress Sox2, while others do not. In contrast, within the sensory epithelium of the cristae (Fig. 7L), all GFP-positive cells coexpressed Sox2. No expression of GFP with the hair cell marker myosin VIIa was observed in any region of the vestibular sensory epithelia at this time.
Proliferative analysis of nestin-expressing cells
We next sought to evaluate if nestin-expressing cells possessed any proliferate capacity in the inner ear in vivo. Nestin-GFP and nestin-CreERT2/tdTomato-reporter mice were used for this analysis. Tissue was evaluated for immunoreactivity for the proliferation markers Ki67 and MCM2 [39 –42] and/or incorporation of EdU. As expected, Ki67-positive cells were observed in the developing otocyst at E11.5 and E12.5 during known proliferative times [36] (data not shown). At E14.5, Ki67 was observed to coexpress with a subset of GFP-positive cells in the osseous spiral lamina and spiral ganglion region of the cochlea. This expression pattern persisted for all samples tested through P12 (Fig. 8A). In agreement with Ki67 staining, some GFP-positive cells in the osseous spiral lamina and spiral ganglion region coexpressed with the proliferation marker MCM2 at E18.5 and P14 (Fig. 8B, C). In general, it appears that a subset of nestin-positive cells may possess some proliferative capacity in the inner ear based on coexpression of Ki67 and MCM2 with GFP; however, this is limited to the osseous spiral lamina and ganglion cell region.

Expression of nestin-GFP with proliferation markers Ki67 and MCM2.
Additional analysis using the mitotic tracer EdU was carried out to corroborate our immunostaining results. Nestin-CreERT2/tdTomato-reporter mice were used for this analysis. In nestin-CreERT2/tdTomato-reporter pups, tamoxifen was administered between E11.5 and E16.5 to lineage trace nestin-expressing cells. EdU was administered 1 day after tamoxifen exposure. Analysis of pups revealed that recombination was restricted to cells in the developing spiral ganglion region. Of these cells, a subset of tdTomato-positive cells was positive for EdU for all samples tested (Fig. 9A–D).
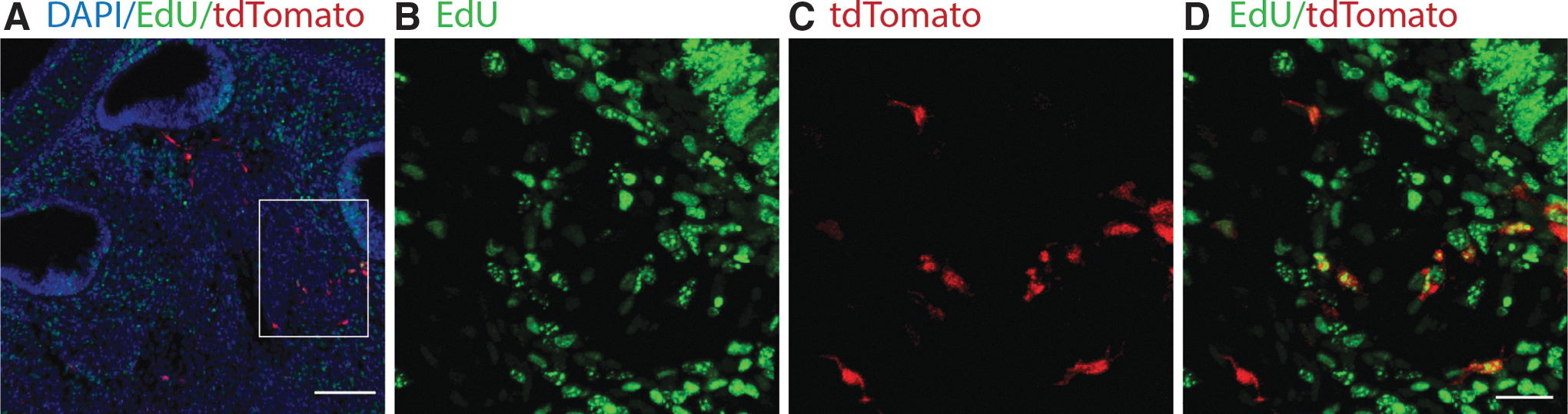
Proliferative analysis in nestin-CreERT2/tdTomato-reporter mice.
Validation of nestin-expressing cells
The expression of nestin in the developing and mature murine inner ear was explored principally using nestin-GFP transgenic mice. Immunostaining was used to verify the fidelity of GFP for nestin. Antibodies to nestin antigens confirmed that GFP-positive cells were nestin positive; however, the antigen retrieval technique to permit use of the nestin antibody resulted in poor tissue preservation (Supplementary Fig. S2A). Because of this, triple transgenic nestin-CreERT2/tdTomato/nestin-GFP mice were generated to verify faithful expression of nestin between the transgenic lines (Supplementary Fig. S2B, C). All tdTomato-positive cells coexpressed GFP; however, additional labeling of GFP was observed in tdTomato-negative cells. This could be explained by the efficiency of the reporter line in nestin-CreERT2/tdTomato double transgenic mice. A recent report found that the recombination efficiency of the nestin-CreERT2 mice used in this study was lower than expected; however, the specificity was more accurate [43]. This would explain the limited labeling of tdTomato relative to GFP. Collectively, these findings suggest that the GFP- and tdTomato-expressing cells are nestin positive; however, tdTomato only labels a limited number of GFP-expressing cells.
Discussion
Nestin was originally discovered as an intermediate filament protein that labels stem cells, but not mature differentiated cell types in the CNS [9]. Since then, its expression has been documented in a number of other organ systems [22 –24,44 –47]. In the present study, we have shown that GFP is expressed in a basal to apical gradient in multiple cell types during inner ear development and into adulthood, and that nestin-positive cells can proliferate in restricted regions of the developing inner ear. We found consistent GFP expression in the cochlea starting at E13.5. GFP was consistently observed in the auditory sensory epithelium starting in the base at E14.5, with continual changes occurring throughout adulthood. Most of these changes progressed in a basal to apical manner, consistent with what is known of the time course of terminal differentiation in the cochlea [36]. Our characterization was sufficiently detailed to demonstrate that gradients of GFP expression occurred in different regions of the cochlea at specific developmental time points. This may account for the disparity between previous reports of nestin expression in the developing rodent cochlea [22 –24,27,28,30,48].
In the current study, we observed nestin expression in similar cell types in the developing cochlea as Lopez et al. and Smeti et al. [22,23]; however, we also observed rapid basal to apical changes during embryonic and early postnatal time points consistent with terminal differentiation in the organ of Corti. In the mature cochlea, we observed GFP in a cell population medial to and below the inner hair cells in a similar region reported by Smeti et al. [23]. Moreover, in agreement with observations by Lopez et al. and Watanabe et al. [22,24], nestin-GFP was detected in the apical region of the cochlea in what appears to be a subpopulation of Deiters' cells. Deiters' cell labeling for GFP exhibited an apical–basal gradient with stronger labeling at the apex. Since this gradient was observed at P66 and at 1 year of age, it is unlikely to reflect a developmental difference.
Compared to the developing auditory epithelium, little has been reported on the expression of nestin in the adjacent vestibular sensory epithelium. Only one report by Lopez et al. describes nestin expression in the crista ampullaris and utricle in postnatal and adult GFP transgenic mice [22]. In the current study, we evaluated nestin expression at embryonic, postnatal, and adult time points. At E15.5, GFP was detected in both hair and supporting cells in the saccule, utricle, and cristae in addition to the vestibular ganglia. This expression pattern persisted until P0. In contrast to Lopez et al., we detected a weak GFP signal in myosin VIIa-positive hair cells in the crista, with strong expression of GFP in Sox2-positive supporting cells at this time. At P12, a small fraction of hair cells and supporting cells colabeled with GFP in the utricle and saccule. Conversely, GFP was only detected in Sox2-positive supporting cells in the crista from P12 through 1 year of age. In agreement with Lopez et al., we observed a decrease in GFP in the vestibular sensory epithelia as maturation proceeded.
A number of rat studies have concluded that nestin may be involved in proliferation of the inner ear sensory epithelia based on in vitro analysis [27,29,30]; however, this has not been evaluated in vivo. Proliferation of the auditory sensory epithelium does not occur spontaneously under normal circumstances following completion of terminal mitosis, so in vivo analysis for proliferation had to be carried out during development at known proliferative times. Immunostaining for proliferation markers Ki67 and MCM2 was evaluated at embryonic and postnatal time points. Widespread Ki67 was observed in the otic vesicle on E12.5 and E13.5. In the developing organ of Corti, GFP-positive cells never coexpressed with Ki67 nor MCM2 at any time point evaluated. This was not unexpected, given GFP was first observed in the developing sensory epithelium after completion of terminal mitosis. We observed GFP coexpression with Ki67 in the vestibular ganglion region as early as E13.5 and in the spiral ganglion at E14.5. Some GFP-positive cells colabeled with Ki67 and MCM2 in the osseous spiral lamina at embryonic and postnatal times, with fewer double labeled cells in the mature inner ear. Taken together, immunostaining for Ki67 and MCM2 suggests that nestin-expressing cells may possess some capacity to proliferate in the spiral and vestibular ganglion regions of the inner ear, but not the auditory sensory epithelia.
It has been reported that after ablation of cochlear hair cells in neonatal mice, some hair cell regeneration occurs, principally at the apex [49]. This may suggest that there is something special or different about cells at the apical end of the cochlea, which may possess a greater capacity for regeneration. A more recent study using nestin β-gal mice found that the area of nestin-expressing cells increased via nonmitotic mechanisms in the apical region of the adult cochlea 2 weeks following noise trauma [24]. Continued analysis at additional times following noise exposure would be helpful in determining the extent of upregulation relative to time after damage. Moreover, additional investigation of other forms of auditory insult, such as ototoxicity, on nestin-expression using the mouse model used in this study may provide insight on any regenerative or inflammatory role that nestin may have in other otological disease processes.
Additional experiments using nestin-CreERT2/tdTomato reporter mice as a means to lineage trace nestin-expressing cells during development were met with unforeseen challenges. Analysis of pups whose promoters were activated with tamoxifen during embryonic times did not result in sufficient recombination. We found recombination was restricted to cells in the auditory and vestibular ganglion regions of the inner ear, with fewer labeled cells the earlier tamoxifen was administered. In general, there was better recombination in neural regions compared to sensory regions. Consistent with other studies [50 –52], reporter expression in nestin-Cre mice was difficult to detect before E15, in which recombination and subsequent expression are observed in a very limited number of cells. This might occur in developmental studies due to insufficient induction of the transgene necessary to excise Cre, which may be further complicated by a lag in the accumulation of the fluorescent reporter [52,53]. Moreover, the specificity and efficiency of reporter expression have been found to vary among nestin-CreERT2 lines [43], with the nestin-CreERT2 line we used having been noted to be more specific but less efficient. Because of this, nestin-CreERT2/tdTomato-reporter mice were only used for proliferative analysis. Insufficient recombination efficiency did not allow us to lineage trace these cells during development as desired, but in cells where sufficient recombination occurred, we could assess for proliferation via EdU incorporation.
Analysis using EdU was carried out to corroborate the proliferative staining of Ki67 and MCM2. Nestin-CreERT2/tdTomato-reporter mice were used to lineage trace nestin-expressing cells. Tamoxifen was administered to label nestin-positive cells with tdTomato, after which EdU was administered to label cells undergoing active proliferation. As expected, EdU was not detected in the organ of Corti when administered after E14.5, at which time terminal mitosis is complete [36]. For all samples evaluated, a majority of tdTomato-expressing cells coexpressed EdU, supporting the conclusion that nestin-expressing cells indeed proliferate in the spiral ganglion region.
In our previous study, nestin-CreERT2/tdTomato-reporter mice were used to lineage trace nestin-expressing cells in the adult murine inner ear [26]. TdTomato localized to a cell population medial to and below the inner hair cell layer in a similar region we observed GFP in this study. However, using nestin-GFP transgenic mice, we also observed GFP in a population of Deiters' cells. TdTomato was not observed in this region in our previous study, which may be due to the limited recombination efficiency (described above) observed for nestin-CreERT2 [43].
An additional marker of adult stem cells is the Wnt target gene Lgr5, and its distribution has been described at various stages in the developing cochlea from E15.5 to P60 [54,55]. Comparison of their results with the current labeling for nestin indicates a possible degree of overlap. In view of the recent labeling for Lgr5-expressing cells in the cochlea, an important question therefore is to assess whether the nestin-positive and the Lgr5-positive cells constitute common or distinct cell populations. It is conceivable that Lgr5 labels a different subpopulation of cells in the organ of Corti and that nestin only labels progenitor cells in the developing embryonic epithelium, which is the case in the olfactory epithelium [52,56,57]. Investigators have shown that Lgr5-positive cells give rise to hair cells and are the source of regenerated hair cells in the neonatal cochlea following hair cell ablation [49,58]. Of interest, there appears to be some heterogeneity that exists among the Lgr5-expressing cell population resulting in only a subset of cells that respond to these experimental conditions [58]. This is also the case in the intestinal epithelium [59,60]. Moreover, findings in the olfactory epithelium, which exhibits continued regenerative capacity into adulthood, indicate nestin and Lgr5 are expressed in different cell populations [52,56,57]. In the olfactory epithelium, Lgr5 labels globose basal cells, which have the potential to become olfactory receptors, whereas nestin-expressing cells are a form of radial glia that do not harness any regenerative capacity [52,56,57]. Future studies to elucidate factors that contribute to the heterogeneity of Lgr5-positive cells in the organ of Corti and whether nestin contributes to this diversity may further inform our understanding of any regenerative capacity the sensory epithelium may possess and whether this persists into adulthood.
Data from our work demonstrate that nestin is expressed in multiple cell types in the developing inner ear. Moreover, a subset of these cells has the capacity to proliferate in the spiral ganglion region, although the exact identity of these cells is unknown. It is possible that nestin-expressing cells represent a unique subtype of cell in the spiral ganglion region that may respond to insult. The use of ouabain to induce auditory nerve degeneration may yield interesting results, given the unique position of nestin-expressing cells between the inner hair cell layer and spiral ganglion region. Ouabain is a well-known Na–K ATPase inhibitor that selectively removes type 1 spiral ganglion neurons [61,62]. In the optic nerve, upregulation of nestin with increased cell proliferation has been observed following insult [63]. Recently, stem cells isolated from the postnatal spiral ganglion have been shown to proliferate and give rise to neuronal and glial cell types [64]. Collectively, it is conceivable that nestin-expressing cells in the spiral ganglion region may respond to insult in a manner similar to cells in the subventricular zone and subgranular zone of the CNS [15]. Based on our findings, additional studies of the regenerative potential of nestin-expressing cells within the spiral ganglion are warranted and may provide insight into novel therapeutic approaches for neuronal forms of hearing loss and optimization of cochlear implant performance.
While the capacity of nestin-expressing cells in the organ of Corti to regenerate on any level is unknown, nestin-expressing cells in the spiral ganglion region possess some regenerative capacity based on the findings of this study. Additional analysis will be necessary to evaluate if nestin labels distinct subpopulations of cells with diverse functions in sensory and neural regions of the inner ear and if targeting these cell populations might provide a novel means for the treatment of hearing loss in the future.
Footnotes
Acknowledgments
We thank D. Oertel, M.J. Phillips, and N.P. Connor for generously allowing us to use their equipment at the University of Wisconsin-Madison; D. Wu, J. Jones, M. Beurg, X. Zhao, N.E. Patzlaff, E.M. Jobe, and R.J.H Smith for their expert discussion, assistance, and advice. This work was supported by the University of Colorado School of Medicine, Department of Otolaryngology; University of Wisconsin-Madison, Department of Surgery, KL2 award, and Type I pilot from the Clinical and Translational Science Award (CTSA) program, previously through the National Center for Research Resources (NCRR) grant 1UL1RR025011, and now by the National Center for Advancing Translational Sciences (NCATS), grant TL1 UL1TR000427; and by grants from the National Institutes of Health (NIH) numbers: NIH/NIDCD R25 DC010019, R03 DC012432-01 (S.P.G.), R01 DC013912-01 (S.P.G.), R01 DC01362 (R.F.), and NIH P30 HD003352.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.