
Abstract
Preterm infants are at high risk of developing severe sepsis. Circulating hematopoietic stem and progenitor cells (HSPCs; CD45+CD34+) have been suggested to play a vital role in the host immunological defense against invading pathogens. The objectives were to investigate the regulation of circulating HSPCs in preterm infants during infection episodes, and to assess the relationship of CD45+CD34+ cells with immunological mediators and differential leukocyte populations. First, we conducted a cross-sectional case-control study comparing these parameters among infected infants (n = 23), gestational and postnatal age-matched noninfected infants (n = 46), and “healthy” control (CTL) infants (n = 12). Second, we investigated the longitudinal change of CD45+CD34+ cell concentrations in infected infants before, during, and after an infection episode, and compared them with the other two groups. Our cross-sectional results showed that CD45+CD34+ cell count and percentage were significantly reduced in infected infants during systemic infection, compared with the noninfected or CTL infants. There were significant positive correlation between levels of CD45+CD34+ cells and lymphocytes or monocytes, and significant negative correlation between CD45+CD34+ cells and neutrophils or interleukin (IL)-6 in infected infants. Longitudinal analysis showed that changes of CD45+CD34+ cells at the onset of sepsis relative to levels 1 week prior and 1 week postsepsis in infected infants were significantly different from those changes in the corresponding time points for the other two groups. Our findings suggested that circulating HSPCs were dynamically regulated during septicemia and could play an important role in the defense mechanism, plausibly contributing to replenishment of leukocytes during sepsis in preterm infants.
Introduction
P
In early life, the innate immune system plays a pivotal role in the host defense against invading pathogens, especially before maturation of the adaptive immune response. Leukocytes contributing to the innate mechanism are endowed with remarkably diverse roles, which are essential for antimicrobial and inflammatory activities, including phagocytosis, generation of oxygen-free radicals, degranulation of antimicrobial proteins, secretion of pro- and anti-inflammatory cytokines, and crosstalking with the adaptive immune response. Most mature leukocytes have a limited lifespan, and are consumed through apoptosis after performing their functions in affected tissues [4]. Thus, during sepsis episodes, hematopoietic stem and progenitor cells (HSPCs), as marked by the CD45+CD34+ cell surface phenotype, are signaled to differentiate into various lineages of mature blood cells that egress from their bone marrow niche into the circulation to replenish immunological effector cells [5]. In rodent models, bacteremia induced by intravenous injection of Escherichia coli or in vivo stimulation with lipopolysaccharide increased HSPCs in the bone marrow and enhanced their mobilization into the peripheral blood [6,7]. Brudecki et al. recently demonstrated that adoptive transfer of CD34+ cells at the late phase of sepsis restored immunoreactivity and improved survival in a murine model [8]. Upon triggering by toll-like receptor (TLR) signals, human and murine bone marrow HSPCs have been shown to respond by an acceleration of differentiation toward the myeloid lineage [9 –11]. In addition to their presence in the bone marrow, there exists a small but consistent population of HSPCs circulating constantly between the bone marrow and blood. The physiological relevance of this pool of HSPCs is uncertain, but has been suggested to act as an immediate and adaptive source of HSPCs that could replenish innate immunological effector cells under both steady-state conditions and in response to inflammatory signals [12].
Although preterm neonatal blood contains relatively high levels of CD45+CD34+ cells [13,14], there has been no knowledge on their regulation in response to systemic infection. Our hypothesis was that circulatory HSPCs could be regulated during sepsis conditions, and might play a role in the host defense mechanism in preterm infants. The objective of the current study was to investigate changes in levels of circulating HSPCs in preterm infants during bacterial infection, and to determine whether this process was associated with stem cell mobilizing and inflammatory cytokines, thereby shedding light on the role of HSPCs in sepsis. Thus, we performed a cross-sectional case-control study to compare the levels of circulating CD45+CD34+ cells, granulocyte colony-stimulating factor (G-CSF) and interleukin (IL)-6 in three strictly defined groups of preterm infants: (1) infants with proven infection, (2) those with suspected infection but subsequently confirmed not to be infected, and (3) control (CTL) “healthy” preterm infants. In addition, we also investigated the longitudinal changes of blood CD45+CD34+ cells before, during, and after an episode of late-onset (>72 h of postnatal age) sepsis during early postnatal life (≤6 weeks of age).
Materials and Methods
Subjects
Preterm (≤32 weeks gestation) infants admitted to the Neonatal Unit (NNU), Prince of Wales Hospital and whose parents had given written consent were eligible for recruitment into the study. Parents could independently consent to the cross-sectional and/or longitudinal arms of the study. Infants with major chromosomal abnormalities, lethal congenital malformations, or family history of immune deficiencies were excluded. Infants who showed signs suggestive of sepsis were subjected to full sepsis evaluation as described previously [15]. The recruited subjects were categorized into three groups: (1) “infected” infants with late-onset systemic sepsis were defined as those with bacteria isolated from clinical specimens, including blood or cerebrospinal fluid cultures, and with concomitant elevation of serum C-reactive protein (CRP) >10 mg/L; (2) the “noninfected” infants were those who met the initial criteria for sepsis evaluation, but were subsequently proven not to be infected. Their blood cultures and other body fluids did not grow any organisms and all had normal levels of CRP ≤10 mg/L. They were later confirmed to have alternative diagnoses, including apnea of prematurity, exacerbation of bronchopulmonary dysplasia, anemia, and heart failure, and (3) “control (CTL)” infants were gestational and postnatal age-matched infants who showed no signs suggestive of clinical sepsis, and did not require sepsis evaluation. These CTL infants had blood levels of CD45+CD34+ measured in conjunction with routine weekly blood taking for monitoring of renal and liver function.
Study design
This study comprised two main components: the cross-sectional case-control study and the longitudinal cohort study. For the cross-sectional investigation, circulating levels of CD45+CD34+ cells, G-CSF, and IL-6 were measured in symptomatic infants, at the onset of clinical features and immediately before antibiotic treatment. Infected infants with proven sepsis (n = 23) were compared with gestational (±1 week) and postnatal age-matched (±1 week) noninfected (n = 46), and “CTL” (n = 12) infants. The CD45+CD34+ cell concentration and percentage of total white cell count were further correlated with CRP, cytokines, and hematological parameters, including total and differential white cell counts.
In the second part of the investigation, weekly longitudinal monitoring of CD45+CD34+ cell levels was performed in 32 preterm infants within 6 weeks of postnatal age. The change of CD45+CD34+ cell levels before, during, and after an episode of infection was evaluated in infected infants (n = 4), and compared with gestational age-matched noninfected infants (n = 8) and CTL infants (n = 8). For data analysis, the time of sepsis evaluation for infected or noninfected infants was assigned as week 0 and used as the reference time point for that particular group. In addition, we included a gestational age-matched CTL group at a postnatally matched time point for comparison.
Sepsis evaluation and blood sampling
According to our NNU protocol, a full sepsis evaluation was performed on each suspected infection episode, which included cultures of bacteria and fungi from: cerebrospinal fluid, blood, urine, endotracheal aspirate (infants on ventilator) and other body fluids (eg, pus, pleural and peritoneal fluids), indwelling central lines or catheters, and surgical and biopsy specimens, as indicated. Chest radiography was routinely performed, and abdominal radiography was requested when patients presented with clinical features suggestive of intra-abdominal pathology, such as necrotizing enterocolitis or intestinal perforation. Hematological and biochemical laboratory investigations, including serial white cell counts with differential cell counts and CRP measurements, were routinely performed. Whole blood samples were collected into a prechilled EDTA bottle, and transported immediately to the laboratory for processing (0.1 mL required for CD45+CD34+ cell concentration measurement). The blood specimens were taken concurrently with routine samples during sepsis evaluation. For the longitudinal study, whole blood samples were collected from preterm infants weekly from week 1 to 6 of postnatal age and coincided with the timing of weekly routine blood sampling for liver/renal function monitoring.
Quantification of plasma cytokines
Plasma samples were obtained by centrifugation of whole blood (1,900 g for 10 min) at 4°C and stored at −80°C until analysis. G-CSF and IL-6 were quantified by the Quantikine ELISA Kits (R&D Systems, Minneapolis, MN) according to manufacturer protocols.
Flow cytometric analysis of circulating CD45+CD34+ cells
Cell suspensions were prepared after red blood cell lysis in buffer EL (Qiagen GmbH, Hilden, Germany). Fc receptors were blocked by FcR reagent (Miltenyi Biotec, Bergisch Gladbach, Germany), and cell surface markers were stained with monoclonal antibodies CD34-FITC and CD45-APC (BD Biosciences, San Jose, CA) for 30 min at 4°C. Dead cells were identified by 7-amino-actinomycin D staining (BD Biosciences) and were excluded from analysis. Fluorescence signals from at least 500,000 events were acquired on a FACS LSRFortessa flow cytometer (BD Biosciences). CD45+CD34+ cells were phenotypically defined as HSPCs and quantified by the standardized and clinically employed ISHAGE gating procedure [16,17]. Staining with respective isotypic control antibodies served as negative control.
Statistical analysis
Mann–Whitney U test was used to compare circulating CD45+CD34+ cells, hematological and immunological parameters in infected, noninfected, and CTL infants in the cross-sectional study. Bonferroni's correction was applied for multiple comparisons. Spearman's correlation was used to assess the association between CD45+CD34+ cell levels and cytokines or hematological parameters. The longitudinal study on CD45+CD34+ cell levels was analyzed by the permutation test to assess differences in the change of CD45+CD34+ cell concentration and percentage (of the total white cell count) among the infected, noninfected, and CTL groups. The permutation test models all possible combinations of the available data, and calculates the probability that the test statistics (ie, the difference between group means) have occurred by chance. As this test makes no assumptions regarding normality of the population or independence of sampling of subjects, either of which is difficult to verify for small sample cases, it is more appropriate for verifying the difference between the means of two or more groups with the permutation test rather than the conventional Student's t-test [18]. Each group comparison was divided into two periods, 1 week before (week −1) or 1 week after (week +1) the sepsis onset time (week 0). The significance level of each test is set to be 0.05 with all computations conducted in R [19].
Patient consent
Written consent was obtained from parents of all recruited infants. This study was approved by the Joint Chinese University of Hong Kong-New Territories East Cluster Clinical Research Ethics Committee.
Results
Patient characteristics
Table 1 summarizes the clinical characteristics of the three groups of infants. There were no significant differences among infected, noninfected, and CTL infants with regard to gestational age, birthweight, sex, Apgar scores at 1 and 5 min, postnatal age of sepsis evaluation versus timing of blood sampling from the other two groups, and mortality rate. Five of the infected infants exhibited features of disseminated intravascular coagulation, compared with none of the noninfected or CTL infants.
Results are expressed as number or median (interquartile range).
Statistically significant after Bonferroni's correction for multiple comparison.
DIC, disseminated intravascular coagulation; NA, not applicable.
Comparison of circulatory CD45+CD34+ cell level, hematological and immunological parameters
Table 2 and Fig. 1 summarize the comparison of CD45+CD34+ cell concentration, hematological and immunological parameters among the three groups of preterm infants. Infected infants had significantly lower CD45+CD34+ cell counts and percentage (relative to CD45+ leukocytes) compared with noninfected or CTL infants. Platelet and lymphocyte counts were significantly lower in the infected group compared with noninfected or CTL infants. Monocyte count and percentage were significantly lower in infected infants compared with noninfected infants. Neutrophil counts, however, were similar among the groups, although there was a significant increase in the percentage of neutrophils in infected infants. As expected, CRP, IL-6, and G-CSF levels were significantly higher in infected infants than both noninfected and CTL infants. There was no significant difference between the noninfected and CTL group in any of the parameters.
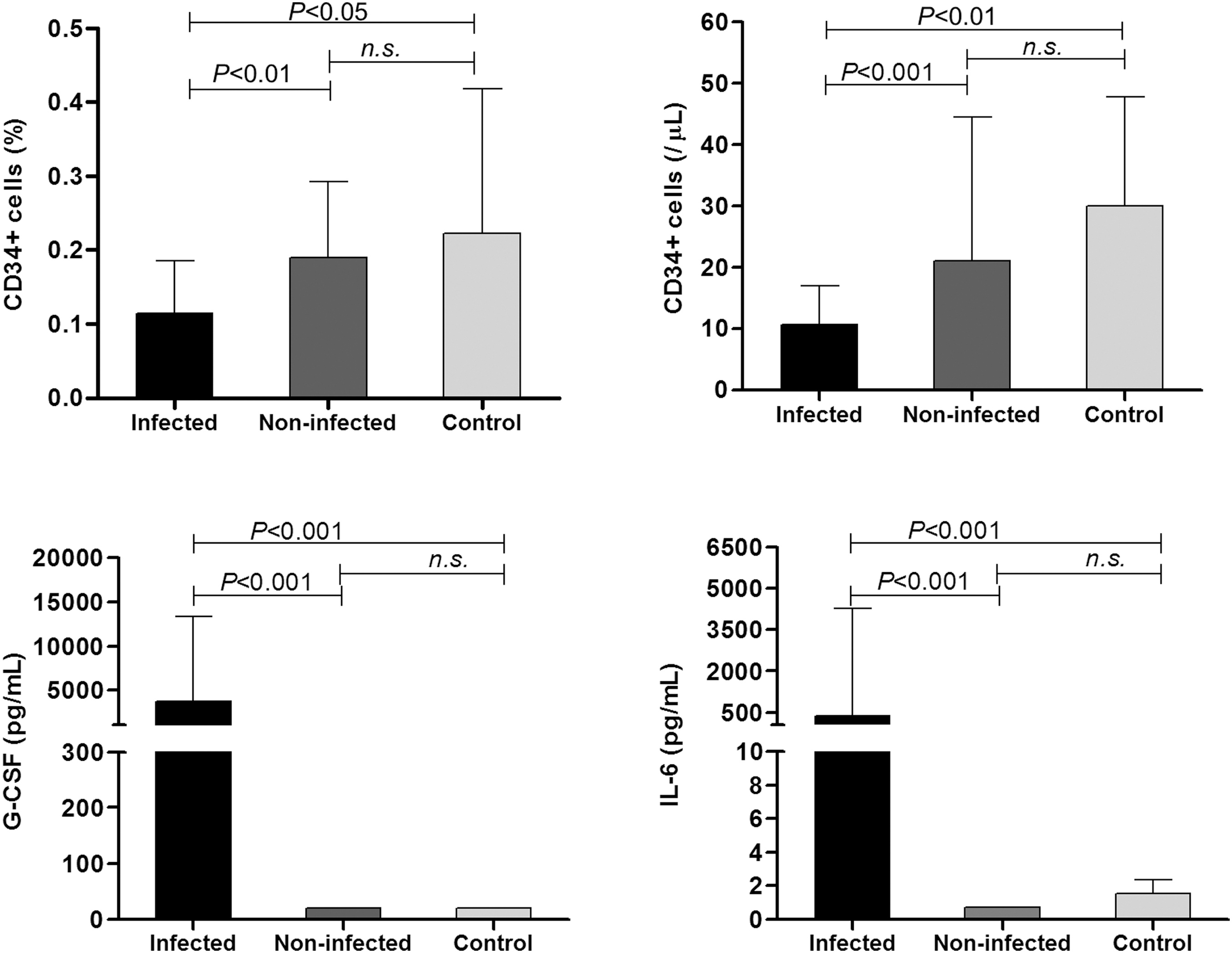
Comparison of circulatory CD45+CD34+ cells, G-CSF, and IL-6 levels in infected, noninfected, and control preterm infants. G-CSF, granulocyte colony-stimulating factor; IL, interleukin.
Results are expressed as median (interquartile range).
Statistically significant after Bonferroni's correction for multiple comparison.
Correlation between CD45+CD34+ cell level and immunological or hematological parameters
Correlation analyses were performed on the absolute count and percentage of CD45+CD34+ cells with respect to immunological and hematological parameters in infected infants (Fig. 2). There were significant positive correlations between CD45+CD34+ cell count and absolute lymphocyte or monocyte counts, and a significant negative correlation between percentage of CD45+CD34+ cells and percentage of neutrophils. In addition, there was a significant positive correlation between G-CSF and IL-6. IL-6 was negatively correlated with CD45+CD34+ cell count. CRP was not correlated with any of the investigated parameters.
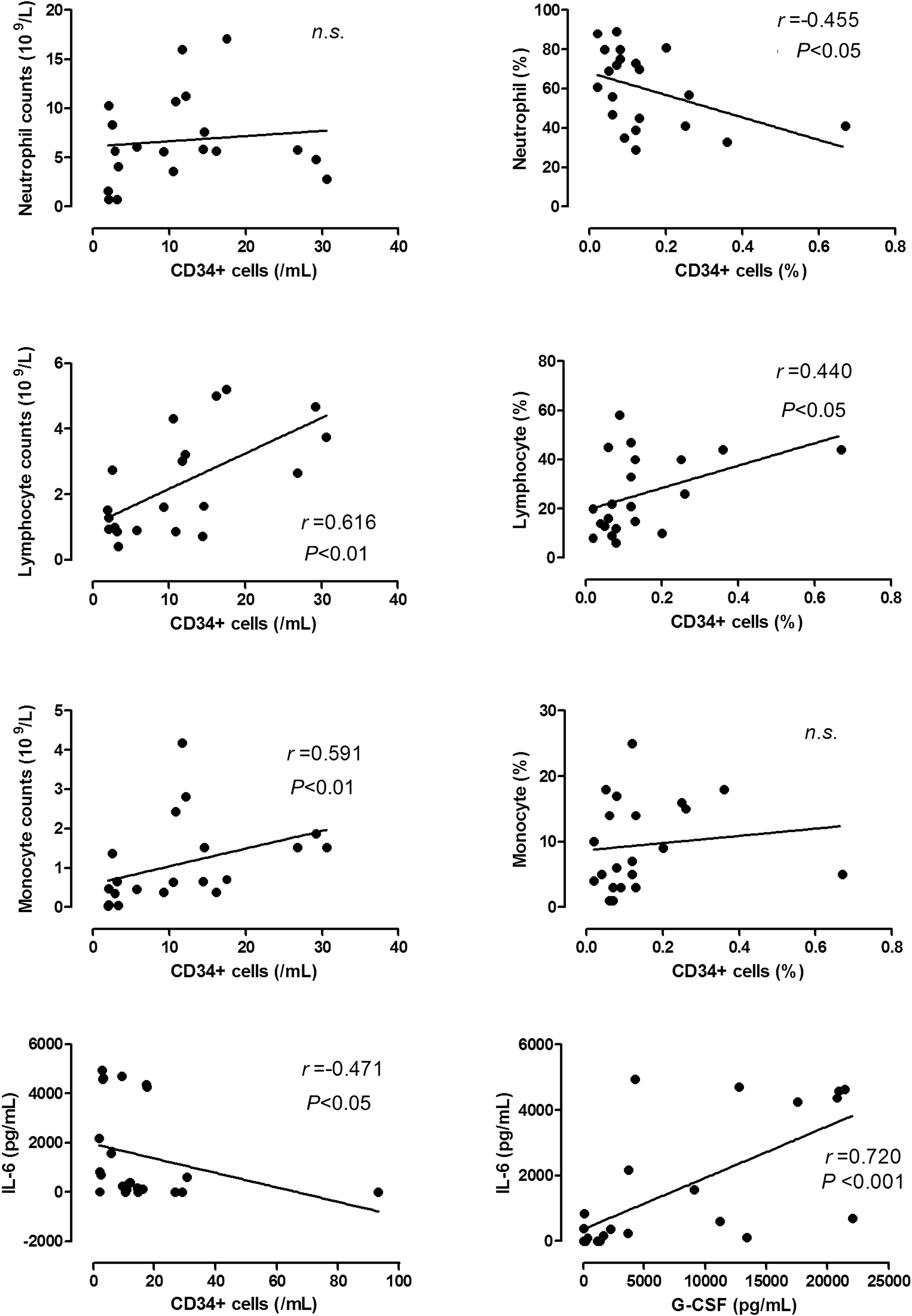
Correlation between CD45+CD34+ cell levels, leukocytes, and cytokines in infected preterm infants. Correlation analyses were performed between CD45+CD34+ cell count or percentage with levels of neutrophils, lymphocytes, monocytes, IL-6 or G-CSF in infected preterm infants (n = 23). n.s., statistically not significant; r, correlation coefficient.
In the noninfected group, only total white cell (r = 0.453, P < 0.01) or absolute neutrophil count (r = 0.370; P < 0.05) was positively correlated with CD45+CD34+ cell count. Concentrations of G-CSF and IL-6 were also positively correlated (r = 0.655; P < 0.001). No significant correlations were found between any of the studied parameters in the CTL group.
Longitudinal assessment of circulatory CD45+CD34+ cell level in infected, noninfected, and CTL infants
In the longitudinal study, we compared the magnitude of changes in circulatory CD45+CD34+ cell levels in the three groups of infants. Changes in cell count and percentage were calculated 1 week before (week −1) and 1 week after (week +1) the clinical presentation of sepsis, relative to the onset of sepsis (week 0). Within each group, the cell level at week 0 was set at 1 (baseline) so as to illustrate the magnitude of fold change between week −1 and week 0, and between week 0 and week +1. Between-group analysis showed significant differences in the fold change of CD45+CD34+ cell levels in infected infants compared with changes in corresponding time points for noninfected and CTL infants (Fig. 3). These results indicated that CD45+CD34+ cell levels were downregulated in infected infants at the onset of sepsis, and recovered after 1 week. In contrast, such regulation was not observed in the noninfected or CTL infants at equivalent postnatal time points.

Longitudinal changes of CD45+CD34+ cell level in infected, noninfected, and control preterm infants. Changes of CD45+CD34+ cell count and percentage were analyzed using sepsis evaluation (week 0) in infected or noninfected infants as the within-group reference time point. A gestational age-matched control group and at a postnatally matched time point was included for comparison. Significant differences in the change of CD45+CD34+ cell levels were observed both 1 week prior (ie, changes between week −1 and week 0) and 1 week after (ie, changes between week 0 and week +1) sepsis in the infected group (n = 4), compared with corresponding changes in CD45+CD34+ cell levels in the age-matched noninfected group (n = 8) and CTL group (n = 8). There was no significant difference in the change of CD45+CD34+ cell levels between the noninfected and CTL group. CTL, control.
Discussion
Our results provided the first evidence that circulatory CD45+CD34+ HSPCs are significantly downregulated in preterm infants during sepsis. We also observed longitudinal changes of the CD45+CD34+ cell level during the course of sepsis episodes. Stem cell mobilizing factor G-CSF and inflammatory cytokine IL-6 were increased in infected infants and the latter was negatively correlated with circulating CD45+CD34+ cell counts. We postulate that circulatory CD45+CD34+ cells could be an integral part of an active defensive mechanism in the innate cellular response even at such early gestations by replenishing mature leukocytes at the tissue level, in particular, providing neutrophils for protection against bacterial infection.
It is generally acknowledged that hematopoiesis during embryonic development is tightly regulated at restricted time periods and in precise locations. Definitive hematopoietic stem cells originate in the embryonic aorta–gonad–mesonephros and successively colonize the fetal liver, spleen, and finally bone marrow [20]. In the later fetal stage and throughout adult life, hematopoietic stem cells reside mainly in the bone marrow niche, where they continue to replenish the blood cellular system, including all immune responsive mature blood cells. In our previous study, we reported that preterm infants had higher levels of circulatory CD45+CD34+ cells compared with full-term infants, and they declined significantly within 48 h after birth [13].
To date, little information has been revealed on the roles of HSPCs in the circulation, especially whether they are actively involved in host defense of preterm infants. Although levels of circulatory HSPCs vary and span a large range among individuals [21], our cross-sectional and longitudinal data consistently revealed significant lower concentration and percentage of CD45+CD34+ cells during infection episodes. This suggests that their levels are actively regulated in response to bacterial pathogens or their products. However, this decrease of CD45+CD34+ cells in infected preterm infants appeared to be in contrast to the increased levels reported in adults [22]. Tsaganos et al. reported that CD45+CD34+ cells were significantly elevated in adult patients with sepsis due to pneumonia at day 1-7 after the onset of symptoms, compared with healthy subjects [22]. They also demonstrated that a high level of circulatory CD45+CD34+ cells (≥310/μL) at disease onset was associated with overall poor prognosis. Furthermore, these investigators observed a significant positive association between circulating CD45+CD34+ cell count and monocyte count. In the current study, circulatory CD45+CD34+ cell levels exhibited positive correlation with lymphocyte and monocyte counts, but a negative correlation with the percentage of neutrophils within the total white cell population. The number of HSPCs in the circulation, similar with other blood cells, is in a balance between consumption and replenishment from the bone marrow. Hematopoiesis and mobilization at the steady-state and sepsis conditions, however, could be very dynamic and influenced by many factors. The discrepancy between the observation in infants and adults could plausibly be due to different kinetics between mobilization and utilization of CD45+CD34+ cells. In preterm infants, the circulating HSPC population might not be sufficiently replenished by mobilization from the bone marrow and thus, appeared to be decreased in absolute number and percentage in response to infection. This phenomenon could indicate that the stem cell protective mechanism, although in operation, might still be compromised due to immaturity of the immunological system.
G-CSF is a multifunction cytokine that plays key roles in the differentiation of CD45+CD34+ cells into the myeloid lineage and mobilization of HSPCs from the bone marrow to the bloodstream [23,24]. It has been reported that G-CSF possesses crucial inflammatory properties by modulating antibacterial activities of neutrophils and monocytes [25]. G-CSF has also been clinically evaluated for treatment or prevention of neonatal infection [26,27]. In accordance with our findings, increase in G-CSF was observed in both infected preterm infants and adults suffering from infection [28]. IL-6 is a proinflammatory cytokine, which upregulates substantially in neonatal sepsis, and the magnitude of increase in IL-10 and IL-6 could be utilized as an indicator for predicting disease severity [29,30]. We observed a significant inverse correlation of IL-6 and CD45+CD34+ cell counts. Although it has been uncertain whether the kinetics of the HSPC response was affected by IL-6 or vice versa, our data did suggest that CD45+CD34+ cells, similar to G-CSF and IL-6, could be an integrated member of the immunological cascade in the host defense mechanism against infection. Our analysis revealed decrease of total lymphocytes and monocytes in infected infants, but neutrophil counts in these infants were similar to those of noninfected or CTL infants. We also observed a significant inverse correlation between the percentage of CD45+CD34 cells and neutrophils, which led us to suggest that differentiation of CD45+CD34+ cells might have occurred at the tissue level to maintain the number of circulatory neutrophils. In addition, changes of HSPC and leukocyte levels could be contributed by simultaneous increase of granulopoiesis and limiting lymphopoiesis in the bone marrow niche as regulated by G-CSF and other inflammatory factors (eg, SDF-1) [31,32]. With limited evidence in the human neonatal setting, the mechanisms governing the balance and association between HPSCs, leukocyte subpopoulations, and the network of interfering cytokines remain speculative and further studies are required to prove this important concept. Our results are in line with those of De et al., who reported in vitro stimulation of cord blood CD34+ cells by TLR agonists and bacterial antigens, which resulted in shifting CD34+ cell differentiation to myelopoiesis, including lineages of neutrophil, monocyte, and dendritic cell [9]. In the mouse model, Massberg et al. demonstrated that migratory HSPCs differentiated into tissue-resident myeloid cells at a steady-state condition and in response to TLR agonists, indicating that these cells might play an active role in immunosurveillance [12].
Unlike investigations in adult subjects, our research approach has been greatly limited by the constraints of collecting blood specimens from preterm infants. Our previous and other investigations have revealed that dysregulated circulatory cytokines/chemokines in infected infants might exhibit significant influences on HSPC and leukocyte functions (eg, G-CSF, IL-6, IL-10, SDF-1, MMP-9) [29,33]. The small volume of neonatal blood sampling, however, has precluded us from conducting a comprehensive analysis on the full spectrum of cytokines that could regulate HSPC mobilization and the innate immune response. Instead, we have focused on two key cytokines, that is, G-CSF and IL-6, which are crucial for HSPC mobilization and play vital roles in the proinflammatory cascade. Longitudinal blood sampling from preterm infants has been particularly challenging. Of 32 infants enrolled into the longitudinal arm, only 4 subsequently developed proven infection with septicemia. In addition, it is not feasible to conduct in vitro studies that require a substantially larger volume of neonatal blood to investigate the mechanism and fate of circulatory CD45+CD34+ cells upon stimulation by pathogens or their products. In spite of these intrinsic limitations, our findings on both the case-control and longitudinal experiments are consistent and in accordance with those of animal models [6,7], strongly suggesting that our data represent actual sepsis responses of preterm infants.
In conclusion, we reported the dynamic regulation of circulating CD45+CD34+ cells in human preterm infants during episodes of proven septicemia, and correlations of HPSCs with proinflammatory cytokine IL-6, and leukocyte counts. These results lead us to propose that circulating HSPC could play an integral role in the host defense, plausibly by timely replenishment of immunological effector cells at the tissue level. As circulatory CD45+CD34+ cells were significantly decreased in infected infants and increased upon recovery, analysis of their levels or subsets could further be explored as a parameter for prediction of sepsis at an early phase in vulnerable preterm infants.
Footnotes
Acknowledgments
This study was supported by the Health and Medical Research Fund, Food and Health Bureau, the HKSAR Government (project no.: 01120826). The funding organization did not play any role in the study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.