
Abstract
Umbilical cord blood (UCB) transplantation can provide a successful therapeutic option for patients that have no suitable related donor. UCB transplantation is often limited by the relatively small hematopoietic stem cell (HSC) numbers in UCB especially for adult recipients. Early neutrophil and platelet engraftment correlates with the stem cell numbers in UCB transplant. Compared to other HSC sources, immune reconstitution following UCB transplant is slower and complicated by increased frequency of opportunistic infections. The effect of HSC numbers in UCB transplant on immune reconstitution was not thoroughly examined. Using immunocompromised mice transplanted with purified UCB CD34+ stem cells, we have demonstrated that increasing the numbers of CD34+ cells in the transplant promotes hematopoietic and immune reconstitution. At early stages posttransplant, high stem cell dose generated relatively more B cells, while lower dose generated more myeloid and T cells. Thus, the size of the stem cell graft appears to modulate the differentiation potential of infused stem cells. In addition, increasing stem cell dose in the transplant improved CD8+ T cell development and delayed late memory T cell skewing in expense of naive T cells highlighting the importance of HSC dose to maintain the pool of naive T cells able to develop strong immune responses. Transplantation of ex vivo expanded CD34+ cells did not promote, but rather delayed immune reconstitution suggesting the loss of primitive lymphoid precursor cells during ex vivo expansion.
Introduction
U
Immunologic reconstitution is a critical factor in promoting a successful outcome following UCB transplantation. The successful reconstitution of donor T and B cells ultimately determines the quality of immune responses following UCB transplantation. Donor grafts contain B and T cells that contribute to adoptive antimicrobial and alloreactive effector cells at early stages posttransplant. At later stages posttransplant, B and T cells de novo generated from donor HSCs provide long-term immune reconstitution in the recipients of UCB transplant.
T cell reconstitution occurs in two phases [7]. For long-term immune reconstitution, a T cell expansion involving thymic production of new naive T cells is necessary to provide T cells with broad T cell repertoire. Hematopoietic progenitors derived from the engrafted HSCs within the BM enter the thymus to form early T cell precursors. During T cell development in the thymus, double positive thymocytes (CD4+CD8+) are exposed to self-major histocompatibility complex (MHC) on the thymic cortical epithelial cells.
Escalon and Komanduri reported delayed recovery of thymopoiesis in UCB transplantation compared to other HSC sources [8]. T cell reconstitution is delayed with a median time to recovery of ∼9–12 months [9]. B cell reconstitution occurs over the first 6 months post-UCB transplantation, although full recovery of immunoglobulins takes longer [10]. As a consequence of delayed immune reconstitution, UCB transplantation is associated with a significant risk of opportunistic infections associated with significant mortality following UCB transplantation [6,11]. Factors affecting lymphocyte reconstitution after UCB transplantation in children were HLA-matched UCB, higher nucleated cell dose, and positive recipient cytomegalovirus (CMV) serology before transplantation [10].
Using purified UCB-derived CD34+ cells rich in HSCs, we show that increasing CD34+ cell dose in the graft acts to promote immune regeneration in immunocompromised mice. Mice transplanted with larger HSC grafts exhibited better multilineage engraftment at early and late stages posttransplant. Increased donor CD34+ cell numbers and numbers of colony forming units (CFUs) were identified in the BM of recipients of larger HSC grafts. Increasing the HSC dose in the graft promoted CD8+ T cell development and delayed skewing toward an effector memory T cell phenotype (Tem) at the expense of naive T cells (Tn) potentially improving T cell–mediated immune responses.
Ex vivo expansion was attempted to increase the numbers of HSCs in the infused product [12 –17]. In this study, we used minimally cultured CD34+ cells based on our previous studies of UCB CD34+ cell expansion [18]. Infusion of ex vivo expanded CD34+ cells did not lead to improved hematopoietic and immune reconstitution; in fact the recipients of expanded grafts exhibited inferior human cell engraftment and delayed B and T cell reconstitution suggesting that ex vivo expansion may compromise the regenerative function of HSCs.
Materials and Methods
Human stem cell processing
UCB was provided by Sydney Cord Blood Bank. All experiments were approved by the Sydney Children's Hospital Human Research Ethics Committee. Mononuclear cells were purified from UCB by Ficoll-Hypaque (Sigma Chemical Co., St. Louis, MO) density-gradient centrifugation. CD34+ cells were separated to greater than 95% purity by positive selection using a magnetic cell purification system (autoMACS Miltenyi Biotec, Auburn, CA).
Stem cell expansion
CD34+ cells were seeded at a density of 0.5–1 × 105 cells/mL in serum-free medium Stemline II (Sigma Aldrich), 2 mM
Human stem cell transplantation
Approval from the UNSW Animal Care and Ethics Committee (ACEC) was obtained for animal experiments. Severely immunocompromised NOD/SCID-IL2Rgnull (NSG) mice were obtained from the Jackson Laboratory and bred in the animal facility at Children's Cancer Institute Australia. Female NSG mice at 6–8 weeks of age were irradiated with a sublethal dose of 2.5 Gy, 12–24 h before being transplanted with human stem cells through intravenous (IV) injection. Weekly bleeding was performed from the tail vein to monitor human cell engraftment in peripheral blood (PB) of transplant recipient mice. Human and murine cells were distinguished by the species-specific expression of pan-hematopoietic marker CD45. The proportion of cells expressing human CD45 (hCD45) was defined as the level of engraftment. Antibodies against human (h) CD19, CD33, and CD3 were used to examine multilineage reconstitution in PB, BM, and spleen (SPL). T cell subsets were analyzed using antibodies against hCD4, CD8, CD62L, and CD45RO (all from Becton Dickinson; San Jose, CA). 1 × 105 events were acquired on a FACSCanto, and data were analyzed with FACSDiva software (Becton Dickinson, San Jose, CA). Viable cells were gated based on forward versus side scatter (FSC/SSC).
Graft-versus-host disease
Transplanted mice were monitored for signs of Graft-versus-Host Disease (GVHD) according to an established mouse GVHD grading system [19]. The symptoms of GVHD were monitored by weight loss, mortality, and clinical parameters. Histopathological examination of the BM, SPL, and liver was conducted postmortem to identify mononuclear cell infiltration [19].
CFU assay
CFU numbers produced by hCD45+ cells derived from the BM of transplant recipient mice were measured using methylcellulose that does not contain human erythropoietin (MethoCult4534). A total of 2 × 104 hCD45+ cells from each mouse were plated in 1.1 mL of methylcellulose and incubated in humidified atmosphere for 14 days at 37°C and 5% CO2. On day 14, all cultures were scored for total colony formation.
Statistical analysis
All experiments were performed at least twice. Six to 12 mice per group were used in transplant experiments. Differences between groups were examined for statistical significance using Student's t-test. Mean and standard deviation were calculated with GraphPad Prism 5 software.
Results
Increasing CD34+ cell dose in the transplant promotes human hematopoietic and immune reconstitution in NSG mice
Human CD45+ cells were identified in PB of transplant recipient mice 4 weeks posttransplant (Fig. 1A). The proportion of hCD45+ cells in PB progressively increased with time and reached the plateau at week 12 posttransplant (Fig. 1A). The majority of hCD45+ cells coexpressed B cell marker hCD19 (Fig. 1B). The proportion of hCD19+ B cells increased over time, reaching the maximum magnitude of up to 50%–60% at week 12–14 and then gradually declined (Fig. 1B). Human CD33+ myeloid cells were detected in PB from week 7, and the percentage of hCD33+ cells gradually increased with time (Fig. 1C). Human CD3+ T cells were identified in PB 12 weeks posttransplant (Fig. 1D). T cell engraftment in PB gradually increased with the time (Fig. 1D). One representative flow cytometry analysis for each cell subset is shown in Supplementary Fig. S1 (Supplementary Data are available online at
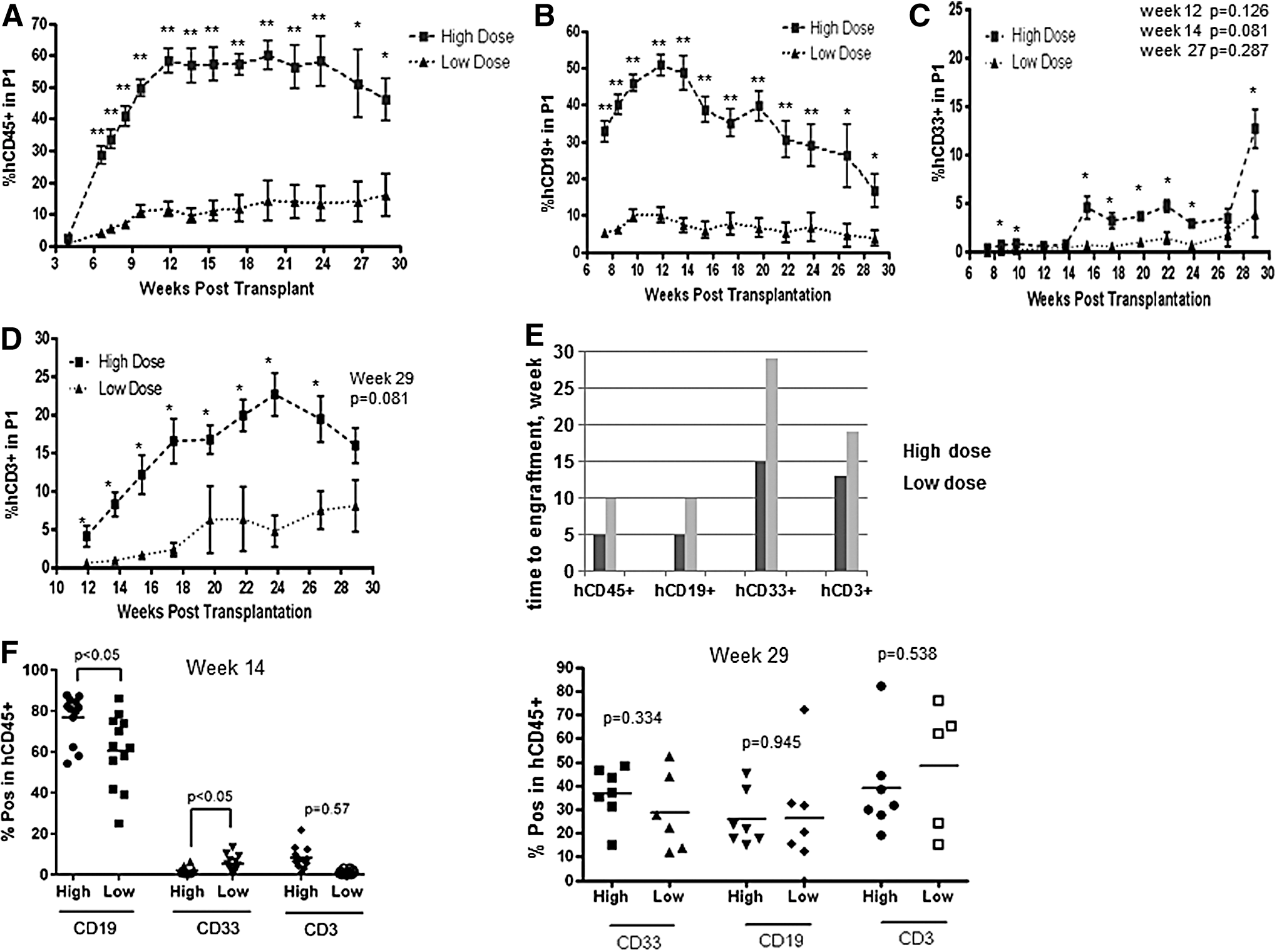
Increasing CD34+ cell dose in the transplant promotes human hematopoietic and immune reconstitution in NSG mice. Sublethally irradiated mice (n = 22) were transplanted with 0.1 or 0.5 × 105 CD34+ cells per mouse (low and high dose, n = 11, respectively). Four mice from each group were analyzed postmortem at week 14 and seven mice at week 29 posttransplant. Proportion of hCD45+
Increasing CD34+ cell dose from 0.1 to 0.5 × 105 cells per mouse promoted multilineage engraftment increasing the maximum level of donor chimerism and reducing the time to engraftment (Fig. 1A–E). In addition, the proportion of hCD19+ cells within hCD45+ gate was significantly higher in mice that received the higher stem cell dose (Fig. 1F, left panel). In contrast, the proportion of hCD33+ cells within hCD45+ gate was significantly lower in high stem cell dose group (Fig. 1F, left panel). At week 29 posttransplant, there were no statistically significant differences between the proportion of B and myeloid cells between two groups (Fig. 1F, right panel). The proportion of hCD3+ cells in hCD45+ gate did not significantly differ between groups at week 14 and 29 (Fig. 1F, left and right panels).
Infusion of larger numbers of CD34+ cells had positive effect on human cell generation in the BM. At week 14 and 29, hCD45+ and hCD19+ cell engraftment was higher in mice infused with larger numbers of CD34+ cells (Fig. 2A, B). There were no significant differences in hCD33+ cell engraftment between two groups at week 14 and 29 (Fig. 2A, B). Human CD45+ and CD19+ cell engraftment in the BM was significantly lower at week 29 compared with week 14 in both groups of mice (Fig. 2A, B). In contrast, hCD33+ cell engraftment in the BM was significantly higher at week 29 compared with week 14 in both groups (Fig. 2A, B).
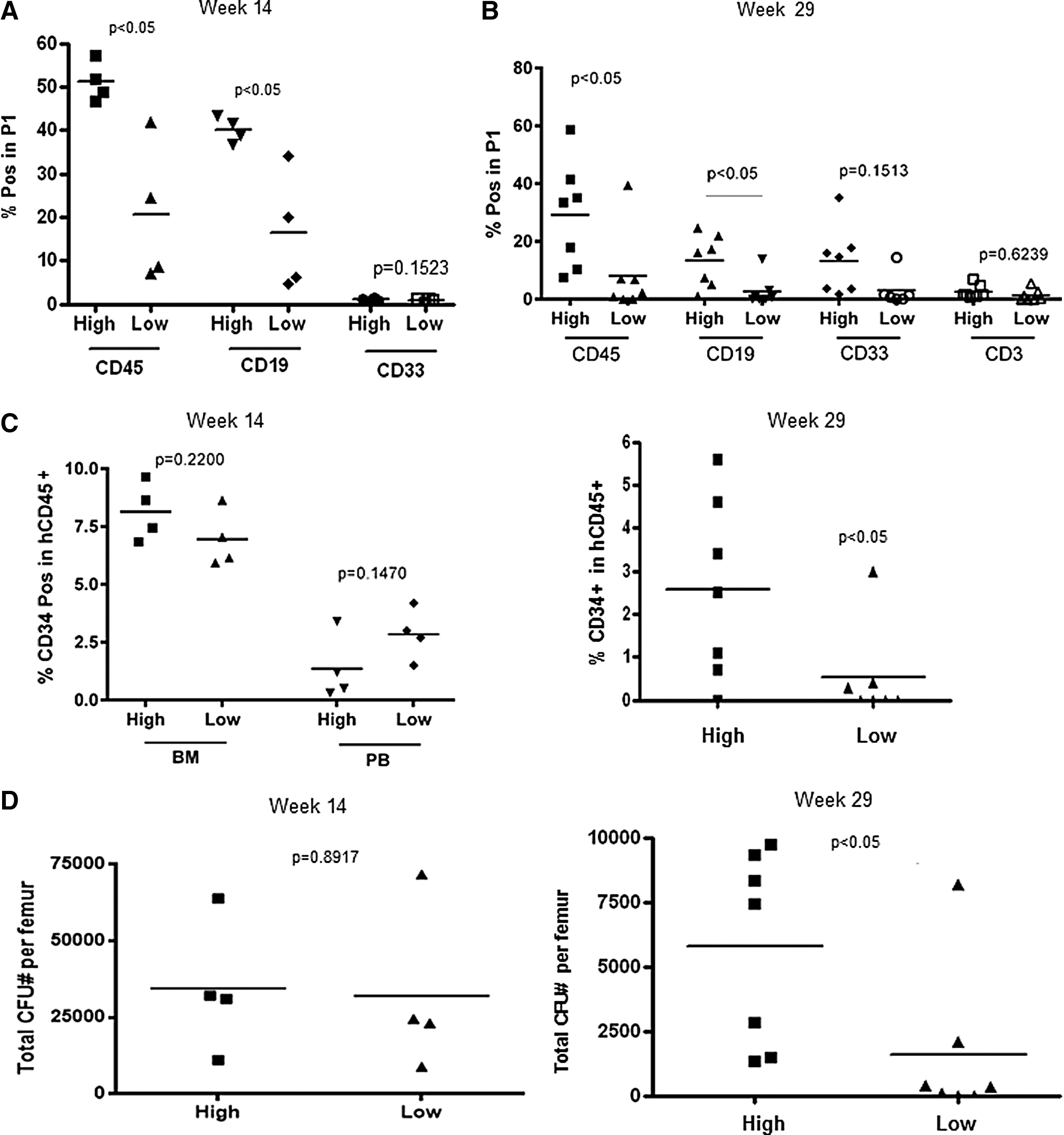
Human multilineage engraftment in the BM.
Human CD3+ T cells were hardly detectable in the BM at week 14 (data not shown). Low T cell infiltration of BM was observed at week 29 posttransplant (Fig. 2B). The proportions of primitive hCD34+ progenitor cells identified in the BM and PB at week 14 were similar in both groups of mice (Fig. 2C). At week 29, the proportions of hCD34+ cells in BM were significantly higher in the recipients of larger stem cell grafts (Fig. 2D). Of note, the proportion of hCD34+ cells in the BM at week 29 was significantly lower than at week 14 in both groups (Fig. 2C, left and right panels). Human CD34+ cells were not detectable in PB at week 29 (data not shown). Total CFU numbers per femur did not differ between groups at week 14 (Fig. 2D, left panel). At week 29, however, there were significantly more CFUs in the BM of mice infused with a larger dose of hCD34+ cells (Fig. 2D, right panel) suggesting that the infusion of larger numbers of stem cells increases the regenerative capacity of transplanted hCD34+ cells in the BM. Infusion of larger numbers of stem cells increased hCD45+ and CD19+, but not hCD33+ cell engraftment in the spleen (SPL) analyzed at week 14 posttransplant (Fig. 3A). Human CD3+ cells were undetectable at week 14 posttransplant (data not shown). In addition, hCD45+, hCD19+, and hCD3+, but not hCD33+ cell engraftment was higher in the SPL of the recipients of high dose compared with low dose of stem cells at week 29 posttransplant (Fig. 3B).
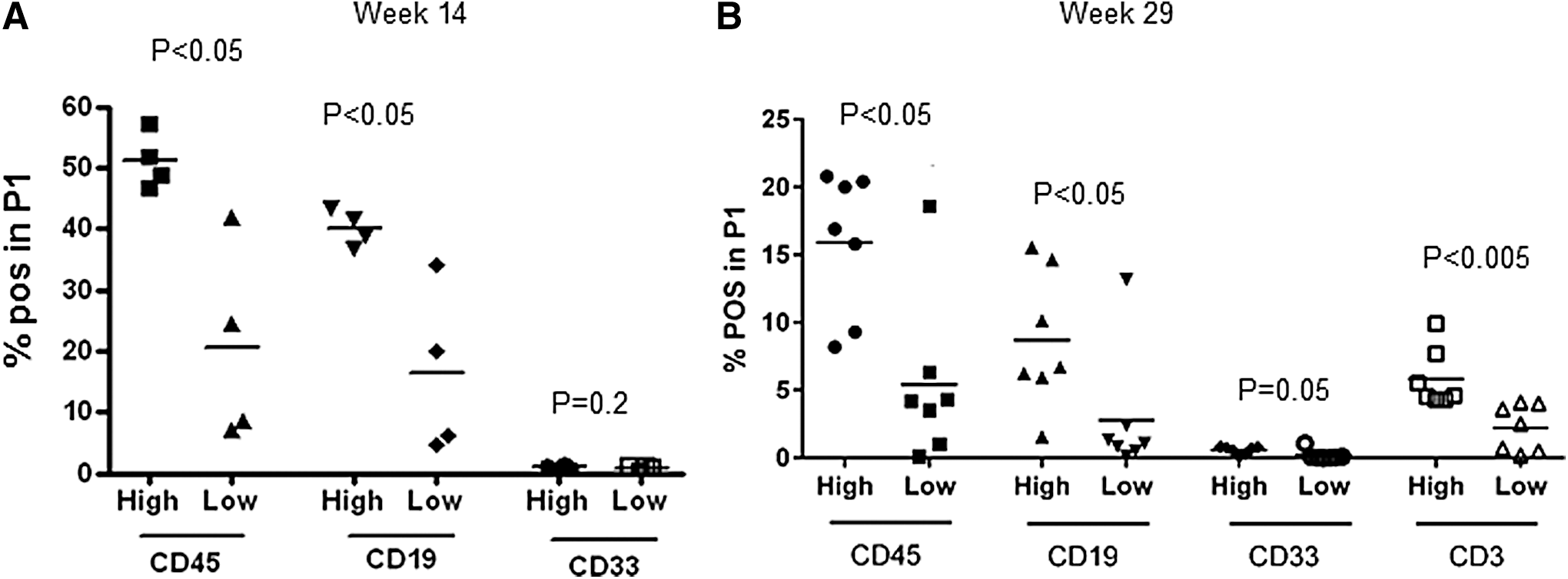
Human multilineage cell engraftment in the SPL of mice transplanted with high and low doses of CD34+ stem cells.
Increasing stem cell dose in the transplant promotes CD8+ T cell development and preserves naive T cell phenotype
Mice infused with larger numbers of human stem cells exhibited higher proportion of hCD8+ T cells and lower proportion of hCD4+ T cells in PB and SPL compared with mice that received smaller stem cell grafts (Fig. 4A, B). In addition, mice transplanted with larger numbers of CD34+ cells exhibited higher proportion of CD62L+CD45RO− naive T (Tn) cells in the CD4+ T cell subsets compared with the mice that received smaller stem cell grafts (Fig. 5A). The proportions of CD62L+CD45RO+ central memory T cells (Tcm) and CD62L−CD45R+ effector memory T cells (Tem) in hCD4+ T cell subset in PB were not affected by the dose of the stem cells infused (Fig. 5A). The proportions of Tcm in CD4+ cell subset isolated from the SPL, however, were higher in high stem cell dose group, while the proportion of Tem was lower than in low stem cell dose group (Fig. 5B). CD8+ T cells exhibited more Tn and less Tcm and Tem cells than CD4+ T cells due to delayed CD8+ cell differentiation in both groups of mice (Fig. 5A, C). The percentages of Tn, Tcm, and Tem in CD8+ T cells from PB were similar between mice infused with large and small numbers of CD34+ cells (Fig. 5C). T cells recovered from the SPL exhibited lower proportion of Tn and higher proportions of Tcm and Tem than T cells isolated from PB (Fig. 5C, D). Importantly, mice infused with the larger numbers of CD34+ cells had a higher proportion of Tn and a lower proportion of Tem cells in CD8+ T cell subsets of the SPL compared with the mice transplanted with lower numbers of CD34+ cells (Fig. 5D).
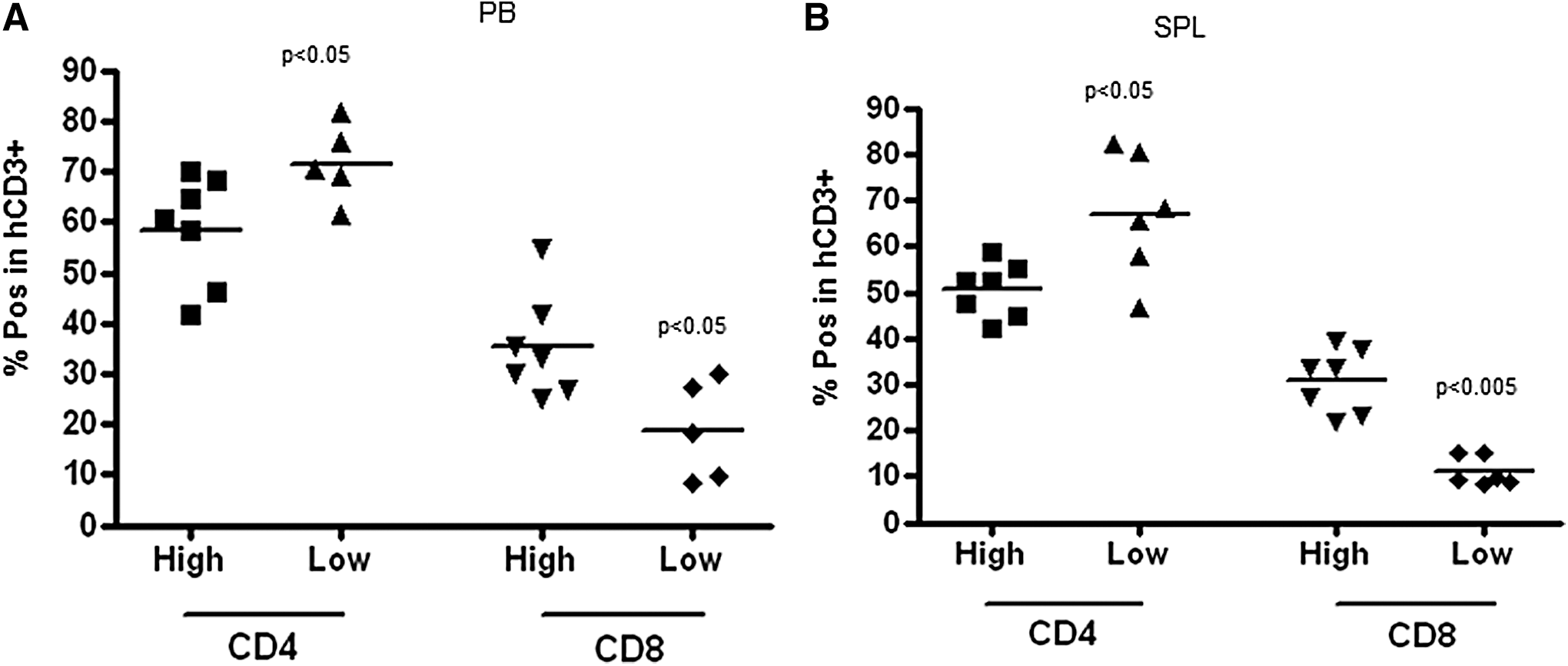
Human CD4+ and CD8+ T cell engraftment in PB
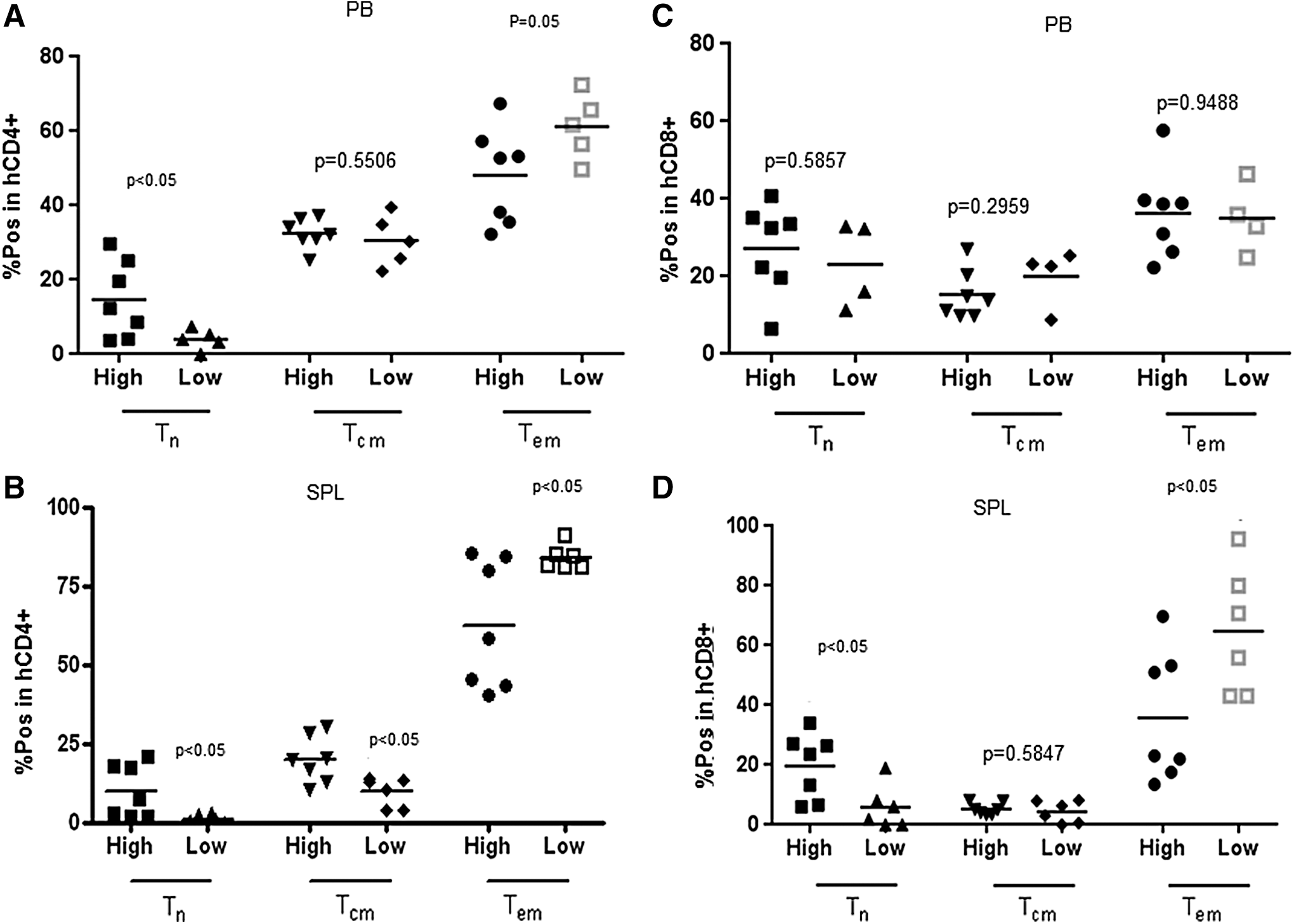
Proportions of CD62L+CD45RO− Tn, CD62L+CD45RO+ Tcm, and CD62L−CD45RO+ Tem cells in CD4+ T cells isolated from PB and SPL (
Ex vivo expansion of CD34+ progenitor cells does not promote immune reconstitution
On the basis of our previous studies, UCB-derived CD34+ cells were cultured for 6 days in the presence of a cytokine cocktail consisting of SCF, Flt3L, and TPO, all at 100 ng/mL [18]. Under these conditions, 3–10-fold increase in the number of CD34+ cells was obtained [18]. In this study, we transplanted mice with 1 × 105 fresh CD34+ cells (unexpanded group) or the ex vivo expanded equivalent of 1 × 105 unexpanded CD34+ cells (expanded group). Ex vivo expanded cell grafts contained 8 × 105 total mononuclear cells 40% of which were CD34+ cells in this particular experiment. Transplantation of ex vivo expanded stem cells did not promote hematopoietic reconstitution, in fact, hCD45+, hCD19+, and hCD33+ cell engraftment was delayed in mice transplanted with ex vivo expanded CD34+ cells (Fig. 6A–C). Given that the mice in “expanded” group were transplanted with 3.2-fold higher dose of hCD34+ cells than mice from “unexpanded” group, we conclude that multilineage engraftment normalized to the input CD34+ stem cells was reduced following ex vivo expansion. The kinetics of CD3+ T cell engraftment in PB was similar in mice transplanted with fresh or ex vivo expanded stem cells at first 24 weeks posttransplant; however, significantly reduced proportion of CD3+ T cells was identified in PB only at week 27 posttransplant (Fig. 6D). Proportions of CD4+ and CD8+ T cells in PB were similar in mice from both groups (Supplementary Fig. S2A), and the proportions of Tn, Tcm, and Tem cells in CD4+ and CD8+ T cell gates were not modulated by ex vivo expansion (Supplementary Fig. S2B). In addition, similar numbers of hCD4+ and hCD8+ T cells were recovered from the SPL of the recipients of expanded and unexpanded stem cell grafts (Supplementary Fig. S2C). Human CD45+ and hCD19+ cell engraftment in the BM and SPL did not significantly differ between the recipients of expanded and unexpanded grafts (Fig. 7A, B). Human CD33+ myeloid cell engraftment in the BM and SPL, however, was significantly higher in mice infused with the ex vivo expanded stem cells (Fig. 7A, B). We speculate that the reduced egress of myeloid cells from the BM to PB may account for the reduced hCD33+ cell engraftment in PB of the recipients of ex vivo expanded grafts; CFU and CD34+CD38− cell numbers generated by ex vivo expanded CD34+ cells were lower compared with unexpanded CD34+ cells (Fig. 7C, D) suggesting that ex vivo expansion compromised long-term regenerative function of the stem/progenitor cells and/or led to the loss of primitive immune precursor cells.
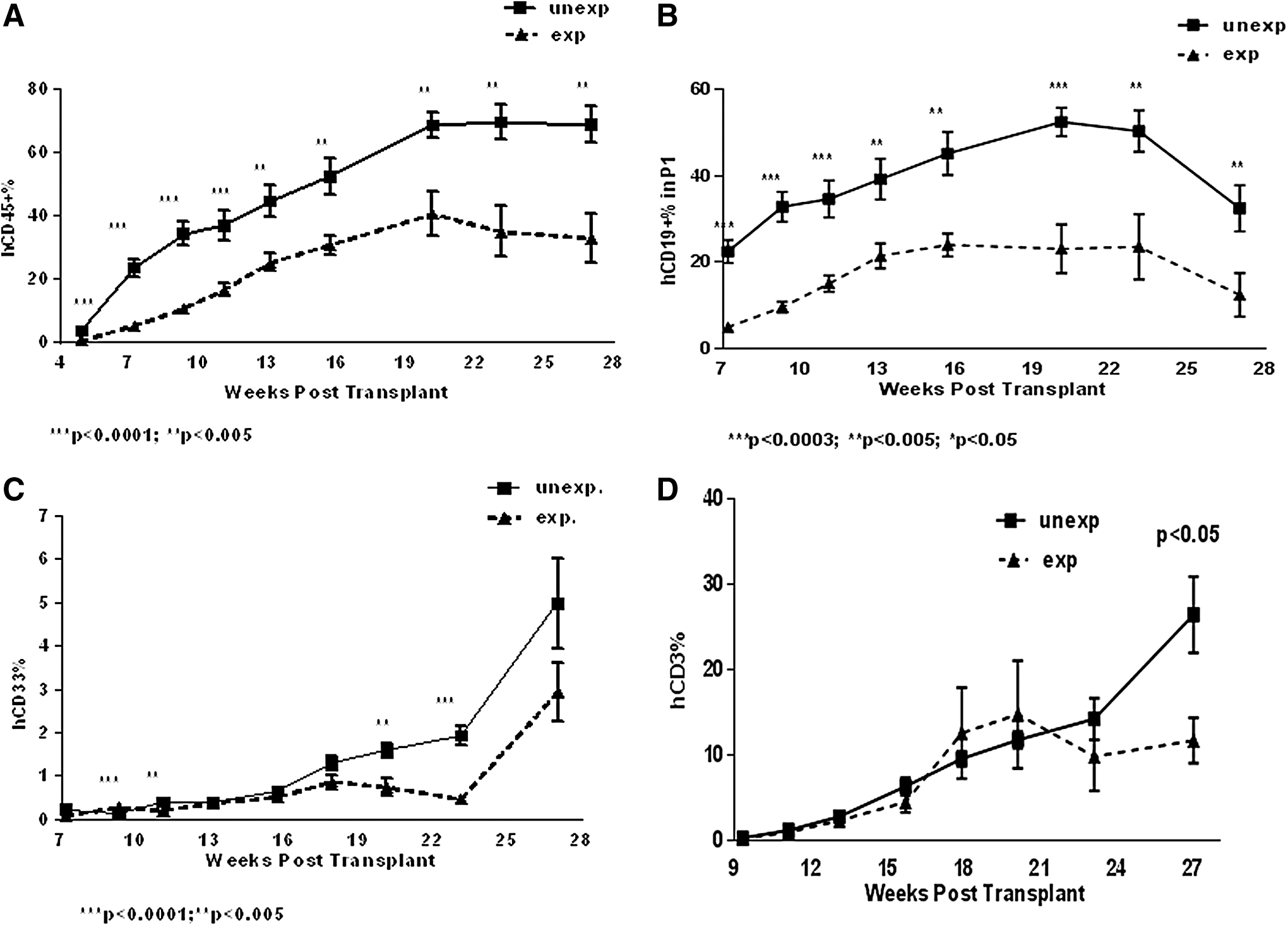
Multilineage reconstitution in PB of mice transplanted with unexpanded or ex vivo expanded stem cells. Mice were transplanted with unexpanded or 6-day expanded CD34+ cells. Twelve mice per group were used in this experiment. Two experiments using CD34+ stem cells derived from two different UCB donors were conducted comparing unexpanded and ex vivo expanded stem cells. Proportions of hCD45+, hCD19+, hCD33+, and hCD3+ cell engraftment in PB (
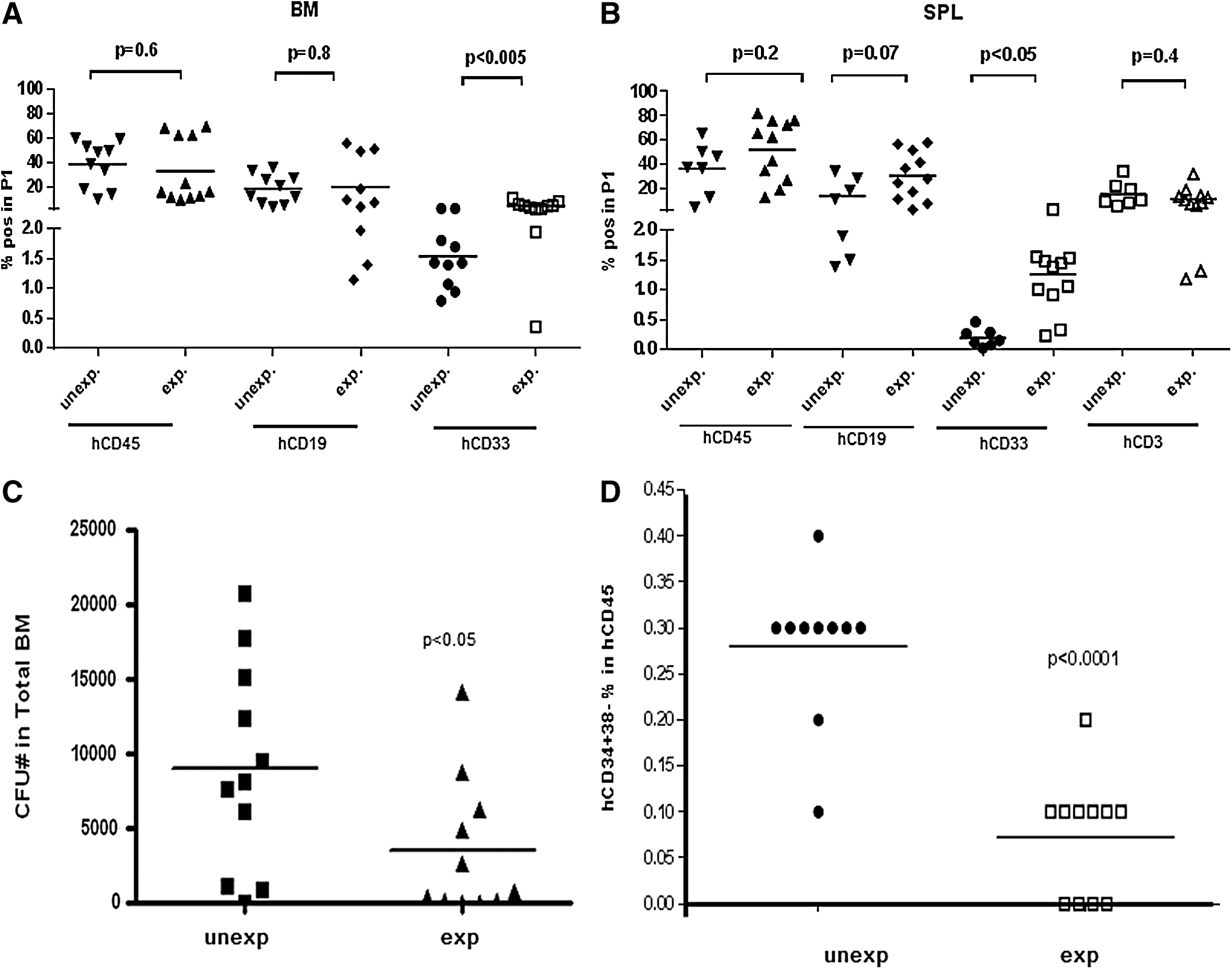
Multilineage reconstitution in BM and SPL in mice transplanted with unexpanded or ex vivo expanded stem cells.
Discussion
UCB is used as a source of HSCs for transplantation for both malignant and nonmalignant diseases [1]. The total number of HSCs in UCB grafts, however, is lower relative to the number in a BM or mobilized PB allografts, which lead to longer engraftment times [1 –5]. The immune reconstitution is also delayed in adult recipients of UCB transplants [8,9]. The impact of CD34+ cell numbers in UCBT has been extensively studied demonstrating the shortening of the time to engraftment of neutrophils and platelets in patients transplanted with higher doses of CD34+ cells.
In this study, we used NSG mice transplanted with highly purified UCB-derived CD34+ cells to examine the impact of CD34+ cell dose on posttransplant immune reconstitution. In this model, B and T cell development was purely derived from the infused CD34+ stem/progenitor cells excluding the impact of mature subsets. The kinetics of human multilineage reconstitution was evaluated in PB at multiple time points at early and late stages posttransplant. Human B cells were detectable in PB as early as 4 weeks posttransplant and then progressively increased with the time posttransplant. Both myeloid and T cell reconstitutions were delayed in NSG mice infused with UCB-derived CD34+ stem cells. In clinical UCB transplantation, however, myeloid reconstitution precedes B and T cell regeneration. Delayed myeloid reconstitution in NSG mice transplanted with human CD34+ cells is likely due to the low cross-reactivity between human and murine myeloid-specific growth factors [18,20]. This may limit the applicability of the model.
Increasing CD34+ cell dose positively affected posttransplant immune regeneration. The recipients of larger stem cell transplants exhibited larger numbers of CD19+ B cells, CD33+ myeloid cells, and CD3+CD4+ and CD3+CD8+ T cells in PB than the recipients of the smaller stem cell grafts. Transplanting more stem cells promoted multilineage hematopoiesis at early stages and increased the magnitude of reconstitution at late stages posttransplant. Our week 14 analysis revealed that transplantation of larger numbers of CD34+ cells increased human B and myeloid hematopoiesis in the BM. Improved T cell reconstitution in mice transplanted with larger numbers of hCD34+ cells also correlated with the increased engraftment of early human progenitors in the BM. Therefore we speculate that transplantation of larger numbers of stem cells increasing the numbers of human progenitor cells populating the recipient's BM may also increase the proportion of human BM progenitors populating the host thymus and promoting human T cell development.
We have also shown that at early stages posttransplant, high stem cell dose generated relatively more B cells, while lower dose of stem cells generated relatively more myeloid and T cells. Human B cells may be less affected by the low cross-reactivity of human and mouse cytokines than myeloid and T cells; therefore, increasing the numbers of infused HSCs may promote B cell regeneration to greater extent compared with myeloid cell and T cell regeneration. It is relevant that increased B cell versus T cell potency was identified in the recipients of larger stem cell grafts in mouse transplantation model suggesting that the size of the stem cell graft appears to modulate the differentiation potential of stem cells [20].
Increasing the numbers of infused CD34+ cells modulated specific T cell subsets in the PB. The T cell compartment in PB is maintained by homeostatic thymus-independent proliferation of Tn cells. Tn cell proliferation in PB leads to premature effector T cell differentiation reducing the Tn cell pool and, therefore, potentially reducing the potency of T cell mediated immune responses [7,8,21]. In this study, we show that the larger stem cell grafts generated bigger absolute numbers of Tn, Tcm, and Tem cells in PB and SPL and delayed Tem cell skewing at the expanse of Tn cells. Transplantation of larger numbers of stem cells is likely to increase the numbers of human progenitor cells repopulating the recipient's BM and thymus and may, therefore, reduce the need of extensive homeostatic Tn cell proliferation in PB preserving Tn cells from differentiation into Tem cells and ultimately increasing the immune potency of the graft. Subset analysis of newly generated T cells also revealed the increased proportion of CD8+ T cells in mice transplanted with the higher dose of human CD34+ cells (Fig. 4A, B). Human CD8+ T cell development in NSG mice is suppressed by the lack of HLA class 1 expression [22]. We speculate that larger numbers of stem cells generated more HLA class 1 expressing cells promoting CD8+ T cell development.
The speed of immune reconstitution in a mouse transplantation model using murine T cell depleted BM stem cells was shown to correlate with the dose of infused stem cells [23]. The improved immune recovery was shown to be due to enhanced thymopoiesis in this study. Analysis of naive and memory T cell phenotypes, however, was not conducted in this study. The impact of stem cell dose on posttransplant hematopoietic and immune reconstitution has been also examined in “humanized” NSG mice infused with LinnegALDHbr fraction of UCB CD34+ cells [24]. This study has provided evidence that LinnegALDHbr CD34+ cell dose influenced both the strength and the pace of hematopoietic chimerism within UCB transplant. The effect of stem cell dose on T cell subsets, however, was not conducted due to insufficient T cell numbers recovered from the SPL and PB of mice transplanted with low stem cell numbers. Thus, to the best of our knowledge, our study is the first report demonstrating that increasing stem cell numbers in transplant promotes immune reconstitution in “humanized” mouse model.
Ex vivo expansion of HSCs represents a strategy to increase the number of HSCs in the graft [12 –18]. There are concerns, however, that ex vivo expansion of UCB may result in differentiation of HSCs and predominant expansion of committed myeloid progenitor cells in expanse of lymphoid compartment, thereby delaying immune reconstitution. CD34+ UCB cells ex vivo expanded in the presence of early acting cytokines for 14 days tested in fetal thymic organ cultures generated only half the CD4+TCR+T cells that was seen in unexpanded CD34+ cells [25]. Later, the same group has shown that 14-day cultured CD34+ cells transplanted to NOD/SCID mice have the capacity to differentiate into mature T cells ex vivo to the same extent as fresh CD34+ cells [26]. Finally, 14-day expanded CD34+ cells transplanted into conditioned NSG mice differentiated into myeloid and lymphoid lineages; however, T cell development required administration of human TNFα before transplantation [27]. The comparative analysis of T cell development in mice transplanted with fresh or ex vivo expanded CD34+ cells was not conducted in this study.
Here we show that transplantation of ex vivo expanded UCB-derived CD34+ cells does not promote immune regeneration, in fact, B and myeloid cell engraftment in PB was delayed in the recipients of ex vivo expanded stem cells. The inferior engraftment in PB, however, did not correlate with reduced B and myeloid cell chimerism in the BM, suggesting that reduced egress of B and myeloid cells from the BM to PB may account for the reduced engraftment in PB of the recipients of ex vivo expanded stem cells. Reduced primitive CD34+ and CD34+CD38− progenitor and CFU numbers in the BM of mice reconstituted with ex vivo expanded CD34+ cells are suggesting that extensive ex vivo proliferation of CD34+ cells before infusion results in the reduced long-term regenerative capacity of the graft. We have previously shown that extensive proliferation of UCB stem/progenitors during ex vivo expansion results in transcriptional dysregulation of signaling networks and impaired regenerative activity [18].
Importantly, ex vivo expansion of stem cells did not benefit T cell reconstitution. CD4/CD8 ratio and naive and memory T cell subsets were not modulated by ex vivo expansion of stem cells before infusion. Collectively, these results are suggesting that the numbers and the “quality” of infused stem cells determine immune regeneration following UCB cell transplantation.
Footnotes
Acknowledgments
The authors thank Children's Cancer Institute Australia for supporting their research. The authors thank the Sydney Cord Blood Bank (Australia) for providing UCB for our research. The authors also thank Inner Wheel Foundation for financial support.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.