
Abstract
Stem cell therapies for tympanic membrane repair have shown initial experimental success using mesenchymal stem cells in rat models to promote healing; however, the mechanisms providing this benefit are not known. We investigated in vitro the paracrine effects of human adipose-derived stem cells (ADSCs) on wound healing mechanisms for human tympanic membrane-derived keratinocytes (hTM) and immortalized human keratinocytes (HaCaT). ADSC conditioned media (CMADSC) were assessed for paracrine activity on keratinocyte proliferation and migration, with hypoxic conditions for ADSC culture used to generate contrasting effects on cytokine gene expression. Keratinocytes cultured in CMADSC showed a significant increase in cell number compared to serum-free cultures and further significant increases in hypoxic CMADSC. Assessment of ADSC gene expression on a cytokine array showed a range of wound healing cytokines expressed and under stringent hypoxic and serum-free conditions was upregulated (VEGF A, MMP9, Tissue Factor, PAI-1) or downregulated (CXCL5, CCL7, TNF-α). Several of these may contribute to the activity of conditioned media on the keratinocytes with potential applications in TM perforation repair. VEGFA protein was confirmed by immunoassay to be increased in conditioned media. Together with gene regulation associated with hypoxia in ADSCs, this study has provided several strong leads for a stem cell–derived approach to TM wound healing.
Introduction
T
One avenue relatively unexplored is stem cell stimulated wound healing. One of the few reports of stem cell healing for TM is the delivery of human mesenchymal stem cells (MSCs) into the middle ear, which promoted healing of acute and chronic perforations in rat ears [8]. Recent work showed that delivery of mouse bone marrow MSC on hyaluronic acid scaffold to perforated mouse tympanic membrane was able to promote healing of acute perforation [9] compared to scaffold alone. The molecular mechanism of stem cell mediated healing was however not discovered in either study. MSCs applied in other wound healing models have been shown to produce mediators that enhance wound healing locally through improved cell proliferation and viability [10,11], tissue remodeling by reepithelialization [8,12 –16], angiogenesis [12,14,15,17,18], and inflammation [14,19], in a paracrine manner. These paracrine mediators may ultimately be able to reactivate the wound healing cascade in chronic wound healing [20,21].
We propose that paracrine mechanisms might prove useful to promote wound healing in TM perforations and explain some of the wound healing activity derived from MSC in rats. In the present study, we evaluated in vitro models for paracrine activity from human adipose-derived MSCs on human tympanic membrane keratinocytes and the HaCaT keratinocyte cell line. Hypoxic conditioning of the MSC was an effective stimulator of proliferative and chemotactic activity. We further studied the effect of hypoxic conditioning on the MSC wound healing transcriptome and identified several candidate factors for future study.
Materials and Methods
Culture of primary hTM keratinocytes and HaCaT cells
Primary hTM keratinocytes derived from normal human tympanic membrane explants were used as previously described [22]. The St John of God HealthCare Ethics Committee approved collection of excess tympanic membrane tissue from patients undergoing otological procedures at St John of God Hospital (Subiaco, Australia). The HaCaT cell line was supplied by the Burns Injury Research Unit, School of Surgery, University of Western Australia. Both cell types were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 4.5 g/L
Culture of adipose-derived stem cell
Human adipose-derived stem cells (ADSCs; Lonza) were cultured in DMEM containing 1 g/L
Collection of ADSC-conditioned medium
ADSCs were cultured to 80%–90% confluence in 75 cm2 tissue culture flasks, rinsed with 1 × phosphate-buffered saline (PBS), pH 7.2 (Gibco), and incubated with 10 mL of serum-free (S−) DMEM (1 g/L
Total RNA isolation and complementary DNA synthesis
Total RNA was extracted from ADSCs and hTM keratinocytes according to manufacturer's instructions (FavorPrep™ Tissue Total RNA Mini Kit; Favorgen Biotech Corp), then RNA concentration and purity were measured by spectrophotometry (Epoch Take3; BioTek Instruments, Inc.). Synthesis of complementary DNA (cDNA) was then performed (RT2 First Strand Kit; Qiagen) according to manufacturer's instructions with 0.5 μg of RNA.
Real-time polymerase chain reaction of human wound healing array
Human Wound Healing polymerase chain reaction (PCR) array was performed according to manufacturer's instruction (RT2 Profiler; Qiagen) on the cDNA of ADSCHx, ADSCNx, and hTM keratinocyte S−. Briefly, cDNA, SYBR green master mix, and water were combined to a final volume of 2,700 μL. A volume of 25 μL per well was added to the 96-well array plate and run on a CFX Connect™ Real-Time PCR Detection System (Bio-Rad) under the following conditions: 95°C for 10 min to activate HotStart DNA Taq Polymerase and 40 cycles of denaturation at 95°C for 15 s and annealing at 60°C for 1 min. Threshold cycles (Ct) were determined using the exponential growth phase and the baseline signal from fluorescence versus cycle number plots. Relative messenger RNA (mRNA) expression levels were determined using the Livak method (2−ΔΔCt) with beta-2-microglobulin (B2M) selected as the most stable housekeeping gene. Each experiment was performed in duplicate array plates repeated for all ADSCHx, ADSCNx, and hTM keratinocyte S−.
Gene ontology analysis of hypoxia-induced gene expression changes in cultured ADSC
Gene ontology analysis was performed using the g:Profiler platform for enrichment analyses to identify biological functions affected by hypoxic conditions in cultured ADSC (
Total protein assay
Total protein concentration in both Hx and Nx CMADSC were quantified using a Bradford Protein Assay (Quick Start; Bio-Rad) according to manufacturer's protocol. Bovine Serum Albumin standards were used over a range of 1.25–10 μg/mL. After incubating the CMADSC with dye reagent, the absorbance was measured at 595 nm using a spectrophotometer (Epoch; BioTek Instruments, Inc.) and protein concentrations of the CMADSC read from the standard curve.
Quantification of specific protein levels
Secreted VEGF-A, IL6, IL1B, FGF2, HB-EGF, and FGF7 proteins in CMADSC were quantified using enzyme-linked immunosorbent assay (ELISA, Quantikine kits; R&D Systems) according to the manufacturer's protocol. Briefly, protein standard curves were prepared by serial dilution. In duplicates, samples were loaded into microplate wells precoated with monoclonal antibodies. After incubation for 2–3 h, wells were washed to remove unbound proteins and then enzyme-linked polyclonal antibody was added to the wells and incubated. Final washing was done to remove unbound antibody-enzyme reagent, then substrate containing hydrogen peroxide and tetramethylbenzidine was added and incubated for 30 min before stopping the reaction by acidification. Optical density was measured at 450 nm on the spectrophotometer with a wavelength correction at 570 nm. Secreted protein concentrations were expressed in ng per mg of total protein, calculated from either linear standard curves or four parameter logistic (4-PL) curve fit by interpolating sample values.
MTS assay for proliferation
Cell proliferation was analyzed by colorimetric assay (CellTiter 96® AQueous One; Promega Corporation). Briefly, keratinocytes were serum-starved for 8 h before experimentation, resuspended in test medium [5,000–10,000 cells in 10% serum (S+), serum free (S−), or 50:50 mix of conditioned media and S− media], and seeded in 96-well plates for a 24-h or 48-h incubation at 37°C, 5% CO2. Following incubation, 20 μL of colorimetric solution was added to each well, and plates were incubated for 2 h. Absorbance was measured at 490 nm wavelength using the Epoch colorimetric plate reader (BioTek Instruments, Inc.). Samples were read in triplicate, averaged, and corrected for background optical density. The effect on proliferation was expressed as a fraction of the S− control treatment for each cell, respectively.
Scratch test assay for migration
A scratch wound assay was performed on hTM keratinocytes and HaCaT cells to assess cell migration when cultured in Hx CMADSC, Nx CMADSC, and S+. Initially, 500,000 keratinocytes were seeded in 12-well culture plates with S+ media and grown to confluence. Following 8 h preincubation in S− medium, cell monolayers were wounded with a sterile 200 μL pipette tip with light vacuum applied to clear debris from the wound. Cells were then washed with PBS and incubated with the corresponding test medium. Microscopic images were taken immediately after scratch wounding (t0) and after 10 h on an Olympus BX60 microscope with a DP70 digital camera (Olympus). Wound gaps were measured using ImageJ (1.48v) software. Each gap was measured 45 times. All measurements were repeated in triplicate and averaged. The migration of keratinocytes into the gap was calculated as:
The results are expressed as a fraction of the S− control treatment as above.
Statistics
Quantitative data are presented as mean ± standard error of the mean (SEM). Microsoft Excel was used for all data analysis. Statistical methods used include: • One-way analysis of variance (ANOVA) for proliferation and migration experiments • Dunnett's test as a post hoc test for individual treatment group comparisons against S− control group for both proliferation and migration experiments • Student t-test for ELISA and PCR data analysis
Statistical level of confidence was set at P < 0.05 unless otherwise stated.
Results
Cell proliferation and migration assays were performed using Hx and Nx CMADSC on hTM and HaCaT keratinocytes with serum-containing (S+) and serum-free (S−) media as positive and negative controls. Figure 1 shows that ADSC cultures were viable before and after conditioning in hypoxic or normoxic conditions.
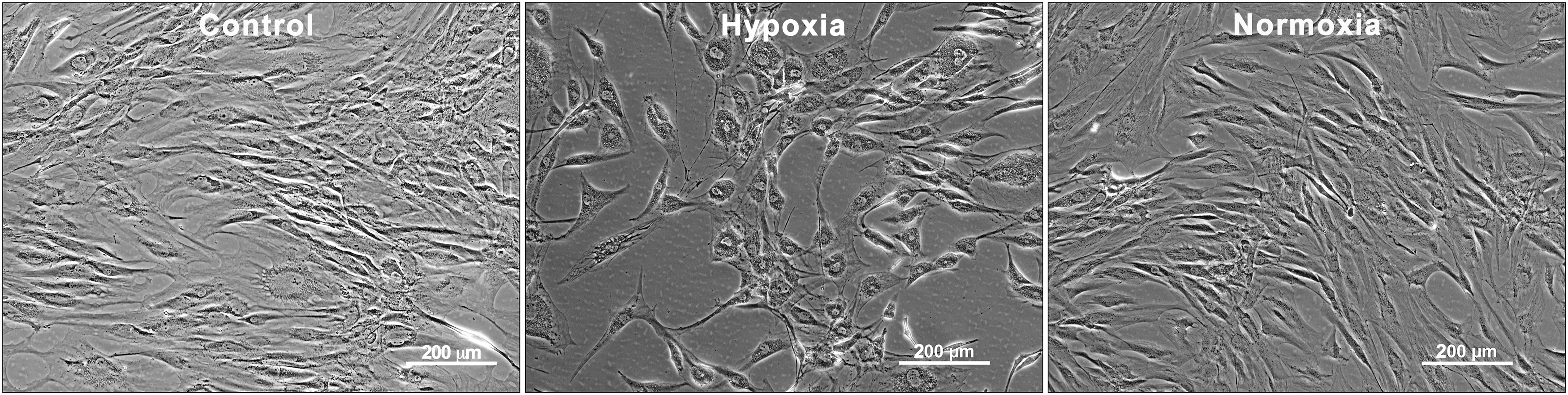
Human ADSC before and after hypoxic and normoxic conditioning for 48 h. Scale bars = 200 μm. ADSC, adipose-derived stem cell.
Effect of CMADSC on keratinocyte proliferation
Representative images of proliferation responses in hTM and HaCaT keratinocytes over 48 h are shown in Fig. 2. In hTM keratinocyte cultures, serum produced a substantial proliferative response at 24 and 48 h compared to serum-free cultures (Fig. 3). Hx CMADSC media produced a significantly greater proliferative response (P < 0.05) than serum-free media and quantitatively as much as 83% and 95% of the serum response at 24 and 48 h, respectively. Nx CMADSC however only showed significant (P < 0.05) proliferative response at 48 h, up to 79% of the serum response. When comparing Hx and Nx CMADSC, there was a significant difference (P < 0.05) between the proliferative effects on hTM keratinocytes at both 24 and 48 h.
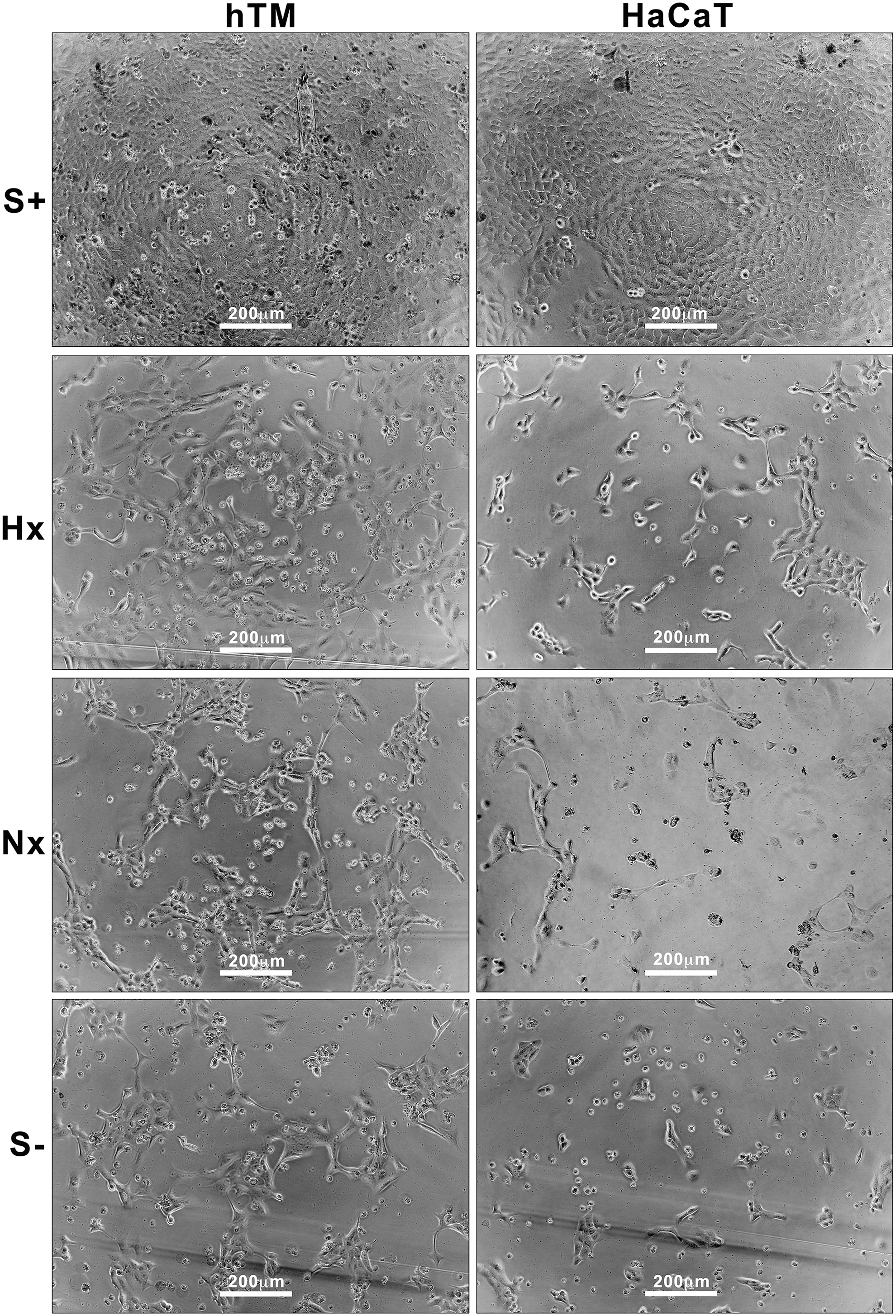
Left columns: hTMt in test medium (S+, Hx and Nx CMADSC, S−) during proliferation test at 48 h; right columns: HaCaTs in test medium during proliferation test at 48 h. Scale bars = 200 μm. CMADSC, ADSC conditioned media.
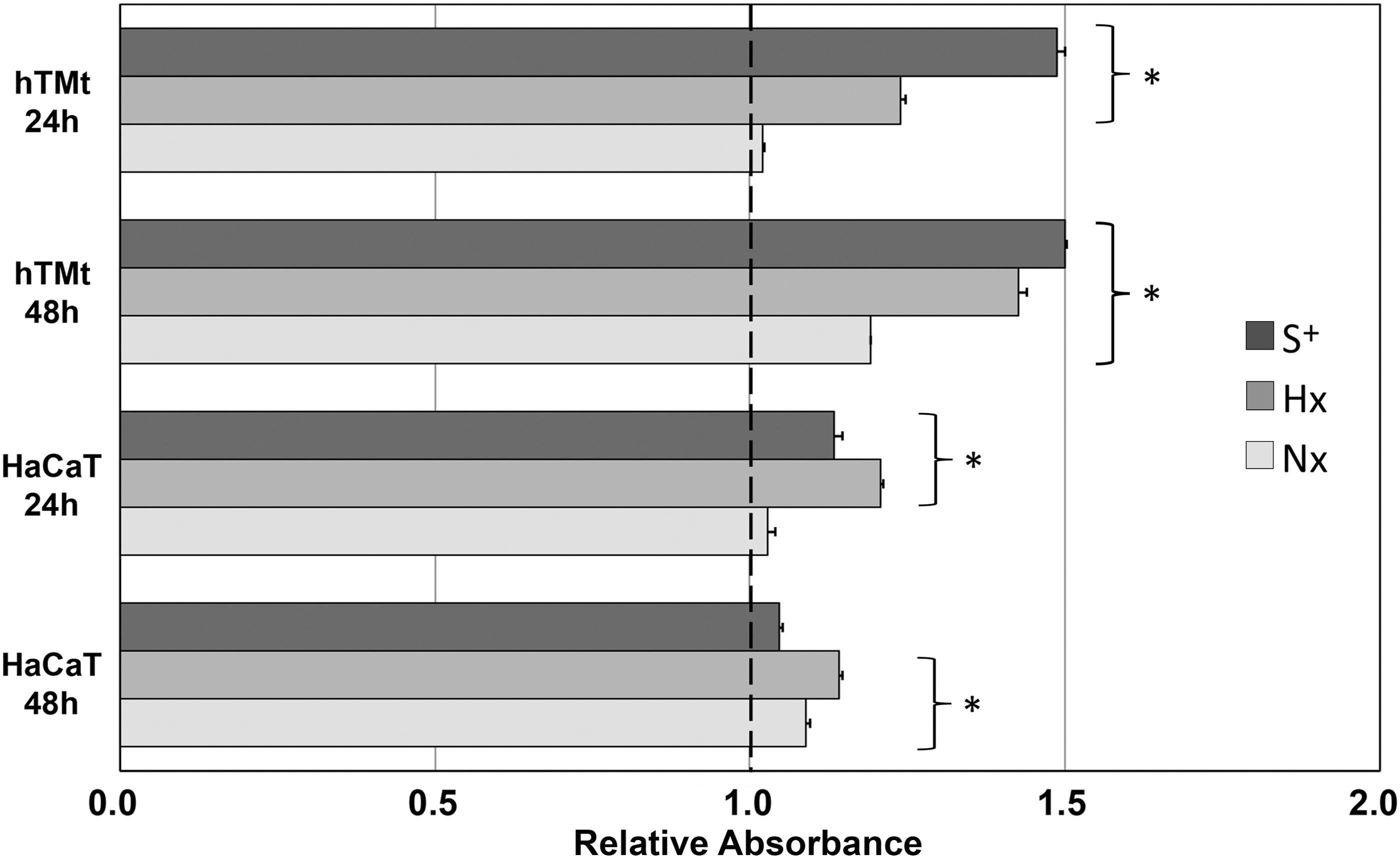
Effect of S+, Hx and Nx, CMADSC on the proliferation of hTM and HaCaT keratinocytes over 24 and 48 h. Graph shows representative data of relative absorbance values normalized against the negative (S−) control. hTM keratinocytes and HaCaT keratinocytes had a greater proliferative response to Hx CMADSC than Nx CMADSC. Test media marked with asterisk (*) were significantly different to S− media (P < 0.05).
In HaCaT keratinocytes, the proliferative effect of serum was significant (P < 0.05) at 24 h, but not at 48 h compared to serum-free controls (Fig. 3). Proliferative responses of HaCaT keratinocytes to Hx CMADSC was significant at both 24 and 48 h, quantitated at 106% and 109% of serum response, respectively. Nx CMADSC treatments were not significant at 24 h, but at 48 h responses were significantly greater than serum-free controls of up to 9% compared to serum-free controls. Both CMADSC treatments produced a proliferative response larger than the serum treatment effect at 48 h. When comparing Hx and Nx CMADSC, there was a significant difference (P < 0.05) between the proliferative effects on HaCaT at both 24 and 48 h.
Effect of CMADSC on keratinocyte migration
Migration assays were performed on hTM and HaCaT keratinocytes over 10 h to compare the effect of Hx and Nx CMADSC measured against serum-free control media (Figs. 4 and 5). Representative images of wound gaps in all test media are presented in Fig. 4, and the quantitative data are presented as the normalized percentage gap closure in Fig. 5.
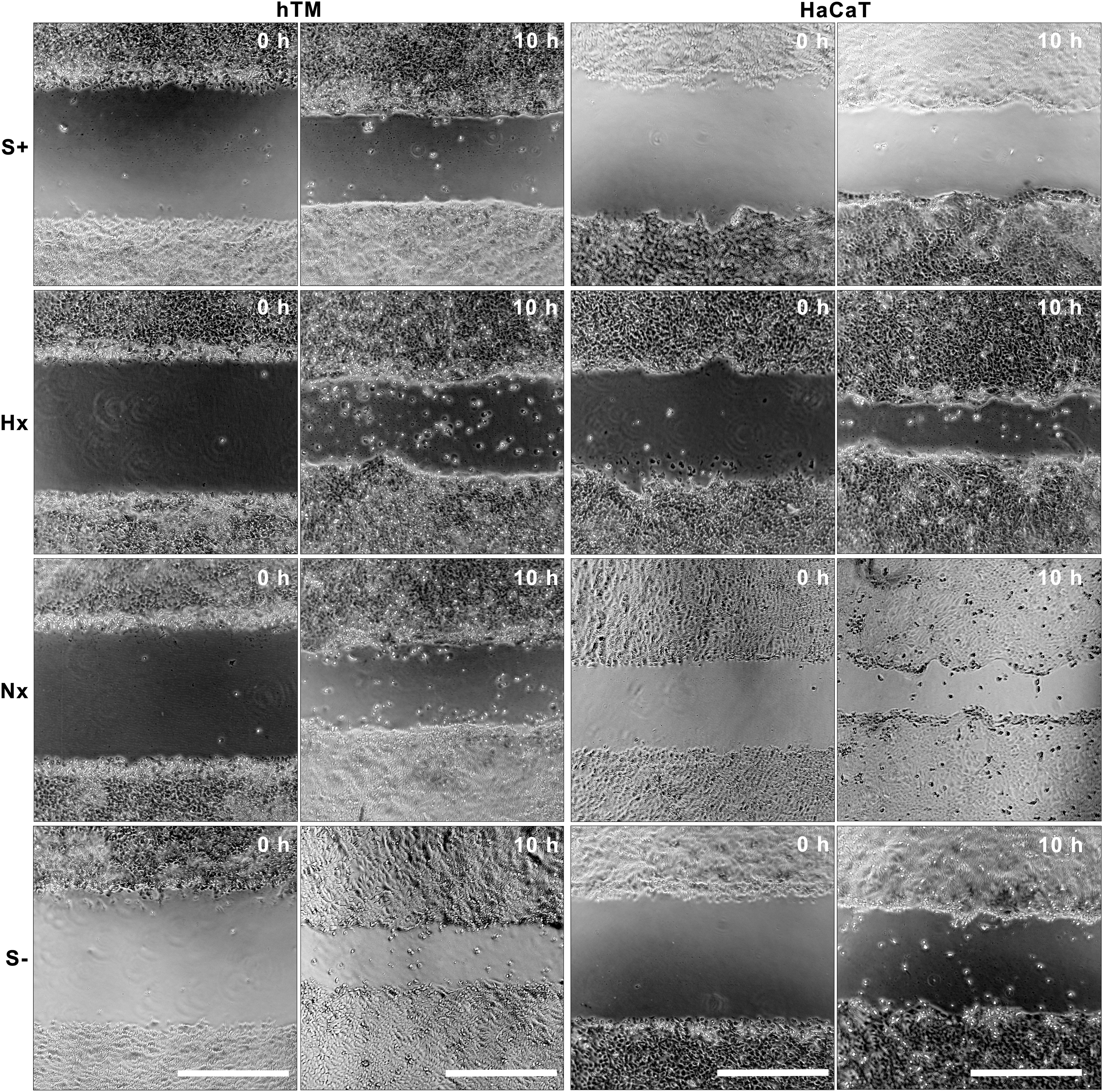
Representative image of migration test over 10 h; left two columns: hTM keratinocyte wound gaps in various test medium; right two columns: HaCaT keratinocyte wound gaps in various test medium. Scale bars = 500 μm.
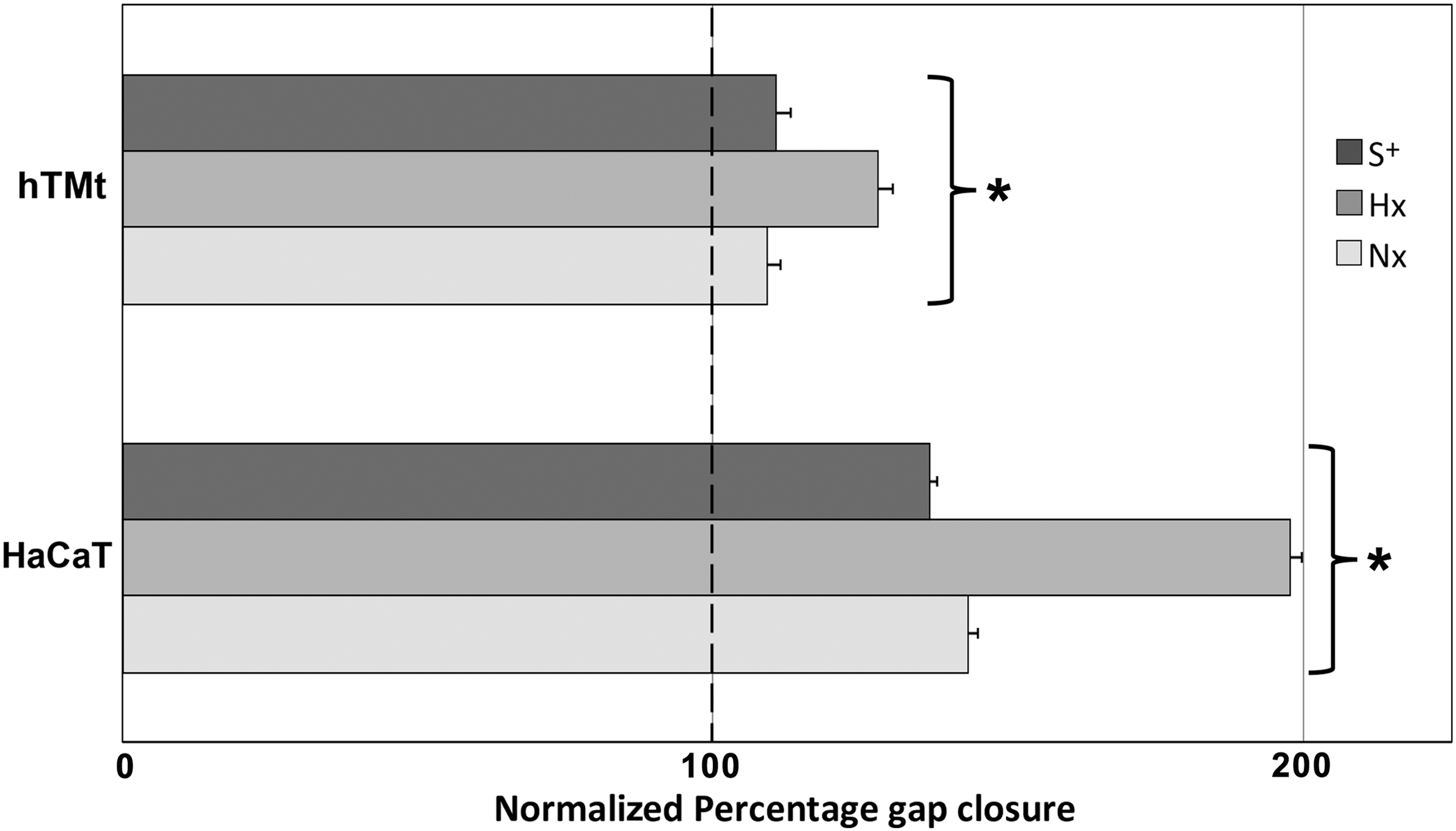
Effect of S+, Hx and Nx, CMADSC on the migration of hTM and HaCaT keratinocytes over 10 h. Migration data over 10 h are represented as normalized percentage gap closure of negative control (S−) wells (at 100%). All test media marked with an asterisk (*) were significantly different (P < 0.05) from the corresponding S− controls.
Hx CMADSC and Nx CMADSC produced a significantly greater (P < 0.05) gap closure effect on hTM keratinocytes compared to S− media controls. Hx CMADSC effects on hTM keratinocytes were significantly higher than Nx CMADSC (P < 0.05). Gap closures were on average 27% and 10%, respectively, greater than in the absence of paracrine activity. In these experiments, hTM keratinocytes were found to maintain a migration capacity in the absence of serum (P < 0.05).
In HaCaT keratinocytes, significant differences in migration were observed for both Hx and Nx CMADSC (P < 0.05). Gap closures were on average 37% in serum controls, 98% in Hx CMADSC and 43% in Nx CMADSC greater than in the S− media controls. HaCaT cells cultured in S− media migrated less well and maintained a 35% gap even after 28 h, while all other test wells were closed at this time.
Molecular identification of paracrine activity
PCR array for wound healing gene expression in ADSCs and hTM keratinocytes
PCR Wound Healing arrays were used to determine which of the 84 genes present on the array were expressed in ADSCs, as well as to perform a quantitative comparison of the effects of hypoxic and normoxic culture conditions. B2M was a stable housekeeping gene for these arrays, so was used to normalize all values, and Ct thresholds were manually set to the same value across all samples.
Gene expression
Among the 84 genes present on the array (Supplementary Table S1; Supplementary Data are available online at
B2M, beta-2-microglobulin.
Increase fold Δ >2-folds
Not regulated by hypoxia
Decrease fold Δ <2-folds
Housekeeping Gene
Not expressed
Bold text shows P < 0.05.
Expression of wound healing genes in hTM keratinocytes was also examined in the same manner, and a largely similar set was found (Table 2), but with notable differences being absence of gene expression in keratinocytes for cell adhesion molecule integrin β3, collagens alpha 1A2, 4A1, 4A3, 5A1, 5A3, and 14A1, WISP1 and WNT5A, and presence of gene expression for integrin β6, TGFα, E-cadherin, MMP7, and TNF.
Hx ADSC, Nx ADSC, and hTM keratinocytes expressed (+) or not expressed (−) where highlighted boxes indicate variations and bold represents secreted genes.
ADSC, adipose-derived stem cell.
Hypoxia in ADSCs
Thirty-eight expressed genes were upregulated (2- to 62-fold) by hypoxia in ADSCs, 12 genes were downregulated (2- to 5-fold), and 17 were expressed, but showed less than twofold difference between the culture conditions. Taking into consideration that we were looking for secreted proteins, genes encoding for nonsecreted proteins were removed from the list of comparison between Hx and Nx ADSCs (Table 1). Genes not regulated by hypoxia, hence not likely related to the hypoxia-induced growth responses, included prominent keratinocyte growth regulators HB-EGF and FGF7. FGF2 was only 1.9-fold higher in Hx. Genes reduced by hypoxia were chemokines and cytokines, including prominently the keratinocyte growth factor TNFα.
Gene expression regulated by hypoxia
Tissue Factor (F3), showed the highest upregulation in this assay with more than 60-fold higher expression in Hx ADSCs than Nx ADSCs. Out of 56 genes coding for secreted proteins, 24 were upregulated by at least twofold; 13 showed similar expression levels; 7 were downregulated between two- to sixfold; and 12 genes did not meet the threshold of detection (Ct values >35 or blank) in the array.
Gene ontology analysis
The results are summarized in Table 3. For a list of regulated genes and all significant gene ontology terms, see Supplementary Figure S1. The genes upregulated by hypoxic conditions are rich in collagens IV and V particularly and, thus, related to basement membrane and fibrillar collagen formation in the extracellular matrix. Among the downregulated genes are multiple chemokine (C-X-C) ligands, showing that ADSC influenced by hypoxia may have a direct impact on the chemotaxis of leukocytes.
Organism: Homo sapiens; query list of significantly up- (19) and downregulated (7) genes; options: significant only, ordered query, no electronic gene ontology annotations, gene list as a statistical background: all genes in the RT2 Profiler PCR array that were expressed by ADSC (69 genes); hierarchical filtering: best per parent; gene ontology analyses; significance threshold: Benjamini–Hochberg FDR corrected P value (P < 0.0005).
BP, biological process; CC, cellular component; FDR, false discovery rate; MF, molecular function; PCR, polymerase chain reaction.
Protein concentration in conditioned media
To determine whether the hypoxia-mediated increase in gene expression of VEGFA, IL6, and IL-1B from the PCR array results was translated into increased protein secretion, CMADSC were analyzed using ELISA. FGF2, HB-EGF, and FGF7 were included in the analysis due to their potential involvement in TM wound healing mechanism. VEGFA, IL6, IL-1B, FGF2, and FGF7 were all observed at detectable levels in CMADSC. HB-EGF was not detected. Figure 6 shows the relative fold change in specific protein levels in Hx CMADSC against Nx CMADSC. VEGFA protein level was sixfold higher in Hx CMADSC than in Nx CMADSC (P < 0.05), while IL6 and IL-1B were not significantly higher (P > 0.05).
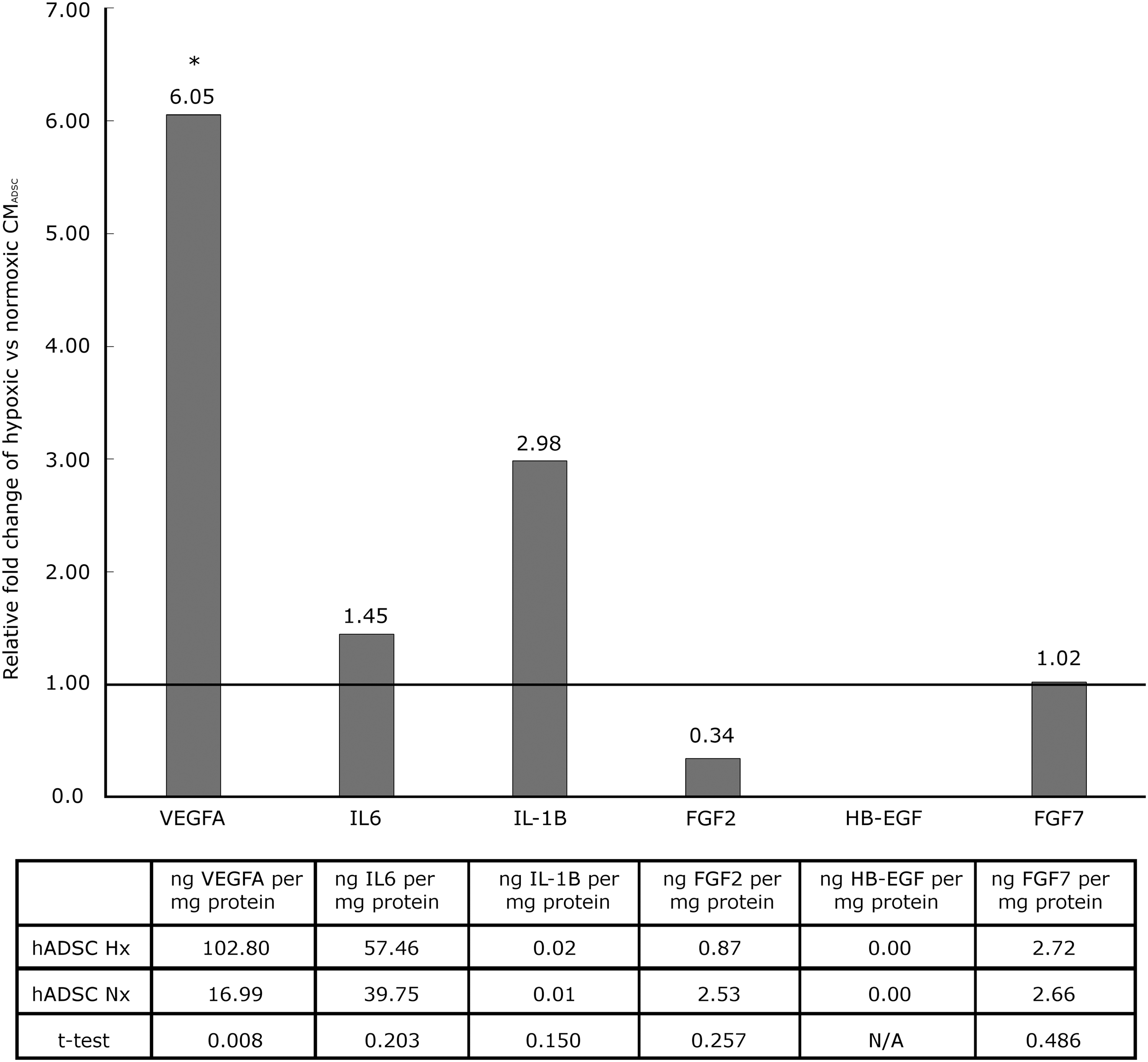
Relative fold change of secreted VEGFA, IL6, IL-1B, FGF2, HB-EGF, and FGF7 in hypoxic and normoxic conditioned media after 48 h culture of ADSC (*P ≤ 0.05).
Discussion
The major finding of this study was that ADSCs produced a paracrine activity to promote wound healing mechanisms in hTM and HaCaT keratinocytes in vitro. Conditioned media from hypoxic ADSC cultures promoted further increase in both proliferation and migration of keratinocyte cell lines compared to normoxic conditioned media. To find active candidates for this paracrine effect, we identified which wound healing genes coding for secreted proteins were expressed by ADSC and were regulated by hypoxic culture conditions. We were also able to exclude some candidates based on a lack of gene expression, or no change with hypoxia. This study is the first to show that ADSC-secreted VEGF might influence keratinocyte growth and wound healing in a paracrine manner and could influence MSC therapy at the tympanic membrane.
ADSCs in TM wound healing and keratinocytes
Recent studies show that human bone marrow-derived MSCs promote healing of chronic TM perforation in rats [8] and that mouse bone marrow-derived MSCs promote healing of acute TM perforation in mice [9]. Neither study identified the mechanism of these effects. In this study, we show for the first time an improved in vitro wound healing of hTM keratinocytes using adipose-derived MSC paracrine activity. These data provide a plausible mechanism for the in vivo findings in rodents and support further investigation into the therapeutic potential of conditioned media or growth factor therapy, rather than the logistically more difficult cell implantation therapy.
Based on the wound-healing literature, keratinocyte proliferation and migration can be regulated by exposure to bioactive molecules from exogenous, paracrine, or autocrine sources. Factors involved include EGF, TGFα, TGFβ1, HB-EGF, FGF2, KGF, IL6, and VEGF A. [16,24 –27]. In TM perforations, however, the molecular mechanisms regulating wound healing are less well studied [5,8,28], and the search for an optimal growth promoting activity is ongoing.
Response to normoxic and hypoxic CMADSC
In the present study, ADSCs in a normoxic environment enhanced proliferation and migration in keratinocytes consistent with previous studies on epithelial cells such as skin keratinocytes [29 –32] or endothelial cells [32 –34] and on nonepithelial cells like fibroblasts [16,33]. Conditioned media from hypoxic culture conditions produced a greater regenerative response in hTM and HaCaT keratinocytes, suggesting that in hypoxia ADSCs secreted more or different paracrine factors to that in normoxia. A hypoxic environment stimulates greater production of some paracrine factors from MSC [34] through activation of hypoxia inducible factor alpha (HIF-1α) transcription factor [2,35 –38]. MSCs using glycolysis rather than mitochondrial respiration in hypoxic environments [39] increase their expression of paracrine factors [40 –42]. The migration response in both hTM and HaCaT keratinocytes to Hx CMADSC was greater than responses to serum, indicating a high degree of responsiveness and specificity for chemotactic mechanisms by the paracrine mediators.
Identity of the paracrine activity from ADSCs
From a wound healing PCR array on ADSCs we identified gene expression of secreted paracrine factors and found genes expressed differently in Hx ADSCs versus Nx ADSCs. Eleven genes expressed but not regulated by hypoxia were set aside as not likely to explain the changes in paracrine activity seen in hypoxia. Among the genes for secreted proteins, 12 were not expressed, 24 were upregulated, and 9 were downregulated in Hx ADSCs. These are all candidates worthy of further consideration, as the differential expression in genes could be associated with the difference in paracrine activity secreted. (Table 4).
Several important keratinocyte growth regulators from the literature such as TGFα [24] and EGF were not expressed by ADSC. HB-EGF, another important regulator of TM wound healing [43,44], was expressed but not regulated significantly by hypoxia, and secreted protein was not detected. FGF-2, also used to promote TM wound healing in other studies [43,45 –47], was expressed and secreted into the CMADSC, but was not regulated by hypoxia in our study. These data suggest that there is significant growth activity from factors other than those already presented in the TM repair literature. Other promoters of keratinocyte growth such as VEGFA and IL6 were upregulated by hypoxia, and secreted VEGF was increased most and so seems the most likely candidate to increase keratinocyte growth response (Table 4). The other cytokines we tested showed no significant effects of hypoxia on protein levels accumulated, so their influence remains uncertain and will require further study.
Consistent with previous findings [17,19,40,48], VEGFA protein levels in CMADSC were significantly increased by hypoxia. Of note in this study, VEGFA had been shown to be upregulated in traumatic TM perforations, in both epithelial and fibrous layers of the TM, and was reduced in nonhealing TM [49]. This could represent a novel finding for the importance of VEGFA in the healing of TM perforations and to improve healing by regulating the production of VEGFA and other factors.
The gene enrichment analysis is another approach that may provide insight; it showed a downregulation of neutrophil chemotaxis, and neutrophils are an important cell type in the inflammation phase of wound healing [50]. HIF-1α has been shown to reduce the recruitment of neutrophils in vitro through nestin-1 induction [51] and similar mechanisms may be apparent in keratinocytes. Our finding of ADSC regulated chemokines during hypoxia indicates that the paracrine effect may be directed toward regeneration through tissue repair mechanisms and reduction of inflammation.
Comparison of these data with TM wound healing microarray [52] may provide further context for these genes; however, further experiments such as proteomic analysis of the conditioned media or PCR arrays on keratinocytes posttreatment will be required to support the PCR and ELISA data. A broader transcriptome/proteome approach would also be valuable, particularly together with the rat TM wound healing transcriptome data already available [53] and our limited data set from human TM keratinocyte gene expression array. Once a set of candidates is refined, receptor inhibitors or recombinant proteins could be used to target specific pathways in vitro or in vivo.
Practical applications of this finding include cell-free conditioned media approaches by topical application in vivo or combining the mediator(s) with scaffolds to promote wound healing in a chronic TM perforation [46]. These data may also support use of fat plug myringoplasty [54,55], where direct benefits might come from ADSC present in the graft. The paracrine activity identified in this study requires further characterization and optimization regarding which tissue MSCs to use, what conditions might optimize the activity, and what is the nature of the molecular entities producing the activity. Both adipose- and bone marrow-derived MSC produce cytokines and growth factors. Adipose tissue can be a preferred source for MSC therapies, due to their low risk in isolation, high cell yields, and ready availability in most individuals.
Conclusion
In summary, this study has identified a wound healing activity from ADSCs that promotes growth and migration of tympanic membrane keratinocytes. The activity was regulated by hypoxia and was associated with expression of a number of secreted proteins that could be major effectors of keratinocyte proliferation and migration. These findings not only revealed the paracrine effects of ADSCs potentially affecting TM keratinocyte growth but also encourage further investigations for stem cells or their conditioned media as possible therapeutics for chronic TM wound healing.
Footnotes
Acknowledgments
The HaCaT cell line was kindly supplied by the Burns Injury Research Unit (School of Surgery, University of Western Australia). The work was funded, in part, by grants from the Garnett Passe and Rodney Williams Memorial Foundation (R.J.D.), National Health and Medical Research Council (R.J.D., M.D.A.), the Gift of Hearing (R.J.M), and Raine Foundation (M.v.U.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.