
Abstract
The life-long story of the heart starts concomitantly with primary differentiation events occurring in multipotent progenitors located in the so-called heart tube. This initially tubular structure starts a looping process, which leads to formation of the final four-chambered heart with a primary contribution of geometric and position-associated cell sensing. While this establishes the correct patterning of the final cardiac structure, it also provides feedbacks to fundamental cellular machineries controlling proliferation and differentiation, thus ensuring a coordinated restriction of cell growth and a myocyte terminal differentiation. Novel evidences provided by embryological and cell engineering studies have clarified the relevance of mechanics-supported position sensing for the correct recognition of cell fate inside developing embryos and multicellular aggregates. One of the main components of this pathway, the Hippo-dependent signal transduction machinery, is responsible for cell mechanics intracellular transduction with important consequences for gene transcription and cell growth control. Being the Hippo pathway also directly connected to stress responses and altered metabolism, it is tempting to speculate that permanent alterations of mechanosensing may account for modifying self-renewal control in tissue homeostasis. In the present contribution, we translate these concepts to the aging process and the failing of the human heart, two pathophysiologic conditions that are strongly affected by stress responses and altered metabolism.
Spatial Control, and Mechanotransduction: Multimodal Regulation of Cell Social Relationships
M
The basics in cell mechanotransduction: the control of social cellular behavior in multicellular contexts
The architecture of multicellular development is under the control of pathways not only related (as thought until now) to mutual chemical signaling by the cells, but also to information depending, in large part, on their spatial or “social” relationships. For example, when considering the primary differentiation events occurring during the early cleavage stages in the mammalian embryo, the initial patterning of cell division is realized by a nonpreferential orientation of the division axes, resulting in an initially spherical geometry [7]. This organization of the cleavage progression leads, in a short time, to an irreversible determination of the blastomere fate, which essentially depends on their positioning, “more inside” or “more outside” of the initial sphere. This phenomenon, in early times called “polarization” of the blastomeres, started to be easily detected more than 30 years ago by staining the extracellular membrane with fluorescent lectins [8]. Due to the lack of a spatial template allowing a predetermination of cell fate resulting from segregation of asymmetrically distributed cytoplasmic determinants (this is the typical situation in invertebrates like Drosophila or Caenorhabditis elegans), the cells in the growing embryo need, in a relatively short time, to take fundamental decisions whether to: (i) divide symmetrically and give rise to two ancestors of the inner pluripotent cells, (ii) divide symmetrically to produce two outer trophoblast cells, or (iii) divide asymmetrically to give rise to one pluripotent cell inside and one trophoblast cells outside. Recent observations suggest that, although these divisions are highly spatially coordinated, they occur with an intrinsically stochastic modality; despite this, they lead to a correct numbering of inner and outer blastomeres, which become restricted to give rise to the pluripotent “inner” cell mass or enter into the first lineage determination resulting in the formation of the “outer” trophoblast [7,9,10].
The mechanism underlying blastomere determination has remained unsolved until few years ago, when the concept of cell mechanics as a potent inducer of (stem) cell determination in a “stand-alone” manner has started to be understood. Specifically, we refer to the strict relationship between cell fate and either the localization of cells into a growing pattern [11,12] or the geometric constraints that a given cell receives, for example, by virtue of tissue architecture and matrix mechanics [13 –15]. The definition of geometric criteria to achieve controlled dynamics of stem cell fate has been, thus, introduced to operate a more controlled cell differentiation modality into conditions promoting niche growth and self-renewal [16,17], to explain the systematics of pathologic transformation [18], or to understand the functional adaptation of stem cells to niche perturbing conditions [19]. This advancement, enabled by the evolution of systems to engineer cell culture environments in two dimensions (2D) or three dimensions (3D), has made it possible to culture cells under architectural and/or spatially regulated signaling, a task that was impossible to be realized with conventional cell culture systems [20,21]. In this way, mechanical forces have definitively entered cell biology with the identification of molecular pathways linking the architectural or the extracellular environment compliance to signal transduction and (epi)genetic programming. This has established a novel and universal vision of the way cells communicate with neighbors and receive short-range information, which, combined with humoral and chemical cues, results in controlled organ growth dynamics, tissue repair, and regeneration by finely modulating cellular plasticity [19], and, as recently discussed by us [5], cardiovascular tissue pathology.
Intracellular control of cell mechanosensing
How is the intracellular machinery connected to the cytoskeleton and the extracellular attachment domains to achieve a highly tunable regulation of cellular physiology? Integrins, G protein-coupled receptors (GPCR), protein tyrosine kinases, stretch-activated ion channels, nuclear factor of activated T cells (NFAT), and, more recently, the cytoskeleton and the nucleoskeleton have been identified as protein transducers able to convert mechanical cues into biochemical responses (Fig. 1). Integrins are heterodimeric proteins consisting of α and β subunits that mediate interactions between cells and either the extracellular matrix or plasma membrane protein ligands and were first identified almost 30 years ago [22]. These proteins are key bidirectional regulators of cell adhesion that mediate both outside-in and inside-out signaling. More than 50 adaptor proteins have been described that mediate the interaction between integrins and the actin microfilaments (reviewed in [23]). Talin, kindlins, vinculin, paxillin, α-actinin, and filamin are organized, following cell adhesion, into focal adhesions, which promote the interaction between integrins and the actin cytoskeleton and regulate signaling molecules Focal Adhesion Kinase (FAK) and Src [23]. Integrin clustering leads to the recruitment of FAK to focal adhesion complexes, followed by its phosphorylation at multiple sites, the major of which is Tyr397. This latter is a docking site for SH2-containing proteins (including Src, phosphatidylinositol 3-kinase, phospholipase Cγ, and Grb7 [23]). Furthermore, FAK interacts with two regulators of small GTP binding proteins (ie, GRAF, a GTPase activating protein for Rho, and ASAP1, a GTP exchange factor for Arf-1/6) through their SH3 domain. Activated Src phosphorylates Cas, an event that has been linked to the transduction of extracellular matrix stiffness into mechanosensitive cell cycling [24]. The FAK-Src complex is also a critical regulator of both small GTP binding proteins, such as Rho, Rac, and Cdc42, which are critical regulators of the actin dynamics [25], and the ERK/MAPK pathway through either RAP1 or Grb2 [26]. With regard to the role played by GPCR and stretch-activated ion channels (SAC) in mechanotransduction, a substantial body of literature has demonstrated the involvement of these receptors in mediating both the response of endothelial cells to shear stress and the myogenic response of smooth muscle cells to stretching, which is responsible for the intrinsic mechanism of regulation of flow (reviewed in ref. [27]). Briefly, membrane depolarization, through the activation of SAC, is required to regulate the myogenic tone. The exact nature of the SAC expressed by smooth muscle cells is still disputed, but may include the transient receptor potential (TRP) channel and the epithelial Na+ channel [27]. A similar role has been shown for SAC (ie, a K+ channel, TRP channels, and P2X4 channels) in transducing the effect of shear stress on endothelial cells (reviewed in ref. [28]). In smooth muscle cells, mechanical loading induces the local generation and secretion of peptides, lipids, nucleotides, and amines that act on GPCR. However, both these latter (eg, Angiotensin II type 1 receptors or bradykinin B2 GPCR) and receptor tyrosine kinases (eg, VEGFR and Tie2) may be activated directly by mechanical stimuli, in an agonist-independent manner [27,28]. With regard to NFAT, recent studies have demonstrated an important role for polycystins (PC) as mechanosensor molecules [29,30]. PC constitute a protein family that comprises eight family members, among which PC1 and PC2 are the most studied. PC1 and PC2 interact at the primary cilia (an antenna-like protrusion that functions as a sensory organelle [31]) and form a mechanosensitive Ca2+ channel [30]. Recent data have shown in human periodontal ligament cells that mechanical stimulation and PC-mediated calcium influx activate the calcineurin/NFAT signaling cascade [32].
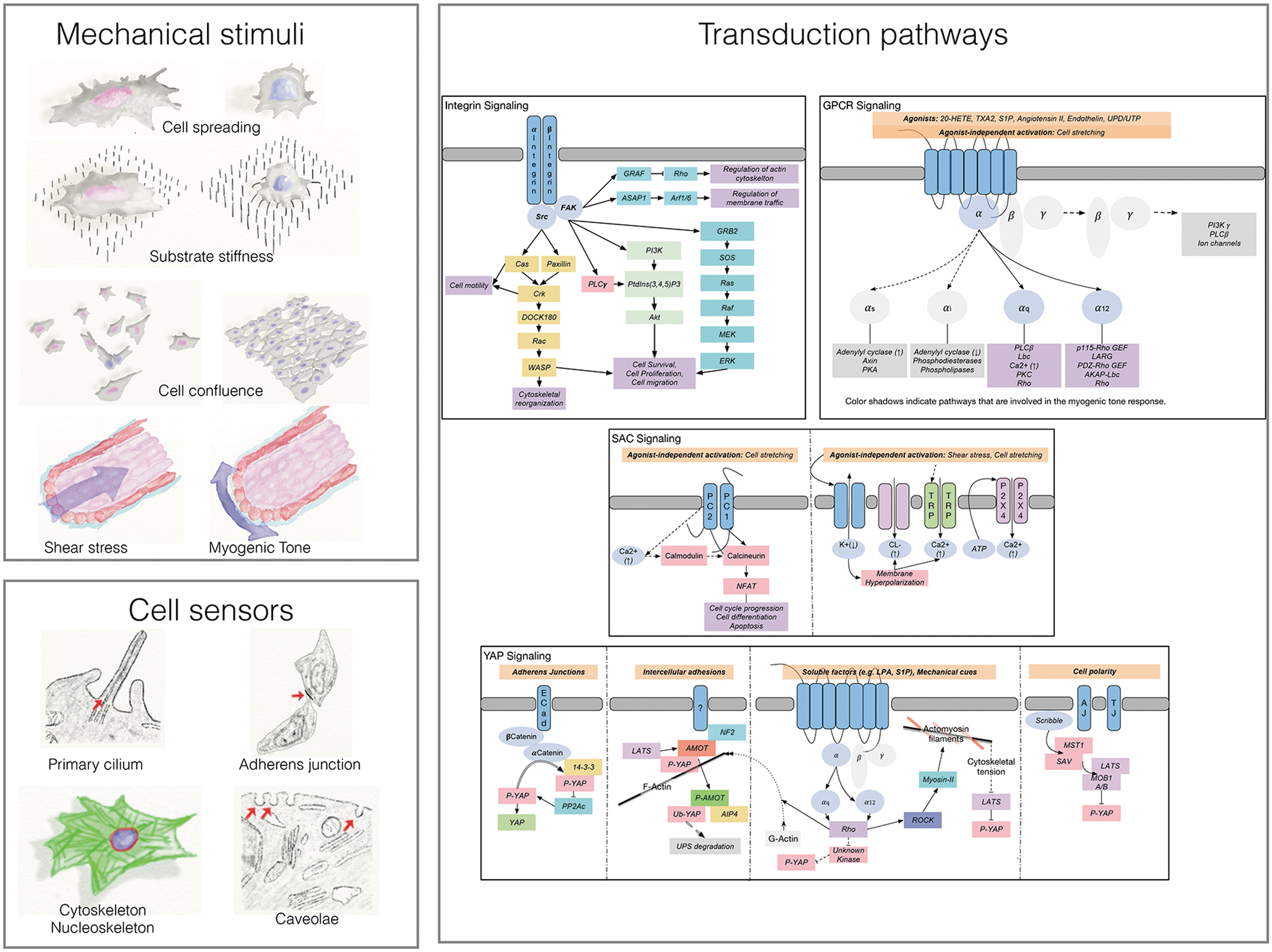
Mechanosensing and mechanotransduction. Cells are able to respond to diverse mechanical stimuli, thus modifying their behavior according to the microenvironment. Cell spreading, stiff substrates, and low degrees of cell confluence have been associated with the nuclear localization of YAP/TAZ (pink nuclei). Furthermore, cells are able to sense both shear stress and cell stretching, and transduce these stimuli by GPCR, SAC, or YAP signaling. Cell sensors such as the primary cilium, intercellular adhesions, the caveolae, the cytoskeleton, and the nucleoskeleton have been identified as supramolecular structures able to orchestrate the cellular response to physical stimuli. Transduction pathways involved in this process, which include integrin, GPCR, SAC, and YAP signaling, are depicted in this cartoon. See text for major details. GPCR, G protein-coupled receptor; SAC, stretch-activated channels; FAK, focal adhesion kinase; TRP, transient receptor potential; PC, polycystin; ECad, E-Cadherin; AJ, adherens junction; TJ, tight junction; UPS, ubiquitin proteasome system.
The Hippo pathway (name referring to the typical large size of Hippopotamus organs) has been at first identified in Drosophila and was subsequently found to be highly conserved in mammals, where it has been implicated in tissue architecture and renewal [33]. Recently, a direct association between the activity of the Hippo pathway and cytoskeletal tensioning has been established, noticing that geometric cues determined, for example, by a spread cell shape, high matrix stiffness, or low confluence, cause the shuttling of the YAP/TAZ transcriptional complex in the nucleus, with a consequent positive regulation of target genes mediated by DNA binding by TEAD1-4 transcription factors [34]. Conversely, the acquisition of a round shape by plating cells onto a low stiffness matrix or by high cell confluence determines phosphorylation of the YAP/TAZ complex by the kinase components of the pathway (eg, LATS1/2; MST1/2), with its consequent sequestration in the cytoplasm and degradation through the Ub/proteasome machinery [35].
The involvement of mechanosensing and the Hippo-dependent pathway in primary differentiation events of the mouse embryo has been highlighted by recent studies, which reported the dependence of the blastocyst cellular compartmentalization events on the differential activation of the Hippo-kinase pathway (sequestering YAP/TAZ in the cytoplasm) in the inner versus the outer blastocyst cells. In particular, it was found that in outer cells, in which intercellular adhesions are more firm and involve, for example, E-cadherin or other adherens junction-associated proteins, the cytoskeleton is under tension, and YAP/TAZ translocates to the nucleus, where it activates target genes involved in trophoblast formation [36 –38]. The cell contacts established in the inner blastocyst cell core activate the Hippo pathway; this typically leads to phosphorylation and inactivation of YAP/TAZ, with consequences for control of naive pluripotency genes, and specification of the inner cell mass (ICM) [38 –40]. A reverse scenario is calling for a role of the YAP/TAZ complex in maintenance of pluripotency as a transcriptional repressor of genes involved in early differentiation events. For example, it has been shown that the YAP/TAZ/TEAD trio participates in a transcriptional complex with Smad2/3, Oct-4, and NuRD, to repress mesendoderm differentiation genes in embryonic stem cells [41]. While this highlights a direct participation of YAP/TAZ in the maintenance of pluripotency, it also supports a crosstalk between mechanosensing-driven and morphogen-dependent pathways such as that of the transforming growth factor (TGF)-β [42]. This role is also consolidated by the function of the phosphorylated YAP as an enhancer of Smad-1 function for TGF-β- and BMP-dependent target gene expression [43].
Hippo inactivation and the consequent YAP activation promote cell proliferation in an undifferentiated state [44] and, conversely, sustain cell reprogramming of fibroblasts into induced pluripotent stem cells [45,46], therefore representing a possible target to modulate stem cell commitment for regenerative medicine applications. A very interesting speculation about the crossroad between stemness maintenance, YAP signaling, and geometry, derives, finally, from the different localization of YAP in the complete embryo compared to embryonic stem cells (ESCs) in vitro. In fact, differences have been observed between the rates of YAP cytoplasm/nuclear localization ratio in the ICM compared to ESCs, once derived from the ICM [38,46], again highlighting a strong interplay between the pluripotency regulation machinery and geometrical information from the environment.
The actions of traction forces transduced to the cells by the surrounding extracellular matrix or subtle differences in mechanical compliance/stiffness of the matrix itself have been found to be potent inducers of pathologic cell transformation [47,48], connecting the YAP/TAZ transcriptional control to cell growth in various solid tumors [49]. In addition, in analogy with the embryonic development, an intersection between mechanically driven and humoral control has been established by the finding that the YAP/TAZ complex can act as a flexible transcriptional coactivator cooperating with other nuclear intracellular cascades such as those activated by morphogen-dependent intracellular signals (eg, the Wnt/β-Catenin or the TGF-β-Smad) [35], metabolic pathways [50,51], and altered oxygen metabolism [52]. Taken together, these observations have established the basis of a direct relationship between mechanosensing-driven pathways and humoral/metabolic cues, thus amplifying the number of control “nodes” potentially involved in pathophysiological conditions associated to acute or chronic diseases.
Geometry and Mechanotransduction in Heart Growth Regulation
Figure 2 represents a schematic view of the relevance of mechanosensing at early stages of development and cardiac morphogenesis, aging, and failure. The heart is the first organ that starts functioning during the embryonic development of vertebrates. It arises from the heart tube (HT) by its looping and twisting [53] and subsequent septation and maturation into a structure with four chambers. The striking morphological changes involved in this process are associated to the beginning of its mechanical functioning as a pump [54]. While a definitive biological and mechanical maturation is needed to complete its morphogenesis, this primitive beating function supports the generalized increase in the oxygen and nutrients need that the whole embryo requires for its definitive growth.
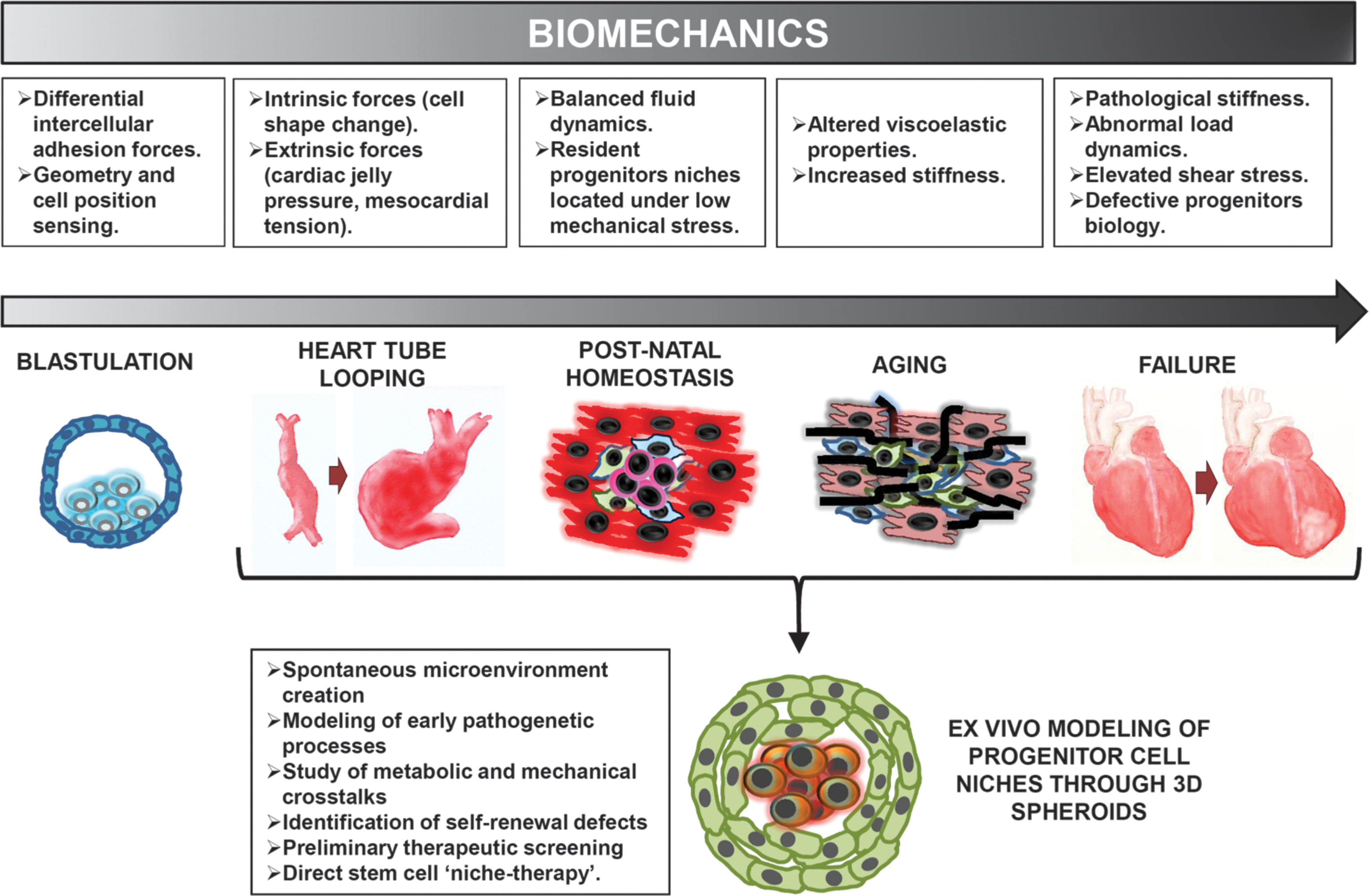
Evolution of biomechanics and position sensing in the normal and pathologic hearts. Modeling of mechanoperception in ex vivo three-dimensional cell aggregates to mimic cardiac development biology and pathologic process.
Physical forces and heart specification/growth
Physical forces are engaged in shaping morphogenesis and cell fate from very early stages. For example, intrinsic and extrinsic mechanical forces drive bending and torsion of the HT, respectively [55], and transmission of primitive flow dynamics in the HT through the endocardium is able to signal crucial developmental cues for the formation of myocardial trabeculae and the shaping of heart valves [56]. The mechanism generating intrinsic forces is becoming gradually clear, as multiple sources of physical forces have been identified, such as cardiac jelly pressure or dorsal mesocardium tension. However, the main bending force seems to stem from active changes in cell shape, which cause cells in the outer curvature to become more elongated than those in the inner curvature. This is a striking example of how intracellular signal transduction pathways associated to differential actin polymerization dynamics create forces able to move and distort structures of higher orders of magnitude, such as the whole HT itself [55,57].
The contribution of physical stimuli to the development of heart shape and function represents a recurrent motif in cardiac morphogenesis in a variety of vertebrate and nonvertebrate species. Subsequent developmental stages, such as s-looping, are again controlled by external forces deriving from other embryonic structures [58]. Once the heart becomes functional and blood flow starts, fluid dynamics become another physical factor significantly conditioning its development and physiology. The blood flow imposes a shear stress parallel to the endocardium and a force perpendicular to the tube wall, resulting in circumferential tension on the entire wall. After HT looping, such forces are not homogeneous anymore and increasing pressure is associated to specific tracts of the looped HT, such as prospective outflow tracts and atrioventricular valves [59]. Well-known pathways, such as that of PECAM1/VE-cadherin/VEGFR2 [60,61], Notch/ephrinB2 [62], or related to cilia formation [63], have been already identified, which could play a significant role in the mechanotransduction of physical stimuli from the endocardium. Progressive pressure loading in the embryo can also affect early cardiomyocyte phenotype, inducing changes in their physiology such as hypertrophy [64]. Although the dissection of this morphogenetic process is not yet completely understood, this influence becomes evident when developmental malformations arise due to abnormal embryonic fluid dynamics, which have been shown to significantly alter gene expression profiles [61], and potentially cause serious defects in heart anatomy at birth [65].
From physical forces to lineage development in heart formation
The employment of cell marking techniques to assess clonal development of developing tissues [66] and the combined use of image-assisted computational analyses have, finally, allowed to explore the dynamics of the cardiac looping as a process containing an intrinsic geometrical regulation. This mostly depends both on the cooperative participation of two independent sources of progenitor cells [67] and the mutual induction of premyocytes and endocardial cells, with a crucial contribution of position-dependent effects governing the formation of the definitive four-chambered structure from a simple tubular geometry with a primitive pulsatile contraction activity [68]. In this process, named cardiac trabeculation, paracrine effects due to endocardium/myocardium mutual induction are important; on the other hand, the dynamics of the cardiac motion is, at this stage, crucial to determine the spatial arrangement of trabeculation with specific regionalization of mechanotransductive molecule distribution [69] and preferentially oriented cell division [70]. Up to now, however, it is not known whether a mechanistic link exists between the anisotropic modality of cardiac cell proliferation and the intrinsic ability of immature cardiomyocytes to sense the mechanical environment, and the known relationship between the Hippo-dependent pathway and the embryonic/fetal heart growth [71]. On the other hand, recent evidences showed that YAP/TAZ functions may have implications in heart (re)generation [72 –75] thus suggesting relationships between cardiac development, homeostasis/renewal, and transduction of mechanical forces. How the extensive manipulation of mechanosensing-responsive pathways might enhance the slow steady renewal at the myocardial level [76,77], in favor of an effective myocardial regeneration, is still a matter of speculation.
The focus on physical factors, such as applied forces, flow strain, tissue geometry, and stiffness, has favored the setup of various in vitro and in vivo experimental models to recreate the complexity of native tissue micromechanics [19]. To this purpose, in vitro modeling of “synthetic” stem cell “niches” appears particularly promising to identify the molecular bases of tissue damage, or defects in self-renewal, screen for possible therapeutic interventions [78,79], find optimized cell–biomaterial combinations [20], or even release stand-alone niche therapeutic applications [80 –82]. First, hypothesized in 1978 by Schofield, the niche describes the limited microenvironment supporting stem cells [83,84]. Alterations of tissue niches have been soon predicted as important initial events in various pathophysiologic processes (ranging from alteration of tissue growth/renewal to insurgence of degenerative/neoplastic diseases). Therefore, the understanding of cues that subject the tissue niches (and the stem cells present in these environments) to these changes contributes to dissect the initial steps of pathogenetic processes or to model the disease in cell culture, to eventually validate intervention strategies and new therapies [85 –87]. In contrast to scaffold- or multisheet-based 3D in vitro cellular systems, used mostly in tissue engineering applications, scaffold-free spheroid models have historically represented a milestone in the study of tissue microenvironment and progenitor niches [88 –91]. From the embryoid bodies made with the hanging drop method of culture of embryonic stem cells to tumor spheroids and tissue-specific progenitor spheres (eg, neurospheres and mammospheres), these models share a natural tissue spherical geometry that recapitulates the primordial embryonic microenvironment, enabling a spontaneous mimesis of the tissue/organ they are derived from. Furthermore, as for natural niches, a primary function of the 3D spheroid in vitro model is to maintain stem cells into a permissive microenvironment, which is normally located in the center of the sphere. This geometrical confinement is particularly convenient for the dissection of the biological characteristics of progenitor cells. These include, for example, the ability of the cells to synthesize their own extracellular matrix components and secrete matrix-linked growth factors, to achieve a short-range control of cellular self-renewal versus differentiation, or the regulation of stem cell function(s) by a spatially regulated control of paracrine functions [92] (eg, IGF1 bioavailability) and oxygen metabolism in the core (more hypoxic) versus the external (less hypoxic) cell layers.
Modeling heart specification in 3D conditions; from developmental biology to pathology modeling
To be fully understood, the complexity of cardiac morphogenesis requires advanced techniques of investigation. In this context, the use of culture systems enabling cardiac niche and tissue self-assembly in 3D conditions are certainly preferable to finely dissect the cell-to-cell interactions. This model may be also useful to possibly dissect pathological evolution of the heart when major modifications in (stem) cell fate occur due to, for example, risk conditions (see below) or, finally, derive cells endowed with heart regeneration potential and use them for therapeutic applications. One of the most investigated 3D models of cardiac (re)generation is the so-called “Cardiospheres” (CSps) [93 –95]. CSps can be derived in culture from cells isolated by human cardiac explants (eg, from atrial appendage tissue), in which cells show distinct proteomic and gene expression profiles involved in cardiac muscle and vascular system development, compared with the corresponding progenitors cultured in 2D, and isolated by different methods and markers [96 –99]. Consistently, CSps have a more robust cardiac-related secretome profile and a higher cardioprotection activity, when administered into experimental models of myocardial ischemia compared to conventionally cultured cells [100,101]. Besides representing a reference tool for cardiac morphogenetic-like processes and a potentially useful tool for cardiac repair, CSps are also an interesting model for the dissection of the involvement of oxygen metabolism in cardiac progenitor cell (CPC) commitment. Indeed, the cells present in CSps (especially those present in the core) have a greater adaptation to hypoxic environment than monolayer-cultured cells [94], with a higher degree of glycolytic metabolic enzyme expression, resulting from adaptation of the cells to limited oxygen diffusion [96].
The mechanosensing machinery, which is particularly relevant for heart development and physiology (see sections above), could also be investigated in the microenvironment of CSps. As a matter of fact, the relevance of mechanosensing in the CSp context was recently disclosed with the finding that extracellular matrix rigidity can affect the differentiation of CSp-derived cells through the crucial involvement of the RhoA-activating intracellular transducer p190RhoGAP for divergence of cardiac versus endothelial lineages [95,102]. In addition, the presence of external and internal cells could ideally connect the differential cell behavior to the variable activation of the Hippo pathway and YAP/TAZ nuclear sequestering by position-dependent signaling. Given the recent implication of the Hippo signaling in the framework of metabolic activated pathways [35,50,51], CSps may be finally employed as tools to dissect the influence of metabolic factors on the susceptibility to mechanosensing or, vice versa, mechanosensing on the response to metabolic signaling. This latest consideration is not trivial, especially in the light of the reported crosstalk between mechanosensing and possible epigenetic driven mechanisms [103], which may connect, for example, the heart failure setting to permanently altered mechanotransduction [104].
Progression into Senescence and Failure: Metabolic Drift and Risk Conditions that Affect Mechanoperceptive Regulation of Cardiac Self-Renewal
The heart undergoes a continuous motion with more than three billion contraction cycles during the average human life span. In this context, the extracellular matrix turnover is crucial to control cardiac homeostasis and a continuous low-level degree of myocardial cell turnover. The maintenance of mechanical integrity of the heart is primarily controlled by cardiac fibroblasts (cFbs). Activation of these cells into pro-fibrotic elements (the so called myo-fibroblasts; myo-Fbs) occurs in response to various conditions promoting inflammation-driven matrix degradation and collagen deposition, leading to enhanced mechanical loading and stress [105]. While the increase in myocardial stiffness involves a change from a 10–20 kPa Young's Modulus physiologic value to a 50–200 kPa range, autocrine feedback signaling (eg, by TGF-β) sustains the mechanically induced fibrosis that ends up in a chronic scarring process [106]. The ability of cFbs to sense the mechanical environment has been considered a downstream component of the myocardium remodeling machinery; on the other hand, recent data, provided by us and others, have shown the susceptibility of human and mouse CPCs to respond to incremental stiffness values and cyclic strain by nuclear sequestering of the YAP/TAZ complex [107,108]. This is of relevance in the view that these cells, by virtue of their mesenchymal nature [109,110] and of the known ability of mesenchymal cells to respond to stiffness “gradients” [111,112], may consistently contribute to a shift in the equilibrium between the cFbs/cMyo-Fbs populations under pathophysiological conditions (eg, at the infarct border zone [107]), thus establishing a novel mechanoperception basis of cardiac remodeling.
Altered biomechanics in heart failure; a clinical perspective
The change in structural and loading alterations that condition the behavior of the resident cell populations depend on both modification of the chemical and physical properties of the myocardium and changes of the loading forces that are actively generated by, or act passively on, the heart. With regard to the first group, for example, hyperglycemia (through nonenzymatic glycation of cellular and extracellular matrix components) [113], hypertension (through alteration of T tubule organization, Ca2+ handling, and tissue fibrosis) [114], ischemia (through the development of scarring) [115], and aging (by means of complex interactions between pathophysiological changes of the cardiovascular system that have been summarized as “cardiovascular continuum”) [116] are all known to affect the viscoelastic properties of the myocardium [114]. Concerning the loading forces, the increase in cardiac preload, as in valvulopathies, or afterload, as in hypertension, are related to myocardial remodeling, ventricular dilation, and wall thinning, which, in turn, determine an increase in the ventricular wall stress, according to the Laplace law [117]. More complex, but equally important, mechanisms of altered cardiac mechanics include cardiac dyssynchrony that may lead to vigorous late-systolic contraction against high left ventricle pressures, imposing load on earlier activated region of the myocardium, which undergoes paradoxical stretch [118]. Finally, the diastolic filling vortex forces, which impose an elevated shear stress on the endocardium and have a compression effect on endocardium and myocardium (as a consequence of the centripetal and centrifugal forces associated with the right ventricle/left ventricle rotatory diastolic flow), are altered in heart disease or following surgical procedures. Intriguingly, a possible impact of this latter alteration and cardiac epigenetics, involving mechanosensing, has been hypothesized [119,120].
Detrimental impact of cardiac pathology on CPC function and mechanosensing-related gene expression pathways
In an effort to dissect whether the properties of CPCs remain unchanged from birth to aging and pathology, we and other authors have investigated the functional and biological features of CPCs isolated from human fetuses, normal hearts donated for cardiac transplantation, and hearts explanted from patients affected by end-stage heart failure [121 –123]. All these reports were in agreement concerning senescent CPCs to accumulate defects associated with basic cell function alterations, such as a reduction in the ability to migrate along a chemotactic gradient [124] and an increased sensitivity to contact inhibition [122]. Accordingly, transcriptome profiling in these cells showed a coherent enrichment of gene expression pathways related to cell motion, migration, invasion, and shape. Interestingly, among the most significantly differentially represented canonical pathways, we observed caveolar-mediated endocytosis signaling, integrin-linked kinase (ILK) signaling, signaling by Rho Family GTPases, Rho GDI signaling, regulation of actin-based Motility by Rho, tight junction signaling, and gap junction signaling [125], all of which point toward a major involvement of the mechanoperceptive apparatus. Whether cells from failing hearts are defective for mechanotransduction machinery under cyclic straining or static conditions is the subject of these investigations (A.P. Beltrami and M. Pesce, unpublished data).
Cooperative effects of mechanosensing and metabolic alterations in CPCs
A timely emerging concept is that cell signaling and transcriptional networks can be potently affected by metabolism and its intermediate products, thus integrating cell functions into complex biological frameworks depending on concomitant presence of risk conditions, often on a personalized basis. In this regard, again, the transcriptional coactivator YAP/TAZ may play a prominent role, being effectors of the Hippo kinase pathway targets of lysophosphatidic acid and sphingosine-1-phosphate [126] as well as of the mevalonate pathway [50,127]. Endosomes and lysosomes may also be primary actors in the integration of metabolic and mechanosensing pathways. Indeed, the chaperone-assisted selective autophagy (CASA) complex senses the mechanical unfolding of filamin and sorts, with the chaperone-associated ubiquitin ligase CHIP, damaged filamin to lysosomes for degradation. Intriguingly, autophagosome formation is increased as a function of the stiffness of the polyacrylamide cell substrate in a CASA-dependent manner. Nonetheless, filamin levels do not vary since a parallel increase in the transcription of filamin gene occurs in a BAG3 (a component of CASA) and YAP-/TAZ-dependent manner. Taken together, these results suggest that tension-induced unfolding of mechanosensors and cytoskeleton activates an adaptation program that is similar to a heat shock response [128]. Even more fascinating is the recently described integration between ligand-occupied integrin internalization, lysosome positioning, and recruitment and activation of mTOR in this subcellular compartment, linking nutrient signaling with cell migratory abilities [129]. We consider these findings of major importance for cardiac senescence process since we have recently shown that TORC1 hyperactivation and lysosomal dysfunction are typical features of CPCs obtained from failing hearts, which dictate their senescent phenotype [125]. It is therefore tempting to speculate that a block in the autophagy/lysosomal pathway may convey into an alteration of the mechanotransduction properties of CPCs residing in failing hearts. Experiments are currently being performed to address this intriguing hypothesis.
Caveola- and Sarcomere-Associated Structures: Novel Players in Cardiac Mechanosensing
Caveolae
Caveolae are 60–80 nm wide pits in the plasma membrane containing oligomeric caveolins that are stabilized upon interaction with cavin. These structures are a specialized form of lipid that act as a compartment into which elements of the endocytic system, lipid transporters, and cell signaling are confined [130]. Location of the caveolae in the cell membrane is not uniform. In fact, for example, they are concentrated in the rear aspect of migrating cells. Moreover, caveolar densities vary in different tissues and are particularly abundant in cells undergoing mechanical stress, such as endothelial cells [131]. Complex interactions between cytoskeleton components and caveolar structures have been described involving the actin cytoskeleton, microtubules, and intermediate filaments [131]. In particular, a primary role is thought to be played by β1 integrin and ILK, which are required for caveolar formation by recruiting the F-actin binding protein IQ-GAP1 to the cell cortex and cooperating with mDIA1 (a formin homology protein involved in actin polymerization) to stabilize the microtubules and allow the insertion of caveolae in the plasma membrane [132]. Notably, Abl tyrosine kinase and mDIA1 cooperate to coordinate stress fiber formation with caveolar anchoring to the cytoskeleton; this process requires filamin A, a protein involved in crosslinking the actin filaments and anchoring them on the cell membrane [133]. The compartmentalization of various cell signaling molecules within caveolae and their interaction with caveolins strongly support the hypothesis that these structures may be important hubs integrating signaling pathways, as first hypothesized in 1994 [134]. Specifically, CAV1 contains a scaffolding domain (CSD) that organizes signaling molecules in functional complexes. For example, it binds and inhibits endothelial nitric oxide synthase (eNOS), G protein-coupled receptors, Ras and Src family members, EGFR, PKA, and PKC. In mechanically stimulated cells, as it occurs to endothelial cells exposed to shear stress, the inhibitory function of CSD may be released and downstream molecules, such as eNOS, activated [135]. A special mention should be reserved to the interaction between caveolae and Rho family GTPases, which are crucial regulators of actin dynamics and cell motility. Specifically, loss of CAV1 is associated with impaired cell polarity, defects in wound closure, reduced Rho activity, and increased Rac and Cdc2 activities, which are secondary to Src activation [136]. With regard to mechanosensing, several reports have demonstrated that caveolae flatten and disassemble in response to cell stretching. This property prevents cell damage or lysis, as opposed to that observed in patients with muscle dystrophy associated with a Cav3 mutation, which is characterized by increased membrane fragility as a result of the absence of a functional caveolar reservoir [137]. The relevance of caveolae in cardiac mechanosensing and its modification under pathologic conditions are intriguing hypotheses that are likely to lead, in the future, to an increased integration of pathology-related pathways. Caveolar flattening, a process that follows changes in mechanical forces applied to cells [137], modulates cell signaling events, such as the activation of mechanosensitive ion channels (eg, the swelling-activated chloride channel ICl, swell ), neutral sphingomyelinase, eNOS, Src-like kinases, and tyrosine kinases [131,138,139]. In addition, both caveolins and cavins can have a direct effect on mechanosensing. For example, CAV1 can be phosphorylated at Tyr14 by Src kinase in response to growth factor stimulation [140] and integrin-mediated mechanotransduction [141]. Phospho(p)-CAV1Tyr14, finally, modulates focal adhesion dynamics, activates Rho/ROCK, and stimulates directional cell migration, by inhibiting Src-dependent activation of p190RhoGAP [136,142], furthermore, it stimulates caveolae formation by a feedback mechanism that involves the inactivation of the transcription factor Egr1, an event that relieves the inhibition of CAV1 and cavin-1 genes [143]. The altered expression of caveolin may be considered a crucial link between pathology and mechanosensing. Indeed, it was shown that upregulation of CAV-1 leads to cell senescence [144] and replicative senescence is characterized by upregulation of CAV-1, CAV-2, and CAV-3 [145]. Whether these molecular alterations are associated with an increased number of caveolae per cell or with increased endocytosed caveolar structures (ie, cavicles) is still a matter of discussion [146]. On the other hand, recent data indicate that Cavin-1, an essential component in the biogenesis of caveolae, is upregulated in human senescent fibroblasts and is required to increase the caveolar number and promote DNA damage in cell senescence [147]. Furthermore, senescent human fibroblasts are characterized by increased expression of β1 integrin and FAK, increased phosphorylation of FAK and paxillin, and increased activity of Rac and Cdc42 in caveolin-rich membrane domains. Intriguingly, downregulation of caveolin-1 in senescent cells inactivated FAK, reduced focal adhesion and stress fiber formation, and reverted the cell morphology from a flattened senescent shape to a smaller polarized phenotype [148]. Last, with regard to contact inhibition of proliferation, when fibroblasts reach confluence, caveolin-1 is shifted to areas of intercellular contact [149], while caveolin-1-deficient fibroblasts are characterized by loss of contact inhibition and anchorage-independent growth [150].
Sarcomeres and mechanosensing
The well-known contractile apparatus of the cardiomyocyte is the sarcomere, which is made of three different filaments (thin, thick, and elastic), whose overlapping and interaction create the typical structures of Z-disks and I/A/M bands. The resulting histological striated appearance is not only the basic force-generating structure but also a crossroad for sensors and signal transduction molecules [151]. In particular, Z-disks and titin elastic filaments bind complicated receptive and modulatory machineries, which are still largely unraveled. Multiple classical transduction pathways (also activated by mechanical stress), such as MAPK, PKC, and calcium signaling, are known to interact with this network [152], and this subcellular localization is most likely functional for their activation and downstream signaling [153]. One of these proteins that seem to be involved in mechanical sensing is muscle-specific LIM protein (MLP), which has been linked to the development of dilated cardiomyopathy both in animal models [154] and patients [155,156]. In fact, MLP is involved in the maintenance of a compact and functional Z-disk structure, contributing to the mechanical elastic properties of cardiac tissue. Interestingly, MLP is also able to interact with multiple muscle regulatory transcription factors, such as MyoD and MRF4 [157], evidencing its potential role as mechanical-to-biological transducer at the Z-disk location. The direct role of titin and elastic filaments, as well as of other candidate transducers (eg, PDZ-LIM protein family), is also emerging as a likely sensor network for mechanical stress in cardiomyocytes.
Intercalated disks are another fundamental structure for the contractile function of cardiomyocytes. They are complex membrane scaffolds for the electromechanical coupling of adjacent myocytes into the cardiac syncytium, whose function and integrity are necessary for creating the mechanical synergy of the tissue. They are another candidate mechanical sensor zone, since they represent the interlink between cell–cell junctions and desmosomes made by different cadherins, granting anchorage continuity through cell membranes between the cytoskeletons of adjacent cells. The potential mechanosensing role of intercalated disks is supported by the colocalization of proteins such as ALP [158], which belongs to the above-mentioned PDZ-LIM family of candidate mechanical transducers. Intercalated disks are enriched in an MLP binding protein, named N-RAP [159]; sequestration of many transcription factors, such as beta-catenin transcriptional regulator [160], has been described as well. Overall, key mechanical structures related to sarcomeres and intercellular junctions in cardiomyocytes seem to have more complicated functions than that of simple force generators or histological scaffolds, and are likely engaged in translating complex physical signals into biological and genetic responses.
Concluding Remarks and Future Perspectives
This contribution has been conceived to offer readers a comprehensive view of the mechanosensing process in the heart as a new cell physiology effector linking apparently separated worlds such as embryonic development and cardiac disease. The remarkable conservation of the cellular machinery transducing mechanical cues makes, indeed, possible to translate the early biological information laid down during embryogenesis and heart development to a disease-related setting that could be decrypted by modern 3D culture systems. While this may help dissect the role of changes in the mechanical characteristics of the myocardium (eg, after a myocardial infarction), this will also contribute to establish a possible link between the changes in metabolism with modification in cardiac mechanoperception, with consequent cardiac disease. This novel interpretation appears timely, given the always more intricate relationship between the functional readout of cell metabolism and cardiovascular risk conditions. Only future integrated approaches establishing links between the biophysical regulation of cardiac (progenitors) cell populations and deviations from the normal metabolic setup will help to follow heart development and disease from its early beats to its aging and disease.
Footnotes
Acknowledgments
This contribution was partially supported by grant 20123E8FH4 from MIUR-Italian Ministry of University and Education to I.C. and by grant RF-2011-02346867 from the Italian Ministry of Health, issued to M.P.
Authors Disclosure Statement
No competing financial interests exist.