
Abstract
Mesenchymal stem cells (MSCs) have a great potential for treating equine musculoskeletal injuries. Although their mechanisms of action are not completely known, their immunomodulatory properties appear to be key in their functions. The expression of immunoregulatory molecules by MSCs is regulated by proinflammatory cytokines; so inflammatory priming of MSCs might improve their therapeutic potential. However, inflammatory environment could also increase MSC immunogenicity and decrease MSC viability and differentiation capacity. The aim of this study was to assess the effect of cytokine priming on equine bone marrow-derived MSC (eBM-MSC) immunoregulation, immunogenicity, viability, and differentiation potential, to enhance MSC immunoregulatory properties, without impairing their immune-evasive status, viability, and plasticity. Equine BM-MSCs (n = 4) were exposed to 5 ng/mL of TNFα and IFNγ for 12 h (CK5-priming). Subsequently, expression of genes coding for immunomodulatory, immunogenic, and apoptosis-related molecules was analyzed by real-time quantitative polymerase chain reaction. Chromatin integrity and proliferation assays were assessed to evaluate cell viability. Trilineage differentiation was evaluated by specific staining and gene expression. Cells were reseeded in a basal medium for additional 7 days post-CK5 to elucidate if priming-induced changes were maintained along the time. CK5-priming led to an upregulation of immunoregulatory genes IDO, iNOS, IL-6, COX-2, and VCAM-1. MHC-II and CD40 were also upregulated, but no change in other costimulatory molecules was observed. These changes were not maintained 7 days after CK5-priming. Viability and differentiation potential were maintained after CK5-priming. These findings suggest that CK5-priming of eBM-MSCs could improve their in vivo effectiveness without affecting other eBM-MSC properties.
Introduction
M
The aim of this study was to assess the effects of cytokine priming on eBM-MSC immunoregulation, immunogenicity, viability, and differentiation potential, in search of a balance between stimulation of their immune-suppressive capacity without related induction of negative effects on other properties.
Materials and Methods
Animals
Four Shetland pony geldings (aged 4–7 years, weight 138–162 kg) were used as bone marrow (BM) donors. All horses were determined to be in good health. All procedures were carried out within the Project License 31/11 approved by the in-house Ethics Committee for Animal Experiments from the University of Zaragoza. The care and use of animals were performed accordingly with the Spanish Policy for Animal Protection RD53/2013, which meets the European Union Directive 2010/63.
Harvesting and characterization of eBM-MSCS
Twenty microliters of BM from sternum were obtained using a 4″ 11G Jamshidi needle in heparinized syringes. Equine BM-MSCs were isolated using a gradient density separation technique as previously described [12]. Cells were seeded in a culture medium consisting of low-glucose Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich) supplemented with 1% Glutamine (Sigma-Aldrich), 1% Streptomycin/Penicillin (Sigma-Aldrich), and 10% Fetal Bovine Serum (FBS; Sigma-Aldrich). Cells were expanded until passage three and characterized by phenotype determination and trilineage differentiation as previously described [12]. Subsequently, cells were frozen in 10% DMSO (Sigma-Aldrich)-90% FBS medium and cryopreserved until experiments started.
Priming of eBM-MSCs
Cryopreserved eBM-MSCs (n = 4) were thawed and seeded at 5,000 cells/cm2 in the basal medium at 37°C and 5% CO2 for 5 days, allowing readjustments of conditions before initiating the experiments. A preliminary assay was conducted to select the optimal time of exposure to cytokines (Supplementary Data and Supplementary Fig. S1; Supplementary Data are available online at
Real-time quantitative polymerase chain reaction
Expression of genes related with immunoregulation, immunogenicity, apoptosis, and differentiation markers was analyzed by real-time quantitative polymerase chain reaction (RT-qPCR). Total mRNA from approximately 106 eBM-MSCs from each sample was isolated with the RNAspin Mini RNA Isolation Kit (GE Healthcare). Genomic DNA was removed with the kit DNAse Turbo (Ambion) and 1 μg of mRNA from each sample was retrotranscripted to cDNA by using the Superscript Reverse Trancriptase Kit (Life Technologies). Isolation of mRNA and cDNA retrotranscription were performed with the kit Cells-to-cDNA II (Ambion) in eBM-MSC samples from differentiation assays and 7-day postpriming assessment. All procedures were performed according to manufacturer's instructions.
RT-qPCRs were performed and monitored using a StepOne Real-Time PCR System device (Applied Biosystems). All reactions were carried out using Fast SYBR Green Master Mix (Applied Biosystems) and 2 μL of cDNA as template. Amplification was performed in triplicate following the protocol: 20 s at 95°C, followed by 40 cycles consisting of 3″ at 95°C and 30″ at 60°C. A dissociation curve protocol was run after every reaction. Levels of gene expression were obtained using the comparative Ct method. Normalization factor (NF) was calculated as the geometric mean of the quantity of two housekeeping genes, GAPDH and B2 M [12]. Primers were designed with the Primer Express 2.0 software based on known equine sequences. Information about primers is shown in Table 1.
GenBank accession numbers of the sequences used for primers design. Primers (F: forward and R: reverse) and length of the amplicon in base pair (bp). Genes were grouped in agreement with the functions and implications of encoded molecules.
Proliferation
Proliferation after CK5-priming was evaluated by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay (Sigma-Aldrich) for 6 days as previously described [13]. Triplicates of each sample were seeded in the basal medium at 5,000 cells/cm2 in 96-well plates. The basal medium was used as a blank. Biotek Synergy HT spectrophotometer was used to measure the optical density (570 nm) in each well.
Viable cell numbers were determined by extrapolating from a standard curve consisting of nine triplicated points of increasing quantity of cells (0–15,000 cells/cm2) as previously reported [13]. Calibration curve equation for cell number extrapolation was y = 1 × 10−5 × −0.0349, r 2 = 0.997 (y = optical density of the well; x = amount of cells). Cell doubling times (DT) were calculated according to the formulae: CD = ln [(Nf/Ni)/ln2] and DT = CT/CD, where DT = cell doubling time; Nf = final number of cells; Ni = initial number of cells; and CD = cell doubling number.
Assessment of chromatin integrity
Chromatin integrity was evaluated by the chromatin dispersion test making use of the D3-Max Kit (Halotech DNA). This methodology was designed to assess DNA damage in MSCs through minor modification of the previously described technique for somatic cumulus cells [14] to MSCs. DNA damage was visualized using a Proyser fluorescent microscope (Proyser) employing SyberGreen II fluorochrome (Biotium) at 40× magnification. Cells showing a large halo of chromatin dispersion were considered to contain highly fragmented DNA (Supplementary Fig. S2). The percentages of normal cells and eBM-MSCs with fragmented DNA were determined in each sample by fluorescence microscope count by two independent observers. Equine BM-MSCs from one control were treated with hydrogen peroxide to perform a positive control of DNA damage through exposition to oxidative stress.
Trilineage differentiation assay
Equine BM-MSCs from both control and CK5 samples were exposed to the induction medium (differentiation) and basal medium (negative control) in triplicates according to each differentiation assay.
Osteogenic differentiation
Equine BM-MSCs were seeded at 20,000 cells/cm2 density in 24-well plates. The osteogenic medium consisted of 10 nM dexamethasone (Sigma-Aldrich), 10 mM β-glycerophosphate (Sigma-Aldrich), and 100 μM ascorbate-2-phosphate-supplemented (Sigma-Aldrich) basal medium. After 7 days, differentiation was assessed by Alizarin Red staining. Cells were fixed with 70% ethanol for 1 h at room temperature (RT), stained with 2% Alizarin Red stain (pH 4.6) (Sigma-Aldrich) for 10 min (RT), and washed with PBS (Gibco Invitrogen Corporation).
Adipogenic differentiation
Equine BM-MSCs were seeded at 5,000 cells/cm2 density in 12-well plates. The adipogenic medium consisted of 1 μM dexamethasone (Sigma-Aldrich), 500 μM 3-isobutyl-1-methylxanthine (Sigma-Aldrich), 200 μM indomethacin (Sigma-Aldrich), and 15% rabbit serum-supplemented (Sigma-Aldrich) basal medium. After 15 days, differentiation was evaluated by Oil Red O staining. The cells were fixed with 10% formalin (Sigma-Aldrich) for 15 min at RT, stained with 0.3% Oil Red O (Sigma-Aldrich) stain (dissolved in 60:40; isopropanol:distilled water) for 30 min at 37°C, and washed with distilled water.
Chondrogenic differentiation
For chondrogenic differentiation assay, both control and CK5-eBM-MSCs were exposed to a chondrogenic induction medium, but due to the number of available cells, culture in the basal medium was not carried out. Approximately, 300,000 eBM-MSCs from each sample were transferred to conic bottom 15 mL tube and 400 μL of chondrogenic differentiation medium was added and centrifuged at 1,750 rpm for 5 min to pellet the cells. The chondrogenic medium consisted of 10% FBS, 10 ng/mL TGFβ-3 (R&D Systems), ITS+ premix (Beckton Dickinson), 40 μg/mL proline (Sigma-Aldrich), 50 μg/mL ascorbate-2-phosphate (Sigma-Aldrich), and 0.1 μM dexamethasone-supplemented (Sigma-Aldrich) high-glucose DMEM (Sigma-Aldrich).
After 21 days, chondrogenic differentiation was evaluated by specific staining with Alcian Blue stain. Pellets were fixed in 10% formalin, embedded in paraffin, and sectioned into 5-μm sections. The sections were hydrated with increasing gradients of alcohols, stained with Mayer's hematoxylin and 3% Alcian Blue dyes, rinsed with distilled water, dehydrated with decreasing amounts of alcohol, and mounted.
Statistical analysis
Statistical analysis was performed using the SPSS 15.0 (SPSS, Inc.). Normality of each data group was tested with the Shapiro–Wilk test. Differences in mean cell numbers between control and CK5 eBM-MSCs along the proliferation assay were analyzed by the nonparametric ANOVA Kruskall–Wallis test with Dunn's post hoc test. Kruskall–Wallis test was also used to analyze differences between control-nondifferentiated, control-differentiated, CK5-nondifferentiated, and CK5-differentiated eBM-MSCs from trilineage differentiation assays. Differences in gene expression between CK5-eBM-MSCs and their controls were analyzed by Mann–Whitney test. Differences in percentages of normal (nonfragmented DNA) and DNA-damaged cells were analyzed between control and CK5-exposed cells by student's t test. Significance level was set at P < 0.05 for all analyses.
Results
Equine BM-MSC immunomodulation after cytokine priming
Cytokine priming induced significant upregulation of the immunoregulatory-related genes Vascular Cell Adhesion Molecule 1 (VCAM-1), Indoleamine 2,3-dioxygenase (IDO), Inducible Nitric Oxide Synthase (iNOS) (P < 0.01), and Interleukin 6 (IL-6) (P < 0.05) by eBM-MSCs. In addition, nonsignificant upregulation of Cyclooxygenase 2 (COX-2) and significant downregulation of Interleukin 10 (IL-10) (P < 0.05) were observed (Fig. 1A). Expression of significantly upregulated genes was evaluated 7 days after CK5-priming and IL-6, IDO, iNOS, and COX-2 expression remained similar to unstimulated control and VCAM expression significantly decreased (P < 0.05) (Fig. 2).
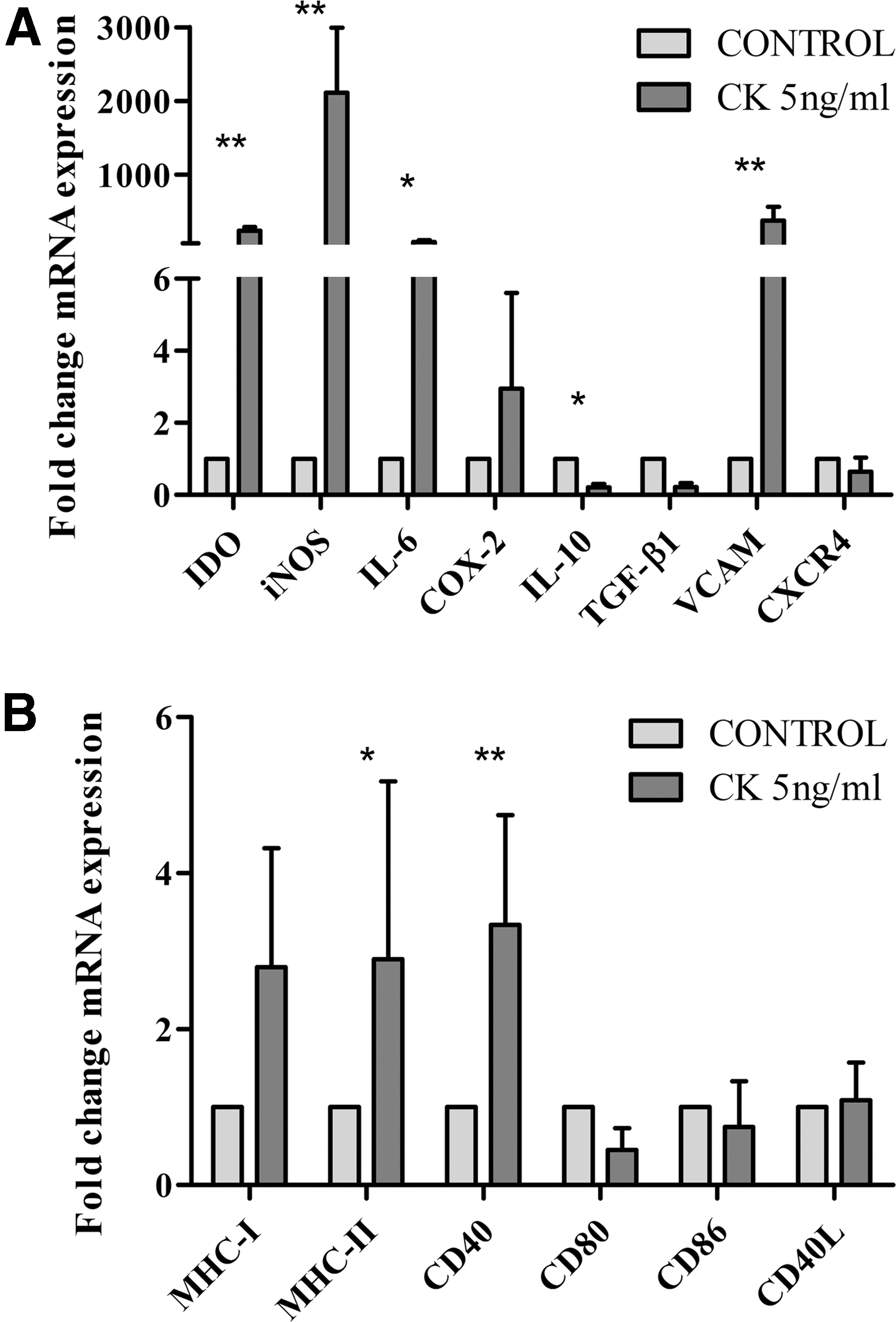
Gene expression data are reported as mean (n = 4) fold increase or decrease of CK5-stimulated eBM-MSC gene expression over unstimulated control eBM-MSC.

Immunomodulatory and immunogenic related molecule expression 7 days after CK5-priming. Gene expression data are reported as mean (n = 4) fold increase or decrease of CK5-stimulated eBM-MSC gene expression over unstimulated control eBM-MSCs. (* = P < 0.05).
Equine BM-MSC immunogenicity after cytokine priming
Cytokine priming induced a significant increase in the expression of MHC-II (P < 0.05) and the costimulatory molecule CD40 (P < 0.01) by eBM-MSCs. MHC-I was upregulated in a nonsignificant manner and the expression level of costimulatory molecules CD80, CD86, and CD40L remained unchanged (Fig. 1B). Seven days after the 12-h cytokine priming, the expression level of most relevant immunogenic molecules was assessed and MHC-I and MHC-II expression decreased, reaching similar values than in control cells (Fig. 2).
Equine BM-MSC viability after cytokine priming
Proliferation assay
Proliferation data are presented in Fig. 3A. CK-exposed cells grew in a similar manner to control cells: significant differences in the number of cells were not observed between control and CK5-exposed cells and cell DT was similar between control cells (3.071 ± 1.316 days) and CK5-stimulated eBM-MSCs (3.280 ± 1.307 days).
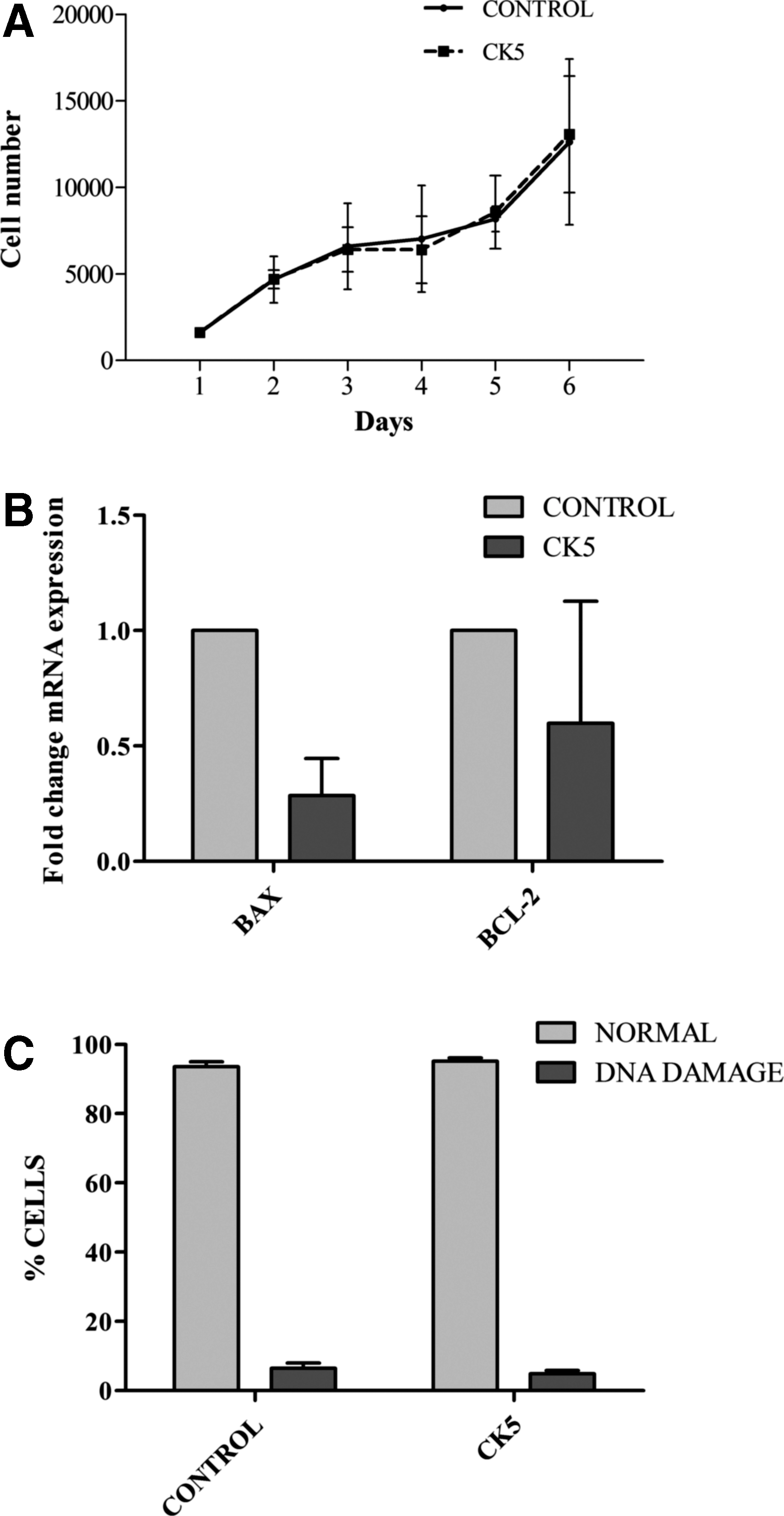
Data from viability assays.
Apoptosis gene expression
Proapoptotic gene BCL2-Associated X Protein (BAX) was downregulated in CK5-eBM-MSCs compared to control cells (P < 0.005), whereas there were no significant differences between control and stimulated cells for the antiapoptotic gene BCL-2 (Fig. 3B).
Chromatin integrity
Percentage of normal cells (nonfragmented chromatin) and DNA-damaged cells (dispersed chromatin fragments) did not show significant differences between control and CK5-eBM-MSC samples (Fig. 3C).
Equine BM-MSC differentiation after cytokine priming
Osteogenic, adipogenic, and chondrogenic differentiation were achieved in control cells from all animals. Spontaneous differentiation was not observed in any of the nondifferentiated controls (Fig. 4A, B).
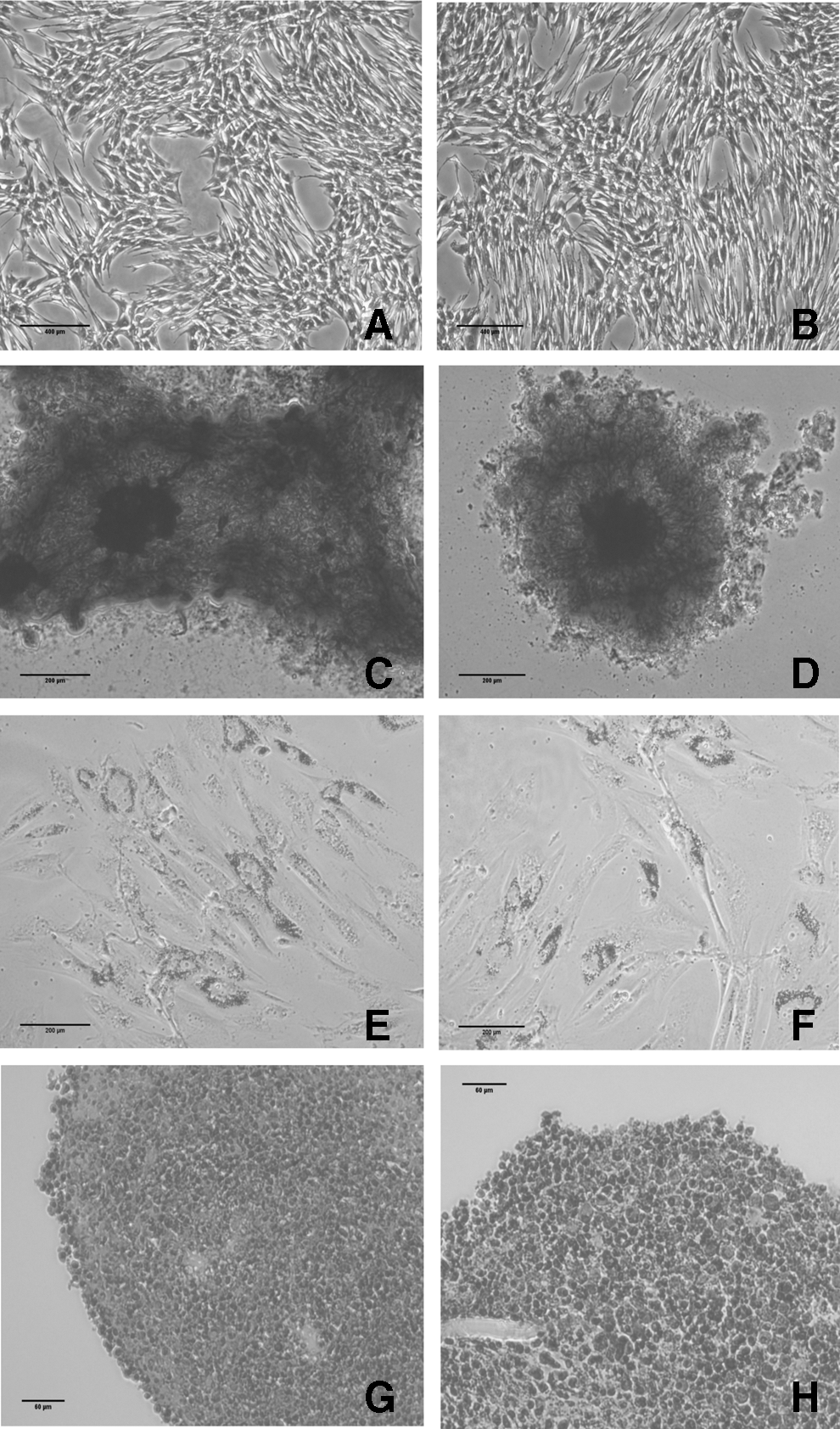
Staining for osteogenic, adipogenic, and chondrogenic differentiation of control and CK5-stimulated eBM-MSCs. Control
Osteogenic differentiation
Osteogenic differentiation was confirmed in both control and CK5-exposed eBM-MSCs by positive staining of the calcium deposits with Alizarin Red (Fig. 4C, D). Alkaline phosphatase (ALP) and Runt-related transcription factor 2 (RUNX2) gene expression was significantly higher (P < 0.05) in both control and CK5-differentiated samples over their corresponding nondifferentiated cells. In addition, expression of RUNX2 was significantly higher (P < 0.05) in differentiated control eBM-MSCs over differentiated CK5-exposed cells (Fig. 5A1–2)
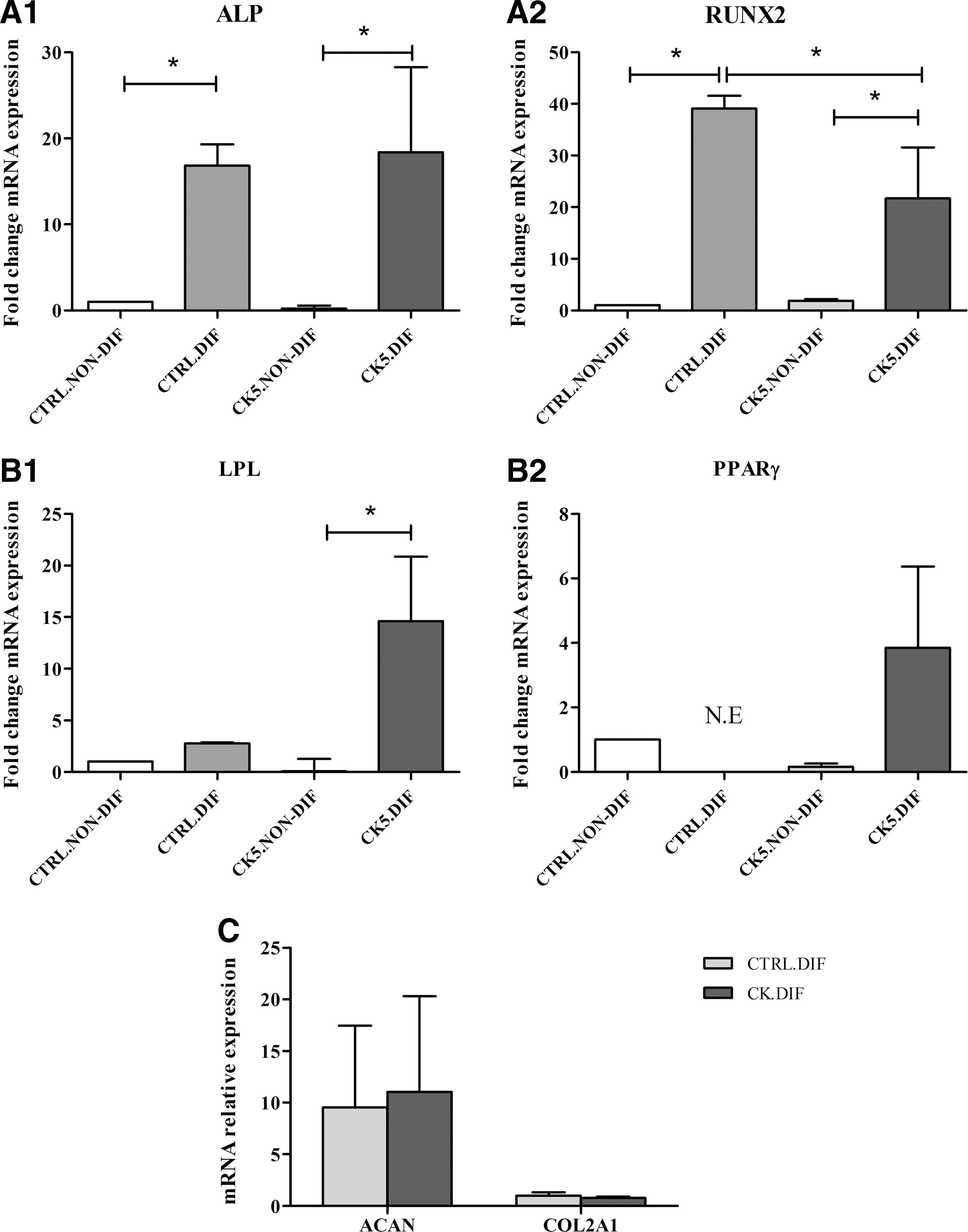
Gene expression results of trilineage differentiation assay. Results from osteogenesis
Adipogenic differentiation
Lipid droplets were detected in both control and CK5-exposed eBM-MSCs under adipogenic induction by Oil red O staining (Fig. 4E, F). Lipoprotein lipase (LPL) mRNA level was significantly higher only in differentiated CK5-eBM-MSCs over correspondent nondifferentiated cells (P < 0.05) (Fig. 5B1).
Chondrogenic differentiation
Chondrogenic phenotype was achieved by both control and CK5-exposed cells, detected by positive blue staining of the extracellular matrix by Alcian Blue staining and lacunae formation (Fig. 4G, H). Both aggrecan (ACAN) and collagen type II alpha I (COL2A1) showed similar level of mRNA relative expression in both differentiated control and CK5-eBM-MSCs (Fig. 5C).
Discussion
TNFα and IFNγ synergistically induce eBM-MSC immunoregulatory abilities [10], but certain cytokine concentrations and time of exposure also produce a decrease in their proliferation and differentiation abilities [11]. Therefore, eBM-MSC stimulation with a lower cytokine dose and a shorter time of exposure was tested to reach immunoregulatory induction without impairing viability and plasticity or increasing immunogenicity. Concentration of cytokines was chosen from a previous report in other species [3] and the time of exposure was set according to a preliminary assay.
Both cell–cell contact and paracrine signaling mechanisms are implied in the immunoregulatory functions of MSCs [2]. The adhesion molecule VCAM-1 participates in immunosuppression exerted by MSCs [15] and it was upregulated by CK5 priming, supporting the participation of direct cell-contact pathways in the eBM-MSC immunoregulatory mechanisms. Adhesion molecules are also related to MSC recruitment and migration, and these processes are particularly complex in inflammatory environments, showing both enhancement [16] and impairment of this ability [17]. The chemokine receptor CXCR4 participates in MSC migration in other species [18], but this is not evident in equine MSCs according to our findings and previous reports [10].
Paracrine signaling immunomodulation mechanism is mainly governed by molecules such as IDO, iNOS, IL-6, and COX-2, eliciting different mechanisms on immune cells [2], and TNFα and IFNγ are reported as inductors of these molecules by MSCs from different species and sources [19,20]. CK5-priming of eBM-MSCs led to a significant increase in the expression of immunomodulatory-related genes, similar to previous reports in equine species using higher cytokine concentrations [10]. IL-10 and TGF-β1 could also participate in MSC immune mechanisms [2,21], but other reports have shown that inflammatory exposure could elicit different effects on these molecules, ranging from increases in their production [22], no change of their expression/secretion [23], or even a significant reduction [24]. CK5-priming did not alter TGF-β1, but downregulated IL-10 expression, similar to previous findings in eBM-MSCs stimulated with higher cytokine doses [10].
Since immunoregulatory mechanisms can vary between different species and MSC sources [20], a variety of factors could participate in the MSC immunosuppression mechanisms. Our findings suggest that IL-10 and TGF-β1 may not have a major role in eBM-MSC immunoregulation, whereas IDO, iNOS, IL-6, or COX-2 would be mainly involved. Changes induced in eBM-MSC immunoregulatory-related gene expression by CK5-priming confirmed that a lower dose and a shorter time of exposure to proinflammatory cytokines can induce a similar enhancement in the eBM-MSC immunomodulatory profile to that described using higher cytokine concentrations and longer exposure [10].
Inflammatory environments could also produce a rise in MSC immunogenicity, with adverse safety and efficacy implications for their allogeneic use. Allogeneic MSC therapy is especially interesting when cell therapy is needed immediately or there are handicaps for autologous MSC use, such as aged and/or diseased patients [25,26].
Therefore, preservation of low immunogenicity after inflammatory stimulation is important to allow safe MSC allogeneic transplantation. Inflammatory stimuli, particularly IFNγ, might induce MHC expression [5,27]. CK5-priming induced a significant increase of MHC-II expression in eBM-MSCs, as it was observed with higher doses and longer exposure [10]. Even so, IFNγ-induced MHC upregulation may not lead to an increase in immunogenicity because of the lack of costimulatory molecules CD40, CD40L, CD80, and CD86 MSC expression, needed for antigenic recognition [28,29]. Nevertheless, inflammatory conditions might upregulate CD40 expression in human MSCs without affecting the expression of CD80, CD86, and CD40L [27,30]. In a similar manner, CD40 significant upregulation was detected after CK5-priming. However, absence of changes on costimulatory molecules CD80, CD86, and CD40L expression, combined with the upregulation of immunoregulatory molecules, suggests that eBM-MSCs might conserve their immune-evasive status after inflammatory priming [31].
Immunoregulatory functions of MSCs might be improved by proinflammatory cytokine priming, but how long these changes are maintained for after the stimulation remains unknown. Our results showed that the overexpression of VCAM-I, IL-6, iNOS, and COX-2 achieved by CK5-priming was not maintained after 7 days. In the same way, MHC-II upregulation did not remain after 7 days. These facts suggest that changes induced by cytokine priming might be short-term events, which may not remain for long after the in vivo implantation.
Furthermore, TNFα and IFNγ may synergistically induce apoptosis in murine MSCs [8]. However, there are conflicting reports showing that these cytokines can also promote human and murine MSC proliferation [24,32]. CK5-priming did not alter the regular growth of eBM-MSCs and relevant changes in the expression of genes involved in apoptosis [33] or chromatin integrity [34] were not induced. These findings suggest that CK5-priming is well tolerated and does not hamper eBM-MSC viability.
Impairment of MSC differentiation ability by inflammatory environment has been well documented [6]. TNFα inhibits RUNX2 expression in human and rat [7,35] and IFNγ might prevent MSC osteogenesis in vivo in allogeneic implants in mice [36]. High level of TNFα leads to a reduction in PPARγ expression by rat MSCs undergoing adipogenesis [37] and might also impair the chondrogenesis of murine and rat MSCs [6]. Possible mechanisms participating in the differentiation inhibition by inflammatory environment have been described in human MSCs [38], but on the other hand, TNFα might have osteogenic promotive effects. Indeed, TNFα enhanced human MSC osteogenic differentiation as measured by increases in the ALP activity and matrix mineralization [7], increasing Osteocalcin [39], RUNX2, Osterix, and bone morphogenetic protein 2 (BMP-2) expression [32].
In eBM-MSCs, adverse effects of TNFα and IFNγ exposure on trilineage differentiation have been reported when using higher cytokine doses and exposure time [11]. Nevertheless, eBM-MSCs were able to differentiate into the three lineages after CK5-priming. Previous studies describing differentiation impairment generally used cytokine concentrations similar to those used in this study, but these were added to the culture medium along the differentiation induction [6,7,35], whereas we pretreated cells and induced differentiation without adding these cytokines. Other studies using cytokine pretreatment and subsequent regular differentiation induction also reported negative effects, but these studies used higher amounts of cytokines for priming MSCs [11,40].
In summary, the effect of proinflammatory cytokines on MSC differentiation is considered detrimental, but under certain conditions, they can promote an enhancing effect. Our results suggest that a short cytokine exposure with low doses is well tolerated by eBM-MSCs, without abolishing their differentiation mechanisms or allowing its recovery after CK5-priming.
In conclusion, priming eBM-MSCs with 5 ng/mL of TNFα and IFNγ for 12 h showed an upregulation of immunoregulatory-related genes without a significant increase of most of the immunogenic ones and without eBM-MSC viability and plasticity impairment. Since the inflammatory environment in a naturally occurring disease, like in equine joint injury, could be heterogeneous and may not consistently induce changes in the immunoregulatory MSC profile [10], eBM-MSC CK5-priming enables this induction.
The stimulation with TNFα and IFNγ of MSC populations with initially different immunosuppressive capacity increases regulatory ability to a similar level, leading to homogenous MSC immunosuppressive capacity between different cell populations [41]. Consequently, prior ex vivo priming of eBM-MSCs could improve the effectiveness of cellular therapy. In addition, immunogenicity, viability, and plasticity of eBM-MSCs after cytokine priming must be considered. Viability and differentiation abilities of eBM-MSCs were preserved after CK5-priming. Furthermore, the possible effect of upregulated MHC-II might be attenuated by the lack of costimulatory molecules and by the enhancement in the immunomodulatory expression profile by eBM-MSCs. This work contributes to clarify the effects of inflammatory exposure on eBM-MSCs and constitutes a previous step to enhance their therapeutic use. Further studies will be needed to clarify the effect of cytokine priming on the safety and efficacy of eBM-MSCs.
Footnotes
Acknowledgments
We acknowledge the Veterinary Hospital of the University of Zaragoza for allowing the use of its facilities for sample collection. Authors thank Dr. J.A. Castillo for facilitating access to fluorescence microscope for chromatin integrity assessment.
Author Disclosure Statement
None of the authors have any financial or personal relationships that could inappropriately influence or bias the content of article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.