
Abstract
Repair of calvarial bony defects remains challenging for craniofacial surgeons. Injury experiments on animal calvarial bones are widely used to study healing mechanisms and test tissue engineering approaches. Previously, we identified Gli1+ cells within the calvarial sutures as stem cells supporting calvarial bone turnover and injury repair. In this study, we tested the regenerative capacity of the suture region compared with other areas of calvarial bone. Injuries were made to mouse sagittal sutures or other areas of the calvarial bone at varying distances from the suture. Samples were collected at different time points after injury for evaluation. MicroCT and histological analyses were conducted. EdU incorporation analysis was performed to assay cell proliferation. Gli1-CreERT2;Tdtomatoflox mice were used to trace the fate of Gli1+ stem cells after injury. Calvarial sutures possess much stronger regeneration capability than the nonsuture bony areas of the calvaria. The healing rate of the calvarial bone is inversely proportional to the distance between the suture and injury site: injuries closer to the suture heal faster. After complete removal of the sagittal suture, regeneration and restoration of normal organization occur within 6 weeks. Gli1+ cells within the suture mesenchyme are the cellular source for injury repair and bone regeneration. These results demonstrate that calvarial bone healing is not an evenly distributed event on the calvarial surface. Sutures contain stem cells and are the origin of calvarial bone tissue regeneration. Therefore, current practice in calvarial surgery needs to be reevaluated and modified. These findings also necessitate the design of new approaches for repairing calvarial bony defects.
Introduction
B
Large defects in the calvarial bones caused by pathological conditions or trauma will remain patent if no intervention is made and are called critical-sized defects (CSDs). The reasons that CSDs fail to repair are unknown, but contributing factors may include injury size, age, systemic conditions, nutrition, sex hormones, and infection [13]. Over time, the injury site is filled with fibrous tissue instead of bone. CSD treatment remains a major challenge for clinicians, and current surgical approaches for large craniofacial CSDs include the use of autologous, allogenic, and prosthetic materials [14 –17]. Neither of the models described above, which attribute the healing capacity of cranial bones to the periosteum or dura, can provide a satisfactory explanation for the persistence of CSDs.
In our previous study, we identified Gli1+ cells within the suture mesenchyme as the stem cells supporting calvarial bone turnover and injury repair [18]. Our study indicated that Gli1+ cells give rise to the periosteum, dura, and osteocytes of the calvarial bones. Stem cells within the suture can be activated and begin to proliferate immediately following injury, whereas cells in the nonsuture regions are incapable of repairing calvarial bone defects. Identification of the suture as the major location of mesenchymal stem cells suggests that the regeneration capacity of the calvarial bones should reside within the sutures. In our current study, we investigated the responses of different regions of the calvarial bones upon injury using mice and rabbit as models. We found that sutures possess much stronger regeneration capacity than other regions of the calvaria. Moreover, the healing rate of calvarial bone is inversely proportional to the distance between the suture and injury site.
Materials and Methods
Animal information
All animal studies were performed in accordance with federal regulations and with approval from the Institutional Animal Care and Use Committee (IACUC) at the University of Southern California. We obtained the following mouse strains from the Jackson Laboratory: Gli1-LacZ (JAX#008211), Gli1-CreERT2 (JAX#007913), and ROSA26LoxP-STOP-LoxP-Tdtomato (JAX#007905).
Suture and calvarial bone injury assays
In the suture injury assay, a sagittal incision was made in the midline region. The scalp was then revealed and the sagittal suture was exposed. A dental round bur of 1.8 mm diameter was used to drill through the sagittal suture. In the calvarial bone injury assay, the parietal bones were exposed and a dental round bur of 1.8 mm diameter was used to drill a hole at various distances from the sagittal suture. A handheld drill (NSK Z500) was used. Extreme care was taken not to damage the underlying dura. The scalp was then sutured with 4–0 nylon or wound clips. The distance from the edge of the injury site to the center of the sagittal suture was measured. The healing ratio was defined as (initial diameter of the injury site−final diameter of the injury site)/(initial diameter of the injury site).
In the rabbit injury experiments, the rabbit was anesthetized and a 5-cm incision was made in the skin. The tip of the scissors was used to expose the suture and bone. A 2-mm round bur was used to make round defects in different areas of the parietal bone. The dura mater remained intact. The incision was closed with an autoclip or using 4–0 nylon sutures.
EdU incorporation
EdU (A10044; Life Technology) was injected intraperitoneally into mice 2 h before sample collection at a dosage of 200 mg/kg. Calvarial samples were fixed with 4% PFA (paraformaldehyde) overnight and decalcified in 10% EDTA for 1–2 weeks. Decalcified bone tissues were embedded in optimal cutting temperature (OCT) compound (TissuTek, Sakura) and sectioned at 12–18 μm thickness. EdU+ signals were detected with a Click-iT® EdU Alexa Fluor® 488 Imaging Kit (C10337; Invitrogen) following the recommended protocol. Cell nuclei were stained with DAPI. For quantification of EdU+ cells, we counted the number of stained nuclei in each section.
Tamoxifen administration
Tamoxifen (T5648; Sigma) was dissolved in corn oil (C8267; Sigma) at a concentration of 20 mg/mL and injected intraperitoneally for 4 days at a dosage of 10 mg per day per mouse.
MicroCT analysis
The calvarial bones were removed and fixed in 4% PFA overnight, then radiographed using a Skyscan1174 compact MicroCT. The images were processed and analyzed using Amira 3D software for Life Sciences.
X-gal staining
Samples from Gli1-LacZ mice were fixed with 0.2% glutaraldehyde for 2 h and then decalcified with 20% EDTA for 1 week. After embedding in OCT compound, frozen sections of 12 μm thickness were cut. Sections were then stained with X-gal overnight at 37°C.
Histology
Calvarial bone samples were dissected under a stereomicroscope (Leica L2) and fixed in 4% PFA. Samples were then decalcified in 10% EDTA for 14 days. Decalcified samples were embedded in OCT compound and cryosectioned at 12–16 μm thickness using a cryostat (CM1850; Leica). Sections were stained with Vector® Nuclear Fast Red (H-3403).
Results
Based on our previous findings that Gli1+ stem cells within the suture mesenchyme support calvarial bone turnover and injury repair, we compared the regeneration capacities of suture and nonsuture regions of calvarial bone after injury. We created injury sites, 2 mm in diameter, in the skulls of 6–8-week-old CD1 mice, centered on the sagittal suture or in the central region of the parietal bone (Fig. 1A). One month after surgery, injury sites in the central region of the parietal bone had not healed (Fig. 1B–D). In contrast, injuries to the sagittal suture healed completely and were indistinguishable from uninjured suture regions (Fig. 1B–D). Histological analysis indicated that the nonsuture injury sites were filled mostly with fibrous tissue and had little bone formation 1 month after injury, whereas the suture injury sites were restored with newly formed bone that had nearly normal organization (Fig. 1E). These results indicate that the suture regenerates faster than nonsuture regions of the calvarial bone following injury.
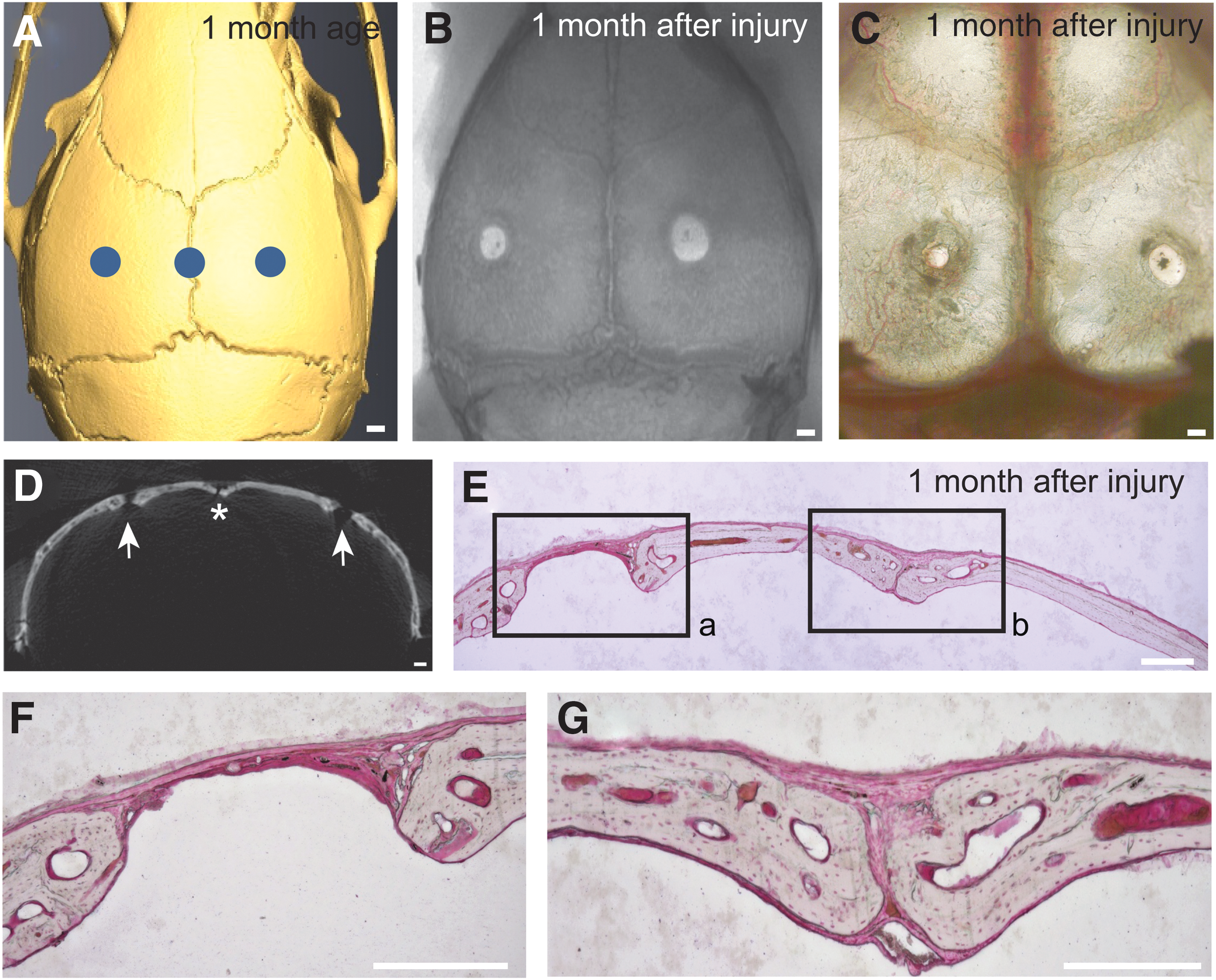
The suture possesses stronger regenerative capacity than other regions of the calvarial bone.
Next, we investigated the time course of the healing process of the suture after injury. After drilling 2 mm diameter round holes in the middle portion of the sagittal suture of 6–8-week-old CD1 mice, we collected calvarial samples at various time points after injury (n = 6 per time point). The injury sites did not change significantly 1 week after injury (Fig. 2A). MicroCT images indicated that significant closure of the injury site occurred by 2 weeks postinjury (Fig. 2B). By 3 weeks, the suture injury site was almost closed (Fig. 2C). Four weeks after injury, the site was completely restored to normal morphology and was indistinguishable from the uninjured portion of the sagittal suture (Fig. 2D). In addition, we analyzed the healing process histologically. One week after injury, the injury site was filled with fibrous tissue (Fig. 2E). The two osteogenic fronts began to approach each other by 2 weeks postinjury, and the soft tissue appears to have been overproliferating (Fig. 2F). Three weeks after injury, the original injury site was filled with new tissue and appeared comparable with a normal suture (Fig. 2G). By 4 weeks, the histological organization of the injury site was restored to normal and the suture remained patent (Fig. 2H).
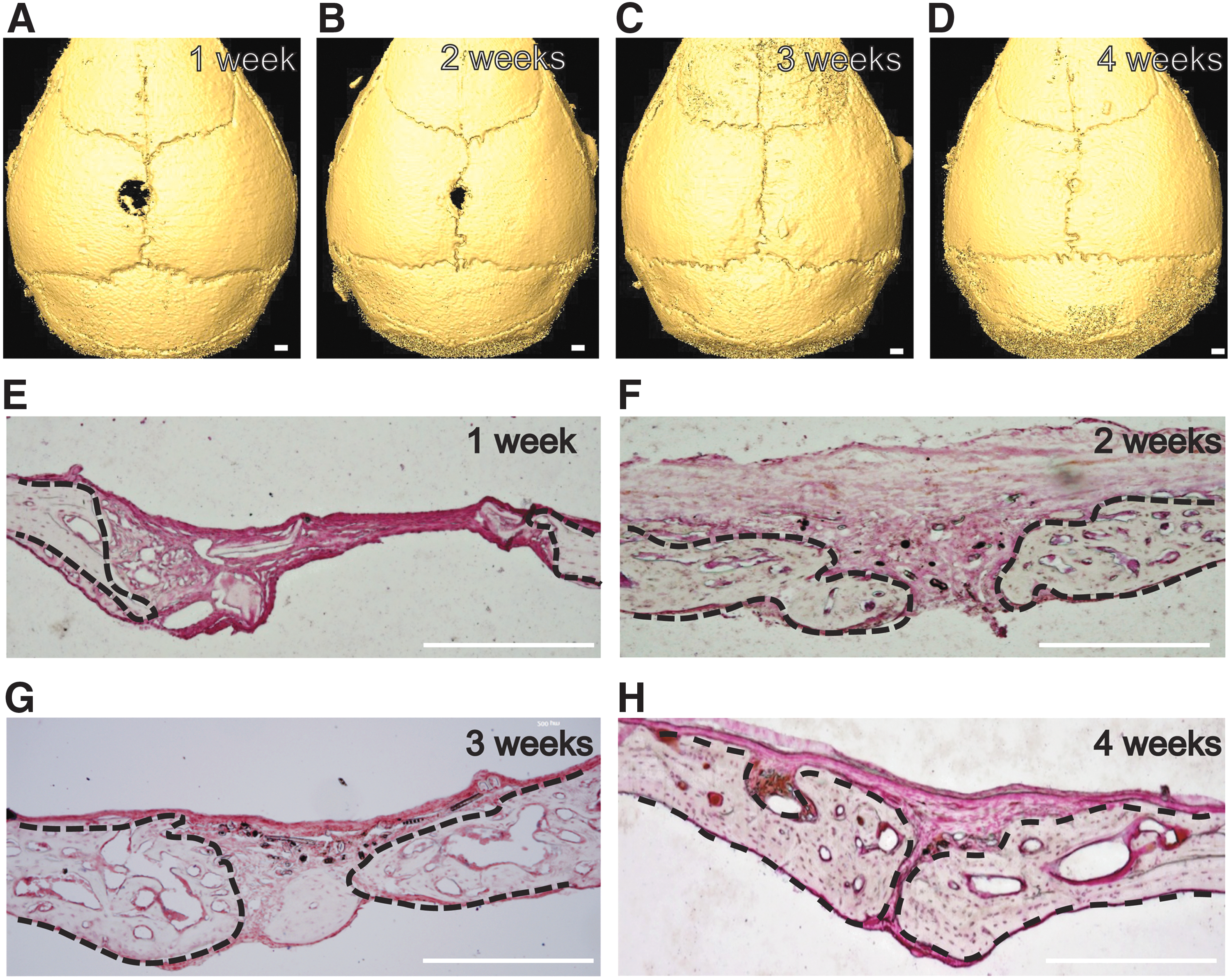
Time course of suture injury repair.
To investigate the effect of removing the entire length of the sagittal suture, we created a 2 mm by 5 mm injury longitudinally along the sagittal suture. One week postinjury, new bone formation was detectable both anterior and posterior to the injury site (Fig. 3A). The injury sites continued to close from both ends after 2 weeks and were completely covered with newly formed bone by 3 weeks (Fig. 3B, C). Six weeks after injury, the sagittal suture was restored to its normal morphology (Fig. 3D). These results indicate that the sagittal suture is capable of regenerating itself even after complete removal.
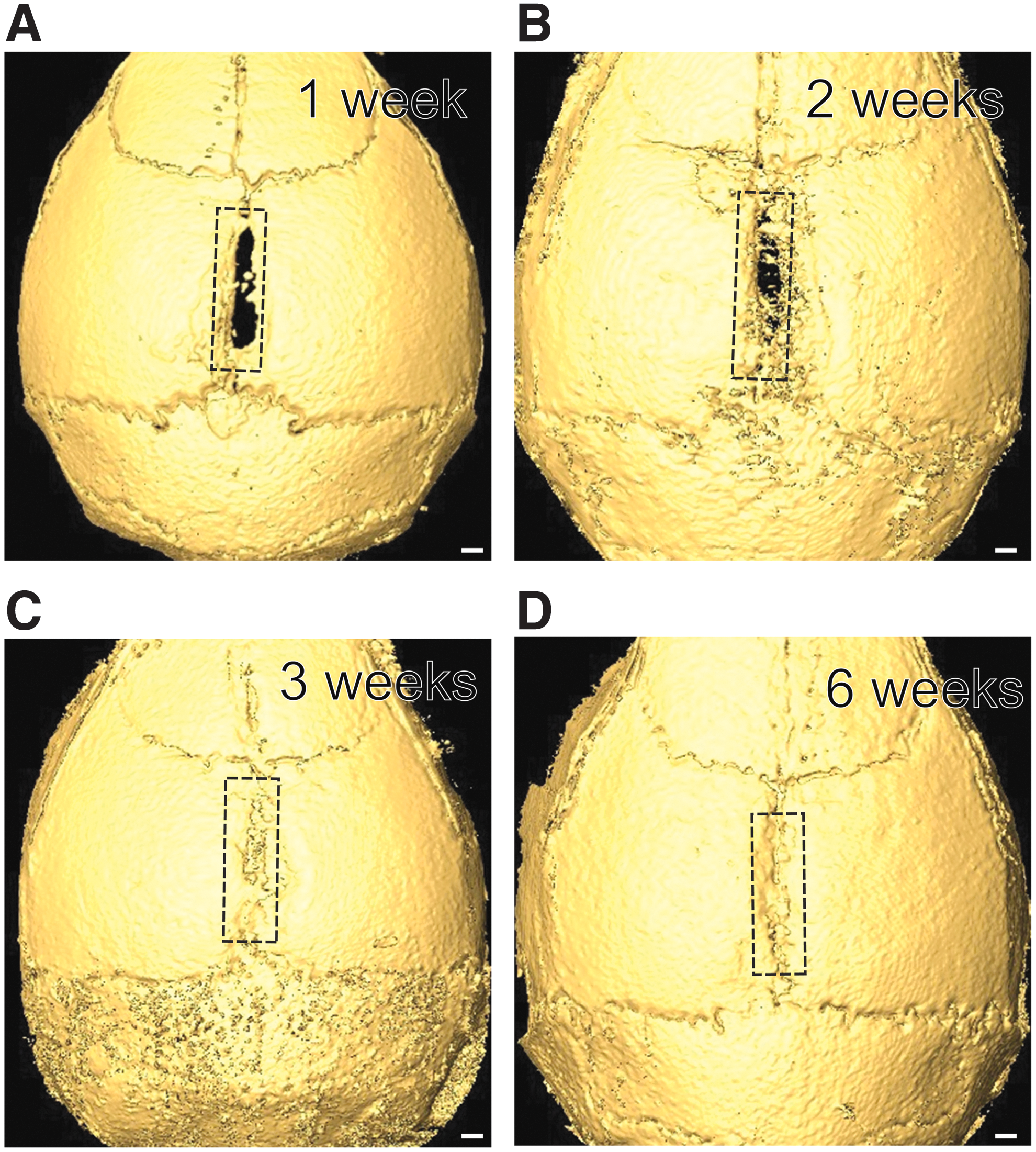
Regeneration of sagittal sutures after complete removal.
We also examined the effect of distance between the injury site and the sagittal suture on regeneration of the calvarial bone. Round holes, 2 mm in diameter, were drilled at varying distances from the sagittal suture (n = 5 per group) (Fig. 4A). One month later, we found that injuries to the suture were completed healed. Injury sites 0.5 mm from the suture were ∼80% filled with bone tissue (Fig. 4B, D), whereas injury sites 1 mm from the suture were ∼50% healed (Fig. 4C, D). Injury sites 2 mm from the suture were <20% healed (Fig. 4C, D). These results indicate that the healing capacity of the calvarial bone is inversely proportional to the distance of the injury site from the suture.
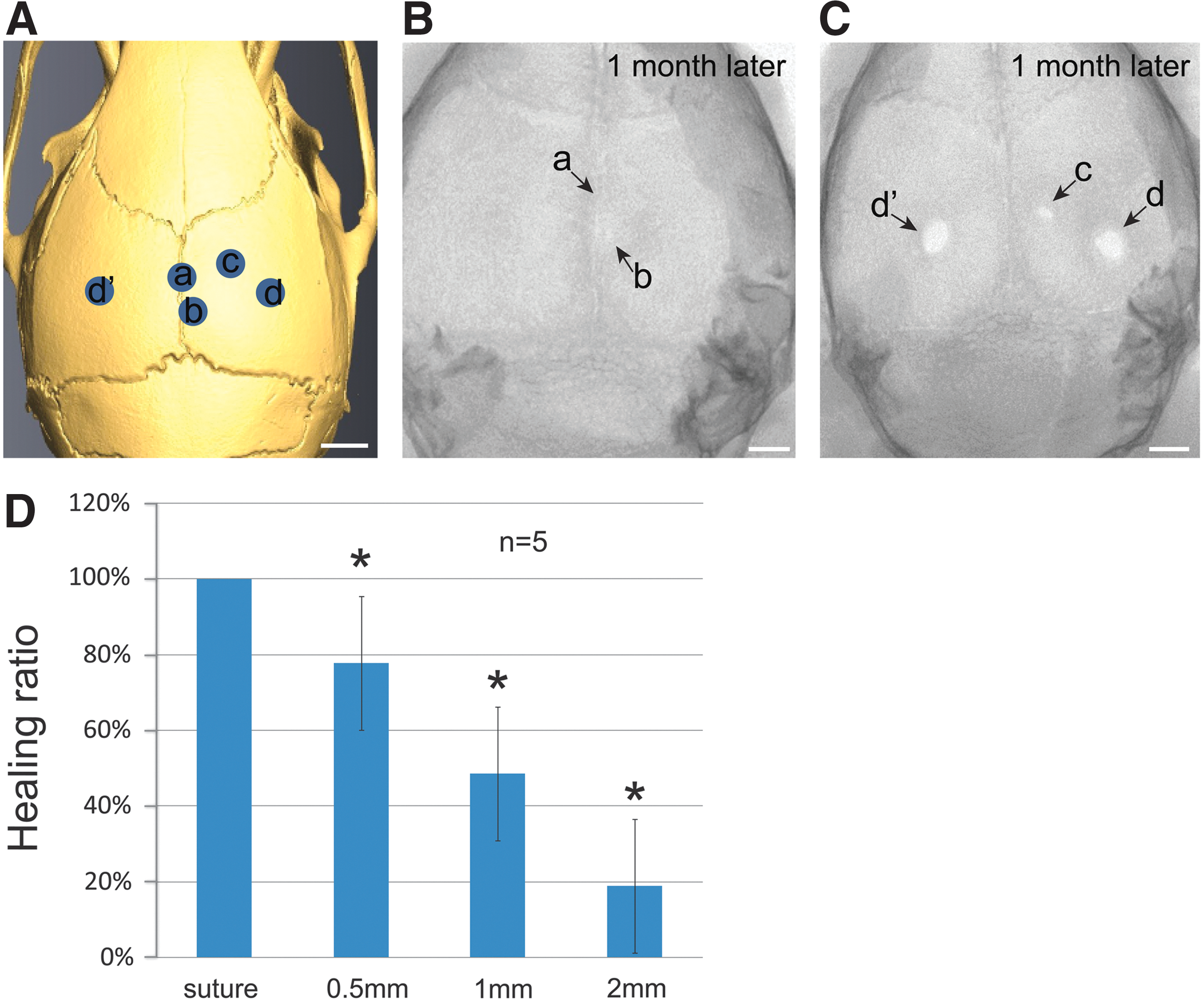
The healing capacity of an injury site is inversely proportional to its distance from the suture.
To investigate the cellular response to injury, we analyzed cell proliferation in suture and nonsuture regions. We performed EdU incorporation analysis and found that few proliferating cells were present within the suture mesenchyme in control mice (Fig. 5A). Upon injury, cells within the suture mesenchyme responded rapidly and proliferated within 48 h (Fig. 5B, C). In contrast, nonsuture regions contained few proliferating cells after injury (Fig. 5B, D). These results suggest that stem cells within the suture are activated rapidly after injury. The lack of increase in cell proliferation in nonsuture regions suggests that cells for repair must be recruited from another site, likely the suture.

The suture responds to injury in a manner distinct from nonsuture regions.
To test whether Gli1+ cells within the suture mesenchyme can migrate to support injury repair in nonsuture regions, we induced Gli1-CreERT2;Tdtomatoflox mice at 1 month of age with tamoxifen and created an injury to the parietal bone 1 mm from the suture. Two weeks later, Gli1+ cells were detectable between the injury site and the suture, consistent with migration of these cells toward the injury site (Fig. 5E). Injury experiments performed on the sagittal sutures of Gli1-LacZ mice at 1 month of age indicated that Gli1+ cells persisted in the suture region after complete healing (Fig. 5F). Thus, our results indicate that Gli1+ cells within the suture mesenchyme can respond to injury rapidly and migrate to support injury repair, whereas the nonsuture calvarial bone does not possess such capability.
Finally, to determine whether suture stem cell-mediated calvarial bone injury repair is unique to mice, we compared the healing rates of injury sites in different regions of the skull bone in rabbits. Rabbits represent a commonly used large animal model for the study of craniofacial bone tissue homeostasis and repair [19]. Round holes, 4 mm in diameter, were drilled either in the center of the parietal bone or the sagittal suture. One month later, injuries to the suture were almost healed completely, whereas injury sites in the center of the calvarial bone had healed poorly (Fig. 6). Thus, our results indicate that the sutures of the rabbit calvarial bone also possess stronger regenerative capacities than nonsuture regions, confirming the results we obtained in mice.
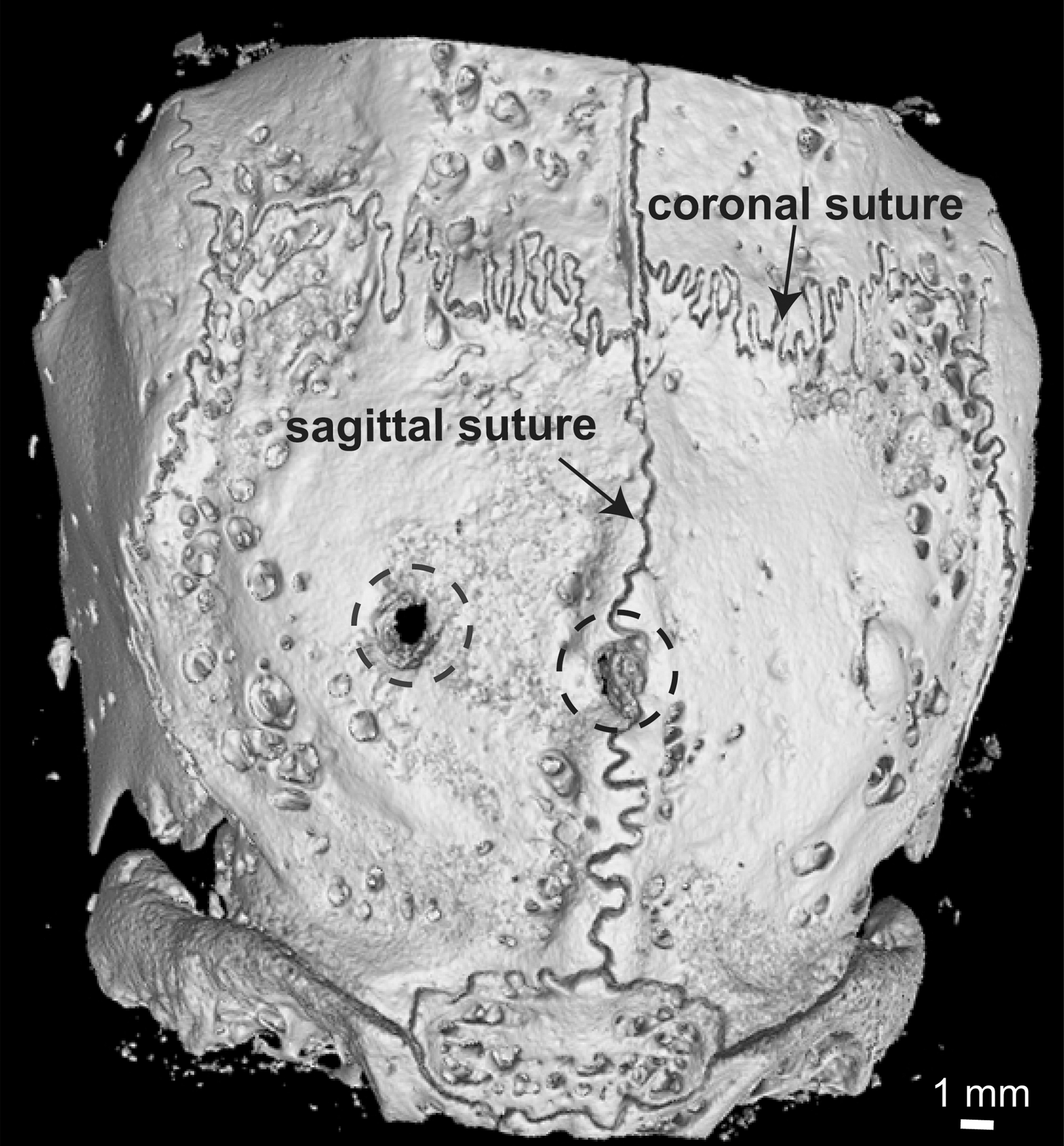
Rabbit calvarial sutures possess stronger regenerative capacity than nonsuture regions. MicroCT imaging of calvarial bones 1 month after injury of 2-month-old rabbits. Holes, 4 mm in diameter, were drilled in the parietal bones. One injury was centered on the sagittal suture and the other on the parietal bone. Dotted circles outline the injury locations. Scale bar, 1 mm.
Discussion
In our current study, we have found that calvarial sutures possess stronger regeneration capacity than other regions of the calvarial bone and that the regenerative capacity of the calvarial bone diminishes with increasing distance from the suture. Our findings support and extend our previous findings that Gli1+ cells within the suture mesenchyme are mesenchymal stem cells supporting craniofacial bone turnover and injury repair [18]. The healing of a calvarial bone injury is not an evenly distributed process on the calvarial bone. The regenerative capacity of calvarial bone diminishes with increasing distance from the suture. The presence of resident stem cells within the suture is likely the reason for the stronger regeneration capability of the suture. These stem cells can be immediately activated into proliferation upon injury and migrate to support the injury repair of nonsuture regions of the calvaria.
Our study provides a reasonable explanation for the presence of CSDs. A CSD is defined as a large bony defect that cannot heal without the assistance from a reparative material [13]. Critical-sized calvarial defects are typically managed through reconstruction with metal implants or other materials [20]. Because the majority of the stem cell population of the calvarial bone resides within the suture, the ability of these mesenchymal stem cells to respond and migrate into an injury area in sufficient numbers for repair is dependent upon the distance of the injury site from the suture.
Strikingly, we found that the sagittal suture can regenerate after complete removal. The newly regenerated suture was restored to its normal configuration and the Gli1+ population returned to a normal level within 2 months after injury, suggesting maintenance of the stem cell population after regeneration. The cellular source for these stem cells could be other neighboring sutures, including the coronal and lambdoid sutures. This model is based on our observation that the closure of the suture wound was initiated from both ends. Furthermore, our recent study has shown that loss of Gli1+ stem cells within the suture leads to compromised healing of calvarial bone injury [18]. The regenerative property of the suture makes it a potential donor tissue for transplantation. In the future, we may be able to utilize autologous sutures as seeds for the repair of large calvarial bony defects.
Calvarial bone injuries have been widely used as models for analyzing the bone repair process and for evaluating tissue engineering approaches due to their accessibility and amenability to direct visualization. Injuries of different sizes or types are made to the calvarial bones and various materials or medications are applied to the injured area for comparison and evaluation [19]. Previous studies showed that the frontal bone and parietal bone possess differential regeneration capability due to their different developmental origins or endogenous fibroblast growth factor levels [16,21,22]. Our current study suggests that the suture responds to injury in a manner distinct from that of other areas of the calvarial bone. Therefore, the injury location on the calvarial bone is an important factor to be considered when designing experiments.
Surgical removal of the suture, followed by reshaping of the calvarial bones, is currently the standard approach for treating patients with craniosynostosis. The purpose of the surgery is to form artificial space between the calvarial bones to allow for future brain growth and expansion. Surprisingly, the natural suture tissue is routinely discarded as surgical waste during these procedures [23,24]. Our results strongly suggest that this surgical practice should be reevaluated due to the essential roles of sutures in calvarial bone tissue homeostasis and regeneration.
Footnotes
Acknowledgments
The authors thank Julie Mayo and Bridget Samuels for critical reading of the manuscript. H.Z. acknowledges training grant support from the National Institute of Dental and Craniofacial Research, NIH (R90 DE022528). This study was supported by grants from the National Institute of Dental and Craniofacial Research, NIH (DE022503, DE020065, and DE012711) to Y.C.
Author Disclosure Statement
No competing financial interests exist.