
Abstract
Duchenne muscular dystrophy (DMD) is a lethal X-linked disorder caused by null mutations in the dystrophin gene. Although the primary defect is the deficiency of muscle dystrophin, secondary events, including chronic inflammation, fibrosis, and muscle regeneration failure are thought to actively contribute to disease progression. Despite several advances, there is still no effective therapy for DMD. Therefore, the potential regenerative capacities, and immune-privileged properties of mesenchymal stromal cells (MSCs), have been the focus of intense investigation in different animal models aiming the treatment of these disorders. However, these studies have shown different outcomes according to the sources from which MSCs were obtained, which raise the question whether stem cells from distinct sources have comparable clinical effects. Here, we analyzed the protein content of the secretome of MSCs, isolated from three different sources (adipose tissue, skeletal muscle, and uterine tubes), obtained from five donors and evaluated their in vitro properties when cocultured with DMD myoblasts. All MSC lineages showed pathways enrichment related to protein metabolic process, oxidation-reduction process, cell proliferation, and regulation of apoptosis. We found that MSCs secretome proteins and their effect in vitro vary significantly according to the tissue and donors, including opposite effects in apoptosis assay, indicating the importance of characterizing MSC secretome profile before its use in animal and clinical trials. Despite the individual differences a pool of conditioned media from all MSCs lineages was able to delay apoptosis and enhance migration when in contact with DMD myoblasts.
Introduction
P
Despite countless advances, there is still no effective therapy for PMDs [3]. Therefore, the potential regenerative capacity, and immune privileged properties of mesenchymal stromal cells (MSCs), have been the focus of intense investigation in different animal models of PMDs [4 –7] aiming the treatment of these disorders. However, these studies have shown different effects according to the sources from which MSCs were obtained [8]. Distinct methodologies to generate MSC lineages and even cryopreservation could modify their proliferation rate, adhesion, and secreted proteins, which would explain the distinct results obtained by us and by several groups [9 –11].
Different reports have confirmed that, even when there is no migration to the site of injury, MSCs exert therapeutic benefits, which could be related to angiogenesis, immune modulation, and regulation of apoptosis [12 –14]. In this scenario, MSCs would work as a local factory of mediators, stimulating cell proliferation and survival in host tissues. In summary, there is a growing body of evidence supporting the paracrine activity of MSCs and suggesting differences in their therapeutic potential according to the tissue that originated from each strain. However, the mechanism by which MSCs act on dystrophic muscle improving the clinical phenotype is still unknown. Thus, the identification of secreted molecules by different MSCs through the analysis of their conditioned media (CM) and their effect on PMDs muscle cells is of great interest [15 –17].
Here, we compared the MSCs secretome protein profile obtained from different tissues and different donors, grown under the same conditions, and their in vitro effect when cocultured with DMD myoblasts. We found that MSCs secreted proteins and their effect in vitro vary significantly according to the tissue and donors and that functional assays with cells obtained from different individuals may show opposite results. The analysis of differently expressed proteins and enriched pathways is of utmost importance to enhance our comprehension on MSCs mode of action. Furthermore, the analysis of their profile and their variability is imperative to optimize their use in therapies for PMDs in a robust and safe manner.
Materials and Methods
Subjects
Tissue samples were obtained from three tissues (adipose tissue, muscle, and uterine tubes) from five women submitted to hysterectomy after written informed consent from the donor's parents. Myoblasts from the DMD patient were obtained after written informed consent from the donor's parents. All research was carried out in the Human Genome and Stem Cell Research Center (Biosciences Institute, University of São Paulo) and at the Research and Development Center from Fleury Group. The project was approved by the Ethical Committee of the University Hospital from the University of São Paulo.
Selection of cell lines
MSCs were obtained from five women undergoing hysterectomy abdominal surgery when it was possible to obtain different tissues. To compare whether the variability between MSCs tissues was related to the sources' origin or from their donors we analyzed different tissues from the same individuals. Due to the interest in investigating the secretome profile of MSCs previously used in preclinical studies in animal models for neuromuscular disorders we focused on adipose tissue and muscle samples [18,19]. Uterine tubes samples were included to further investigate tissue variability. MSCs immunophenotypic profiles were evaluated by flow cytometry [6,20] (Supplementary Data; Supplementary Data are available online at
Cell culture and CM production
The cells were cultured until they reached passage 6. They were plated in six 225 cm2 bottles (Corning) at a concentration of 5,000 cells per cm2 and maintained until a confluence of ∼80%. When collecting the CM, cell viability was assessed by dilution in a solution of trypan blue 0.4% (Life Technologies) and quantification using automated cell counter Countess (Invitrogen). Dulbecco's modified Eagle medium (DMEM) low glucose with 10% HyClone (Thermo Scientific) and 1% antibiotic was used. The cells were washed three times with phosphate-buffered saline and then incubated with 32 mL of culture medium per bottle without fetal bovine serum (FBS). After 8 h of conditioning, a new washing for cell lysates removal and further conditioning was done for 20 h, which was used for further analysis.
After the conditioning period, the culture medium was centrifuged at 500g for 10 min to remove cellular debris, and immediately filtered on a 0.22 μm membrane (Steritop & Stericup; Millipore). The protein content was concentrated from the CM using Amicon Ultracel 3K (Millipore) following the manufacturer's protocol to a final volume of 1.5 mL. DMEM was subjected to the same process and the resultant volume (concentrated DMEM) was used as a control in further experiments. The protein concentration recovered from the CM was quantified by the fluorescence based on the natural tryptophan fluorescence [21].
Glycoprotein enrichment and proteomic analysis
All cell lines were subjected to the isolation of glycosylated proteins by affinity to concanavalin A and wheat germ agglutinin followed by tryptic digestion. Coupled liquid chromatography mass spectrometry (LC-MS/MS) were performed on a nanoAcquity (Waters) coupled to a Q-Exactive hybrid mass spectrometer (Thermo) (for further details, see Supplementary Materials and Methods).
The acquired data were processed by MaxQuant 1.4.0.8 proteomics data analysis workflow [22]. Protein identification was performed by Andromeda search tool using the database of the human proteome UniProtKB (SWISSPROT, November 2014). The following criteria were applied to protein identification: allowed maximum of two incomplete cleavages by trypsin, fixed modification by carbamidomethylation of cysteines, and variable modification by acetylation of the N-terminal portion and methionine oxidation.
Quantification was based on two label-free methods: Label Free Quantification (LFQ) [23] and an adaptation of iBAQ strategy [24] in which UPS standards (universal proteomics standard, Sigma Aldrich) were replaced by a mixture of 4 fmol of yeast enolase and 40 fmol of yeast alcohol dehydrogenase (MassPREPWaters™) added to samples before LC-MS/MS to obtain two points calibration.
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE [25] partner repository with the dataset identifier PXD003978.
DMD migration assay
DMD myoblasts at fourth passage were first stained with CFDA-SE (Life Technologies) as per manufacturer's instructions. Briefly, the cells were harvested using trypsin, washed and incubated with the dye (5 mM) for 20 min. Subsequently, the cells were plated (2 × 104 cell per well) in a 96 wells Oris cell Migration assay and incubated overnight with growth media (DMEM 10% FBS 1% anti-anti; Gibco). The inserts were then withdrawn, the cells were washed twice with DMEM. Then, three sets of experiments were performed: (1) 25 μg of CM from pools of each tissue (adipose, tube, or muscle) were added and completed up to 100 μL of solution with DMEM low without FBS; (2) 25 μg of CM from the three tissues from the same donors were added and completed to 100 μL of solution with DMEM low without FBS; (3) 25 μg of all 15 samples were added and completed up to 100 μL of solution with DMEM low without FBS. Concentrated DMEM was used as control. The area free of myoblasts was recorded by the acquisition of images of the cell cultures at time point zero and 24 h, using the AxioVision LE software, version 4.8.2.0.
Statistics and enriched pathway analysis
Proteomic LFQ was analyzed by one-way analysis of variance (ANOVA) followed by t-test comparing all groups against each other in Perseus module (version 1.2.0.17). Threshold value for the significance was set to 0.05. Hierarchical clustering was performed using the Euclidean distance and the average linkage based on differentially expressed proteins of different lineages. Time lapse acquisition was tested using two-way ANOVA comparing all groups against each other. Migration assay was analyzed by one-way ANOVA comparing all groups against each other. All analysis were done with Bonferroni correction. Enrichment pathways analysis was carried out with the assistance of DAVID software and highlighting the seven pathways with higher P value given by GOTERM_BP [26]. Venn's diagrams were made using
Results
Establishment and characterization of cell lines
All cell lines (passage No. 6) were negative for CD31 (endothelial cell marker), CD34, CD45, CD117 (hematopoietic cell markers), and HLA-DR (human leukocyte differentiation antigen class II) expression, but positive for the expression of CD29, CD44, CD90, (adhesion markers) CD73, CD13 (mesenchymal markers), and HLA-ABC (human leukocyte differentiation antigen class I). Cell plasticity was assessed by in vitro differentiation capacity (Supplementary Fig. S1). These results confirmed the mesenchymal nature of the isolated cells and their multipotent potential. Additionally, the viability of cells after FBS deprivation was 95%—99%, for all lineages.
MSCs secretomes are able to induce cell migration and protection against apoptosis when cocultured with DMD myoblasts
Since one of the properties described for MSC is the regulation of apoptosis and cellular migration [27] we investigated whether MSCs' secretome (pooled from of all our samples) have the ability to modulate those cellular functions in vitro. For this experiment, we used myoblasts derived from a DMD patient and a read out based on the activation of apoptotic process through tert-butylhydroperoxide (Fig. 1) [28].
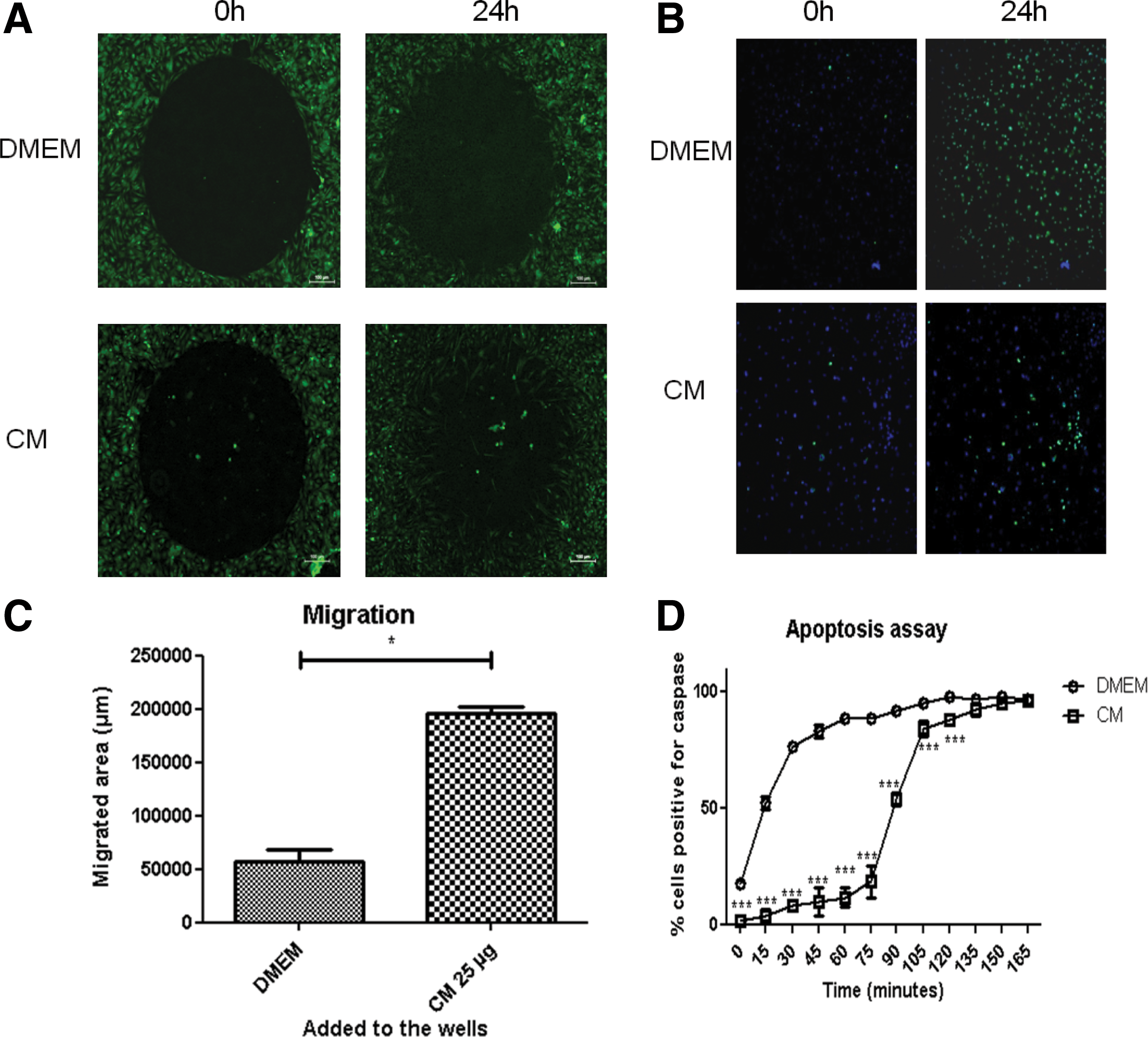
CMs in vitro evaluations. Effect of a pool of secretomes in DMD myoblasts migration and apoptosis.
When added to the culture media, the secretome pool significantly increased the migration and reduced the apoptosis process of human DMD myoblasts (Fig. 1), as compared to the control (concentrated DMEM). We next investigated whether the migration and apoptosis delay was due to proteins exclusive to a particular tissue or donor. To address this question we incubated DMD myoblasts with either a secretome pooled from different tissues or from different donors. As shown in Fig. 2, the ability of both the tissue of origin, and different donors in modulating cell migration and apoptosis of human DMD myoblasts differed significantly when compared to the control. This difference was more pronounced according to the donors than according to the tissues, as illustrated by the apoptosis assay (Fig. 2A–C).
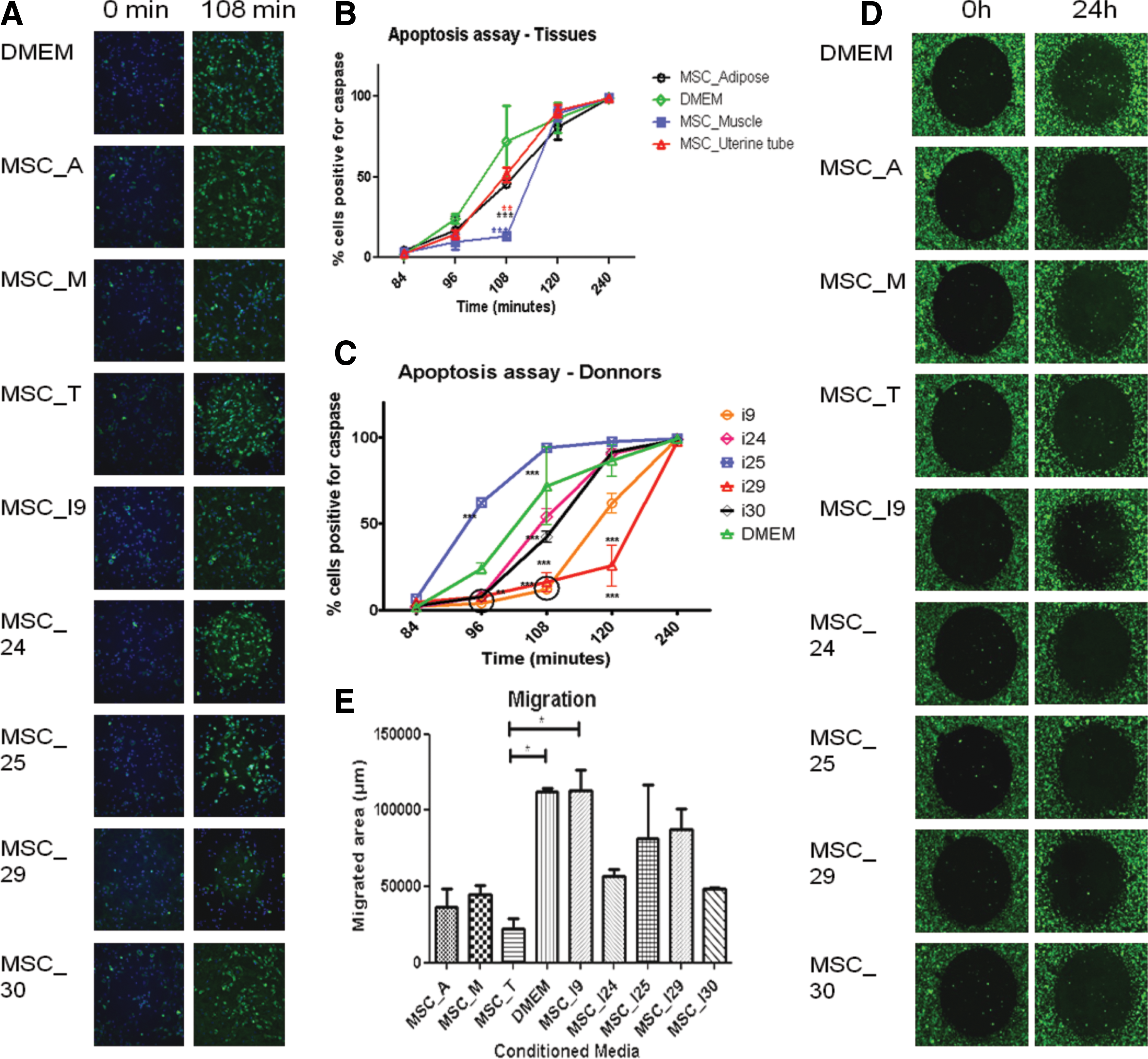
Effect of tissue of origin and donors in the modulation of cell migration and apoptosis by different secretome samples. MSC_A, _M, _T: secretomes pooled regarding the tissue of origin, from adipose, skeletal muscle, or uterine tube, respectively. MSC_I9, _I24, _I25, _I29, and _I30: secretomes pooled regarding donors 9, 24, 25, 29, and 30, respectively.
Since there was no significant effect on migration, we focused our analysis in the apoptosis read out. To investigate the proteins secreted by MSC that may be responsible for the observed apoptosis delay, all CM were analyzed by shotgun proteomics. Furthermore, to evaluate the secreted proteins, which are expected to suffer post-translational modification, we isolated glycoproteins, as previously described [29]. This strategy allowed the identification of 1,079 proteins: 618 proteins were identified in the total CM, 155 in the glycoproteins and 306 proteins in both (Supplementary Tables S1 and S2).
Donors' secretomes
When we compared the secretome of all five donors a large number of shared proteins were found in all samples (Fig. 2B) despite a significant differential expression between individuals that was further analyzed (Supplementary Tables S3–S6).We next analyzed the enriched pathways in secretomes that delayed apoptosis (individuals I9, I24, I29, and I30) as compared to the individual I25, whose secretome enhanced apoptosis (Figs. 2A, C and 3E).
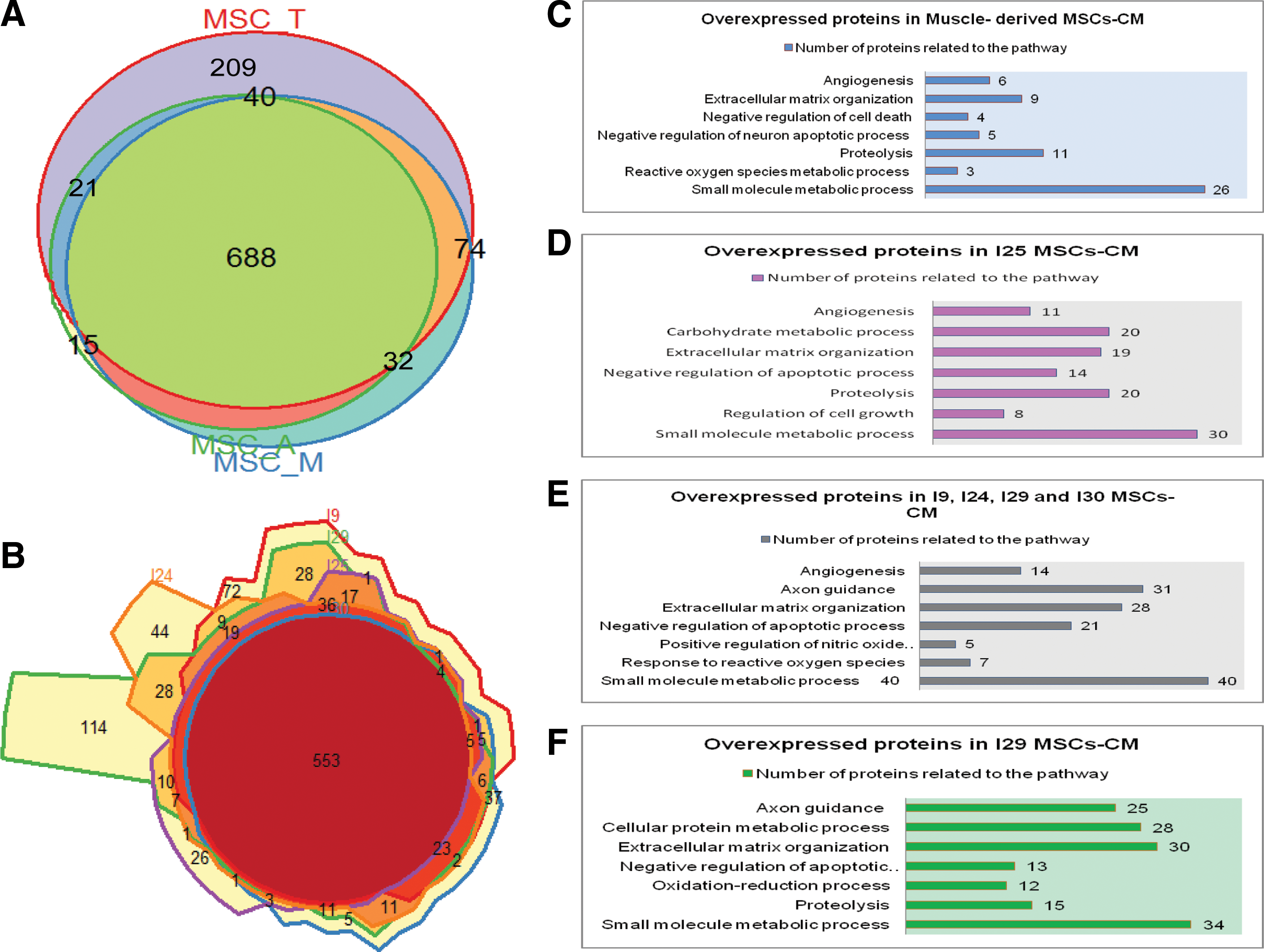
Comparison of proteins between different tissues or donors. Comparison of proteins between different tissues n = 5 biological replicates each tissue
Particularly, the striking differences on the percentage of nuclei caspase 3/7 positive cells (which is a marker of apoptosis) between two individuals, donor I25 and donor I29 called our attention. After 108 min of incubation, 100% of the cells were caspase-positive when cultured with individual I25 secretome but only 10% of the DMD myoblasts showed this activation in response to individual I29 secretome (Fig. 2A, C). When we compared these samples, we observed that some enriched pathways related to protein metabolic process, axon guidance, oxidation-reduction process and positive regulation of nitric oxide (NO) biosynthesis were present only in the secretomes that delayed apoptosis (Fig. 3C–F).
Furthermore, in one sample, from individual I29, which delayed apoptosis more efficiently than the MSCs-CM from other donors, we identified pathways related to small molecule and protein metabolic process, proteolysis, extracellular matrix organization, negative regulation of apoptosis, axon guidance, and oxidation-reduction process (Fig. 3F). It is noteworthy that one of the highly expressed proteins was jagged-1.
Tissues' secretomes
The comparative analysis of the tissue source showed a significant difference in the apoptosis delay between the control and all sources of CM (adipose, muscle, and uterine tubes), which was more pronounced for muscle-derived MSCs (Fig. 2A, B).
Looking for the differences in the protein profile between the different tissues, we observed that protein content in MSCs CM was very similar among the different sources of MSCs (Fig. 3A), since they share 63% (688/1,079) of the identified proteins.
In the apoptosis assay, we could observe that muscle-derived MCSs-CM delayed apoptosis more efficiently than the other sources. Then, we investigated pathways related to proteins that were significantly more expressed in the muscle-derived MSCs-CM compared to adipose and uterine tube-derived MSCs-CM (Supplementary Tables S4 and S6). We identified enrichment for pathways related to small molecule metabolic process, proteolysis, extracellular matrix organization, negative regulation of cell death and apoptosis of neurons, angiogenesis, and reactive oxygen species (ROS) metabolic process (Fig. 3C).
MSCs according to the donor or tissue of origin
A total of 83 MSCs proteins were present in all our samples whose pathways enriched were related to immune response, protein metabolic process and stabilization, oxidation-reduction process, gene expression, cell proliferation, and regulation of apoptosis (Fig. 4A and Supplementary Table S7).
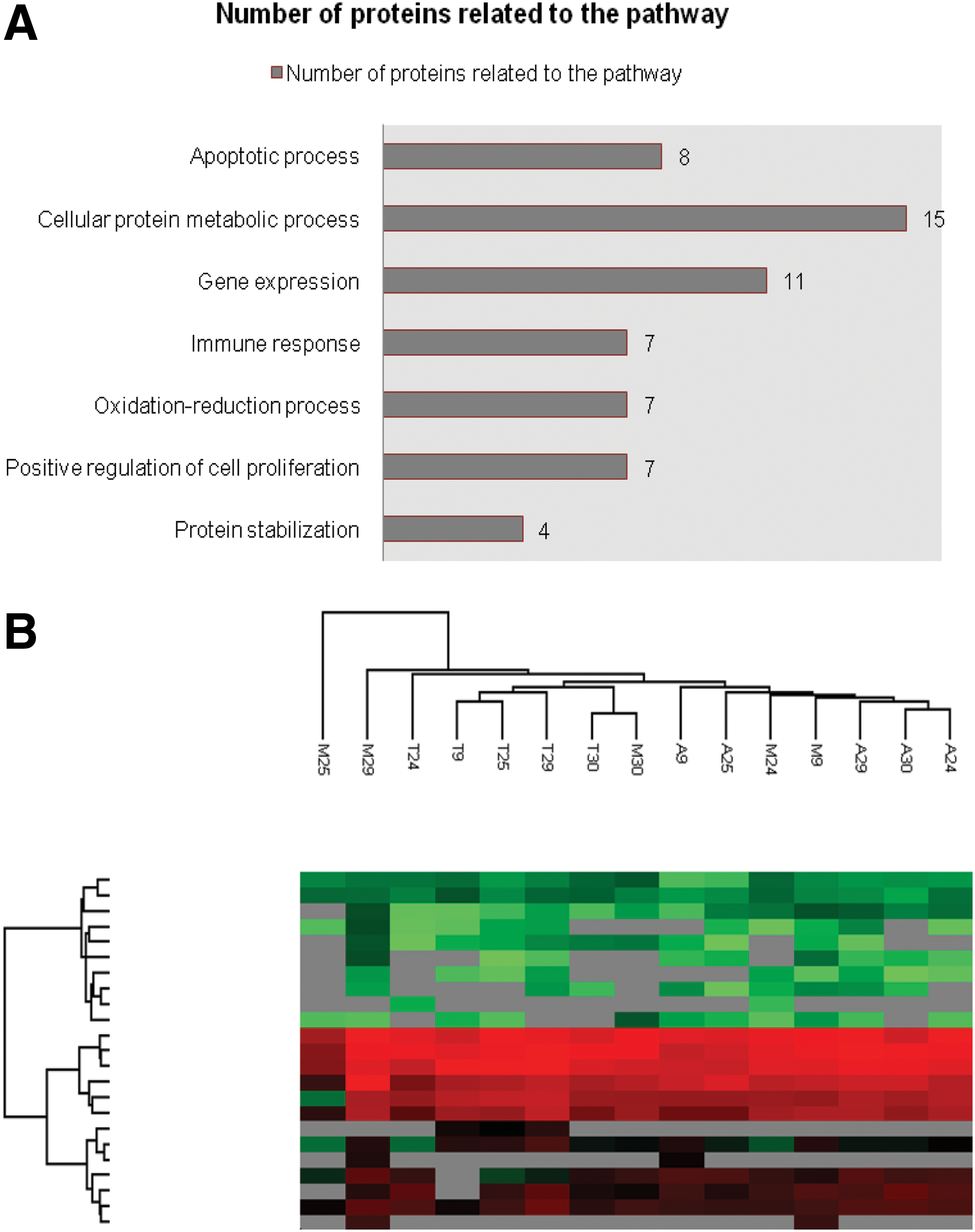
Analysis of enriched pathways and proximity between cell lines. Enriched pathways of proteins of MSCs CM
We next asked whether the MSCs lineages could be clustered either by donor or tissue of origin based on their protein expression pattern. Using the Euclidean distance and the average linkage we generated a hierarchical clustering based on differentially expressed proteins of different lineages. This analysis showed that the proximity of the cell lines CMs does not depend exclusively on the donors or exclusively on the tissue of origin (Fig. 4B).
Total CM and micro vesicle protein content
To further investigate whether the proteins found in MSCs CM are secreted through exosomes, we used shotgun proteomics to evaluate the protein content from CM and microvesicles (MVs), obtained from the same MSC lineage (Supplementary Fig. S2 and Supplementary Table S8).
We found 193 proteins in common between CM and MVs (Supplementary Fig. 2A), which are related to cell adhesion, migration, wounding response, and negative regulation of apoptosis, among other functions. A total of 867 proteins mainly related to cell homeostasis and metabolic functions were exclusive to the CM, suggesting that most proteins present in the complete CM are not secreted by exosomes.
MVs protein content was less variable as compared to total CM profile, and they are related to the same functions as the proteins shared by CM and MV (Supplementary Figs. S2A–D and S3).
Discussion
The present results have demonstrated the secretome variability of MSCs sourced from different human tissues and donors. These findings highlight the importance of matching the secretome from donor MSCs to the recipient to develop a successful stem cell therapy for DMD. It may also be important to closely match the genetics, viability, and plasticity of donor MSCs to the recipient, as mismatches could hinder progress. Comparative analysis of MSC obtained from distinct sources from the same donor and therefore the same genetic background are very difficult to perform due to tissue availability. Here, we have isolated MSCs from three different biological sources, from five women undergoing hysterectomy, and analyzed their in vitro therapeutic potential in DMD myoblasts cocultures.
We show for the first time that the secretome from MSCs is able to modulate the apoptosis process of DMD myoblasts in vitro. However, the heterogeneous nature of a given MSC population could explain the significant differences between donors and tissues and previously reported preclinical discordant results. Secretome protein content analysis through shotgun proteomics confirmed the variability among samples and hierarchical clustering showed that there was no greater proximity between MSCs lines according to specific tissues origin or in accordance with donors. According to Mo et al. (30) the percentage or even the presence or absence of a particular subpopulation within an MSC population can alter their modulatory potential and secretory profile.
It is known that the secretory profile of MSCs can be altered according to the composition of the microenvironment [31,32]. Although it is possible that the “in vitro” secretory profile of MSCs differs from that observed “in vivo,” our results show that the CM obtained without any stimulation significantly increased cell migration and delayed the apoptosis process in human DMD myoblasts (Fig. 1). Other “in vivo” studies have also reported better skeletal muscle regeneration with secretome injections as compared to cell transplantation [33]. Moreover, the importance of in vitro MSCs secretome characterization and its consequences on in vivo outcome of cell therapy has been shown in other studies such as Kim et al. who identified some molecules in MSCs cultures that act as predictors of response to autologous stem cell transplantation in patients with amyotrophic lateral sclerosis [34]. These results highlight the importance of a careful in vitro characterization of MSCs secretome for therapeutic use.
One of the secondary mechanisms that contribute DMD progression is the depletion of muscle progenitor's pool due to repeated cycles of degeneration/regeneration. In addition to the pool depletion caused by the extensive cell division, increased apoptosis might contribute to the failure of regeneration [35]. Here, the reduced apoptosis in the coculture of MSCs secretome and DMD myoblasts provides an explanation for the beneficial effect of MSCs transplantation in animal models for PMD [18,33]. Additionally, the proteins identified in the MSCs CM showed enriched pathways that could be related to these beneficial effects.
Besides negative regulation of apoptosis, enriched pathways related to extracellular matrix organization, axon guidance, antigen processing, protein metabolic process, oxidation-reduction process, and positive regulation of NO may also play an important clinically beneficial role. NO is a diffusible short-lived free radical and a signaling molecule with a number of important physiological functions such as smooth muscle relaxation, neurotransmission, inhibition of platelet aggregation, and host defense mechanisms. It also plays a role in the pathology of several inflammatory diseases and other pathological conditions such as cancer, diabetes, and neurodegenerative diseases [36].
Lai et al. demonstrated that neuronal nitric oxide synthase (nNOS) was able to partially restore cardiac function in aged mdx mice, a murine model of Duchenne cardiomyopathy [37]. Additionally, it was previously reported that various pathologies can result from oxidative stress-induced apoptotic signaling as a consequence from ROS increment and/or antioxidant decrease [38]. Previous reports demonstrated that the extracellular matrix protects cells against apoptosis [39,40]. In short, the proteins identified in MSCs secretomes are consistent with our findings according to which the CM from MSCs is able to protect cells from apoptosis, which could also explain previous results in preclinical studies [8,33].
In this study, many enriched pathways were found in common in all samples but a greater individual expression of some specific proteins could explain different clinical outcomes. For example, among the differentially expressed proteins found in individual I29, jagged-1 called our attention since we recently observed that overexpression of this protein rescued the Duchenne phenotype in GRMD (golden retriever muscular dystrophy) dogs [41].
When we compared the enriched pathways between the individuals that delayed with the one that enhanced apoptosis, the pathways present only in the secretomes that delayed apoptosis were found to be related to protein metabolic process, axon guidance, oxidation-reduction process, and positive regulation of NO biosynthesis (Fig. 3C–F). Further investigation of these pathways is of utmost importance aiming to enhance our comprehension on their putative role to reduce muscle degeneration and decrease the rate of progression in DMD or other forms of muscular dystrophy.
Current clinical trials aiming the treatment of PMDs (NCT02285673, NCT01610440, NCT02235844) with umbilical cord MSCs include transplantation of autologous or allogeneic cells obtained from different donors. The choice of cells that are currently used in trials is mainly based on the ease of collection and abundance. Since it is hypothesized that the predominant mechanism by which MSCs function is related to its paracrine activity our results indicate that the reasons by which the MSCs are chosen for a specific use must be beyond the ease of collection and abundance.
The present results have shown the effectiveness of MSCs to delay myoblast apoptosis in DMD, even when the MSCs have been sourced from different donors. The effectiveness of MSCs to delay myoblast apoptosis should be studied in more detail to identify the functional receptors, bioactive molecules, and molecular pathways, which might lead to the development of novel stem cell therapies, or drugs for the treatment of DMD and other degenerative diseases.
Footnotes
Acknowledgments
The collaboration of the following persons is gratefully acknowledged: Dr. Oswaldo Keith Okamoto, Karina Helena M. Cardozo, Carlos Frederico Martins, Mariz Vainzof, Maria Rita dos Santos e Passos Bueno, Vanessa Sato, Wagner Falciano, Mariane Secco, and Fabio Mitsuo. FAPESP, CNPq, AACD. This work was supported by FAPESP-CEPID under award no. 2013/08028-1, CNPq under award No. 705019/2009, INCT under award no. 2008/578997, AACD, and Fleury.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.