
Abstract
The origin and evolution of molecular mechanisms underlying the self-renewal and differentiation of spermatogonial stem cells (SSCs) are fundamental questions in stem cell biology as well as reproduction medicine. In mammals, glial cell line-derived neurotrophic factor (GDNF) is crucial for SSC self-renewal and maintenance. However, in nonmammals, the role of Gdnf in SSCs still remains unknown. In this study, we report that the two GDNF homologs from medaka fish (Oryzias latipes), namely OlGdnfa and OlGdnfb, can promote proliferation activity and retain the spermatogonial property of SG3, a spermatogonial cell line derived from adult medaka showing the intrinsic property of SSCs by self-renewal and differentiation potential during 2 years of culture. Cloning and sequencing led to the identification of two cDNA sequences as Olgdnfa and Olgdnfb, which are 780-nt and 744-nt in length for 253 and 245 amino acid residues, respectively. Both are homologs of mammalian GDNF and share over 45% identity with the other known vertebrate homologs. Importantly, in a well-defined condition, the recombinant proteins, OlGdnfa and OlGdnfb, can significantly promote the proliferative activity of SG3 cells and retain the spermatogonial gene expression pattern and alkaline phosphatase activity. Meanwhile, both of the two recombinant proteins can upregulate the mRNA expression level of bcl6b, one of the prominent GDNF-regulated genes involved in SSC self-renewal and maintenance in mammals. Taken together, our findings suggest that just like the mammalian counterpart, the nonmammalian Gdnfs might mediate the self-renewal and maintenance of SSCs; moreover, Bcl6b might be a conserved regulator in SSC self-renewal across vertebrate taxa. This study extends our knowledge of GDNF functions in SSC biology during evolution.
Introduction
S
Comprehensive bioinformatic analyses by us reveal that gdnf homologs widely exist in teleost fishes; additionally, two homologs might be present in several fishes, including fugu (Takifugu rubripes), Nile tilapia (Oreochromis niloticus), and medaka (Oryzias latipes). Recent studies have shown that gdnf in rainbow trout (Oncorhynchus mykiss) is highly expressed in type A spermatogonia and the gdnf-gfra1 pathway is expressed in a spermatogenetic-dependent manner [17,18]. Moreover, studies indicate that the expression of Gfra1 occurs limitedly in undifferentiated spermatogonia from rainbow trout, Nile tilapia, and dogfish (Scyliorhinus canicula) [19,20], suggesting that this signaling pathway might imply SSC development. Although previous studies have shown that a cocktail of recombinant mammalian growth factors containing GDNF enhances the proliferation of spermatogonia from fish species, including zebrafish and dogfish [21,22], no effect of rat GDNF or rat GFRA1-Fc fusion protein on the proliferation of rainbow trout type A spermatogonia has been reported [23]. To date, functional studies using fish Gdnf have not been conducted. Therefore, the role of Gdnf in this vertebrate class is not yet elucidated.
A culture system for SSCs in a well-defined culture condition is enormously valuable to identify crucial growth factors for SSC regulation and investigate the underlying mechanisms (such as self-renewal vs. differentiation). In medaka, a spermatogonial stem cell line (SG3) derived from a mature testis was successfully established in a feeder-free culture condition by Hong et al. [24], which is also the only fish spermatogonial cell line to date. The SG3 cell line retains a normal karyotype and shows the defining property of SSCs by long-term, self-renewing cell divisions and sperm production in vitro over 140 passages during 2 years [24]. However, what factor (s) is crucial for the proliferation and maintenance of SG3 cells remains unknown. In the present study, two Gdnfs from the medaka, namely OlGdnfa and OlGdnfb, were first characterized and then obtained through prokaryote expression, purification, and renaturation. Most importantly, further experiments indicate that both the recombinant proteins, OlGdnfa and OlGdnfb, can mediate the self-renewal and maintenance of SG3 cells and upregulate the expression of bcl6b in vitro.
Materials and Methods
Animals
Medakas were kept in recirculating aerated water tanks at 26°C. All animal experiments were carried out in accordance with the Guidelines for Care and Use of Laboratory Animals prescribed by the Regulation of Animal Experimentation of Chongqing, China.
Complementary DNA cloning
Total RNAs were extracted from the brain and testis of adult medaka using RNAiso PLUS (Takara) according to the manufacturer's instructions. The quality and concentration of total RNAs were analyzed by agarose gel electrophoresis and optical density reading at 260 and 280 nm. One microgram of total RNA was reverse transcribed into cDNA using the PrimeScript II 1st Strand cDNA Synthesis Kit (Takara).
By searching the GenBank, two putative sequences with GenBank accession numbers, XM_004074776.2 and XM_004072340.2, were obtained. After comprehensive bioinformatic analysis, PCR primers for the two gdnfs in medaka were designed (Supplementary Table S1; Supplementary Data are available online at
Sequence analyses
After the cDNA sequences containing the full open reading frame of the two gdnfs were obtained, the similarity with other known sequences was analyzed by BLAST program (
Reverse transcription PCR
Total RNAs were extracted from groups of 10 embryos at the desired stage and adult dissected tissues (including brain, eye, heart, spleen, liver, intestine, ovary, and testis) using TRIzol reagent (Takara). Total RNAs were prepared individually and reverse transcribed to cDNAs. Primer sets were previously validated (Supplementary Table S1). Only reactions generating single products of the correct size were used for further analyses.
Prokaryote expression, purification, and renaturation
The recombinant proteins were produced by Detai Biologics Co., Ltd. (Nanjing). The cDNA sequences encoding the full lengths of OlGdnfa (234 aa) and OlGdnfb (226 aa) depletion of each signal peptide (19 aa) were amplified by reverse transcription PCR (RT-PCR) with specific primers containing NdeI/HindIII restriction enzyme sites and then inserted into prokaryotic expression vector pET-30a digested with the same restriction enzymes (Supplementary Table S1). The constructed plasmids were confirmed by sequencing and then were used to transform competent Escherichia coli BL21 (DE3). The recombinant proteins were induced to express by isopropyl β-D-1-thiogalactopyranoside (IPTG) and then purified by Ni-NTA affinity chromatography. After renaturation by gradient dialysis and purification by ion exchange chromatography (SP Sepharose™ FF), the recombinant proteins were dissolved in Tris buffer (pH 8.0) and then detected by SDS-PAGE and western blotting using anti-His tag antibody, respectively. The purity of recombinant proteins was determined by analyzing the band intensity in SDS-PAGE using ImageJ software.
Cell culture
SG3 cells were incubated in ESM4 as previously described [24], which is the DMEM with 25 mM Hepes, 100 U/mL penicillin and streptomycin, 15% fetal bovine serum, 2 mM
CCK8 cell viability assay
The CCK8 assay was used to quantitatively assess cell survival. Briefly, SG3 cells (approximately 5,000 cells/well) were seeded in gelatin-coated 96-well plates with 5 N, 5 N containing different concentrations of OlGdnfa or OlGdnfb (1, 10, 50 ng/mL) as described in the Cell culture section, four replicates in each group. At the indicated time, CCK8 solution (Dojindo) was added to each well, and then SG3 cells were incubated for 4 h at 28°C. Absorbance of each sample at 450 nm was measured with a microplate reader (Thermo Labsystems), while absorbance at 630 nm was also measured as reference. The proliferative activity of each sample is arbitrarily indicated by the relative absorbance value. The relative absorbance value = (the absorbance value at 450 nm–the absorbance value at 630 nm) at indicated time/(the absorbance value at 450 nm–the absorbance value at 630 nm) at 0 h. Each experiment was repeated at least three times.
EdU labeling
To further evaluate the proliferative activity of SG3 cells, the cells in the different culture conditions were first incubated in 48-well plates for 24 h at the concentration of about 2 × 104 cells/well, then 10 μmol 5-ethynyl-2-deoxyuridine (EdU; RIBOBIO) was added to each well and incubated for 2 h at 28°C. EdU staining assay was carried out according to the manufacturer's instructions. Briefly, cells were washed with PBS, followed by fixation and permeabilization with PBS containing 2% formaldehyde, 0.5% Triton X-100, and 300 mM sucrose for 20 min. After extensive washing with PBS, cells were blocked with 10% FBS in PBS for 30 min and incubated with fluorescent azide probe Apollo®567 for 30 min. Cells were washed and counterstained with Hoechst 33342. The fluorescence signals were observed under a Nikon Ti–S inverted fluorescence microscope.
Immunofluorescence and AP staining
SG3 cells (approximately 1 × 106 cells/well) were incubated in gelatin-coated six-well plates for 14 days with ESM4, 5 N containing 50 ng/mL OlGdnfa or 50 ng/mL OlGdnfb, respectively. Then, the Pou5f3 expression of SG3 cells was detected by immunofluorescence staining as previously described [25]. Briefly, SG3 cells were fixed with 4% paraformaldehyde for 15 min and permeabilized with 0.1% Triton X-100 for 25 min. After blocking in 5% BSA/PBS for 2 h, the cells were incubated with primary antibody overnight at 4°C. After being washed several times with PBS, the cells were incubated with the PE-conjugated goat anti-rabbit IgG (Vector Laboratories), and the cell nuclei were stained in Hoechst dye solution (Molecular probes). The fluorescence signals were observed under a Nikon Ti–S inverted fluorescence microscope. Meanwhile, the alkaline phosphatase (AP) activity was detected as previously described [24].
Real-time PCR
The mRNA level was quantified by real-time PCR using delta–delta Ct (threshold crossing value) calculations. Primer pairs used in this study were gene specific and intron spanning (Supplementary Table S1). Before use in real-time PCR, each primer set was validated for use by gel analysis of RT-PCR products. Real-time PCR was performed on the StepOnePlus™ Real-Time PCR System (Life Technologies) as previously described [26].
Statistical analyses
Statistical analyses were performed with Student's t-test for comparison between two groups. P < 0.05 or P < 0.01 is considered as significant or very significant difference from the control, respectively.
Results
Complementary DNA sequences and analyses
The cDNAs of Olgdnfa and Olgdnfb were successfully obtained through RT-PCR and sequencing, which are 780 and 744 nt in length, respectively (Supplementary Fig. S1). The GenBank accession numbers of the two cDNAs are BankIt1928842 KX397593 and BankIt1928842 KX397594, respectively. The two cDNAs encode 253 and 245 amino acid residues (AA) for OlGdnfa and OlGdnfb, respectively (Supplementary Fig. S1). Each of the two putative proteins was predicted to contain a 19-aa signal peptide, a propeptide domain, and a TGF-β domain, together with the conserved functional residues characteristic of the TGF-β superfamily, including a cleavage motif of proteolysis (RXRR), 2 or 1 potential N-glycosylation sites, and seven conserved cysteine residues in the TGF-β domain (Supplementary Fig. S1).
Olgdnfa and Olgdnfb are separately located at chromosomes 12 and 9, and each has just two coding exons like the other vertebrate homologs (Fig. 1A). Surprisingly, the amino acid identity of OlGdnfa and OlGdnfb to each other is similar to the other known vertebrates, which is only 49% in the mature protein part and 72% in the TGF-β domain. Meanwhile, OlGdnfa/OlGdnfb have over 49/45% identity in the mature protein part and 69/65% in the TGF-β domain to the other vertebrates, respectively (Supplementary Table S2).
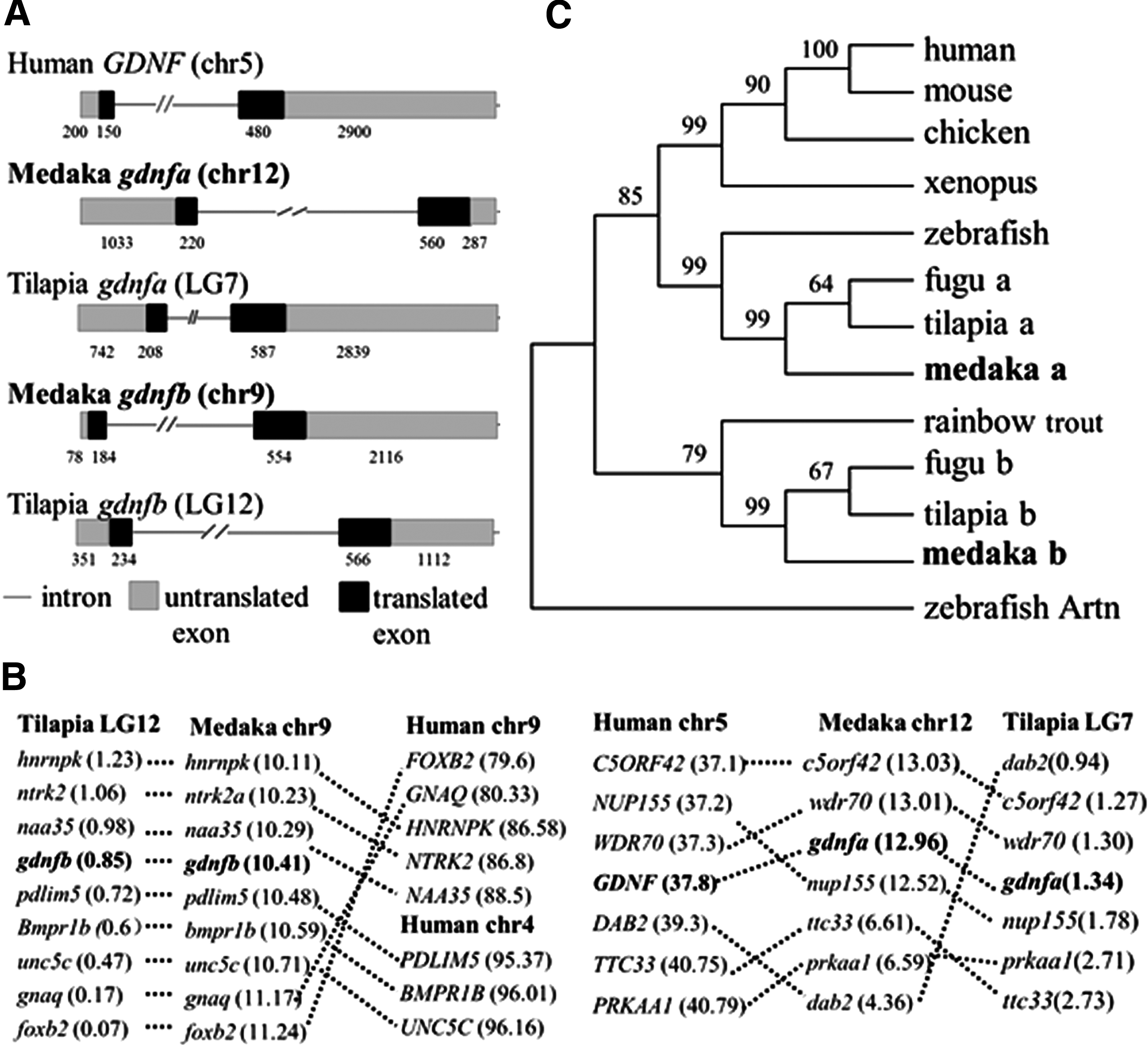
Genomic organization, chromosomal synteny, and phylogenetic tree.
A cross-species comparison of chromosomal location analysis indicates that the gdnfa-containing chromosome 12 in medaka is syntenic to tilapia gdnfa-containing LG7 and human GDNF-containing chromosome 5, while the gdnfb region in medaka chromosome 9 is syntenic to tilapia gdnfb-containing LG12 and human chromosomes 4 and 9. These results suggest that Olgdnfa has good synteny to mammalian GDNF (Fig. 1B). In the phylogenetic tree, OlGdnfa and OlGdnfb formed two distinct clades. OlGdnfa together with Gdnfa from fugu and tilapia was clustered with the tetrapod clade, while OlGdnfb was clustered with Gdnfb from fugu and tilapia and formed a separate clade using zebrafish Artn as an outgroup (Fig. 1C).
The mRNA expression
Relative high expression levels of Olgdnfa and Olgdnfb were observed in different stages of embryo development through morula to fry. The expression of Olgdnfa was readily detectable in various adult tissues, including testis; while relative high expression of Olgdnfb was observed in ovary, low expression level was observed in eye and testis (Fig. 2).
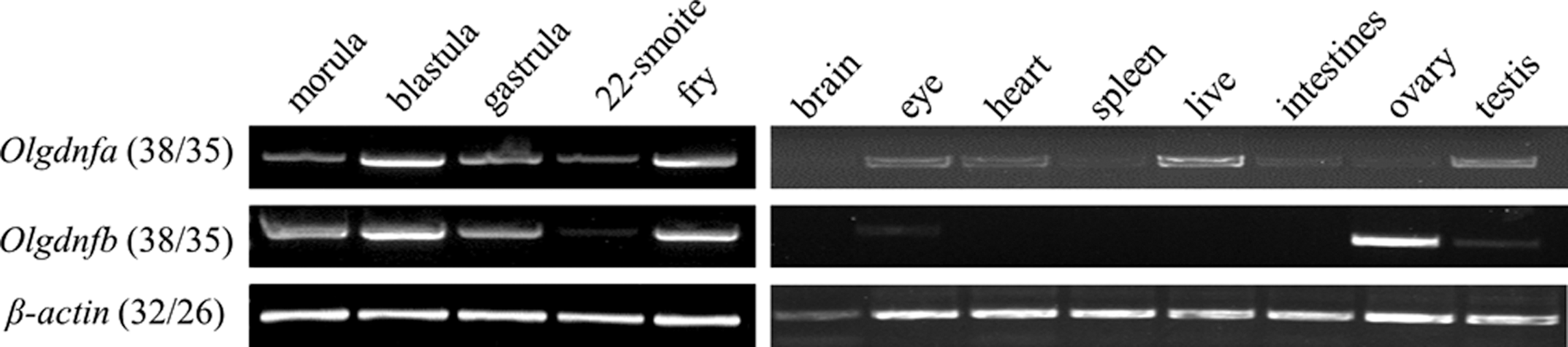
The mRNA expression of Olgdnfa and Olgdnfb in embryos during indicated development stages and adult tissues. β-Actin was used as the internal control. Numbers of PCR cycles are in parentheses for embryos and adult tissues, respectively.
Production of recombinant proteins
Through induced expression of the constructed pET30a, purification, and renaturation, the recombinant proteins, OlGdnfa and OlGdnfb, were obtained. SDS-PAGE and western blotting detection indicated that there was a strong band of the molecular weight compatible with the predicted size of OlGdnfa and OlGdnfb (approximately 27.4 and 27.2 kDa), respectively (Fig. 3). Meanwhile, a band with approximately 55 kDa was also observed for OlGdnfa and OlGdnfb, respectively, which was compatible with the predicted size of the dimer (Fig. 3). The purity of the two recombinant proteins is above 90%, evaluated by ImageJ software.
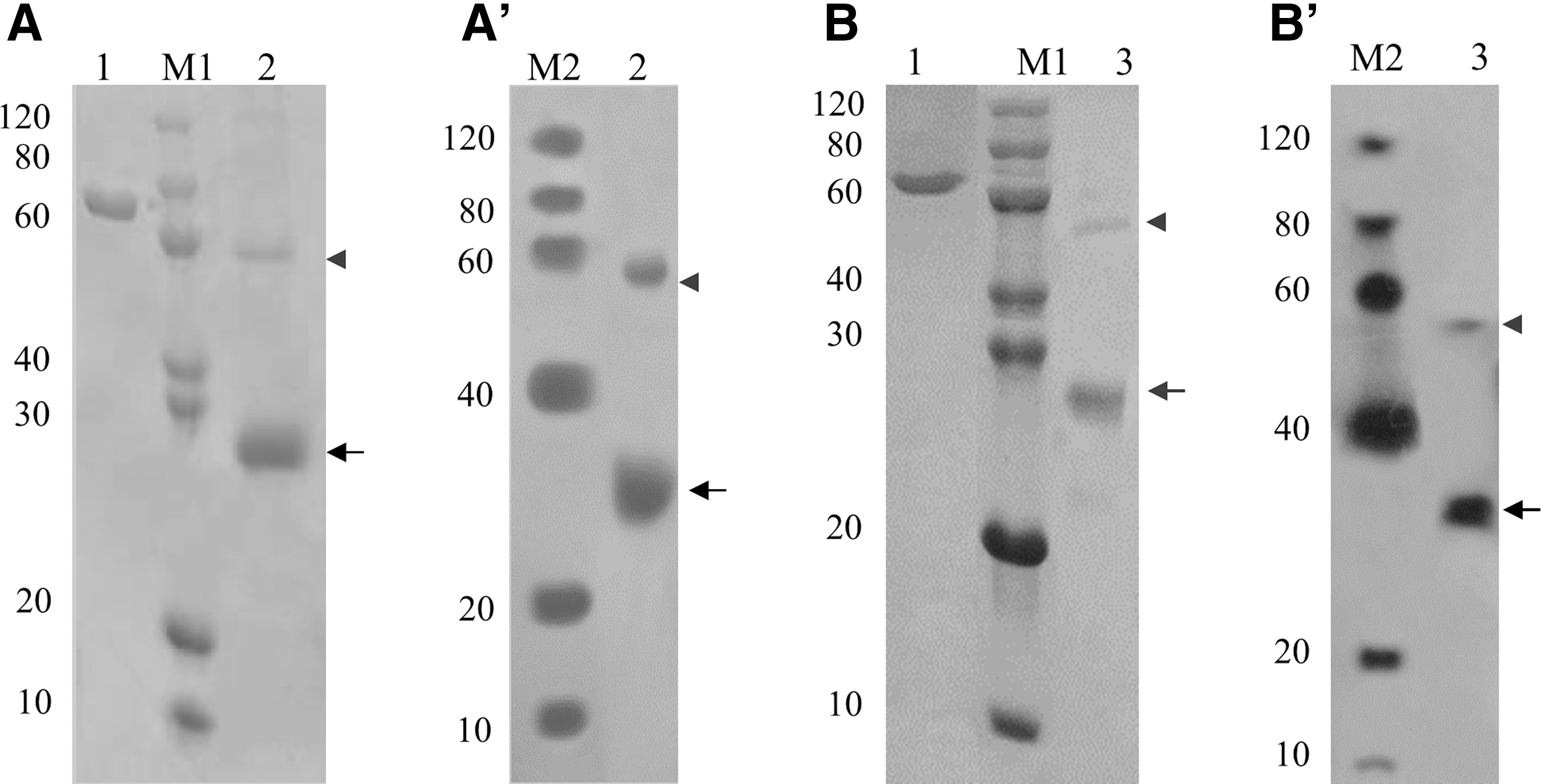
SDS-PAGE and western blotting analyses of the recombinant proteins, OlGdnfa
Proliferation activity of OlGdnfa and OlGdnfb
The proliferative activity of SG3 cells in 5 N containing the recombinant protein OlGdnfa or OlGdnfb was separately measured by EdU incorporation staining and CCK8 cell viability assay. Our data consistently indicate that both OlGdnfa and OlGdnfb can significantly promote the proliferation of SG3 cells (Fig. 4). In 5 N, with the recombinant protein OlGdnfa or OlGdnfb for 24 h, there were about 35% or 34% proliferative cells after a 2-h EdU incubation, while just 22% in the control 5 N (Fig. 4A, B). In a 96-well plate with approximately 5 × 103 cells per well, SG3 cells showed evidently proliferative activity in all of the sampled groups, including the control at 4 days. However, at 5 days onward, SG3 cells showed almost no proliferative activity in the control, while significant proliferative activity was observed in 5 N with OlGdnfa or OlGdnfb at the concentration of 1, 10, or 50 ng/mL. In addition, the proliferation of SG3 cells was more prominent at OlGdnfa with 1 ng/mL than 10 or 50 ng/mL (Fig. 4C).
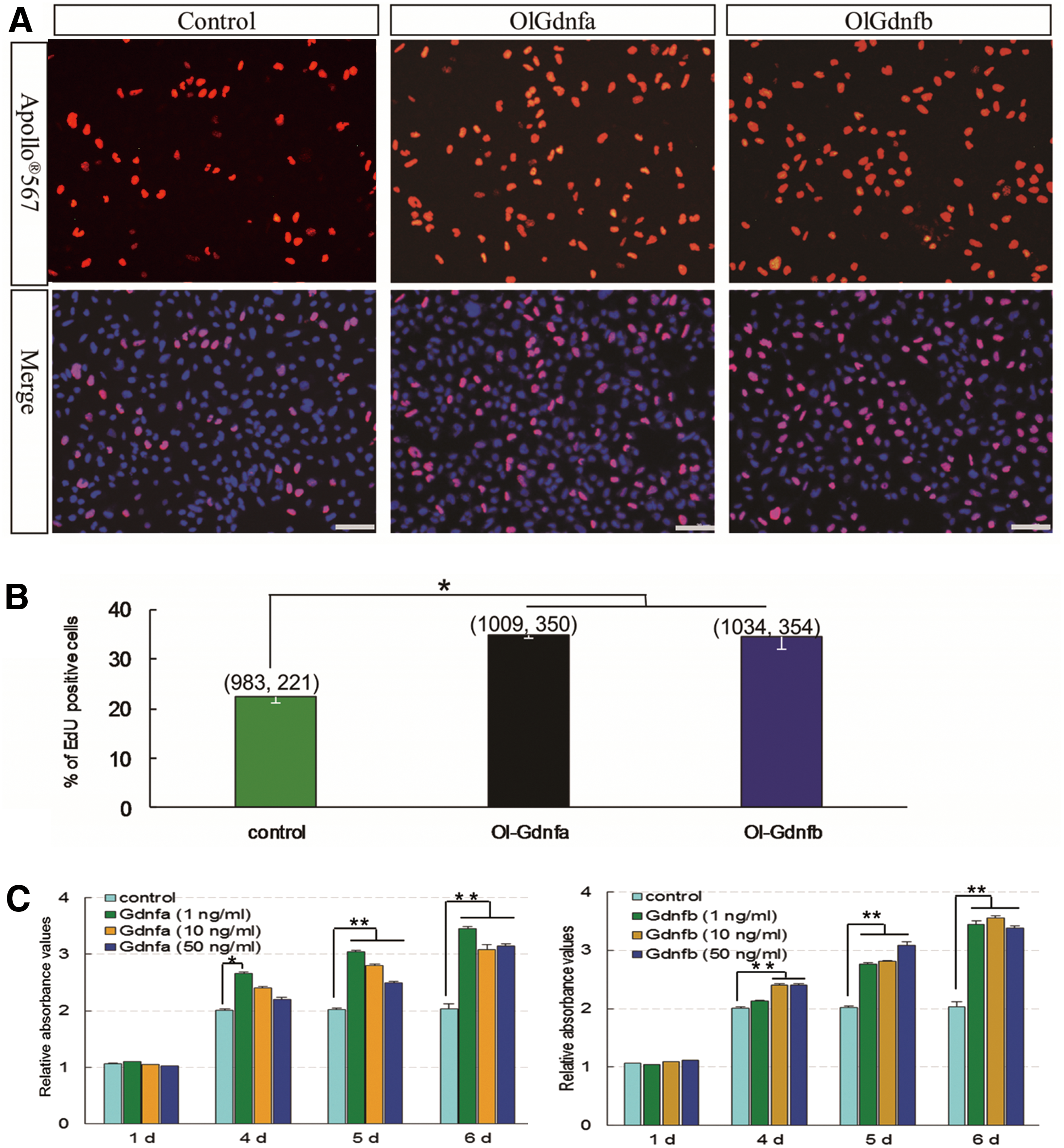
Proliferative activity of OlGdnfa and OlGdnfb.
The spermatogonial property of SG3 cells
To investigate whether OlGdnfa or OlGdnfb might maintain the spermatogonial property of SG3 after long-term incubation, SG3 cells were separately incubated in the complete medium ESM4, 5 N (ESM4 depletion of bFGF, medaka embryo extract, and fish serum) with the addition of 50 ng/mL OlGdnfa or OlGdnfb, and then analyzed after a 14-day incubation. The expression of spermatogonial genes, pou5f3 and nanos2, as well as germline-specific gene vasa was abundant in 5 N with the addition of OlGdnfa or OlGdnfb, which was similar to that in ESM4 (Fig. 5A). The Pou5f3 expression at the protein level was further confirmed by immunofluorescence staining (Fig. 5B). Moreover, the AP activity of SG3 in 5 N with OlGdnfa or OlGdnfb was as evident as in ESM4 (Fig. 5C). Taken together, these results suggest that SG3 cells could retain spermatogonial property in the 5 N with the addition of OlGdnfa or OlGdnfb during the 14-day incubation.
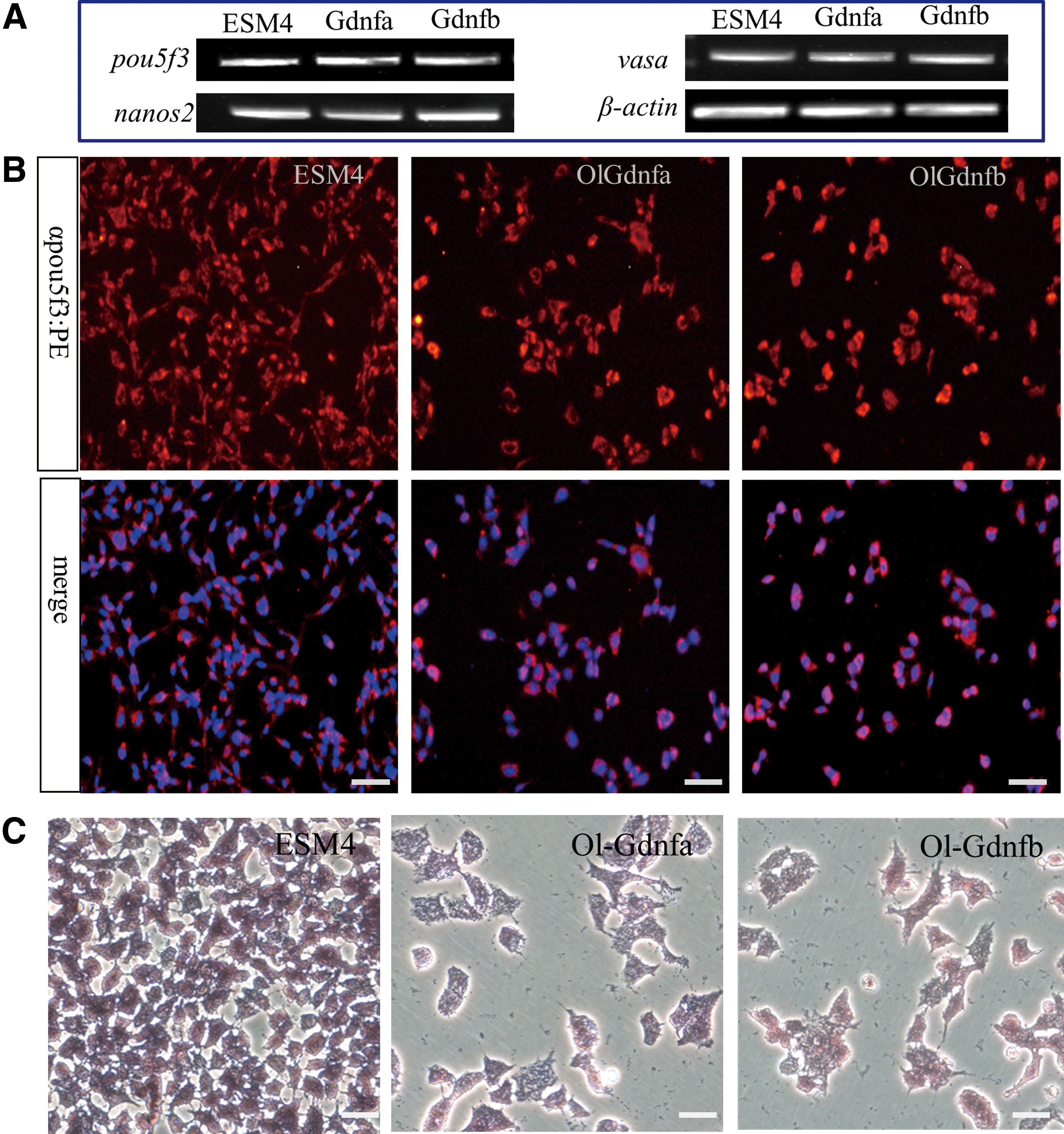
Spermatogonial property. After being separately incubated in ESM4, 5 N supplemented with 50 ng/mL OlGdnfa or OlGdnfb for 14 days, SG3 cells were harvested and further analyzed.
The regulated genes by OlGdnfa or OlGdnfb
In rodents as well as primates, BCL6B is the most responsive gene regulated by GDNF and essential for SSC self-renewal and survival [27]. Meanwhile, Etv5 is also dramatically upregulated by GDNF and important for SSC self-renewal in mice and rats [10]. In the present study, SG3 cells were incubated in 5 N for 18 h, and then supplemented with 50 ng/mL OlGdnfa or OlGdnfb. After a further 4-h incubation, the expression level of bcl6b was significantly upregulated, comparing with that in the control 5 N (P < 0.05), while etv5 displayed no significant change (P > 0.05) (Fig. 6).
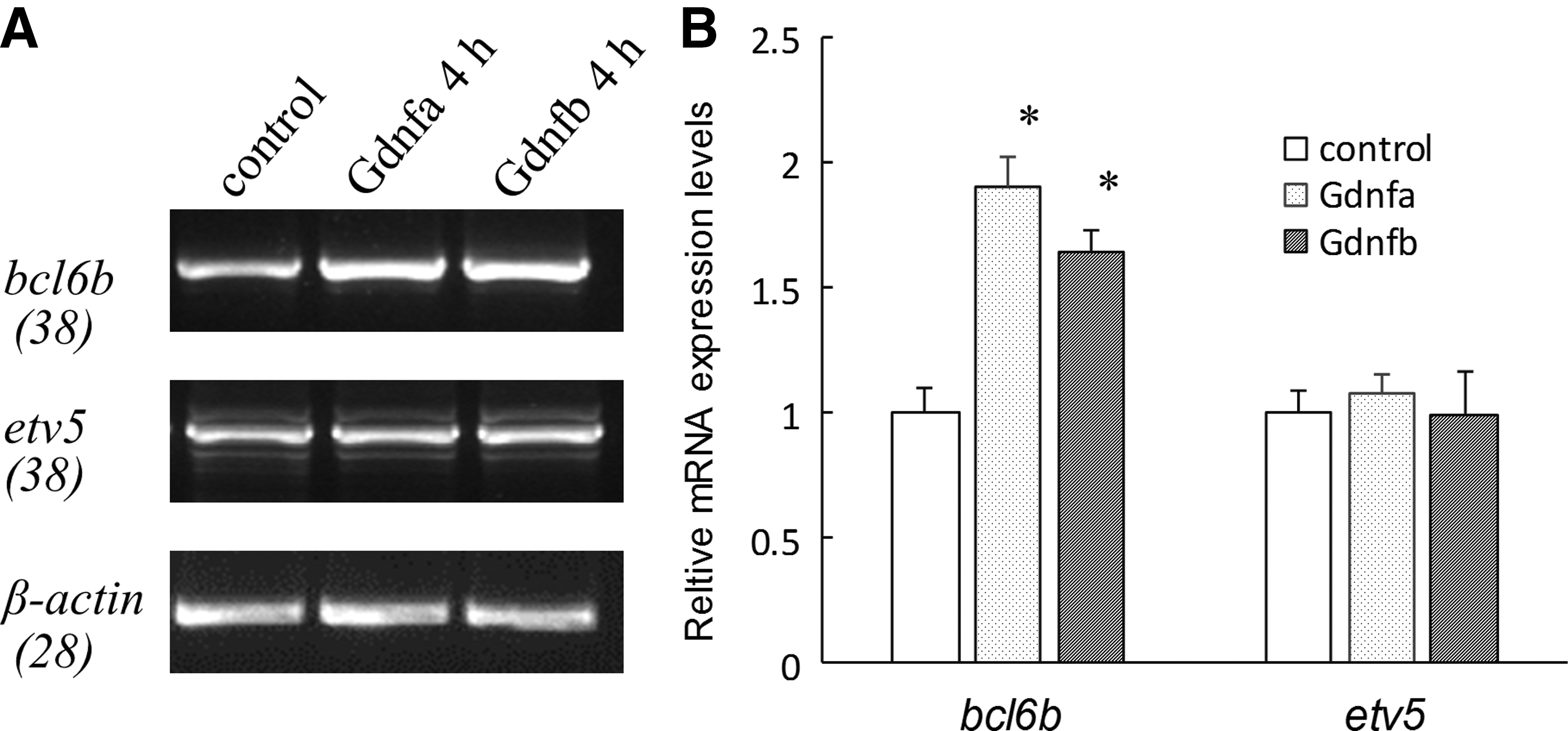
The mRNA expression levels of bcl6b and etv5 regulated by OlGdnfa or OlGdnfb. The SG3 cells were incubated in 5 N for 18 h, subsequently supplemented with 50 ng/mL OlGdnfa or OlGdnfb and incubated for further 4 h, finally harvested, and analyzed. The cells incubated in 5 N were used as the control.
Discussion
Comprehensive bioinformatic analyses suggest that the genes and cDNAs of Gdnf homologs widely exist in teleost fishes. Nonetheless, the study of gdnfs from teleost fishes is just limited to rainbow trout at the mRNA expression levels [17,18]. In the present study, two homologs of GDNF from medaka, namely OlGdnfa and OlGdnfb, were identified, both of which share 49% identity to each other. The mRNA expression of OlGdnfa and OlGdnfb occurred in different stages of embryo development and various tissues, including testis. Both of the recombinant proteins, OlGdnfa and OlGdnfb, can promote the proliferative activity of SG3 cells and retain the spermatogonial property by spermatogonial gene marker (pou5f3, nanos2, vasa, etc.) expression and AP activity. The SG3 cell line retains a diploid karyotype, the phenotype and gene expression pattern of SSCs, long-term self-renewing cell divisions, and differentiation potential (sperm production) after 140 passages during 2 years of culture. Thus, it is reasonable to speculate that OlGdnfa and OlGdnfb could mediate the self-renewal and maintenance of SSCs just like the mammalian ortholog.
Comprehensive analyses suggest that both OlGdnfa and OlGdnfb are homologs of mammalian GDNF by sequence and phylogenetic analyses. Like other members of the transforming growth factor-β (TGF-β) family, both OlGdnfa and OlGdnfb have a 19-aa signal peptide and an RXRR proteolytic processing site. The mature protein is released after removal of the signal peptide and activated by proteolytic cleavage at the conserved proteolytic RXXR site. Additionally, both OlGdnfa and OlGdnfb have seven conserved Cys residues located at the bioactive C-terminal domain, which is characteristic in the TGF-β superfamily (refer to Supplementary Fig. S2) [5]. On a phylogenetic tree, OlGdnfa is clustered with GDNF from tetrapod vertebrates, whereas the putative Gdnfb proteins from medaka and other fish species form a distinct clade with zebrafish Artn as an outgroup. This suggests that two Gdnfs might exist in fish lineage before tetrapod vertebrate lineage divergence occurred, which is something like the whole genome duplication event in the common ancestor of all extant teleosts [28]. Consistently, the amino acid sequence identity of OlGdnfa and OlGdnfb to each other is similar to the other known vertebrates (approximately 50% in the mature protein part and 70% in the TGF-β domain). Recently, it has been reported that there is no Gdnf, but a homologous sequence Artn, in dogfish, a chondrichthyan model, suggesting that teleost fish Gdnf might represent the ancestral prototype in vertebrates [21]. However, the functions of Gdnf in fish SSCs so far remain unclear.
In mammals, it is well established that the GDNF-GFRα1 signaling pathway mediates the self-renewal and maintenance of SSCs [2 –5]. Interestingly, recent studies indicate that the expression of Gfra1 occurs in the undifferentiated spermatogonia of Nile tilapia, dogfish, and rainbow trout [17,19,21]. In the present study, the expression of Gfrα1 in medaka was observed predominantly in the spermatogonia in adult testis as well as in vitro culture SG3 cells (our unpublished data), suggesting that the Gdnf/Gfrα1 signaling pathway might have a role in the fish SSC regulation. In the present study, to investigate the biological activity of fish Gdnf, bioactive recombinant proteins, OlGdnfa and OlGdnfb, were successfully obtained with over 90% purity and evaluated in an in vitro culture system.
Studying SSC in vitro culture provides a valuable model in identifying crucial growth factors for self-renewal and deciphering the mechanisms that regulate SSC development. So far, our knowledge about the underlying mechanisms of SSC self-renewal and maintenance has been largely attributed to the cell culture system in a limited number of mammalian species (particularly mouse and rat) [6,7,9,10,12]. In nonmammalian vertebrates, SG3 provides a valuable tool to investigate single growth factors crucial for self-renewal and molecular regulatory mechanisms in SSCs. In a feeder-free and conditional culture medium (ESM4), SG3 cells show continuous proliferation over 140 passages and sperm production in vitro [24]. It also needs to be mentioned that this culture condition, ESM4, has enabled the derivation of medaka diploid embryo stem (ES) cells, haploid ES cells, and the blastula-derived primordial germ cells capable of colonizing the chimeric germline [29
–32]. Hence, it will be interesting to determine what factors are crucial for the cultivation of medaka stem cells, especially germline stem cells. In the present study, a culture medium (5 N) was used to investigate the potential roles of medaka Gdnfs in SG3 cells. The 5 N is the DMEM supplemented with fetal bovine serum,
To date, because of lack of molecular markers highly specific for piscine SSCs, some highly conserved vertebrate markers such as Pou5f3 (also called Oct4, Pou5f1) and Nanos2 have been used as important candidates for evaluating SSCs in fish [19,33,34]. A recent study indicates that medaka Pou5f3 is not only essential for pluripotency in blastula formation and ES cell derivation but also strongly expressed in undifferentiated spermatogonia, barely detectable in primary and absent in secondary spermatocytes, spermatids, and sperm [35]. In trout, nanos2 expression is restricted to a subpopulation of undifferentiated spermatogonia, which are authentic SSCs verified by transplantation [34]. Additionally, a previous report indicates that in the medaka ES cells, spermatogonia have strong AP staining, while little in spermatids and sperm [24]. In the present study, the spermatogonial property of SG3 cells after incubation in different culture conditions for 14 days was measured through gene expression analyses and AP staining. The candidate spermatogonial genes such as pou5f3 and nanos2 and germline-specific gene vasa were highly expressed in all groups of SG3 cells. The protein expression of Pou5f3 was further confirmed by immunofluorescence staining. Meanwhile, SG3 cells of all groups displayed evident AP activity after a 14-day incubation. Taken together, these findings suggest that OlGdnfa or OlGdnfb could promote the proliferation of SG3 during a 14-day duration without compromising the spermatogonial property.
Notably, the expression level of bcl6b in SG3 cells was significantly upregulated after a 4-h treatment with the addition of OlGdnfa or OlGdnfb, while the expression of evt5 appeared to have no change. Both Bcl6b and Etv5 are the most responsive genes of SSCs regulated by GDNF treatment in mice, while Bcl6b is a conserved regulator of SSC self-renewal among mammalian species [7,27,36,37]. In agreement with the biological significance in SSCs, Bcl6b-null mouse testes display an impairment of spermatogenesis, in which many tubules contained degenerating stages of spermatogenesis or Sertoli cell-only phenotypes; the reduction of Bcl6b transcript levels dramatically affects the number and maintenance of mouse SSCs in vitro [36].
In conclusion, our results imply that GDNF from mammals to fish taxa might have not only conserved functional roles but also conserved regulatory molecules such as BCL6B in SSC self-renewal and maintenance. This study broadens and extends our understanding of GDNF functions in SSC biology, which would facilitate the SSC culture and the development of new biotechnologies for endangered or commercially valuable fish species.
Footnotes
Acknowledgments
This work was supported by a grant, cstc2016jcyjA1844, from the Natural Science Foundation Project of CQ CSTC; grant 31630082 from the National Natural Science Foundation of China; grants, XDJK2015A004 and XDJK2016E095, from Fundamental Research Funds for the Central Universities (Ministry of Education of China); and grant, 31201979, from the National Natural Science Foundation of China.
Author Disclosure Statement
There are no financial or competing interests in the subject matter or materials in the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.