
Abstract
Prolonged or high-dose exposure to anesthetics, such as propofol, can cause brain cell degeneration and subsequent long-term learning or memory deficits, particularly in the developing brain. However, the cellular and molecular mechanisms underlying the deleterious effects of propofol at certain stages of development remain unclear. In this study we found that propofol inhibited the proliferation, neuronal differentiation, and migration of neural stem cells (NSCs) while upregulating miR-141-3p. Silencing of miR-141-3p abrogated the effects of propofol on NSC neurogenesis. Propofol treatment downregulated IGF2BP2, a direct target of miR-141-3p, whereas overexpression of IGF2BP2 attenuated the effects of propofol and miR-141-3p on NSC neurogenesis. In short, propofol inhibits NSC neurogenesis through a mechanism involving the miR-141-3p/IGF2BP2 axis. Our results may provide a potential approach for preventing the neurodegenerative effects of propofol in the developing brain.
Introduction
G
Neural stem cells (NSCs) are a subset of undifferentiated precursor cells; they have the capacity to self-renew and can generate the three main cell types in the central nervous system, namely neurons, astrocytes, and oligodendrocytes [7,8]. NSCs have therefore gained attention for their therapeutic potential in neural repair. In addition, their self-renewal and differentiation properties have made NSCs a valuable experimental tool for the study of neurodegeneration in many diseases.
MicroRNAs (miRNAs) are noncoding RNAs of approximately 20–22 nucleotides that inhibit gene expression at the posttranscriptional level by binding to complementary sequences in the 3′-untranslated region (3′-UTR) of target genes [9,10]. miRNAs play pivotal roles in diverse developmental and physiological processes, including cell growth, differentiation, apoptosis, organ development, and the immune response [11]. In recent studies, the variable expression of miRNAs has been shown to play an important role in many different disease processes, including neurological diseases [12].
In the present study, we explored the mechanisms underlying the effects of propofol using a rat embryonic NSC model. Our results demonstrate that propofol inhibited the proliferation, migration, and neuronal differentiation of NSCs through a mechanism involving miR-141-3p and its direct target IGF2BP2. These findings suggest a potential mechanism for the deleterious effects of propofol on the developing brain.
Materials and Methods
Isolation and culture of NSCs and cell differentiation assays
All animal experiments were approved by the Institutional Animal Care and Use Committee of Shanghai Jiao Tong University. NSCs were harvested from the hippocampus of embryonic day 15 (E15) Sprague-Dawley rat embryos. Briefly, the hippocampus was collected, cut into smaller pieces, and dissociated mechanically into single cells. Cells were cultured in serum-free Dulbecco's modified Eagle's medium/F12 (DMEM/F12) supplemented with 20 ng/mL basic fibroblast growth factor (bFGF; R&D, Minneapolis, MN), 20 ng/mL epidermal growth factor (EGF; R&D), 2% B-27, and 1% N-2 supplement (Gibco, Carlsbad, CA). Neurospheres, formed after 4–7 days in culture, were dissociated into single cells using Accutase and plated on poly-
To verify the identity of NSCs, single neural cells were immunostained with anti-nestin antibody (Abcam, Cambridge, MA). To induce differentiation, the medium was changed to neuron differentiation medium (Neurobasal medium, 2% B-27 Serum-free supplement and 2 mM GlutaMAX; Gibco) or astrocyte differentiation medium [DMEM, 1% N-2 supplement, 1% fetal bovine serum (FBS), and 2 mM GlutaMAX; Gibco]. Cells were allowed to differentiate for 7 days, followed by immunostaining for neuronal and glial markers: anti-β-tubulin III (Proteintech Group, Wuhan, China) or anti-GFAP (Sigma-Aldrich, St Louis, MO). Cells were analyzed under a laser-scanning confocal microscope (Olympus, Tokyo, Japan) and counted in 6–10 random fields.
Propofol exposure
Propofol (Sigma-Aldrich) was diluted in dimethyl sulfoxide (DMSO; Sigma-Aldrich) for in vitro assays. The dose of propofol used clinically in humans ranges from about 4 to 20 μg/mL with higher doses used for induction of anesthesia and lower doses used for maintenance [13]. Thus, NSCs were exposed to the clinically relevant concentrations of propofol (0, 5, 10, or 20 μg/mL) or to equal volumes of DMSO as the vehicle control for 6 or 24 h. The mechanistic experiments in this study were performed in NSCs following a single exposure to 20 μg/mL propofol for 6 h.
Immunostaining
Cells were fixed for 20 min in 4% paraformaldehyde in phosphate-buffered saline (PBS) at room temperature. After extensive washing, cells were treated with 0.2% Triton X-100 in PBS for 8 min at room temperature followed by PBS containing 10% FBS (Gibco) for 1 h. Fixed cells were then exposed to primary antibodies at 4°C overnight and then incubated with fluorescently labeled secondary antibodies. Nuclei were counterstained with DAPI.
Lentivirus infection
NSCs were infected with miR-control, miR-141-3p, anti-miR-control, or anti-miR-141-3p lentivirus (Hanbio, Shanghai, China). At 24 h after infection, virus-containing medium was removed and replaced with normal maintenance medium. Cells were used for experiments at 3 days post-transduction.
Plasmid transfection
The IGF2BP2-overexpression plasmid pcDNA3-IGF2BP2 OE-3′-UTR was generated by inserting the IGF2BP2 complementary DNA (cDNA) (with 3′-UTR) into the pcDNA3 vector (Invitrogen, Carlsbad, CA). To construct a luciferase reporter vector, the wild-type (WT) 3′-UTR of IGF2BP2, containing the putative binding sites for miR-141-3p, was amplified by polymerase chain reaction (PCR) and cloned into the pGL3-luciferase reporter plasmid (Promega, Madison, WI). The corresponding mutant (MUT) constructs were generated by mutating the seed sequences of the miR-141-3p binding sites. The constructs were verified by sequencing.
Real-time PCR
Total RNA was extracted using TRIzol reagent (Invitrogen). The reverse transcriptase-PCR and PCR primers for miR-141-3p and U6 were purchased from RiboBio Co., Ltd. (Guangzhou, China). The cDNA synthesis was performed using the PrimeScript RT Reagent Kit (TaKaRa, Dalian, China) according to the manufacturer's instructions. Real-time PCR was performed using SYBR Green PCR Master Mix Kit (TaKaRa) on an ABI 7900 system (Applied Biosystems, Foster, CA). The expression of U6 or β-actin was used as the internal control.
Cell proliferation
Cells were plated in 96-well plates at a density of 2 × 103 per well for 2 days and treated with the specified concentration of propofol for the indicated times. The culture medium was removed, and 50 μL of a 0.1 mg/mL solution of 3-[4, 5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) was added to each well. The mixture was incubated at 37°C for 4 h, and the precipitate formed was dissolved in DMSO at room temperature for 30 min. The absorbance was read at 490 nm using a microplate reader.
Wound healing assay
Cells were seeded into six-well plates and grown to confluence. A wound was created by scraping the cell monolayer with a pipette tip, and cells were allowed to migrate from either side of the gap. Wound closure was monitored in the same field at different times. The extent of healing was determined using the ImageJ program as an average closed area of the wound relative to the initial wound area at 24 h after wounding.
Western blotting
Cells were harvested and lysed in RIPA buffer (Beyotime, Haimen, China). Protein samples were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto PVDF (polyvinylidene difluoride) membranes (Millipore, Billerica, MA). Membranes were blocked with 5% nonfat milk in Tris-buffered saline containing 0.1% Tween-20 (TBST), washed thrice in TBST, and incubated overnight at 4°C with primary antibodies against β-tubulin III, IGF2BP2, or β-actin (Proteintech Group). After incubation with the corresponding peroxidase-conjugated secondary antibodies, immunoreactive bands were detected by enhanced chemiluminescence (Millipore). The protein bands were quantitatively analyzed with NIH ImageJ software.
Luciferase reporter assay
HEK293T cells were seeded 24 h before transfection in 24-well plates at 50% confluence. The WT or MUT IGF2BP2 3′-UTR reporter constructs were cotransfected along with anti-miR-141-3p or scrambled control using Lipofectamine 2000 (Invitrogen). After 48 h, luciferase activity was assessed using the Dual-Luciferase Reporter reagent (Promega) following the manufacturer's instructions. Renilla luciferase was used for normalization.
Statistical analysis
All of the experiments were conducted at least in triplicate, and data are expressed as the mean ± standard deviation (SD). Significant differences between two groups were determined using two-tailed Student's t-test. Significant differences among more than two groups were assessed using one-way analysis of variance. Values of P < 0.05 are considered statistically significant.
Results
Characterization of rat embryonic NSCs
NSCs isolated from rat embryonic hippocampus were cultured, and they formed neurospheres after 4 days (Fig. 1A), and more than 90% of the cells were positive for nestin (a marker for progenitor cells) (Fig. 1B). Culture in specific induction media resulted in the differentiation of NSCs into neurons and astrocytes, as determined by positive immunostaining against the neuronal marker β-tubulin III and the astrocyte marker GFAP (Fig. 1C, D). These results confirmed the identity of the isolated cells as NSCs.
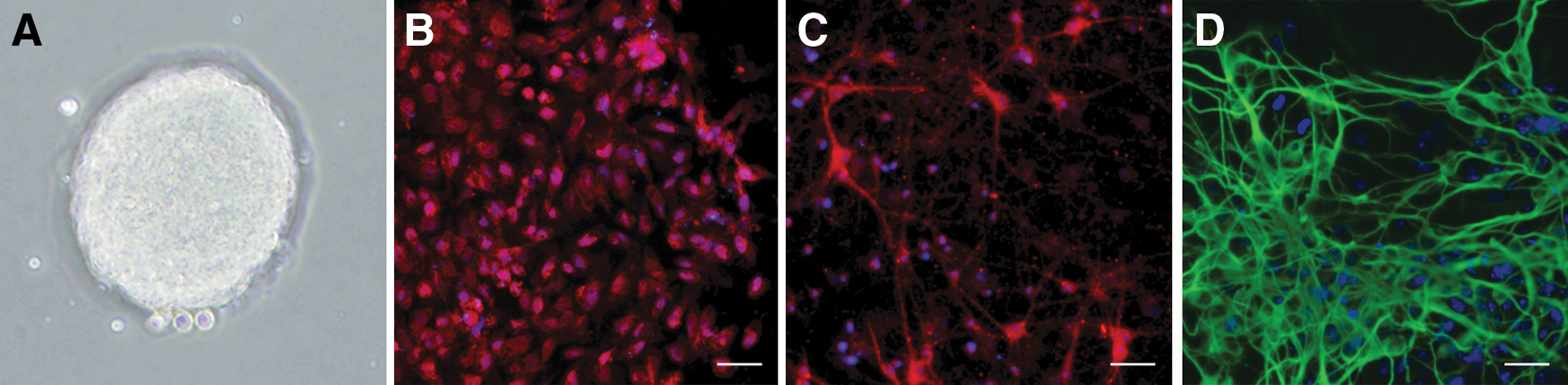
Isolation and identification of rat embryonic NSCs.
Propofol inhibits neurogenesis in rat NSCs
NSCs were exposed to various concentrations of propofol, and cell proliferation was assessed using the MTT assay at 6 and 24 h after treatment. The results showed that propofol inhibited NSC proliferation in a dose-dependent manner, with significant inhibitory effects at high doses and prolonged exposure times (20 μg/mL for 6 h, 10 and 20 μg/mL for 24 h) (Fig. 2A). In addition, cell culture and animal studies have shown that propofol can induce toxicity at high doses [2,14]. So, we choose the high dose (20 μg/mL) as the final concentration in the next study. To investigate whether propofol exposure effected the neuronal differentiation of NSCs, we examined the cellular phenotype using the neuronal marker β-tubulin III after propofol exposure (20 μg/mL for 6 h). As shown in Fig. 2B, exposure of NSCs to propofol decreased the percentages of neurons, indicating that propofol inhibited neuronal differentiation. This was further confirmed by western blot analysis of β-tubulin III expression (Fig. 2C).
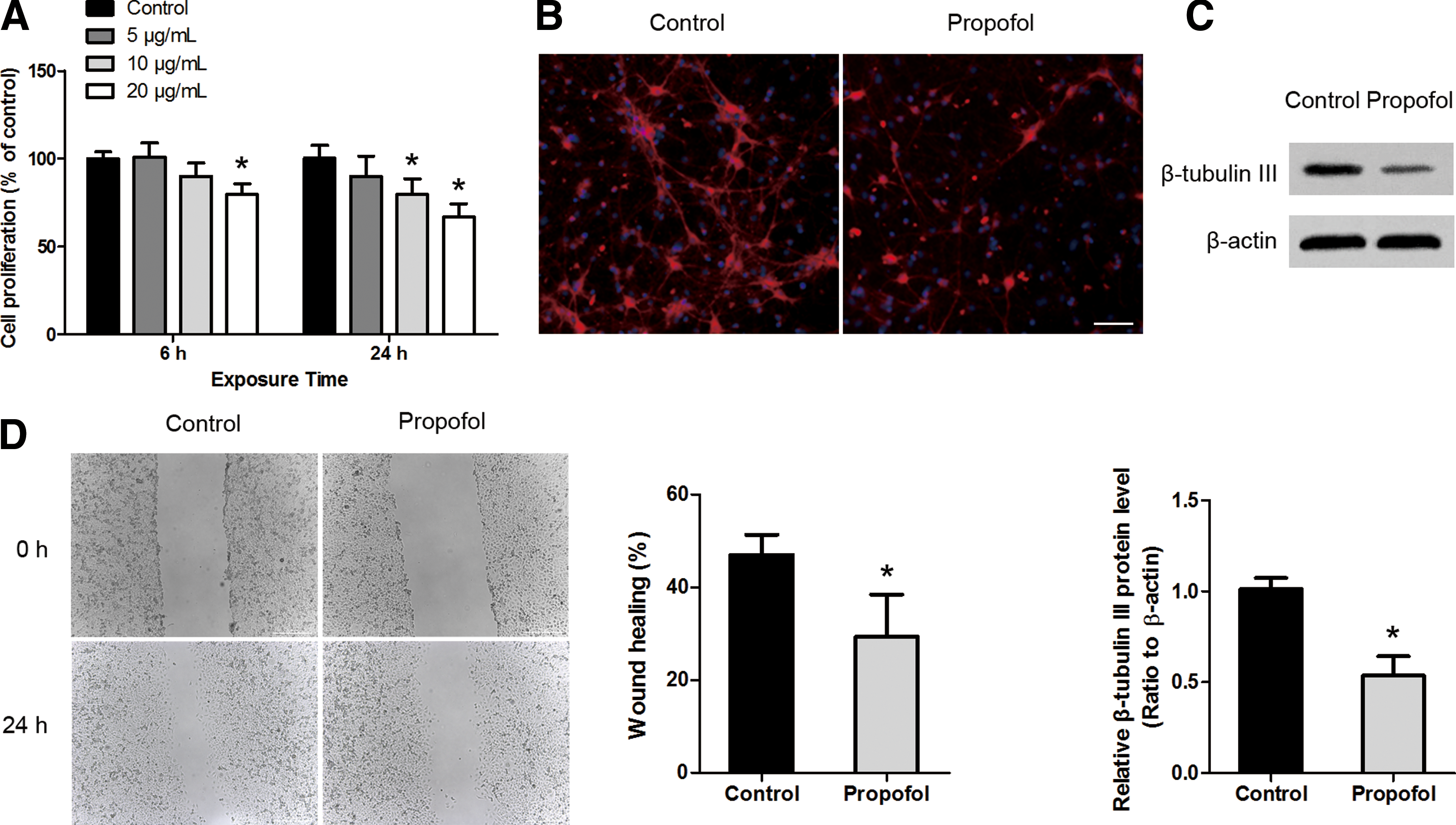
Propofol inhibits the proliferation, neuronal differentiation, and migration of rat NSCs.
In addition to NSC proliferation and differentiation, NSC migration is also a critical aspect of neurogenesis. Using a wound-healing assay, we demonstrated that there was decreased migration of NSCs in the presence of propofol (Fig. 2D). Taken together, these results indicate that propofol inhibits NSC neurogenesis.
Propofol inhibits neurogenesis of rat NSCs by upregulating miR-141-3p
Next, we treated NSCs with 20 μg/mL propofol for 6 h and then detected that miR-141-3p was upregulated by approximately fivefold (Fig. 3A). To explore the role of miR-141-3p in propofol-mediated inhibition of NSC proliferation, differentiation, and migration, NSCs were transduced with a lentivirus expressing anti-miR-141-3p, which effectively downregulates miR-141-3p expression (Fig. 3B). Cell viability was assessed in anti-miR-141-3p-transfected NSCs. We found that knockdown of miR-141-3p abolished the effect of propofol on cell viability (Fig. 3C). Western blot analysis to detect the neuronal marker in NSCs transfected with anti-miR-141-3p indicated that β-tubulin III was upregulated in the anti-miR-141-3p-transfected group compared with the anti-miR-control group in the presence of propofol (Fig. 3D). We further performed the wound-healing experiment and determined that knockdown of miR-141-3p abrogated the influence of propofol on NSC migration (Fig. 3E). These results indicate that propofol influences NSC neurogenesis by upregulating the level of miR-141-3p.
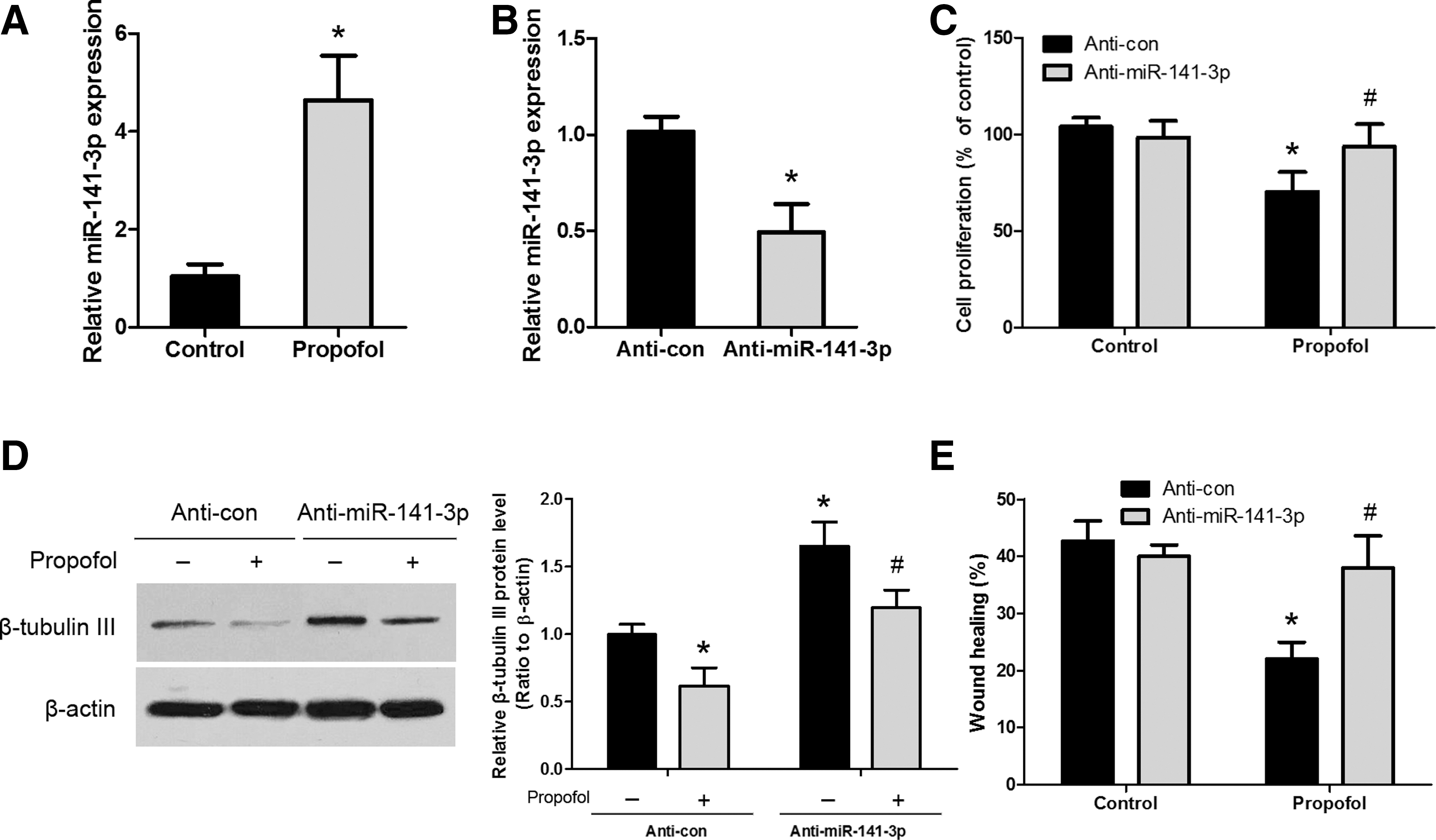
Involvement of miR-141-3p in propofol-mediated NSC proliferation, differentiation, and migration.
IGF2BP2 is the direct target of miR-141-3p
A search of the TargetScan, miRanda, and miRBase databases identified IGF2BP2 as a potential target of miR-141-3p, as shown by the conserved presence of the miR-141-3p binding sequence in the 3′-UTR of IGF2BP2 in different species (Fig. 4A). To confirm the binding of miR-141-3p to the 3′-UTR of IGF2BP2, a luciferase reporter assay was performed, and it showed that knockdown of miR-141-3p upregulated the luciferase activity of the WT, but not that of the MUT IGF2BP2 3′-UTR (Fig. 4B). Furthermore, miR-141-3p silencing was able to upregulate the expression of IGF2BP2 at both the messenger RNA (mRNA) and protein levels (Fig. 4C, D). However, whether IGF2BP2 expression is regulated by propofol remained unknown. We thus examined the effect of propofol on the expression of IGF2BP2. As shown in Fig. 4E and F, following propofol exposure, IGF2BP2 mRNA and protein levels were significantly downregulated. Taken together, these results indicate that propofol, through upregulation of miR-141-3p, downregulates IGF2BP2 expression in NSCs.
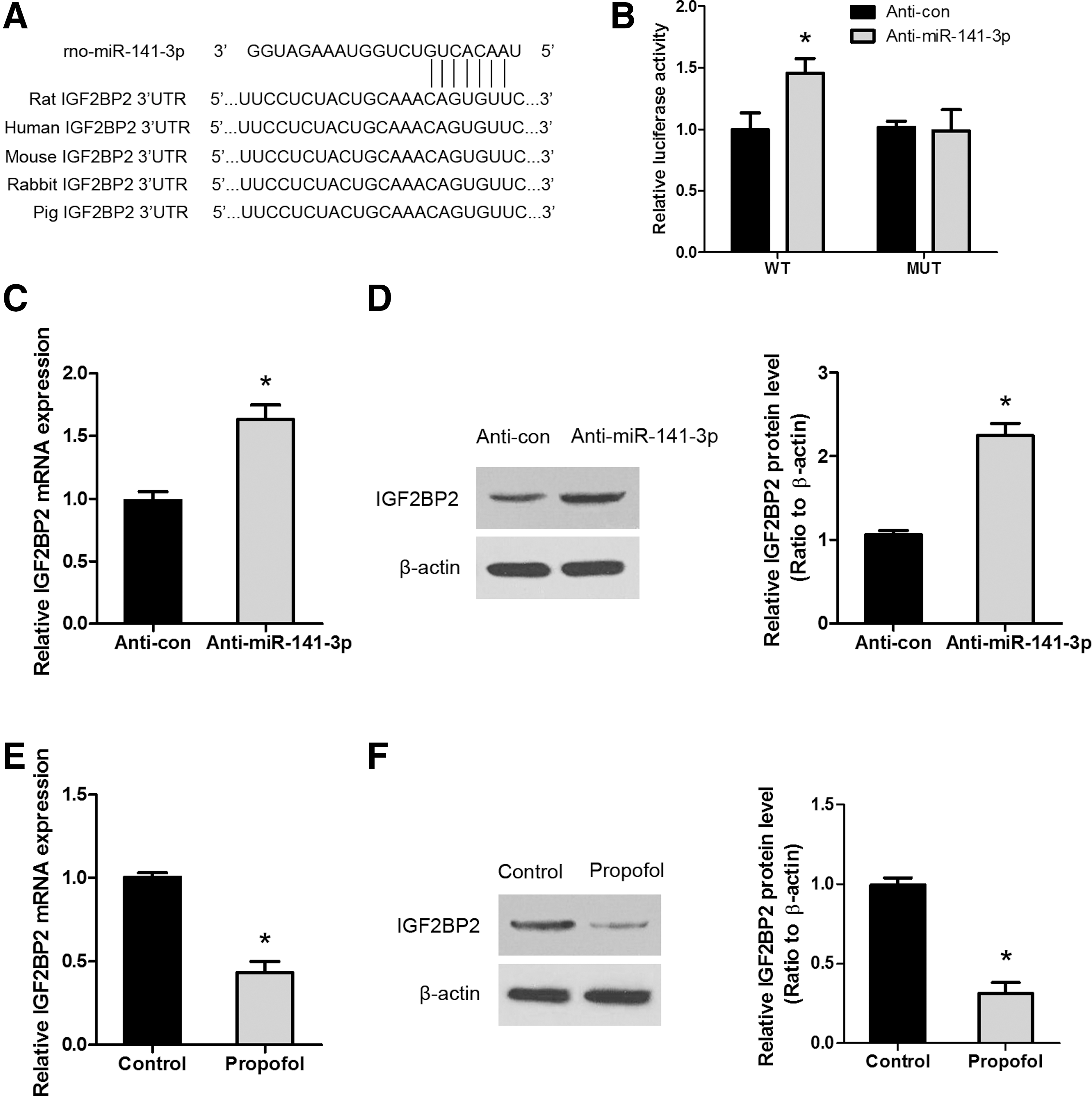
Propofol modulates the expression of miR-141-3p and its direct target, IGF2BP2.
Propofol inhibits neurogenesis of NSCs through miR-141-3p/IGF2BP2 signaling
To determine whether propofol influences NSC neurogenesis by regulating IGF2BP2 expression, we performed a rescue experiment. An expression construct encoding the IGF2BP2 coding sequence (with 3′-UTR) was generated, and NSCs were transfected with either vector control or IGF2BP2 overexpressing construct. Western blot analysis confirmed that IGF2BP2 overexpression vector can promote the expression of IGF2BP2 in NSCs (Supplementary Fig. S1; Supplementary Data are available online at
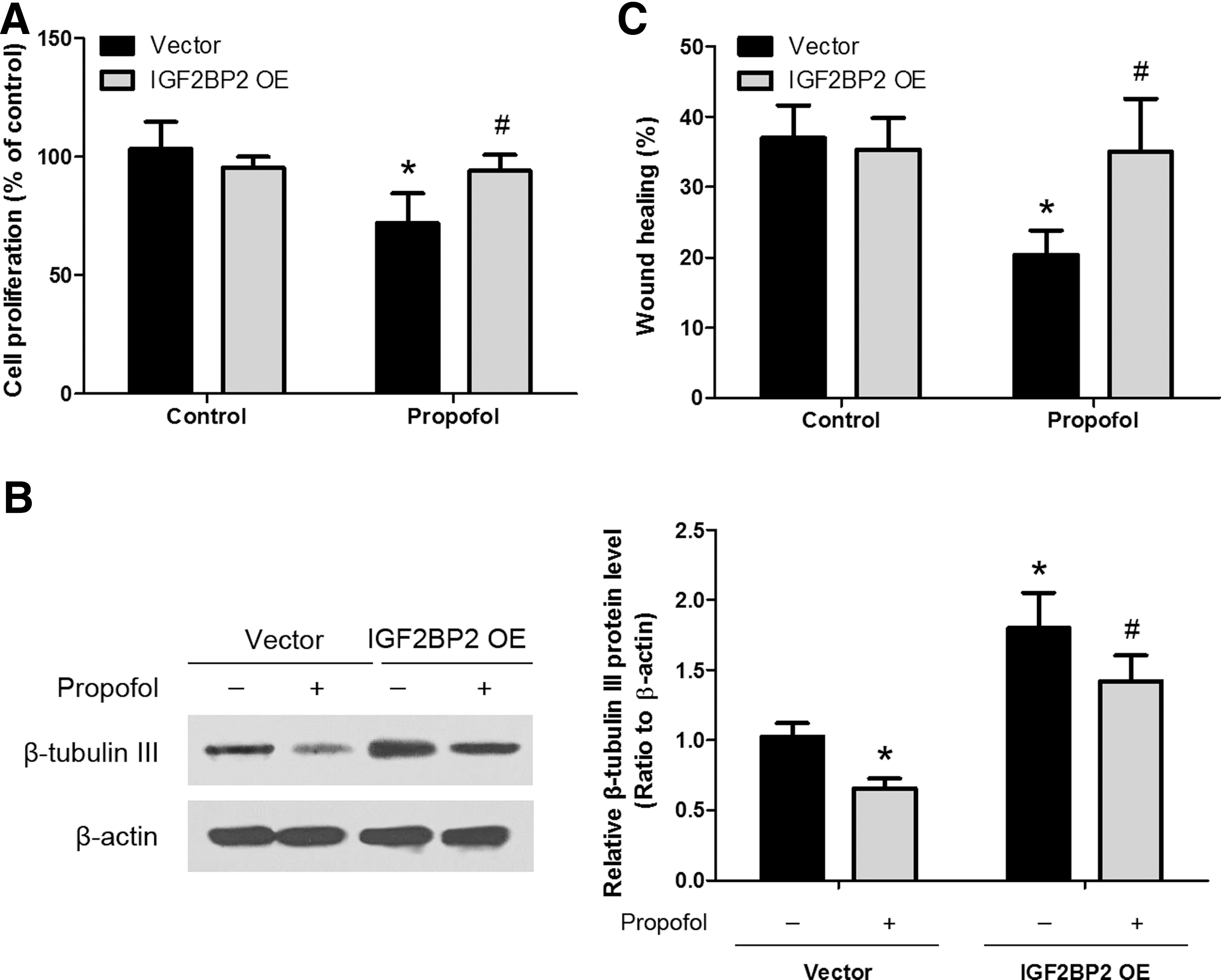
Involvement of IGF2BP2 in propofol-mediated NSC proliferation, differentiation, and migration. Transfection of NSCs with IGF2BP2 overexpression vector (IGF2BP2 OE) abrogated the effects of propofol on NSC proliferation
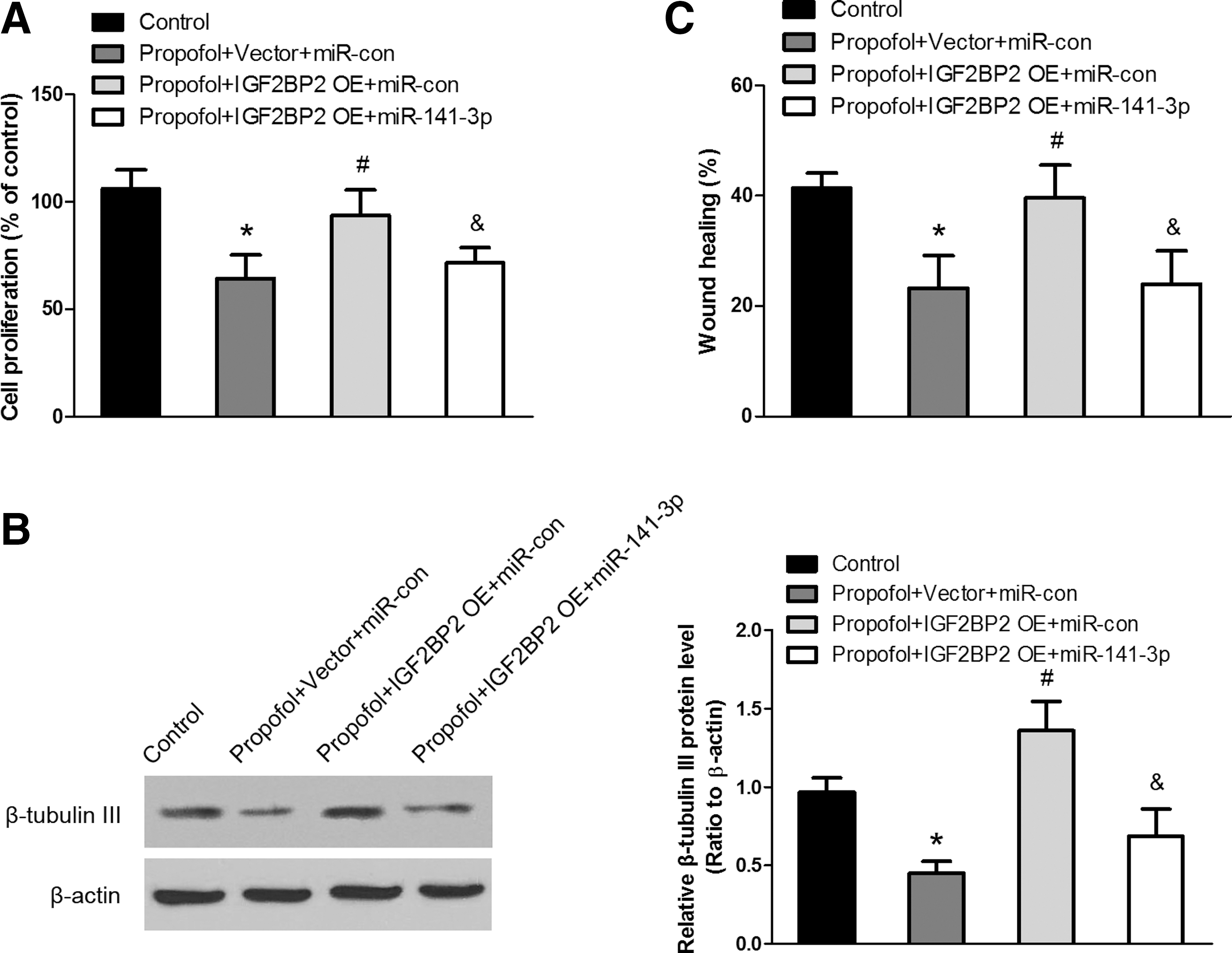
Propofol inhibits neurogenesis of NSCs through miR-141-3p/IGF2BP2 signaling. Overexpression of IGF2BP2 restored the propofol- or miR-141-3p-induced inhibition of proliferation
Discussion
New dentate granule cells are continuously generated from NSCs and integrated into the existing hippocampal circuitry in the brain through an orchestrated process termed neurogenesis [15]. For normal neurogenesis, a complex network of interacting neurons must be established within the brain. This process requires regulated NSC proliferation, differentiation, and migration, which are affected by a variety of pathological, as well as physiological, stimuli [16,17]. Although propofol is known to impair neurogenesis [18 –20], the mechanisms underlying regulation by specific miRNAs and their targets remain incompletely understood. In the present study, we elucidated the involvement of the miR-141-3p/IGF2BP2 axis in the deleterious effects of propofol on NSC proliferation, differentiation, and migration. These findings suggest that the miR-141-3p/IGF2BP2 axis may be a desirable target for research into possible neuroprotective strategies.
Many miRNAs are expressed at high levels in the central nervous system, where they contribute to maintaining normal neuronal functions and homeostasis, affecting neuronal differentiation, synaptic plasticity, and neurogenesis [21,22]. Among brain-specific miRNAs, miR-9 is involved in early neurogenesis and proliferation, while miR-124 plays a role in neuronal differentiation by targeting polypyrimidine tract-binding protein 1, promoting nervous system development and the differentiation of progenitor cells to mature neurons [23,24]. miR-134 has been shown to regulate dendrite formation and the development of the spinal cord [25]. miR-381 promotes NSC proliferation and differentiation into a neuronal lineage by modulating the expression of its target HES1 [26]. miR-21 is involved in propofol-induced cell death in human embryonic stem cell-derived neurons through the signal transducer and activator of transcription 3 (STAT3)/Sprouty2/Akt pathway [13]. In this study, we show that propofol upregulates miR-141-3p and that knockdown of miR-141-3p abrogates the effects of propofol on NSC proliferation, differentiation, and migration. These findings suggest that miR-141-3p may be involved in the deleterious effects of propofol in the developing brain. Our findings are consistent with previous reports demonstrating a similar effect of miR-141-3p in human mesenchymal stem cells [27,28].
Another novel finding of this study was that IGF2BP2, a newly identified molecule, is a target of miR-141-3p. IGF2BP2 belongs to a conserved family of RNA-binding proteins with important roles in cell polarization, migration, proliferation, and differentiation [29]. IGF2BPs 1–3 are mainly expressed during early development, from the zygote until embryonic stages, and are necessary for nerve cell migration and morphological development by controlling cytoskeletal remodeling and dynamics [30,31]. IGF2BP2 expression is upregulated in early-stage neural precursor cells (NPCs), and overexpression of IGF2BP2 induces neurogenesis and inhibits astrocytic differentiation in late-stage NPCs, suggesting its role as a regulator of NPC differentiation [32]. In this study, a bioinformatics analysis using TargetScan, miRanda, and miRBase suggested that miR-141-3p could bind to the 3′-UTR of IGF2BP2. Luciferase reporter assays confirmed IGF2BP2 as a direct target of miR-141-3p. IGF2BP2 overexpression attenuated the effects of propofol or miR-141-3p overexpression on NSC proliferation, differentiation, and migration. These results indicate that the effects of propofol are mediated by the miR-141-3p/IGF2BP2 axis. Future work in human NSCs experimental studies should be conducted to further elucidate this underlying mechanism.
In conclusion, the present study demonstrates that propofol inhibits NSC proliferation, neuronal differentiation, and migration by regulating miR-141-3p/IGF2BP2 signaling. Development of therapeutic strategies regulating the miR-141-3p/IGF2BP2 axis could be considered as a potential approach for preventing the neurodegenerative effects of propofol.
Footnotes
Acknowledgments
The work was supported by Science and Technology Commission of Shanghai Municipality (grant no. 14XD1402900) and the National Science Funds of China (Beijing, China; grant no. 81671058).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.