
Abstract
The use of allogeneic bone marrow-derived mesenchymal stem cells (BMDMSCs) may provide an effective alternative to autologous BMDMSCs for treatment of equine musculoskeletal injuries. However, concerns have been raised regarding the potential safety and effectiveness of allogeneic BMDMSCs. We conducted studies to assess the immunological properties of equine allogeneic BMDMSCs compared with those of autologous BMDMSCs. For assessment of inherent immunogenicity, the relative ability of allogeneic and autologous BMDMSCs to stimulate spontaneous proliferation of equine lymphocytes was compared. The immunosuppressive activity of the two cell types was evaluated by adding autologous or allogeneic BMDMSCs to activated lymphocytes and assessing suppression of lymphocyte proliferation and IFNγ production. Fifty-six allogeneic and 12 autologous combinations were evaluated. Studies were also done to elucidate mechanisms by which equine mesenchymal stem cells (MSCs) suppress lymphocyte function. Potential mechanisms evaluated included production of prostaglandin E2 (PGE2), nitric oxide, transforming growth factor-beta, and indoleamine 2,3-dioxygenase. We found that autologous and allogeneic BMDMSCs both induced mild but equivalent levels of spontaneous lymphocyte activation in vitro. In in vitro assays assessing the ability of BMDMSCs to suppress activated lymphocytes, both allogeneic and autologous BMDMSCs suppressed T cell proliferation and IFNγ production to an equal degree. The primary mechanism of equine BMDMSC suppression of T cells was mediated by PGE2. We concluded that allogeneic and autologous BMDMSCs are equivalent in terms of their immunomodulatory properties, and stimulated peripheral blood mononuclear cells appear to trigger the immunosuppressive properties of MSCs. Therefore, both cell types appear to have equal potency in modulating inflammatory processes related to acute or chronic musculoskeletal injuries in the horse.
Introduction
M
There are many sources of equine mesenchymal stem cells (MSCs), but BMDMSCs are by far the most commonly used as a clinical treatment in the horse [6,7]. BMDMSCs can be easily collected from either the ilium or the sternum of horses, using minimal restraint or sedation. A recent publication by Lombana et al. was unable to identify a difference in cell characteristics between BMDMSCs from ilial and sternal origin [8]. BMDMSCs for clinical use are typically culture expanded over a period of 2–4 weeks, at which time 10 to 20 × 106 BMDMSCs are typically administered to diseased tissues (eg, joints or injured tendons or ligaments) [6].
When autologous BMDMSCs are administered intra-articularly, the risk of immune reaction is minimal, but not negligible (∼9% rate of reactions) [7]. Likewise, a study utilizing intra-articular allogeneic MSCs reported a low occurrence of joint inflammation subsequent to injection (1.8%) [9]. Culture expansion of autologous BMDMSCs for clinical use has multiple clinical disadvantages, including the delay in obtaining cells, the expense of the procedure, and the effects of donor age on MSC functionality [10,11]. Expansion techniques have improved in efficacy over the last several years, and studies have suggested that only small volumes of bone marrow aspirate are necessary [12]. However, the delay from aspiration to administration of the treatment remains a shortcoming of autologous culture-expanded BMDMSCs [11 –13]. In addition, autologous bone marrow expansion is costly due to high labor and material costs.
A readily available source of allogeneic BMDMSCs would allow clinicians to choose the optimal time of injection following injury without the restriction of individual culture and expansion. In addition, a cryostored supply of BMDMSCs could reduce material costs and labor associated with cell expansion. Finally, there is speculation in the regenerative medicine field that MSCs from aged or injured patients may have reduced regenerative abilities [10,11]. Therefore, allogeneic BMDMSCs obtained from young healthy animals may provide a more optimal cell source for routine clinical usage. Moreover, more thorough cell characterization and quality controls are possible with allogeneic MSCs.
Allogeneic BMDMSCs may have several potential advantages, but further research is necessary to ensure that allogeneic BMDMSCs are safe and effective for clinical use. Evidence is mounting that BMDMSCs have potent anti-inflammatory and chondroprotective effects [14,15]. Experimental osteoarthritis studies have demonstrated an increase in comfort level after administration of BMDMSCs [14]. Likewise, a rabbit model of osteoarthritis using anterior cruciate ligament transection revealed less cartilage loss, decreased surface abrasion, increased cartilage content, and improved histologic scores when rabbits were administered intra-articular allogeneic BMDMSCs [15]. In terms of their anti-inflammatory properties, it is important to determine if allogeneic BMDMSCs produce the same degree of immunomodulation as that of autologous, as this could be an important characteristic for clinical use [16]. Guest et al. assessed the safety and survival of allogeneic MSCs in surgically induced lesions of the equine superficial digital flexor tendon and reported no detectable difference in the apparent immunogenicity of autologous versus allogeneic BMDMSCs [17]. Intra-articularly administered allogeneic umbilical cord blood-derived MSCs also appear to reduce inflammatory infiltrate in lipopolysaccharide (LPS)-stimulated joints [18], although there was no comparison with autologous MSCs in this study.
Clinically, the use of allogeneic MSCs has been described in studies by Broeckx et al. and Van Loon et al. where clinical cases of equine degenerative joint disease and equine tendon and ligament injury were treated with allogeneic MSCs and minimal adverse effects were reported [9,19 –21]. Likewise, a phase II clinical trial using allogeneic BMDMSCs in human patients with knee osteoarthritis has reported promising results of increased comfort and cartilage health [22]. Furthermore, 55 people received allogeneic MSCs following partial medial meniscectomy, resulting in significantly increased meniscal volume and increased comfort compared with a hyaluronate control [23]. A number of in vitro studies in other species indicate clearly that allogeneic BMDMSCs are immunomodulatory [24 –35]. However, studies that directly compare the immunomodulatory properties of autologous and allogeneic equine BMDMSCs have not, to our knowledge, been reported previously.
Therefore, the aim of the current study was to compare the immunomodulatory properties of autologous and allogeneic equine BMDMSCs and to identify mechanism(s) for the immunosuppressive properties of allogeneic BMDMSCs.
Materials and Methods
The experimental protocol described complied with the policies of the Institutional Animal Care and Use Committee at Colorado State University (Protocol 12–3483).
Bone marrow collection and stem cell expansion
Sternal bone marrow aspirates were collected from adult horses using 1,000 U/mL of heparin, as previously described [13]. Nucleated cells were then maintained on polystyrene culture flasks (Corning®) in Dulbecco's modified Eagle's medium (DMEM; Gibco®, Invitrogen) with 10% fetal bovine serum (Hyclone, Sigma-Aldrich®), 1% penicillin–streptomycin (Gibco), and 1 M HEPES buffer (Gibco™) in 5% CO2 at 37°C.
Once stem cell colonies (MSCs) were established, cells were harvested using Accumax™ cell dissociation solution (Sigma-Aldrich) as per the manufacturer's instructions. Cells were then maintained in culture in Minimum Essential Medium (MEM) Alpha Medium (αMEM) with 10% fetal bovine serum (Hyclone, Sigma-Aldrich), 1% penicillin–streptomycin (Gibco), and 1 M HEPES buffer (Gibco) in 5% CO2 at 37°C through a second passage. BMDMSCs were then cryopreserved in freeze media consisting of 95% Hyclone fetal bovine serum (Sigma-Aldrich) and 5% dimethyl sulfoxide at a concentration of 10 × 106 cells/mL.
Blood collection and preparation of peripheral blood mononuclear cells
Equine peripheral blood mononuclear cells (PBMCs) were prepared from whole anticoagulated blood collected by jugular venipuncture and density gradient centrifugation, as previously described [25]. PBMCs were cryopreserved in 95% fetal bovine serum and 5% dimethyl sulfoxide at a concentration of 10 × 106 cells/mL and stored in liquid nitrogen before use. For analysis, PBMCs were labeled with mouse anti-horse CD5 (Clone CVS5; Abd Serotec®) and mouse anti-human CD3 (Clone CD3-12; Abd Serotec) and secondarily labeled with anti-mouse FITC (Abd Serotech). CD3 and CD5 analysis was used to create a gate for lymphocytes. This gate was then used to select lymphocytes in future assays.
Lymphocyte proliferation assay
MSCs were recovered from cryopreservation and cultured in DMEM with 10% fetal bovine serum (Hyclone, Sigma-Aldrich), 1% penicillin–streptomycin (Gibco), and 1 M HEPES buffer (Gibco) in 5% CO2 at 37°C. MSCs were then harvested and placed in a 48-well plate at a concentration of 50,000 cells per well and cultured overnight. At least 12 h following plating of BMDMSCs, PBMCs were removed from liquid nitrogen, thawed in a 37°C hot water bath, and washed with phosphate-buffered saline. PBMCs used as negative controls were treated with 1 μM hydroxyurea for 1 h before labeling with carboxyfluorescein succinimidyl ester (Cell Trace™; Thermo Fisher Scientific). All other PBMCs were immediately labeled with 5 μM CFSE and added to MSCs at ratios of 1 MSC per 10 PBMCs, 1 MSC per 50 PBMCs, 1 MSC per 100 PBMCs, 1 MSC per 500 PBMCs, 1 MSC per 1,000 PBMCs, and 1 MSC per 2,000 PBMCs. Four allogeneic and four autologous combinations were analyzed for each ratio of MSCs to PBMCs. All cells were incubated in DMEM with 10% fetal bovine serum (Hyclone, Sigma-Aldrich), 1% penicillin–streptomycin (Gibco), and 1 M HEPES buffer (Gibco) for 4 days in 5% CO2 at 37°C. Cell cocultures were maintained for 4 days. Flow cytometry was performed using the CyAN™ ADP flow cytometer (Beckman Coulter®) to measure lymphocyte cell proliferation using CFSE staining by FITC fluorescence. Fluorescence data were analyzed using Summit Software (Beckman Coulter).
MSC titration for suppression of lymphocyte proliferation
MSCs were recovered from cryopreservation and cultured in DMEM with 10% fetal bovine serum (Hyclone, Sigma-Aldrich), 1% penicillin–streptomycin (Gibco), and 1 M HEPES buffer (Gibco) in 5% CO2 at 37°C. MSCs were then harvested and placed in a 48-well plate at a concentration of 50,000 cells per well and cultured overnight. At least 12 h following plating of the BMDMSCs, PBMCs were removed from liquid nitrogen, thawed in a 37°C hot water bath, and washed with phosphate-buffered saline. PBMCs used as negative controls were treated with 1 μM hydroxyurea for 1 h before labeling with CFSE. All other PBMCs were immediately labeled with 5 μM CFSE and stimulated with 2.5 μg/mL concanavalin A (Sigma-Aldrich). MSCs were added to PBMCs at a ratio of 1 MSC per 10 PBMCs, 1 MSC per 100 PBMCs, and 1 MSC per 1,000 PBMCs. Fifty-six allogeneic and 12 autologous combinations were evaluated. During the last 4 h of incubation, the cocultures were treated with 25 ng/mL phorbol 12-myristate 13-acetate (Sigma-Aldrich), 1 μM ionomycin (Sigma-Aldrich), and 10 ng/mL brefeldin A (BioLegend®), after which the cells were fixed in 2% paraformaldehyde. Cells were then permeabilized with 1% saponin buffer before staining using an intracellular mouse anti-bovine IFNγ:Alexa Fluor®647 (Clone CC302; AbD Serotech). Flow cytometry was performed using the CyAN ADP flow cytometer (Beckman Coulter) to measure lymphocyte cell proliferation using CFSE staining by FITC fluorescence and intracellular IFNγ production by APC647 fluorescence. Fluorescence data were analyzed using Summit Software (Beckman Coulter).
Assessment of mechanism of lymphocyte suppression by MSCs
MSCs were recovered from cryopreservation and cultured in DMEM with 10% fetal bovine serum (Hyclone, Sigma-Aldrich), 1% penicillin–streptomycin (Gibco), and 1 M HEPES buffer (Gibco) in 5% CO2 at 37°C. MSCs were plated at a concentration of 50,000 cells per well in a 48-well plate overnight. Allogeneic PBMCs were collected the following day by jugular venipuncture and processed using density gradient centrifugation [25]. Cells were used immediately without cryopreservation. PBMCs used as negative controls were treated with 1 μM hydroxyurea (Sigma-Aldrich) for 1 h before labeling with CFSE. All other PBMCs were immediately labeled with 5 μM CFSE and stimulated with concanavalin A (2.5 μg/mL). MSCs were tested at various ratios of MSC:PBMCs (1:5, 1:10, 1:100, 1:1,000) to identify an optimal ratio, which was determined by 1:10. To assess mechanisms of MSC inhibition of T cells, known biochemical inhibitors of immunosuppressive pathways in other species were evaluated [16,34 –36]. The inhibitors were used at previously published concentrations for indomethacin [16], L-NMMA [32], and 1-MT [33] and at two times the IC50 for SD208 [36].
Inhibitor concentrations included 10 μM indomethacin (PGE2 inhibitor, Sigma-Aldrich), 1 mM 1-methyl-
After 96 h in culture, the supernatants were removed from each of the wells and stored at −80°C for analysis. The cells were then fixed with 2% paraformaldehyde. Flow cytometry was performed using the Gallios™ flow cytometer (Beckman Coulter) to measure lymphocyte cell proliferation using CFSE staining by FITC fluorescence. Fluorescence data were analyzed using FlowJo (Version 10.0.8).
ELISA assay for prostaglandin E2
Prostaglandin E2 (PGE2) levels in the supernatants of untreated MSCs and PBMC coculture as well as the wells treated with indomethacin were evaluated using the prostaglandin E2 Parameter Assay Kit (R&D Systems®), as per the manufacturer's recommendations. Samples were read using a microplate reader (Molecular Devices®, SpectraMax M3) at 540 nm and quantified with a standard curve.
MHCII expression of MSCs exposed to lymphocyte-conditioned media
This assay was performed to analyze the effects of MSC exposure to PBMC-conditioned media from stimulated or unstimulated PBMCs on expression of MHCII by MSCs. Allogeneic PBMCs were collected by jugular venipuncture and processed using density gradient centrifugation [25]. A total of 500,000 PBMCs were added to each well of a 48-well plate. PBMCs were cultured in DMEM with 10% fetal bovine serum (Hyclone, Sigma-Aldrich), 1% penicillin–streptomycin (Gibco), and 1 M HEPES buffer (Gibco) in 5% CO2 at 37°C. Stimulated PBMCs were exposed to10 μg/mL of concanavalin A for 96 h. After 96 h, supernatants were collected from both stimulated and unstimulated PBMC wells.
Four BMDMSC lines, at passage 3, were recovered from cryopreservation and cultured in DMEM with 10% fetal bovine serum (Hyclone, Sigma-Aldrich), 1% penicillin–streptomycin (Gibco), and 1 M HEPES buffer (Gibco) in 5% CO2 at 37°C. When MSCs were 70%–80% confluent, they were dissociated from culture using Accumax™ and resuspended in a single-cell solution at 1 × 106 cells/mL of media (DMEM with 1% HEPES buffer). Cells were then added to a 96-well plate at a seeding density of 25,000 cells per well. Cells were allowed to attach overnight. The following day, cells were exposed to conditioned media from previously unstimulated or stimulated PBMC cultures. Conditioned media were diluted with control media to concentrations of 0% (control), 10%, 50%, and 100% conditioned media. After 96 h, supernatants were removed and all wells were washed with phosphate-buffered solution before dissociation using Accumax. Cells were resuspended in complete media before two concurrent washes and staining with a monomorphic mouse anti-horse MHCII:RPE antibody (Clone CVS20; BioRad®). All wells were blocked with 5% equine serum for 10 min before antibody exposure. Mouse IgG1: PE (Clone IS5-21F5; Miltenyi Biotech) was utilized as an isotype control. Cells were analyzed for MHCII expression using the FL2 channel. Results were reported as the % of cells expressing MHCII.
Statistical analysis
The continuous data were evaluated for normality. Immunogenicity data were evaluated using a nonparametric test due to low sample size. A Mann–Whitney test was used to compare immunogenicity outcomes with the PBMC-positive and -negative controls. Likewise, immunosuppression proliferation and interferon gamma data were found to be non-normal and evaluated using a Mann–Whitney test to compare proliferation of PBMCs cocultured with MSCs and the positive control (stimulated lymphocytes). For evaluating the cell mechanism data, a Kruskal–Wallis test was used with Dunn's multiple comparisons test. Finally, MHCII expression was evaluated using a two-way ANOVA with Tukey's multiple comparisons test. A P value of 0.05 was used to determine statistical significance. GraphPad Prism v 6.0 (La Jolla, CA) was used to perform the statistical analysis.
Results
Relative immunogenicity of allogeneic and autologous MSCs
To assess the ability of MSCs to stimulate spontaneous proliferation of resting T cells, both allogeneic and autologous MSCs were cocultured with PBMCs at ratios of 1 MSC to 10 PBMCs, 1 MSC to 50 PBMCs, 1 MSC to 100 PBMCs, and 1 MSC to 1,000 PBMCs for 96 h, and T cell proliferation assessed by flow cytometry. We observed a small but statistically significant increase in lymphocyte proliferation when both autologous and allogeneic MSCs were cultured with T cells at a ratio of 1 MSC per 10 PBMCs (P < 0.05) (Fig. 1). Importantly, however, the degree of T cell proliferation induced by autologous MSCs was not statistically different from that of allogeneic MSCs. Similar results were observed at other MSC-to-T cell ratios (data not shown). Thus, we concluded that equine allogeneic MSCs were not inherently more immunogenic than autologous MSCs in terms of T cell activation.
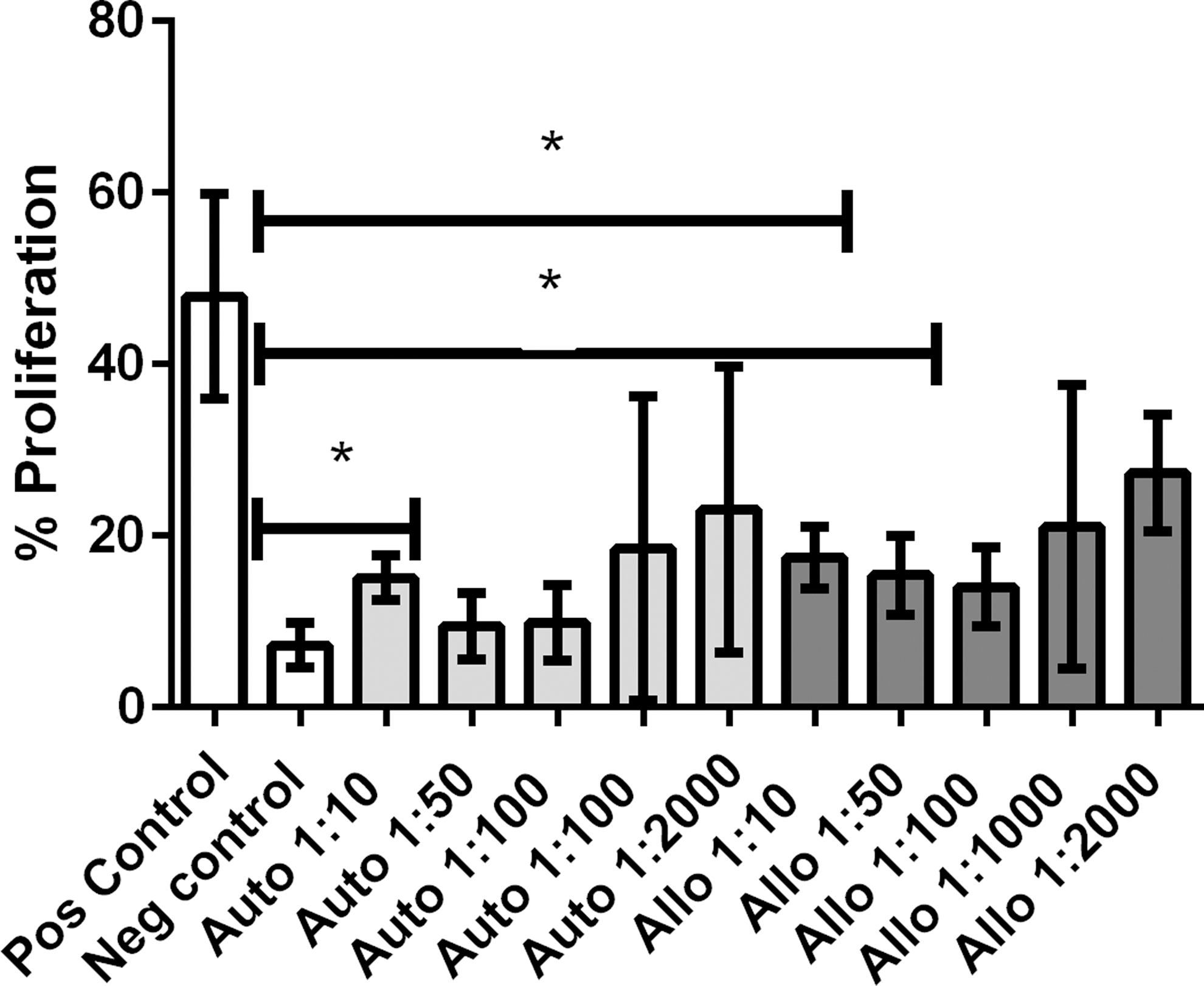
Lymphocyte proliferation assay: Relative immunogenicity of allogeneic and autologous MSCs. Allogeneic and autologous MSCs were nonimmunogenic at low ratios of MSC:PBMC. No difference was noted between the immunogenicity of allogeneic and autologous MSCs regardless of dilution. A mild increase in lymphocyte proliferation was noted when autologous BMDMSCs were cocultured with unstimulated lymphocytes at a ratio of 1 MSC per 10 PBMCs and when allogeneic BMDMSCs were cocultured with unstimulated lymphocytes at a ratio of 1 MSC per 10 PBMCs and 1 MSC per 50 PBMCs. Boxes indicate the average value ± SEM. P values <0.05 are marked by *. BMDMSCs, bone marrow-derived mesenchymal stem cells; MSC, mesenchymal stem cell; PBMC, peripheral blood mononuclear cell.
Suppression of T cell proliferation by autologous and allogeneic MSCs
Next, studies were conducted to determine whether allogeneic MSCs suppressed activated T cell responses to the same degree as autologous MSCs. We found that there was a significant and statistically equivalent degree of suppression of T cell proliferation by both allogeneic and autologous BMDMSCs (Fig. 2). For example, at an MSC-to-PBMC ratio of 1:10, there was significant decrease in lymphocyte proliferation induced by both autologous and allogeneic MSCs. However, there was no statistically significant difference in the degree of lymphocyte proliferation suppression induced by the two different MSC populations (autologous vs. allogeneic).

Immunosuppression assay (lymphocyte proliferation). Allogeneic and autologous BMDMSCs were immunosuppressive, demonstrated by a decrease in lymphocyte proliferation compared with the positive control. No difference was noted between allogeneic and autologous MSCs regardless of concentration. The greatest immunosuppression was noted at 1 MSC per 10 PBMCs. Boxes indicate the average value ± SEM. P values <0.01 are marked by **.
In addition, coculture with autologous and allogeneic BMDMSCs both resulted in significantly decreased IFNγ production by T cells (<0.05) (Fig. 3). However, there was no statistically significant difference in the degree of suppression of IFNγ production by autologous BMDMSCs when compared with suppression induced by allogeneic BMDMSCs regardless of dilution.
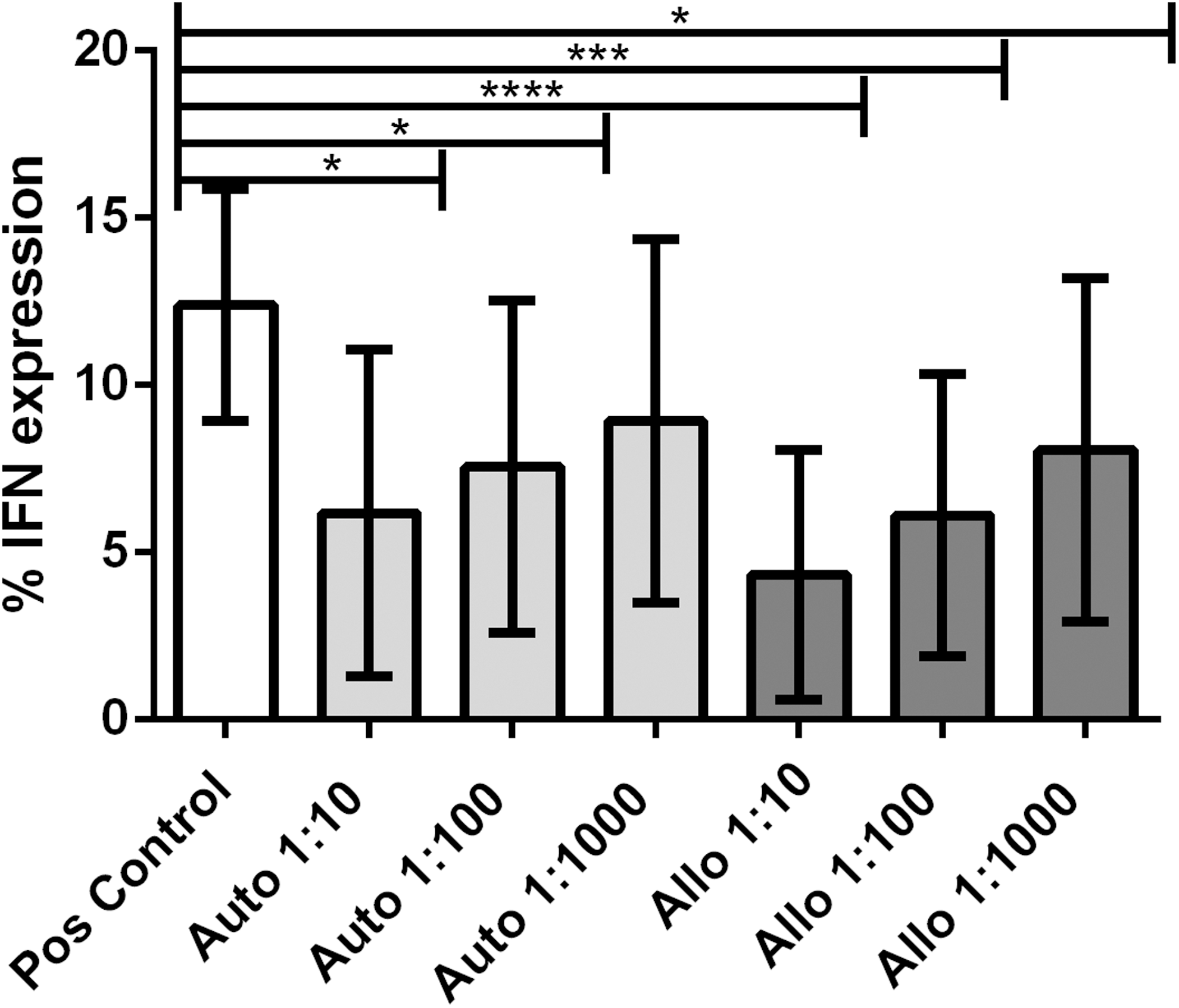
Immunosuppression assay (IFNγ expression). IFNγ expression decreased with increasing ratios of BMDMSCs to PBMCs. The greatest decrease in IFNγ expression was observed at a ratio of 1 MSC per 10 PBMCs. P values <0.05 are marked by *. Boxes indicate the average value ± SEM. P values <0.001 are marked by ***, and P values <0.0001 are marked by ****.
Mechanisms of T cell immunosuppression by equine MSCs
T cell proliferation, along with specific inhibitors of known pathways of MSC suppression in other species, were used to interrogate possible pathways of immunosuppression in the equine system. We found that incubation of MSCs with activated T cells in the presence of indomethacin (an inhibitor of the cyclooxygenase pathway) resulted in significant (P < 0.01) reversal of MSC-induced T cell proliferation suppression (Fig. 4). Moreover, we also found that incubation of MSCs with activated equine T cells resulted in production of significant amounts of PGE2 in vitro (Fig. 5). The amount of PGE2 in the supernatant of the MSC-T cell cocultures was significantly reduced by the addition of indomethacin (P < 0.05) (Fig. 5). In contrast, there was no effect of addition of inhibitors of the NO pathway (L-NMMA), the transforming growth factor-beta (TGFβ) pathway (SB208), or the IDO pathways (1-MT) when added at concentrations known to reverse MSC suppressive effects in other species [16,32,33] or at two times the IC50 for SD208 [36].
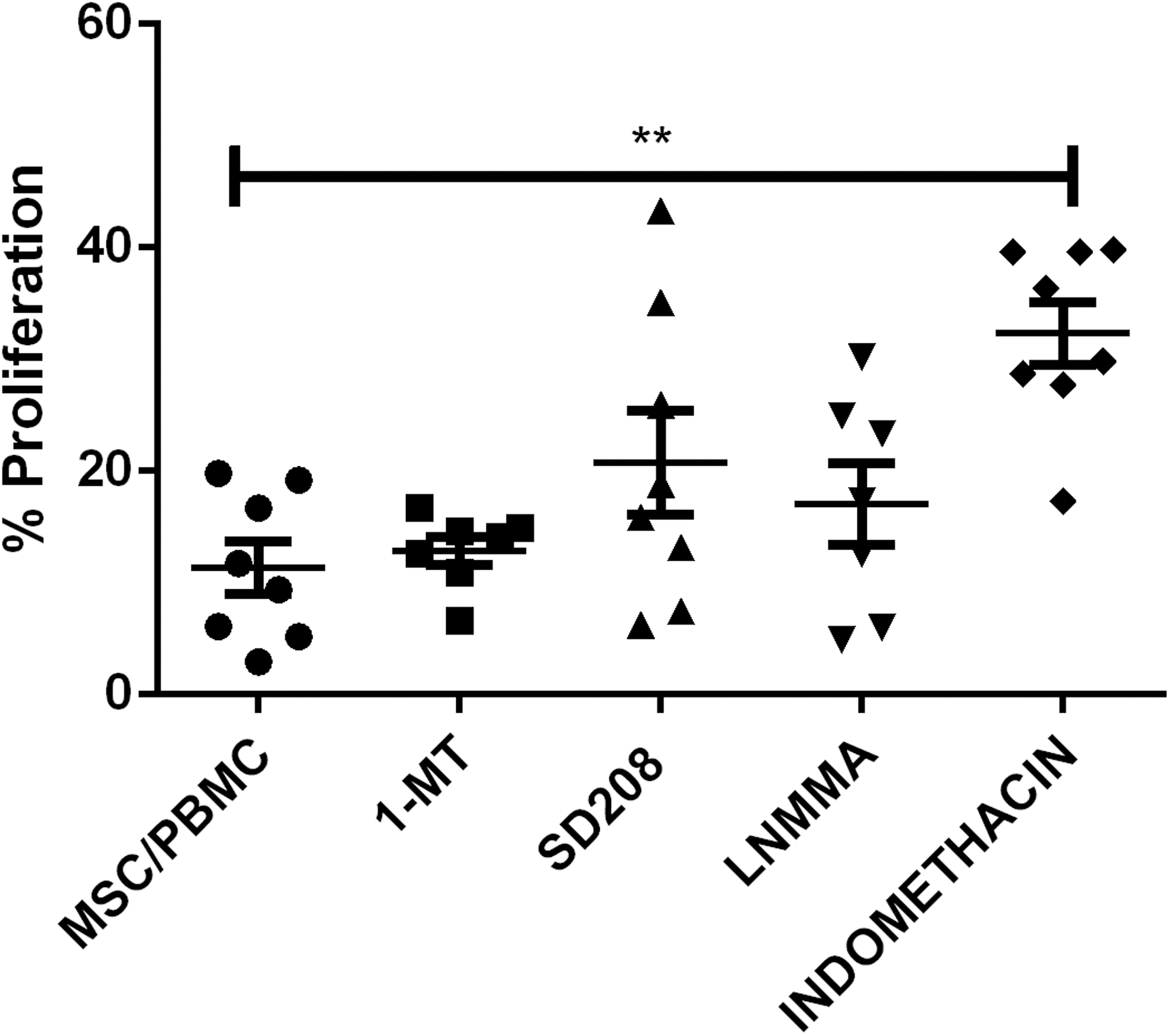
Immune mechanism assay. Inhibition of potential mediators of immunosuppression by 1 MT (indoleamine pathway), SD208 (TGFβ pathway), L-NMMA (nitric oxide pathway), and indomethacin (PGE2 pathway) revealed a reversal of immunosuppression only when cocultures were treated with indomethacin. The lines indicate the mean ± SEM. P values <0.01 are marked by **. PGE2, prostaglandin E2; TGFβ, transforming growth factor-beta.
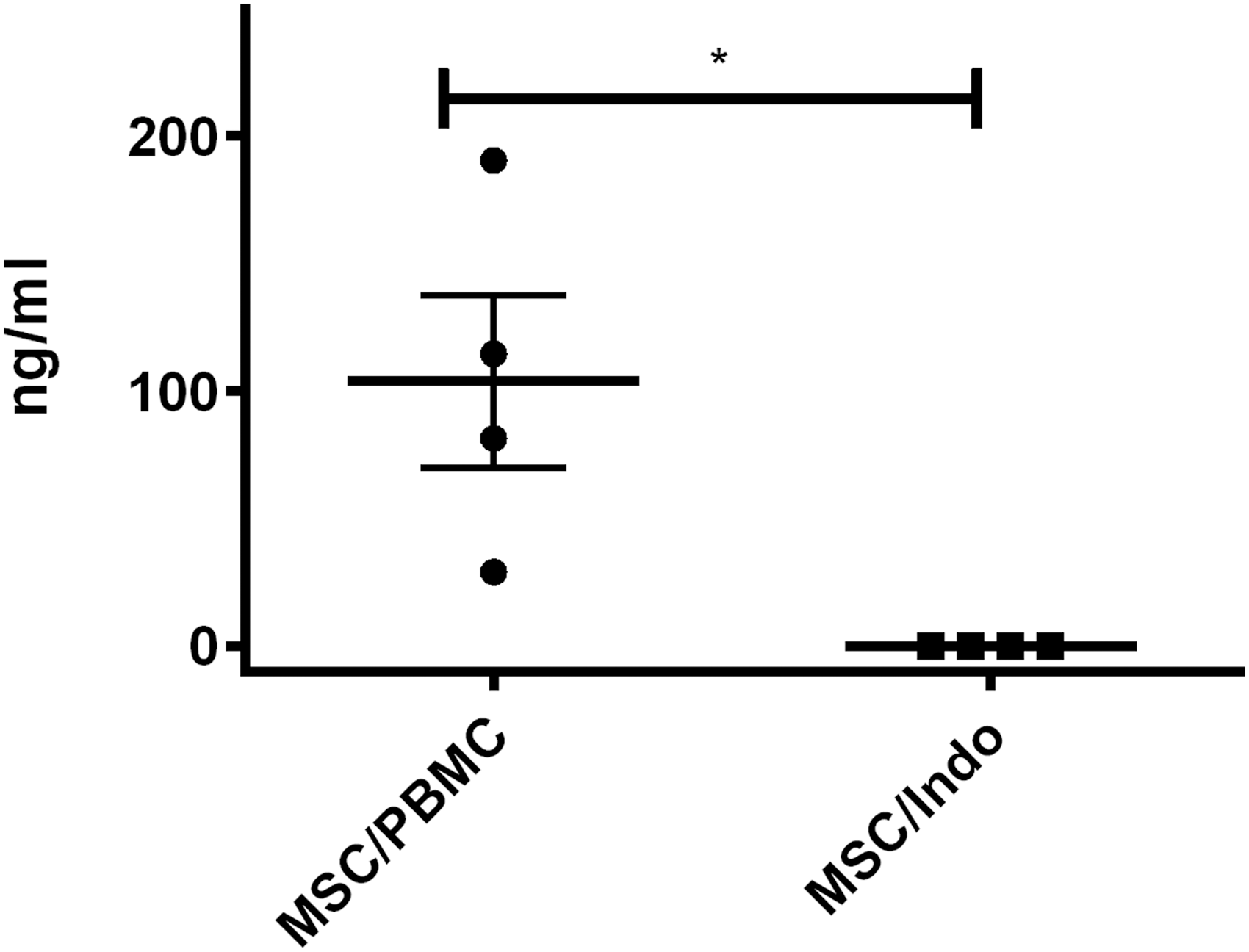
PGE2 levels in supernatants from BMDMSC/PBMC cocultures and indomethacin-treated cocultures. PGE2 levels were significantly increased in BMDMSC/PBMC cocultures compared with cocultures treated with indomethacin. The lines indicate the mean ± SEM. P values <0.05 are marked by *.
MHCII expression of MSCs after coculture with stimulated PBMCs
Flow cytometry was used to evaluate the effects of cytokines released from activated T cell cultures on MHCII expression by MSCs. MHCII expression by MSCs exposed to conditioned media from unstimulated PBMCs or MSCs exposed to conditioned media from stimulated PBMCs was determined using MSCs from four different horses. It should also be noted that the PBMCs were collected from horses unrelated to the MSC donors. The level of MHCII expression was found to be significantly increased in MSCs exposed to 50% and 100% conditioned media from stimulated PBMCs (Fig. 6). Exposure of MSCs to conditioned media from unstimulated PBMCs (100%) also caused an increase in MHCII expression compared with MSCs grown in control media (Fig. 7). Less than 2% of MSCs expressed MHCII when incubated in control media, and expression of MHCII remained under 10% for all conditions (Figs. 6 and 7).
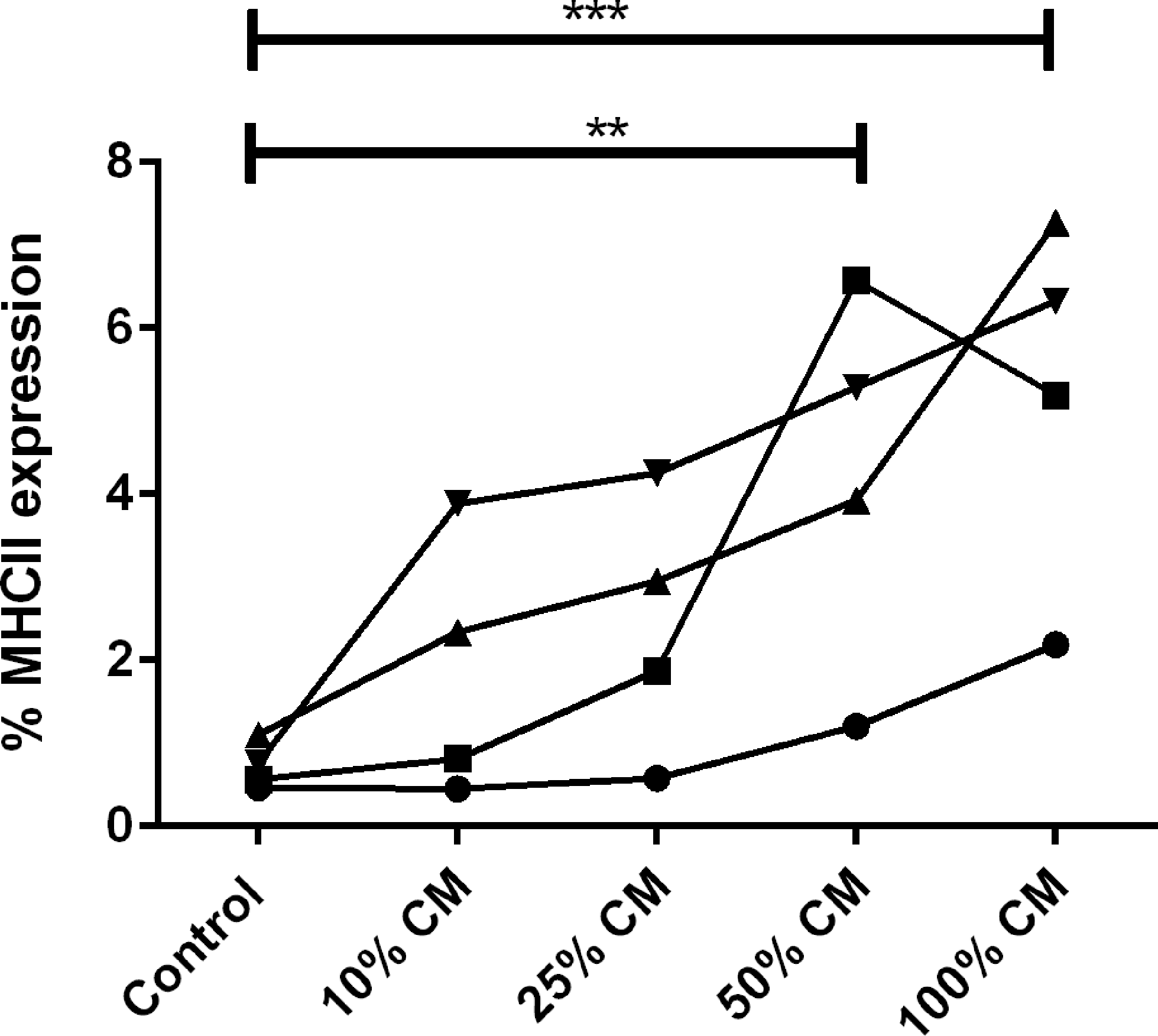
MHCII expression of MSCs following exposure to conditioned media from stimulated PBMCs. MHCII expression was found to increase in MSCs exposed to conditioned media from stimulated PBMCs when 50% and 100% conditioned media were used. P value <0.01 is marked by **, and P value <0.001 is marked by ***.
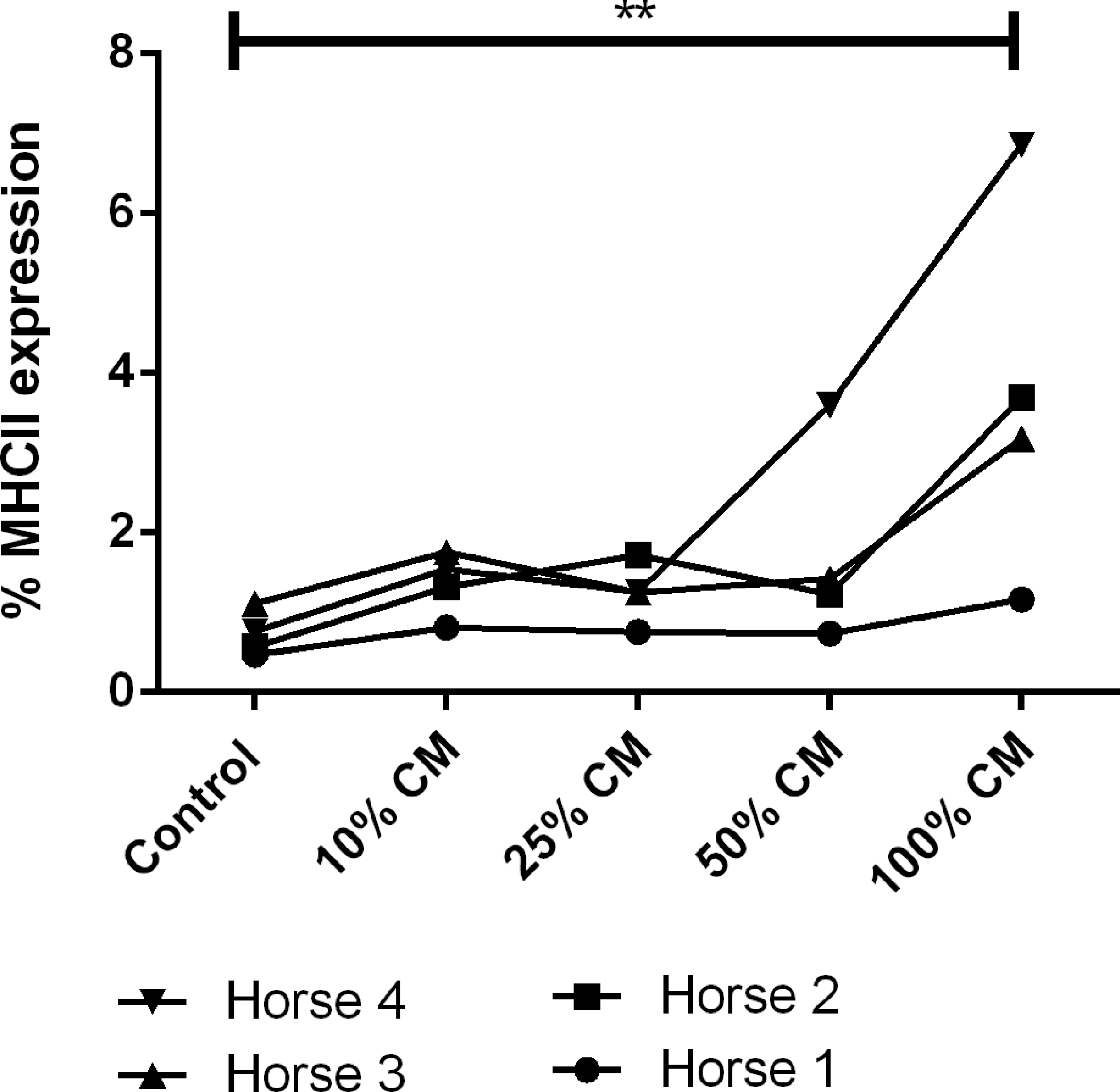
MHCII expression of MSCs following exposure to conditioned media from unstimulated PBMCs. MHCII expression was found to increase in MSCs exposed to conditioned media from unstimulated PBMCs when 100% conditioned media were used. P value <0.01 is marked by **.
Discussion
Autologous BMDMSCs are extensively used for the treatment of osteoarthritis, tendonitis, and desmitis in the horse. However, little is known about the immunomodulatory properties of allogeneic equine BMDMSCs. Previous studies have shown that equine allogeneic MSCs are immunomodulatory in vitro [16,37,38], but these studies did not directly compare their immunosuppressive properties with those of autologous MSCs in side-by-side assays. Without such a direct comparison, the relative immunomodulatory effectiveness of the two cell sources cannot be determined with certainty.
To determine whether equine allogeneic MSCs can activate resting T cells, allogeneic BMDMSCs were mixed with resting lymphocytes from unrelated donor animals (allogeneic reaction) and with lymphocytes from the same animal (autologous reaction). A small degree of lymphocyte activation was observed with both MSC types, but the degree of activation was similar with both (Fig. 1). Thus, allogeneic MSCs do not appear to be capable of generating strong alloreactive T cell responses in previously untreated horses and the small degree of activation with allogeneic MSCs is not more than autologous MSCs.
Previous in vitro studies using human and equine BMDMSCs support this finding; Mancheño-Corvo et al. demonstrated an increase in the immunosuppressive properties of MSCs preactivated with IFNγ, suggesting that local inflammation is an important trigger for MSC immunosuppression [28]. Likewise, Paterson et al. revealed a decrease in the production of inflammatory cytokines (IL-6, INFγ, TNFα) by antigen-stimulated PBMCs when they were cocultured with MSC supernatants [39]. In agreement with in vitro studies [28], a recent in vivo equine study demonstrated that allogeneic umbilical MSCs cause a decrease in inflammation on joints treated with LPS [18]. A local inflammatory environment may be important for the activation of MSC anti-inflammatory mechanisms and should be further investigated.
In our study, coculture of allogeneic and autologous BMDMSCs with stimulated lymphocytes resulted in equivalent immunosuppression as defined by a decrease in lymphocyte proliferation (Fig. 2). Our study indicates that the source of BMDMSCs (allogeneic vs. autologous) is not an important variable in determining the degree of immunosuppression elicited in vitro. This finding could have significant clinical implications as allogeneic BMDMSCs may provide a more convenient and less expensive product for the treatment of musculoskeletal disease in the horse. These findings suggest that further in vivo studies are warranted to compare the behavior of allogeneic and autologous cells within the inflamed joint environment.
The pathways of the immunosuppressive properties of human, murine, and canine BMDMSCs have been previously investigated [29,40,41]. Our study examined the role of TGFβ, PGE2, indoleamine, and reactive nitrogen species as mediators of immunosuppression and found that only PGE2 was an important mediator of immunosuppression by allogeneic equine BMDMSCs (Fig. 4). This finding is in agreement with a previous study that investigated interleukin-6 (IL-6), nitric oxide (NO), and PGE2 as mediators of immunosuppression by allogeneic BMDMSCs [16] and found only PGE2 to be important.
We also observed that the concentration of PGE2 was significantly increased in cocultures of allogeneic BMDMSCs and PBMCs as a result of MSC cytokine production, confirming its upregulation during immunosuppression (Fig. 5). PGE2 has been extensively studied in conditions of acute and chronic inflammation. PGE2, a naturally occurring prostaglandin, suppresses acute inflammation by influencing neutrophils, macrophages, and T cells [42]. Understanding how allogeneic BMDMSCs create an anti-inflammatory environment is important to harnessing their full potential as a therapeutic modality and directing their clinical use. This study was able to identify a single paracrine factor, PGE2, utilized by allogeneic BMDMSCs.
We found no significant effect of inhibitors of the nitric oxide pathway (L-NMMA), the TGFβ pathway (SD-208), or the IDO pathways (1-MT) when added to the MSCs and T cell cocultures. Previous equine studies have demonstrated L-NMMA and indomethacin to be active against the nitric oxide and indomethacin pathways, respectively [16]. However, other equine studies have been unable to detect a difference in the level of TGFβ in cocultures of MSCs and lymphocytes when compared with lymphocytes alone, suggesting that TGFβ is unlikely to be involved as with immunomodulation [37]. We performed a TGFβ-1 ELISA (Human TGF-β1 Immunoassay; R&D Systems), which revealed no difference in the levels of TGFβ in the coculture system with and without SD-208 (data not shown). In addition, 1-MT has not been previously used as a modulator of indoleamine secretion in the horse. However, in agreement with our findings, previous studies have not detected indoleamine in equine cocultures of MSCs and PBMCs, suggesting that this pathway may not be functionally active [37].
Our study focused on detecting soluble paracrine factors associated with immunomodulation by BMDMSCs. Recent literature suggests that BMDMSCs may utilize multiple pathways to achieve immunosuppression, including a combination of direct cell-to-cell contact and paracrine stimulation [43]. Although multiple mechanisms are likely involved, a recent in vitro study using human MSCs suggests that the majority of immunomodulation occurs through paracrine secretion, as opposed to the effects of direct cell-to-cell contact between lymphocytes and MSCs [44].
Previous studies have used intra-articular administration of allogeneic MSCs to treat joint disease in horses [9]. Broeckx et al. used immature and chondrogenically induced allogeneic MSCs in 165 horses with a complication rate of only 1.8% consisting entirely of joint flares [9], a rate similar to that published for intra-articular administration of autologous BMDMSCs [7]. In humans, a multicenter phase II clinical trial using allogeneic MSCs for knee osteoarthritis has reported improvement in algofunctional indices and cartilage quality compared with a hyaluronic acid control [22]. Likewise, a rabbit model of osteoarthritis has reported an increase in cartilage health with intra-articular BMDMSC administration, demonstrating allogeneic stem cell survival and engraftment [15]. Evidence is mounting that allogeneic MSCs may be safe and effective in vivo.
Recent publications have examined the expression of major histocompatibility complex II (MHCII) by equine BMDMSCs [45 –47]. Expression of MHCII may lead to immune recognition and, therefore, is a potential concern for allogeneic use. Direct contradiction exists in the literature with the majority of publications demonstrating a lack of MHCII expression by BMDMSCs [17,45,47], while an alternate publication reports MHCII expression in 11 of 13 lines of BMDMSCs [46]. In our laboratory, MSCs are routinely MHCII negative (data not shown). However, a prior study by Schnabel et al. found that exposure of MSCs to 100 ng/mL of recombinant IFNγ would cause MSCs that lacked MHCII expression to markedly increase expression of MHCII [46]. Lymphocytes in culture produce IFNγ, and stimulation with concanavalin A causes an increase in IFNγ production. Therefore, it was our expectation that MSCs would increase their expression of MHCII when exposed to conditioned media from concanavalin A-stimulated PBMCs and unstimulated PBMCs. As expected, we found an increase in MHCII expression after MSCs were exposed to conditioned media from PBMCs (Figs. 6 and 7). However, despite evidence of an increase in MHCII expression after MSCs are exposed to conditioned media PBMCs, 56 allogeneic coculture combinations were found to demonstrate immunosuppression in vitro. This indicates that either the utilized allogeneic BMDMSCs had low MHCII expression despite coculture with stimulated lymphocytes or that MHCII status cannot be directly linked with the ability to cause immunomodulation in mixed lymphocyte reactions.
Based on the findings from our study, we suggest that further research should be conducted in vivo to compare the relative clinical benefits of the anti-inflammatory and immunomodulating properties of allogeneic BMDMSCs with autologous BMDMSCs. If allogeneic BMDMSCs are found to have comparable immunomodulating properties with autologous BMDMSCs in vivo, they may provide a beneficial off-the-shelf therapy for musculoskeletal disease in horses, eliminating the need for time-consuming, individual culture techniques and the added expense of individual culture.
In conclusion, allogeneic BMDMSCs and autologous BMDMSCs appear to be equally immunosuppressive in vitro. It also appears that equine MSCs principally use the cyclooxygenase pathway for suppression of T cell function.
Footnotes
Acknowledgment
The authors would like to acknowledge the Grayson Jockey Club Foundation for their generous support of this work.
Previous publication: portions of the results from this study have been previously presented in oral presentation and abstract form for AO North America Conference, Las Vegas, NV, February 5–7, 2014, and the American College of Veterinary Surgeons Surgery Summit, San Diego, CA, October 16–18, 2014. In addition, data from the study were presented in poster form at the Orthopaedic Research Society Annual Meeting, Las Vegas, NV, March 24–28, 2015.
Author Disclosure Statement
Dr. Laurie R. Goodrich and Dr. C. Wayne McIlwraith are currently shareholders in Advanced Regenerative Therapies.
The other authors report no conflicting financial interests.