
Abstract
High post-transplantation cell mortality is the main limitation of various approaches that are aimed at improving regeneration of injured neural tissue by an injection of neural stem cells (NSCs) and mesenchymal stromal cells (MStroCs) in and/or around the lesion. Therefore, it is of paramount importance to identify efficient ways to increase cell transplant viability. We have previously proposed the “evolutionary stem cell paradigm,” which explains the association between stem cell anaerobic/microaerophilic metabolic set-up and stem cell self-renewal and inhibition of differentiation. Applying these principles, we have identified the main critical point in the collection and preparation of these cells for experimental therapy: exposure of the cells to atmospheric O2, that is, to oxygen concentrations that are several times higher than the physiologically relevant ones. In this way, the primitive anaerobic cells become either inactivated or adapted, through commitment and differentiation, to highly aerobic conditions (20%–21% O2 in atmospheric air). This inadvertently compromises the cells' survival once they are transplanted into normal tissue, especially in the hypoxic/anoxic/ischemic environment, which is typical of central nervous system (CNS) lesions. In addition to the findings suggesting that stem cells can shift to glycolysis and can proliferate in anoxia, recent studies also propose that stem cells may be able to proliferate in completely anaerobic or ischemic conditions by relying on anaerobic mitochondrial respiration. In this systematic review, we propose strategies to enhance the survival of NSCs and MStroCs that are implanted in hypoxic/ischemic neural tissue by harnessing their anaerobic nature and maintaining as well as enhancing their anaerobic properties via appropriate ex vivo conditioning.
Ischemic Stroke: Epidemiology, Pathophysiology, and Therapeutic Approaches
S
This gives rise to two distinct tissue zones: (1) The ischemic core, initially comprising necrotic tissue, gradually being converted into a region characterized by pan-necrosis and the emergence of a pseudocyst, which represents non-salvageable tissue; (2) The penumbra, that is, the peri-infarct area between the lesion core and the intact region of the brain, representing tissue [5,7,8] that can be rescued by treatment.
The penumbra is characterized by mild-to-moderate ischemia, depending on maintained thresholds of cerebral blood flow energy metabolism, and protein synthesis [9].
In the acute phase, clinical management of stroke aims at restoring cerebral blood-flow in affected brain areas and may involve recanalization with stent retrievers and aspiration thrombectomy [10] or intravascular thrombolysis by administration of tissue plasminogen activator (tPA) [11]. In the subacute phase, secondary brain edema may cause a life-threatening increase in intracranial pressure, necessitating hemicraniectomy.
Despite the fact that such interventions may dramatically improve survival and/or functional outcomes, their efficacy is largely dependent on the time elapsed between the stroke incident and medical treatment. It is estimated that 2 million neurons get lost every minute after the stroke onset. In cases of large vessel occlusions, this means that for every hour without clinical intervention, 120 million neurons, 830 billion synaptic connections, and 714 km of myelinated fibers are lost [12].
Considering the high metabolic demands of the brain and the rapid tissue loss after ischemia, thrombolytic treatment can achieve optimal results only if administered within 3–4.5 h after stroke onset. This effectively means that only <10% of patients are eligible for thrombolytic treatment [13]. In some patients, thrombolysis can cause post-ischemic cerebral reperfusion injury such as cerebral edema, symptomatic intracranial hemorrhage, BBB disruption, or penumbral deterioration [3,14]. A number of caveats, including serious other unwanted side-effects, cause the majority of the new thrombolytic or neuroprotective compounds, aimed at limiting neuronal cell death, to fail in clinical trials [15 –18].
In the sub-acute phase, stroke patients may demonstrate a degree of spontaneous improvement, which may be related to genetic factors such as single-nucleotide polymorphisms, including Vall66Met, in the brain-derived neurotrophic factor (BDNF) gene, APOE4, COX-2, IGF1, and GPIIIα [19,20]. Spontaneous improvement can also be further enhanced with appropriate rehabilitation in the chronic phase. Observed functional outcomes appear to be consistent with a degree of rewiring of surviving neuronal networks and recruitment of intact synapses, which occurs mainly contralateral and, to a lesser degree, ipsilateral to the lesion [21].
Depending on the stroke severity and type of clinical/rehabilitation treatment, these mechanisms may account for the spontaneous functional recovery observed in some stroke patients 30–90 days post-ischemia. This is especially with regard to alleviation of language and cognitive impairments, as well as motor impairments such as voluntary maximum arm extension [22]. Furthermore, evidence from animal studies provides valuable insights into molecular mechanisms of spontaneous recovery after stroke, including structural remodeling at the axonal, dendritic, and synaptic level, activation and migration of endogenous neural stem cells (NSCs), angiogenesis, as well as changes in the extracellular matrix (ECM) [23 –30].
In the previous decade, increased responsiveness in terms of onset-to-treatment times, in combination with enhanced rehabilitation strategies, has improved the overall survival rate of stroke patients by 20%. However, despite this progress, 50% of stroke survivors suffer from severe motor, sensory, and cognitive impairments, with significant implications for their quality of life and high associated socioeconomic costs [31]. As a result, ongoing research efforts focus on further development of alternative or complementary therapies.
Cell Therapy for Stroke: Problem of Cell Survival on Injection in Ischemic and Hypoxic Zone
Stem cell therapy is one of the new therapeutic approaches that potentially can extend the therapeutic window from the acute into the sub-acute or chronic stage. NSCs as well as mesenchymal stem cells (MSCs) and mesenchymal stromal cells (MStroCs) are highly relevant in this context. Numerous experimental studies have demonstrated the potential of NSCs and MStroCs in promoting transplant-mediated repair with functional restoration through such diverse mechanisms as neuroprotection, cell replacement, remyelination, neovascularization, and de novo neurogenesis [20,32,33].
The efficacy of these approaches largely depends on the lesion topology and site of transplantation. Stem cell engraftment typically targets the ischemic penumbra/peri-infarct area, where the host tissue microenvironment is less harsh to the transplanted cells, rather than the ischemic core [34,35]. Despite the fact that the ischemic penumbra represents salvageable tissue, survival of transplanted cells is still very poor, resulting in low engraftment efficiency [20,31]. Furthermore, to restore brain function, an effective transplantation strategy will also need to overcome the barrier of regeneration at the lesion site [32]. In situ tissue engineering strategies aim at addressing this issue by utilizing natural and synthetic biopolymer matrices to promote stem cell graft survival as well as lesion scaffolding and targeted, controlled delivery of pharmacotherapeutics [32,36 –39].
Although such strategies hold great promise as they can potentially address multiple aspects of stroke lesion pathology, their utility may, indeed, be limited unless the cells of choice have been processed in an optimal manner before transplantation. Here, we review how ex vivo cell culture conditions may affect post-transplantation cell survival and discuss how physiological pre-conditioning, which takes into account the metabolic state of the stem cells as well as that of the host tissue microenvironment, may optimize engraftment efficiency in the ischemic core or penumbra/peri-infarct area.
Harnessing Stem Cell Anaerobic Properties to Improve Neuronal Tissue Cell Therapy
Stem cell-based approaches for stroke patients may address cell replacement, enhanced trophic/regenerative support from the transplanted cells, immunomodulation, and stimulation of endogenous brain repair processes (such as angiogenesis, arteriogenesis, neurogenesis, synaptogenesis, and white matter remodeling) [40]. However, in clinical settings to date, stem cell transplantation into infarcted regions has been characterized by low graft survival and/or cell differentiation [41]. Application of physiological low oxygen (O2) preconditioning (usually inaccurately called “hypoxia”) of stem cells for therapeutic purposes in central nervous system (CNS) lesion models in vivo has yielded some positive results.
For quite a long time, the standard conditions for ex vivo culture of human neural and stromal cells have been set to the atmospheric 21% oxygen concentration, which is in stark contrast with the physiological levels of O2 within the niches in which stem cells, including MSCs, usually reside in the body [42].
The two niches in the body where MSCs can be found in most abundance, the bone marrow and the adipose tissue, have an O2 concentration between 1%–7% and 10%–15%, respectively [43 –46]. Even stem cells in the developing fetus seem to be exposed to low O2 concentrations. The female reproductive tract was shown to be physiologically characterized by low O2 concentrations (1.5%–8% O2). These conditions prevail throughout fetal development, with a dissolved oxygen concentration in fetal circulation rarely exceeding 5% [47 –49]. Thus, a low O2 concentration characterizes the microenvironment of all fetal cells, especially that of stem cell niches, including MSC.
Thus, the efficacy of stem cells in neural tissue repair can also be significantly influenced by the ex vivo oxygenation conditions, considering that effective oxygen concentration in the developing and adult brain is several times lower than the atmospheric O2 levels of 20%–21% [50,51].
In the human brain, oxygen concentration is highest in the alveolar space (14%) and lowest within tissues, with values ranging from 8% (19–40 mmHg) in the pia down to 0.55% (4.1 mmHg) in the midbrain (reviewed in Zhang et al. [52]). Large variations in oxygen concentration have been measured over small distances in the dura: 3% (23.2 mmHg) at a depth of 22–27 mm, as opposed to 4% (33 mm Hg) at 7–12 mm [53]. Furthermore, ex vivo cellular responses involving hypoxia-inducible factor 1-alpha (HIF1-α) and Notch signaling, which are traditionally termed as “hypoxic,” occur at O2 concentrations of <5% [54], that is, at physiological brain tissue conditions [55]. It is important to distinguish this physioxia from the real hypoxia, which is a term denoting a pathological state. Unfortunately, in the literature, the term “hypoxia” tends to be used for physiologically relevant O2 concentrations, which results in confusion.
It has been demonstrated that a state of reduced tissue oxygenation represents the state of normoxia (physioxia) for stem cells, particularly with regard to brain tissue as the host microenvironment [53,56].
Indeed, atmospheric O2 levels (20%–21%) represent hyperoxia (Fig. 1) for the stem cells and enhance their commitment and differentiation rates, leading to the exhaustion of stemness [56]. The cellular mechanisms that take place in the context of aerobic adaptation result in the transformation of cell populations naturally exhibiting anaerobic/facultative aerobic metabolic features (stem cells) into aerophylic/aerobic ones (committed progenitors and precursors). This process is reminiscent of the evolution of single-celled eukaryotes from simple anaerobic to increasingly specialized aerobic ones (see “Evolutionary stem cell paradigm” in Ivanovic and Vlaski-Lafarge [56]). In fact, even a short exposure of cells to atmospheric high O2 concentration during their collection and manipulation negatively affects their primitiveness and high proliferative capacity [57].
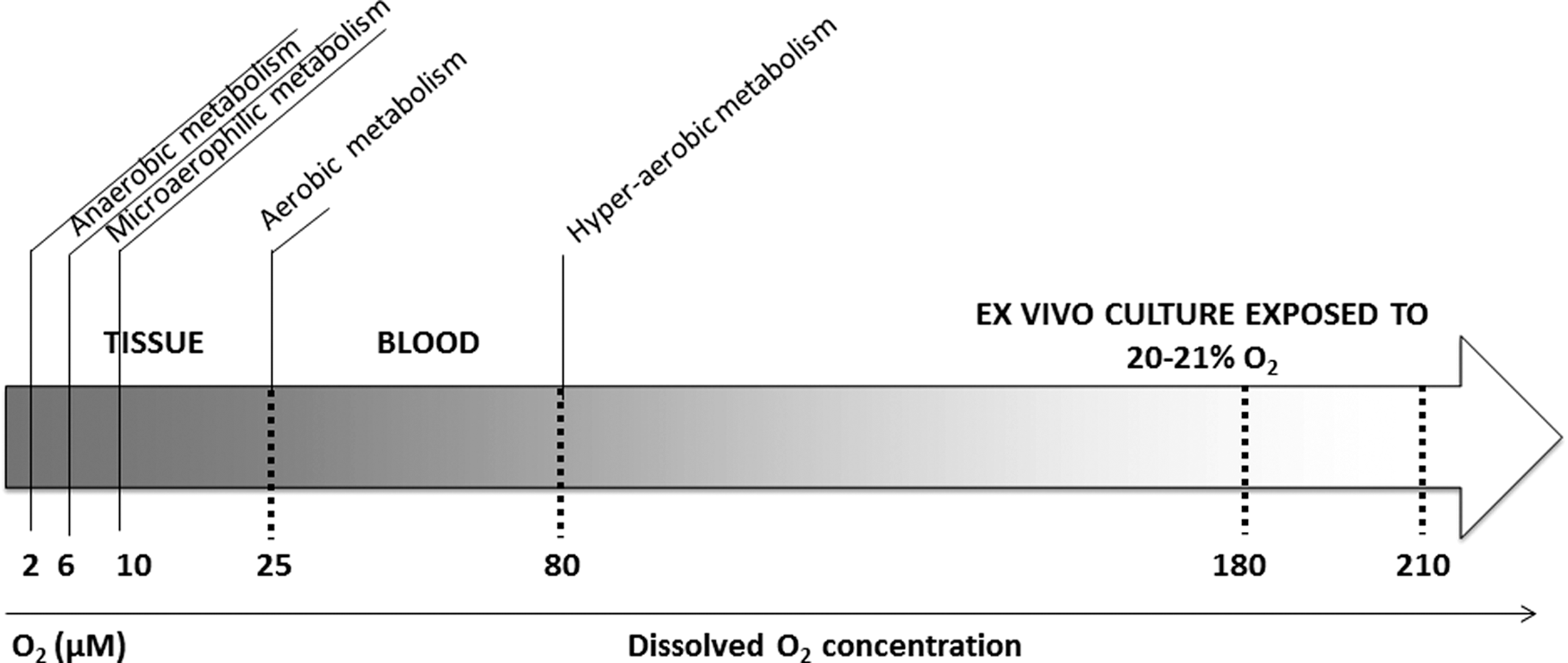
Types of energy metabolism in mammalian cells in relation to dissolved O2 concentration (at the cellular level). The data used for this figure are reviewed in Chapter 2 (“In situ normoxia versus ‘hypoxia’”) of our recent book [56].
These observations led to the conclusion that exposing MStroCs to low oxygen concentrations during ex vivo cell culture before transplantation can better approximate physiological conditions. In this way, preconditioning these cells, especially their MSC subpopulation, may enhance cell survival and function after transplantation into healthy brain tissue and, importantly, ischemic brain regions.
Experimental findings from animal models have, indeed, demonstrated the benefits of “preconditioning” of MStroCs by exposure to low O2 concentration before transplantation. This procedure can modulate the paracrine effects of MSCs, causing upregulation of various secretable factors, including vascular endothelial growth factor (VEGF) and interleukin-6 (IL-6). Moreover, preconditioning can improve MSC mobilization and homing, primarily by inducing stromal cell-derived factor-1 expression along with its receptor, CXC chemokine receptor 4 (CXCR4) [58,59].
Hu et al. have shown that even short-term (24 h) preconditioning at a low, that is, a physiologically relevant O2 concentration of 0.5% O2 significantly enhances bone-marrow (BM)-derived MStroC migration via the Kv2.1 channel. The Kv2.1 channel activates focal adhesion kinase (FAK), enhances cell motility, induces the FAK-Kv2.1 complex formation, and upregulates CXCR4 in BM-MSCs [59,60]. Similarly, Wei et al. showed that exposure of BM-MStroC to 0.5% O2 for 24 h increased their survival and improved functional recovery in a rat stroke model, compared with BM-MStroCs that were cultured at atmospheric O2 levels [61]. Exposure to a low O2 concentration upregulated HIF-1α and trophic/growth factors such as BDNF, glial cell-derived neurotrophic factor (GDNF), VEGF and its receptor FIK-1, erythropoietin (EPO) and its receptor EPOR, and stromal-derived factor-1 (SDF-1) and its CXCR4 [40].
Wei et al. also showed that many pro-inflammatory cytokines/chemokines were downregulated in MSCs that were cultured under physiologically relevant low O2 concentrations. Compared with control cells that were cultured at atmospheric O2, the transplanted low-O2-preconditioned cells showed enhanced homing ability due to increased expression of CXCR4 [61,62]. Berniakovich and Giorgio showed that the culture of mouse BM-MStroCs at 3% O2 augmented cell proliferation, promoted colony formation [colony-forming unit fibroblast (CFU-F) assay], and also increased the percentage of quiescent cells and the expression of stemness markers Rex-1 and Oct-4 [63]. In contrast, intrinsic differentiation processes were inhibited by 3% O2. Furthermore, these authors found that culture at 3% O2 affected BM-MSCs differentiation in p53- and reactive oxygen species (ROS) independent pathways. Overall yield in terms of differentiation was contingent on the adjustment of O2 tension to the specific stage of BM-MSCs culture. The authors concluded that a suitable strategy for efficient BM-MSC ex vivo differentiation should involve an initial phase of cell propagation at 3% O2, followed by a differentiation stage at 21% O2 [63].
In addition to bone marrow, MStroCs can also be derived from the mucoid connective tissue of the human umbilical cord that is referred to as Wharton's jelly (WJ). The cells from this source are very interesting for regenerative medicine given that they can be obtained from medical waste without any risk to the donor. Compared with BM-MstroCs, under a low-oxygen atmosphere, WJ-MstroCs exhibited a higher proliferation rate, greater expansion capability, enhanced neurotrophic factor expression, and a tendency for spontaneous commitment and differentiation toward neural lineage, as confirmed by protein and gene marker induction [64]. Drela et al. showed that preconditioning by exposure of WJ-MStroC [the term “WJ-MSC” (Wharton's jelly Mesenchymal Stem Cells) is used by the authors] cultures to 5% O2 led to increased CFU-F frequencies as well as to expression of Oct4 and Nanog along with Hif1-α and Hif2-α. This was paralleled by a reduction of further spontaneous differentiation into a neuronal phenotype [65].
Teixeira et al. examined the effects of “normoxic” (21% O2) and “hypoxic” (5% O2) conditions on the neuroregulatory gene expression profile of human WJ MStroC secretome [66]. Subsequently, they examined the impact of the WJ MStroC secretome at 5% O2 versus 21% O2 on neuronal differentiation and survival of human neural progenitor cells (NPCs). Their results indicate that hWJ-MStroC secretomes collected from “normoxic” and “hypoxic” conditions had similar effects on neuronal differentiation of human NPCs both ex vivo and in vivo. However, proteomic analysis revealed that the use of “hypoxic” preconditioning led to the upregulation of several proteins within the WJ-MStroC secretome, including GDN, Cys C, UCHL1, clusterin, Prx1, 14–3-3, thymosin-beta, and EF-2, all of which are involved in the inhibition of apoptosis and the promotion of neuroprotection, angiogenesis, and neuronal cell survival and differentiation.
In fact, this so called “hypoxic” upregulation rather approximates a physioxic secretome, that is, it corresponds to the secretome of cells exposed to O2 concentrations that are equivalent to physiological in situ ones. On the other hand, the secretome is altered during adaptation of the cells to a hyperoxic atmospheric O2 concentration (20%–21% O2).
The protective and immunosuppressive potential of 5% O2 preconditioned WJ-MStroC [the term “WJ-MSC” (Wharton's jelly Mesenchymal Stem Cells) is used by the authors] and endothelial progenitor cells derived from WJ-MStroC (WJ-EPCs) was recently documented by Obtulowicz et al. [67] in an ex vivo oxygen–glucose deprivation (OGD) stroke model of organotypic hippocampal slices (OHC). WJ-EPCs were shown to secrete neuro-, angio-, and immunomodulatory molecules such as IL-6, TGF-b1, and VEGF. Furthermore, soluble factors released by both WJ-MStroCs and induced WJ-EPCs primed at 5% O2 had a protective effect on the hippocampus against OGD-induced damage. The innate immunosuppressive potential of WJ-MStroCs and induced WJ-EPCs, evidenced by basal TGF-b1, IL-6, and VEGF secretion, was only minimally affected by proinflammatory LPS and poly(I:C) stimulation.
It should be emphasized that in most of the experiments cited earlier, the MStroC were isolated and ex-vivo expanded in atmospheric O2 concentrations (20%–21%), that is, effectively hyperoxic conditions, before being re-exposed to the physiologically relevant low O2 concentrations post-transplantation into the brain parenchyma. Only the data of Obtulowicz et al. [67] are based on WJ-MStroC cultures expanded at 5% O2; however, the final co-culture experiments with the OGD hippocampal slides had to be performed (24 h) at the ambient (21%) oxygen level, due to the lack of the organotypic culture stability in 5% O2 ex vivo conditions.
As explained in detail in Chapter 4 of our recent book [56], ex vivo MStroC expansion in ambient O2 concentration results in aerobic adaptation of the heterogeneous stromal cell population. In such a population, only a small percentage of cells are multipotent MSCs and lineage-committed progenitors whereas more than 50% are post-progenitor stromal cells, which can still differentiate into mature cells. Clearly, re-exposure to physiologically relevant (low) O2 concentrations may be deleterious for cells that are highly adapted to aerobiosis during culture in atmospheric O2 concentration, whereas cells still preserving some anaerobic and microaerophilic properties will better maintain their proliferative and differentiation capacity.
Thus, the best strategy for maintaining the stemness of these cells would be to avoid oxygenation during and after collection as well as during ex-vivo manipulation. Indeed, this approach has been shown to promote the maintenance of a primitive MSC population, coupled with an extensively high proliferative and differentiation potential [68,69]. It is usually pointed out that the stem cells in physiologically relevant O2 concentrations exhibit a glycolytic metabolic profile. However, recent findings suggest that stem cells under the total absence of O2 (anoxia) as well as in conditions of reduced oxygen and nutrients (as in brain hypoxia/ischemia) may engage mitochondrial anaerobic respiration (“fumarate respiration”) (Chapter 9 [56]) to produce energy, a mechanism known to be employed by some anaerobic parasites and tumor cells. This can be a very important point considering the survival of stem cells that are engrafted in hypoxic/ischemic brain regions where neither glucose nor O2 is available.
In addition to MS/StroCSs, a number of studies concluded that physiologically relevant O2 concentrations can also strongly influence NSC growth and differentiation mechanisms both in vivo and ex vivo [41,55,70,71]. In their review, De Filippis and Delia conclude that the physiological conditions of 2.5%–5% O2, which is typical of neural tissues, particularly promote NSC self-renewal. Furthermore, physiologically relevant O2 values favor the success of engraftment when ex vivo expanded NSCs are transplanted into the brain of experimental animals [53]. This is in line with recent findings showing that ex vivo preconditioning with 5% O2 for 72 h promotes the proliferation of NSCs that are isolated from the embryonic day 14 rat cerebral cortex [72].
In addition to what has been described earlier, Jäderstad et al. demonstrated that “hypoxic” preconditioning increases gap-junction intercellular communication between grafted NSCs and host tissue cells, that is, one of the key functional interactions after NSC engraftment that can promote CNS repair. Gap-junction communication is also known to increase in the CNS under pathological conditions such as brain hypoxia and ischemia. Specifically, the authors hypothesized that controlled “hypoxic” preconditioning of murine NSCs (C17.2) before engraftment is a reliable method to increase connexin 43 expression and to improve subsequent graft-host communication. Their findings demonstrated that NSC exposure to physiologically relevant O2 concentrations for 3 h increased the number of connexin 43 aggregates by 31%. This was paralleled by enhanced hemichannel function shown by faster calcein dye efflux and an augmentation of the early functional graft-host communication [73].
In line with this scenario are the ex vivo findings showing that the effects of the “hypoxic” preconditioning were attenuated in murine NSCs by pharmacological inhibition of cystine glutamate exchanger system Xc-activity with S4-carboxyphenylglycine [74]. This glutamate exchanger system has been demonstrated to exhibit neuroprotective effects on “hypoxic” stimulation [75]. These data are particularly relevant when discussing the metabolic considerations related to anaerobiosis and stem cells, especially with regard to NSCs.
It is interesting to mention a study by Cheng et al., who generated NPCs by combining a treatment with chemical cocktails and exposure to “hypoxia,” that is, a physiologically relevant O2 concentration. Specifically, they treated mouse embryonic fibroblasts with a chemical cocktail, namely VCR (V, VPA, an inhibitor of HDACs; C, CHIR99021, an inhibitor of GSK-3 kinases; and R, Repsox, an inhibitor of TGF-β pathways), under 5% O2. This led to the generation of chemically induced NPCs, which closely resembled mouse brain-derived NPCs in terms of their proliferative and self-renewing abilities, gene expression profiles, and multipotency for different neuroectodermal lineages not only ex vivo but also in vivo, as evidenced 7 days after transplantation in the brain of E13.5 pups [76].
An overview of the effects of various preconditioning protocols is presented in Table 1.
Designation used in the original article.
Designation of cell population according to the terminology employed in this article.
BDNF, brain-derived neurotrophic factor; BM, bone marrow; CFU-F, colony-forming unit fibroblast; CXCR4, CXC chemokine receptor 4; EPC, endothelial progenitor cell; EPO, erythropoietin; GDNF, glial cell-derived neurotrophic factor; HIF, hypoxia-inducible factor; IL-6, interleukin-6; MSC, mesenchymal stem cell; MStroC, mesenchymal stromal cell; NSC, neural stem cell; SDF-1, stromal-derived factor-1; VEGF, vascular endothelial growth factor; WJ, Wharton's jelly.
Anaerobic/Microaerophilic Regulation of MSCs and NSCs
The molecular machinery employed in the maintenance of stemness, particularly the mechanisms implicated in self-renewal, is aligned with the anaerobic/microaerophilic metabolic environment (Table 2 [56, Chapter 7]). The associated properties originate in early single-cell eukaryote evolution and are, in fact, a feature that is directly related to the maintenance of a primitive cell character.
Low O2 concentration ≤5% O2; high O2 concentration 20%–21% O2.
mTOR, mammalian target of rapamycin; RNS, reactive nitrogen species; ROS, reactive oxygen species.
HIFs, predominately HIF-1 and HIF-2 transcription factors, are the key mediators of cellular response in a low (<5%) O2 environment [77,78]. HIF deficiency, which leads to the functional disruption of stem cells, is associated with metabolic changes, suggesting that HIF-mediated “hypoxia adaptive cellular response” underlies pluripotent and adult stem cell maintenance that is relevant to both MStroC and NSCs.
The metabolic functions of HIF-1α are mostly associated with stimulation of the glycolytic metabolic shift [79,80], lipid [81] and nucleotide biosynthesis [82], and protection from oxidative stress [83]. Moreover, HIFs are implicated in the control of the critical molecular regulators of stemness at low O2 concentrations. [56]. Namely, HIF-1α modulates Wnt/β-catenin signaling by enhancing β-catenin activation and expression of other downstream effectors (such as LEF-1 and TCF-1). This causes a reduction in quiescence and self-renewal in pluripotent, embryonic stem cells (ESCs) as well as in adult cell populations containing stem cells, including MStroCs and NSCs, particularly in the hippocampal subgranular zone (SGZ), which is one of the NSC niches in the brain [84,85]. Interestingly, such regulation is not observed in differentiated cells [86].
Another gatekeeper for the self-renewal of stem cells is Notch signaling. In the nervous system, induction of Notch initiates progenitor quiescence. On the other hand, if Notch signaling is blocked, NSCs start dividing and committing toward neuronal fate [87]. This illustrates the importance of Notch signaling in maintaining a balance between quiescence and activation to commitment of adult NSCs.
In the bone marrow, Notch signaling usually acts to maintain a pool of mesenchymal progenitors while suppressing osteoblast differentiation [88]. In the low O2 environment of the stem cell niche, Notch signaling is stimulated in either an HIF-1-dependent or an HIF-1-independent manner [89,90]. HIF-1α potentiates Notch signaling by stabilizing the Notch intracellular signaling component (Notch intracellular domain, N ICD). HIF-1α physically interacts with Notch 1 ICD and accompanies it to Notch-responsive promoters to activate target genes. In addition, a low O2 environment also stimulates expression of the Notch ligand Delta4, which, in turn, stimulates further Notch signaling [89].
The next critical signaling pathway associated with stem cell self-renewal is Sonic Hedgehog (Shh) signaling [91], which is induced at low O2 concentrations and is mediated by HIF-1 [92]. Shh has been shown to mediate the activity of resveratrol (trans-3, 5, 4′-trihydroxystil), a naturally occurring polyphenolic phytoalexin with anti-inflammatory and neuroprotective properties. Shh can also enable NSC proliferation in anoxic/ischemic conditions ex vivo [93]. Similarly, Shh signaling is involved in the maintenance of MSC proliferation and clonogenicity [94].
In conditions characterized by low O2 availability, additional signaling pathways are influenced, which subsequently affect stem cell quiescence [95]. For example, anaerobic stabilization of HIF-1α promotes p53 activity [96], which is associated with the maintenance of NSCs as well as of MStroCs quiescence [97,98].
In addition, the genes that maintain ESCs in the pluripotent state (ie, Sox, Oct4, Nanog, KLF4, and Rex1) are also expressed in MStroCs [99] and are induced at low O2, thus being associated with the maintenance of the undifferentiated state of MStroCs [65,100,101].
In addition to what has been stated earlier, the mammalian target of rapamycin (mTOR)-regulated activity, which is implicated in fine-tuning the balance between NSC and MStroC self-renewal and differentiation, is modulated by a low O2 environment [102 –104].
Another important aspect of stem cell maintenance is their high vulnerability to oxidative stress that takes place when ROS production overcomes antioxidative defense activity. This situation is potentiated at O2 concentrations that are higher than physiological ones at the tissue site/niche in question [56]. To counteract these harmful effects, stem cells are equipped with highly expressed anti
For example, Ataxia telangiectasia mutated (ATM) is a key regulator of the cell cycle checkpoint in response to DNA damage. It is directly activated by ROS-induced cysteine residue oxidation, which, in turn, stimulates an anti-oxidative cell response. Loss of ATM activity, leading to an increase in the levels of ROS, coincides with impaired NSC self-renewal [106]. The underlying mechanisms have been shown to involve ROS-mediated activation of p38 mitogen-activated protein kinase (MAPK). In addition, p38 MAPK oxidation is implicated in the regulation of MStroC proliferation and differentiation [107].
Furthermore, FOXO transcription factors also play a critical role in NSC and MSC fate decisions, by activating expression of genes that encode enzymes that are required for detoxification of ROS/reactive nitrogen species (RNS) [108 –111], thereby influencing the balance between stem cell proliferation and commitment/differentiation.
Taken together, MSCs and NSCs as well as other stem cells have an intrinsic anaerobic molecular set-up that determines their survival and functionality. This set-up preserves essential cellular and molecular mechanisms that regulate pluripotency and self-renewal and render the cells resilient in adverse conditions. By the same token, an understanding of these mechanisms lends the cells malleable to manipulations that can effectively maintain stemness and, hence, function and survival. Thus, manipulations that aim at promoting the intrinsic “primitive” anaerobic molecular set-up before transplantation in the ischemic/hypoxic brain can be expected to significantly promote integration with the damaged host tissue and the potential for functional restoration (Fig. 2).
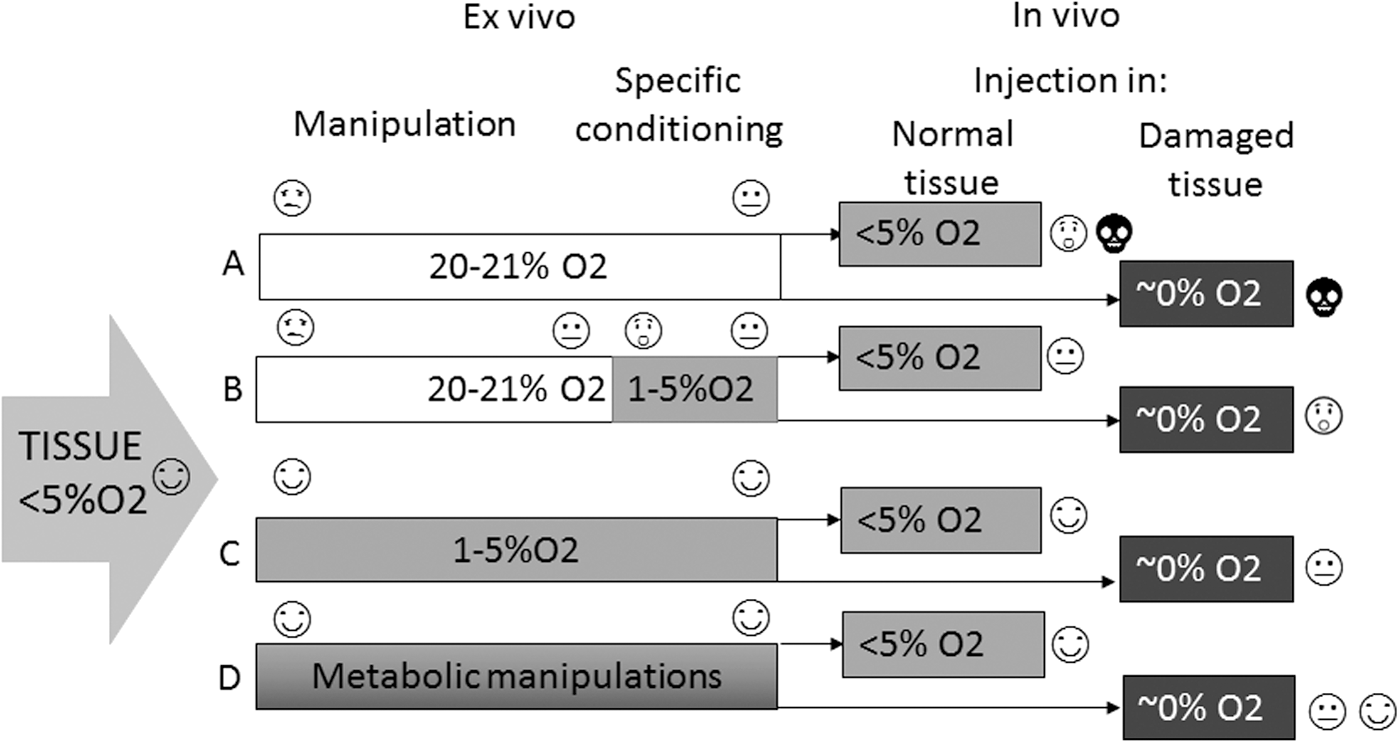
Impact of ex vivo O2 concentration on the survival of cell suspensions containing stem cells.
Conclusions and Perspectives
Efficient engraftment in transplant-mediated repair of stroke-induced lesions must take into account ex vivo optimization and preconditioning of the stem cells in accordance with intrinsic metabolic needs and in anticipation of the physiological conditions of the host microenvironment. This approach can be expected to enhance the viability, differentiation potential, and functionality of the transplanted stem cells per se, and to further promote the efficacy of relevant in situ tissue engineering strategies.
Importantly, such an approach directly addresses key recommendations of the NIH/FDA presented in the “Stem Cells as an Emerging Paradigm in Stroke” (STEPS) I − III, guidelines [112 –115] in the context of improving the validity of preclinical stroke models for safe clinical translation of transplant-mediated therapy. Among other requirements, the STEPS guidelines strongly emphasize the need for rigorous evaluation of the therapeutic cell type, including a better understanding of the relevant mechanism(s) of action post-transplantation.
To standardize conditions to obtain therapeutically efficient MStroCs or NSCs, one may need to prevent the emergence of the most adverse events associated with prolonged cell culture, such as failure of proliferation or cell genome transformation. This can be achieved, simply by changing culture conditions from a 21% to a ≤5% O2 concentration, which physiologically occurs in the majority of tissues and stem cell niches in vivo [116]. Clearly, optimization of the cells' physiological condition and their anaerobic/ischemic conditioning before transplantation will not only provide a better starting point as to predicting engraftment efficacy but also constitute a more appropriate basis for elucidation of their mechanism of action in situ.
In addition to this, the principles presented in this review can be expected to enable a better understanding of both endogenously and exogenously triggered restorative responses of endogenous stem cells that can act synergistically to promote brain repair. These often overlooked responses include neuronal plasticity, neurogenesis and angiogenesis [117], spontaneous functional recovery [117], rewiring of surviving neural networks and axonal ramification, the recruitment of intact synapses post-lesioning [23], and the activation of the adult neurogenic niches, that is, the subependymal zone of the lateral ventricle and the SGZ of the hippocampus [30], followed by subsequent migration of neural progenitors to the injury site, and potential maturation into functional neurons with concommittal integration into the local circuitry [118]. Consideration of the physioxic state of the niche microenvironment [56—Chapters 3 and 5,119] and of the dynamic cytogenic and metabolic changes of the niche in response to ischemic injury [29,30,120] may inform researchers on how to best explore stem cell-based treatments and their potential efficacy in promoting repair after ischemic stroke.
Finally, there is a brief reference to induced pluripotent stem cells (iPSCs), whose discovery by Takahashi and Yamanaka in 2006 [121] hailed a new era in the field of regenerative medicine. Patient-specific iPSCs can be obtained from minimally invasive skin biopsies, thus enabling the generation of theoretically unlimited numbers of autologously derived cells with multipotency and potential therapeutic benefits that are akin to those of ESCs and fetal cells, effectively circumventing some of the ethical and practical considerations involved in obtaining and using the latter. In addition, more recent advances in reprogramming technologies have enabled the generation of transgene-free patient-specific iPSCs using non-integrated viruses or with direct delivery of reprogramming proteins entirely in the absence of viral vectors, thus rendering iPSCs a very attractive candidate cell type for clinical application [122,123].
Equally promising approaches for stroke repair include the direct conversion of somatic cells into neurons [124] and the reprogramming of iPSCs into specific neuronal subtypes [125 –128]. Furthermore, different genome editing strategies, for example, selective expression and activation of suicide genes, can be applied to eliminate potential tumorigenic risks of iPSCs as well as of ESCs [20,129], thus improving their safety for clinical application. Transition of bioenergetics from an oxidative to an anaerobic state precedes the induction of pluripotency [130], which is accompanied by a reversion of somatic mitochondria to an ESC type in terms of morphology, expression of nuclear factors involved in mitochondrial biogenesis and distribution, and mitochondrial DNA content ([131]; reviewed in Saito et al. [132]). This evident anaerobic nature of iPSCs suggests the relevance of preconditioning at low O2 for their maintenance. Conversely, directing the energetic metabolism of iPSCs toward an aerophylic and aerobic profile is associated with commitment and differentiation of iPSCs [131].
Therefore, the anaerobic nature of MSCs, NSCs, and iPSCs should not be neglected when exploring multiple factors that can influence amplification, commitment, and subsequent differentiation to specific neuronal subtypes. Such a perspective can significantly maximize engraftment efficacy in stroke repair.
Footnotes
Acknowledgments
The authors acknowledge the help of Ms. Elisabeth Doutreloux-Volkman in writing and editing this article. The work was supported by: the French Blood Institute (Etablissement Français du Sang) (M.V.-L., L.R., Z.I.), the Liaison Committee between the Central Norway Health Authority (RHA) and the Norwegian University of Science and Technology (NTNU) (I.S., A.S.), the Norwegian Financial Mechanism 2009–2014 and the Czech Ministry of Education, Youth and Sports under project contract no. MSMT-28477/2014 (S.I., S.A.), the Polish National Centre for Research and Development grant no. Strategmed1/234261/2/NCBR/2014. (L.B., A.S.), and the Ministry of Education and Science of Serbia (grants ON174028 and III41007) (I.G.). The D.L. was supported by a French Government PhD fellowship.
Author Disclosure Statement
No competing financial interests exist.