
Abstract
HMGB1 and HMGB2 proteins have been implicated in numerous cellular processes, including proliferation, differentiation, apoptosis, and tumor growth. It is unknown whether they are involved in regulating the typical functions of pluripotent human embryonic stem cells (hESCs) and/or those of the differentiated derivatives of hESCs. Using inducible, stably transfected hESCs capable of shRNA-mediated knockdown of HMGB1 and HMGB2, we provide evidence that downregulation of HMGB1 and/or HMGB2 in undifferentiated hESCs does not affect the stemness of cells and induces only minor changes to the proliferation rate, cell-cycle profile, and apoptosis. After differentiation is induced, however, the downregulation of those proteins has important effects on proliferation, apoptosis, telomerase activity, and the efficiency of differentiation toward the neuroectodermal lineage. Furthermore, those processes are affected only when one, but not both, of the two proteins is downregulated; the knockdown of both HMGB1 and HMGB2 results in a normal phenotype. Those results advance our knowledge of regulation of hESC and human neuroectodermal cell differentiation and illustrate the distinct roles of HMGB1 and HMGB2 during early human development.
Introduction
H
Of the four HMGB-type proteins in humans and mice (HMGB1–4), HMGB1 is the most studied. HMGB2, HMGB3, and HMGB4 are selectively downregulated during embryonic development and are expressed only in certain tissues in adult organisms, whereas HMGB1 continues to be expressed in all embryonic and adult cells (reviewed in Müller et al. [4]). HMGB1 resides primarily in the nucleus and serves as a DNA chaperone to regulate transcription, replication, recombination, repair, and genome stability [5]. HMGB1 can also translocate to the cytoplasm and affect apoptosis/autophagy, or it can even be released to the outside of cells, where it binds several cell-surface receptors to regulate immunity, inflammation, tumor growth, and metastasis (reviewed in Andersson and Tracey [6]). In vivo experiments on HMGB1−/− or HMGB2−/− knockout mice suggest that HMGB proteins might be, to some extent, redundant [7,8], as the single-knockout pups are born viable. A number of other reports show that HMGB1 is highly expressed in isolated human inner cell mass (ICM) [9] and physically interacts with OCT4 [10], although the role of such interaction has never been addressed in detail. HMGB2 has also been linked to stem cell biology, interacting with Oct4 in mouse ESCs [11] and with cyclin-dependent kinase 2 (CDK2) in hESCs [12]. Studies of mouse models revealed that HMGB1 and HMGB2 are involved in embryonal neurogenesis [13 –15]. Comparison of those results to human neuroectodermal development is lacking, however, as is any information on the putative interaction between HMGB1 and HMGB2 during embryonic development and stem cell differentiation.
We aimed to uncover the roles of HMGB1 and HMGB2 in undifferentiated hESCs and in human neuroectodermal cells (hNECs) differentiated in vitro from hESCs. We prepared doxycycline (Dox)-inducible, stably transfected hESCs capable of shRNA-mediated HMGB1 and/or HMGB2 knockdown. Altogether, our data reveal that the nonhistone, DNA-binding proteins HMGB1 and HMGB2 are dispensable for hESC self-renewal but play important, possibly opposing, modulatory roles during the early onset of mammalian differentiation.
Materials and Methods
Construction of inducible HMGB-shRNA plasmid vectors
Smith-Waterman and BLAST sequence alignments were used to design a number of sequences for HMGBs knockdown by plasmid-based shRNA interference and the shRNA constructs were tested for the most efficient HMGB silencing. The specificity of our shRNA HMGB knockdown constructs was further verified by studying the impact of at least two different constructs for a given HMGB gene, and by using the corresponding “scrambled” shRNA constructs in several human cell lines ([16,17] and unpublished data, see also Discussion section). The outcome of the latter experiments was also compared with the impact of HMGB1 or HMGB2 knockout on telomerase activity in mouse embryonic fibroblasts [18]. Top and low oligonucleotides derived from human HMGB1 or HMGB2 cDNAs (for the preparation of specific shRNAs for HMGB1, HMGB2, or HMGB1/2 silencing; Supplementary Table S1; Supplementary Data are available online at
Culture of hESCs and generation of stable transfectants
Undifferentiated hESCs (line CCTL14) used in this study were previously thoroughly characterized for the expression of pluripotency transcription factors and their ability to differentiate as described [19]. CCTL14 line of hESC is also registered in the global registry for human pluripotent stem cell lines (
Differentiation of neuroectodermal cells from hESCs
hESCs were seeded at a density of 103 cells/cm2 as single cells on culture dishes coated with Matrigel and cultured in neurodifferentiation media (described in Supplementary Table S2) at 37°C and 5% CO2 in a humidified atmosphere. After 5 days, typical neuroectodermal cell morphology was observed. Neural rosettes appeared after 8–10 days.
Crystal violet cell-growth assay
Approximately 4 × 103 cells were seeded in triplicate in a 96-well plate, incubated at 37°C for 24–96 h, washed twice with phosphate-buffered saline (PBS), and fixed with 4% paraformaldehyde for 20 min at room temperature (RT). The cells were then washed twice with PBS and incubated with 0.1% crystal violet (in Milli-Q H2O) for 20 min at RT. The cells were then washed thrice with dH2O to remove excess stain. The crystal violet was then extracted using 33% acetic acid for 20 min at RT with gentle shaking. The optical density of the extracted dye was determined by measuring the absorbance at 570 nm using a microplate reader (Tecan Spectrofluoroplus;
Western blotting
To prepare total cell lysates, cells were washed twice with PBS and resuspended in lysis buffer [50 mM Tris-HCl (pH 6.8) containing 10% glycerol and 1% sodium dodecyl sulfate (SDS)]. Western blotting was performed as described previously [21,22]. The primary and secondary antibodies used are listed in the Supplementary Table S3. Protein expression was visualized using a SuperSignal West Dura Kit (ThermoFisher Scientific). The intensities of the bands were quantified using the ImageJ software (
Flow cytometry analysis of the cell cycle and proliferation index count
Cells were harvested using TrypLE Select (ThermoFisher Scientific), washed with PBS, and immediately fixed with ice cold 70% ethanol (v/v). After fixation, the cells were washed with PBS and treated with RNaseA (final concentration 0.02 mg/mL; Boehringer Ingelheim,
Immunocytochemistry
Cells were washed with PBS and fixed with 4% paraformaldehyde at RT for 20 min, permeabilized with 0.1% Triton X-100 for 20 min, and incubated in 1% bovine serum albumin in PBS for 1 h. The cells were then washed with PBS and incubated with primary antibodies against HMGB1, HMGB2, and PAX6 at 4°C overnight. The PBS-washed cells were then incubated with AlexaFluor-conjugated secondary antibodies (listed in Supplementary Table S3) for 2 h. Cell nuclei were then counterstained with DAPI (Sigma-Aldrich).
Annexin V-FITC/propidium iodide double-staining assay
Apoptosis was assessed by flow cytometry using an Annexin V/propidium iodide double-staining kit (ApoFlowEx® FITC Kit; EXBIO,
Teratoma formation assay
A teratoma formation assay was used to assess the capacity of the hESCs to differentiate in vivo. Undifferentiated hESCs were kept in Dox-containing media for three passages. Subsequently, an inoculum of uniform clumps of hESCs (1 × 106 cells in 100 μL culture medium) was injected into the hindlimb muscle of 8-week-old NOD/SCID/IL2Rg-null mice. Dox was added to the mice's drinking water at a concentration of 0.2 mg/mL throughout the study. When tumor diameters reached ∼1 cm, the mice were euthanatized by cervical dislocation. The tumors were dissected from the surrounding tissue, quickly frozen at −80°C, and then sectioned and stained with hematoxylin and eosin to visualize the tissue composition. All animal studies were carried out according to approved guidelines (Project No. 02/2010).
Image acquisition and analysis
Images of cells grown in 24-well plates were acquired using an Image Xpress XL automated microscope (Molecular Devices, Sunnyvale, CA) with a Plan Fluor 10 × /0.3 objective. To cover a maximal area of the wells, 49 images were acquired for each single well in a DAPI channel. Neural rosettes were quantified using the open source image analysis software Cell Profiler (
Telomerase activity assays
hESCs and hNECs cells were collected at ∼80% confluence and extracted with CHAPS lysis buffer from the TRAPeze®XLTelomerase Detection Kit (Millipore) according to the manufacturer's instructions. Telomerase activity was then measured by real-time polymerase chain reaction (PCR)-based TRAP assay as described previously [18,23]. The PCR products were also analyzed on 10% polyacrylamide gels with SYBR Green staining (Roche, Basel, Switzerland;
Statistical analysis
Quantitative data are presented as the mean ± standard deviation. Statistical differences among groups were evaluated by one-way analysis of variance followed by Tukey's Multiple Comparison Test or Bonferroni post-test. In some cases, data were analyzed by t-test. For all statistical analyses, P value <0.05 was considered significant (*P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001). All statistical analyses were performed with GraphPad Prism version 5.0c for Windows (GraphPad Software;
Results
Normal morphology and self-renewing capacity of undifferentiated hESCs do not depend on HMGB1 and HMGB2
It was previously reported that the expression of HMGB-type proteins is very high in human ICM, in established hESC lines, and in mouse embryos at early stages of development [9,24]. The functional roles of HMGB1 and HMGB2 in the cells of early embryos have remained largely unaddressed. Our initial analyses confirmed that HMGB1 and HMGB2 localize in the nuclei of hESCs and are distributed evenly within colonies of hESCs under standard culture conditions (Fig. 1A). We prepared hESC lines with stable, Dox-inducible shRNAs specifically targeting the HMGB1 and/or HMGB2 gene products. We confirmed that the addition of Dox to the culture media of control (empty vector-transfected) cells did not influence the basal levels of HMGB1 and HMGB2 for at least 12 passages (Fig. 1B). We then selected the clones with the greatest shRNA-mediated inhibition of HMGB1 and/or HMGB2 expression and used them for all subsequent analyses. Compared with those in control cells, the levels of HMGB1 and HMGB2 in the Dox-induced hESCs were reduced by an average of ∼90%–95%, as determined by western blotting (Fig. 1C, D, left panels) and verified by immunostaining of HMGB1 and HMGB2 (Fig. 1C, D, right panels). We demonstrated that the knockdown of both HMGB1 and HMGB2 could be sustained in the hESCs for up to 12 passages in the continuous presence of Dox (Supplementary Fig. S1B).
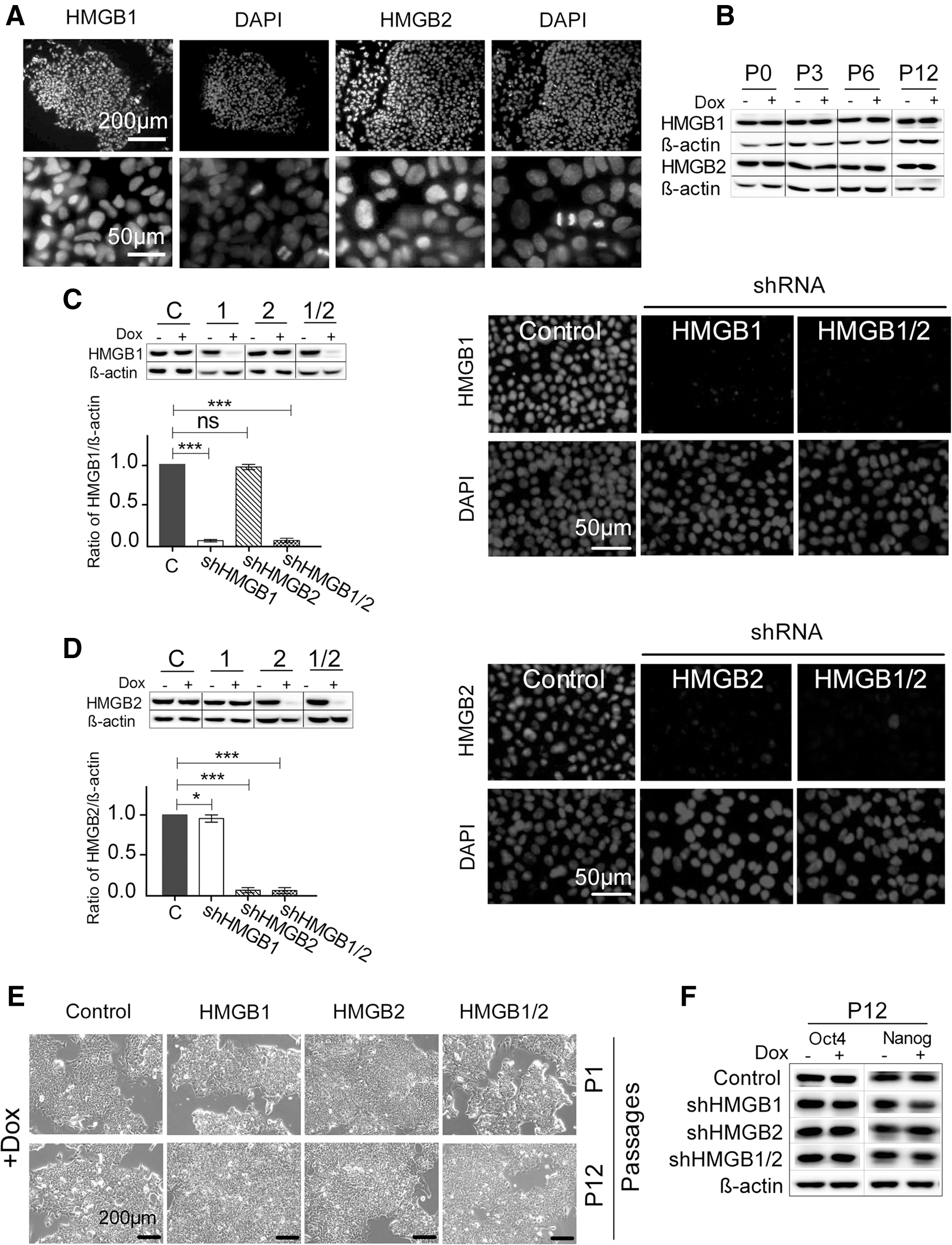
Stably transfected hESCs with downregulated expression of HMGB1 and/or HMGB2 display the normal morphology and self-renewing capacity of undifferentiated hESCs.
The inhibition of HMGB1 and/or HMGB2 expression for 7 days (passage P1) did not alter the morphology of the undifferentiated hESCs (Fig. 1E). Even after 12 consecutive passages in the presence of Dox (passage P12; ∼50 days of culture), the cells displayed no changes in morphology or other signs of differentiation (Fig. 1E). The unchanged expression of the transcription factors OCT4 and NANOG (Fig. 1F) further confirmed that the downregulation of HMGB1 and/or HMGB2 did not compromise the pluripotency of the hESCs.
Downregulation of HMGB1 and HMGB2 induces changes to the proliferation rate and cell-cycle profile of undifferentiated hESCs
Our initial analyses of hESCs with downregulated HMGB1 and HMGB2 expression did not reveal any changes to the cells' undifferentiated characteristics. Undifferentiated hESCs typically display a high rate of proliferation and a specific cell-cycle profile [25]. Therefore, we investigated whether the HMGB silencing influenced those two characteristics in hESCs. In the absence of Dox, the shRNA constructs did not cause any changes to the proliferation rate of the hESCs, as revealed by growth curves (Fig. 2A). Importantly, the differences in an overall proliferation rate among the controls and the cells carrying HMGB shRNA constructs remained marginal (nonsignificant) also upon addition of Dox (Fig. 2A). Subsequent flow cytometric analysis of the cell-cycle profiles of the hESCs showed that compared with empty-vector control cells (Fig. 2B and Supplementary Fig. S2), the cells with downregulated HMGB1 were more likely to be in G1 phase and less likely to be in S phase. In contrast, the hESCs with downregulated HMGB2, and also those with simultaneously downregulated HMGB1 and HMGB2, were less likely to be in G1 phase and more like to be in the S and G2/M phases compared with control cells (Fig. 2B, C). The proliferation index, as calculated from the distribution of cells among the cell-cycle phases, further confirmed changes in the cell division rate (Fig. 2D). Taken together, our results indicate that HMGB1 and/or HMGB2 can modulate the progression of hESC division by an yet-unknown mechanism without compromising the hESC morphology and levels of OCT4 and NANOG, which are associated with the undifferentiated state. Also, the opposing effects of HMGB1 silencing and HMGB2 silencing on the cell cycle might suggest that those proteins play very different roles in the biology of hESCs.
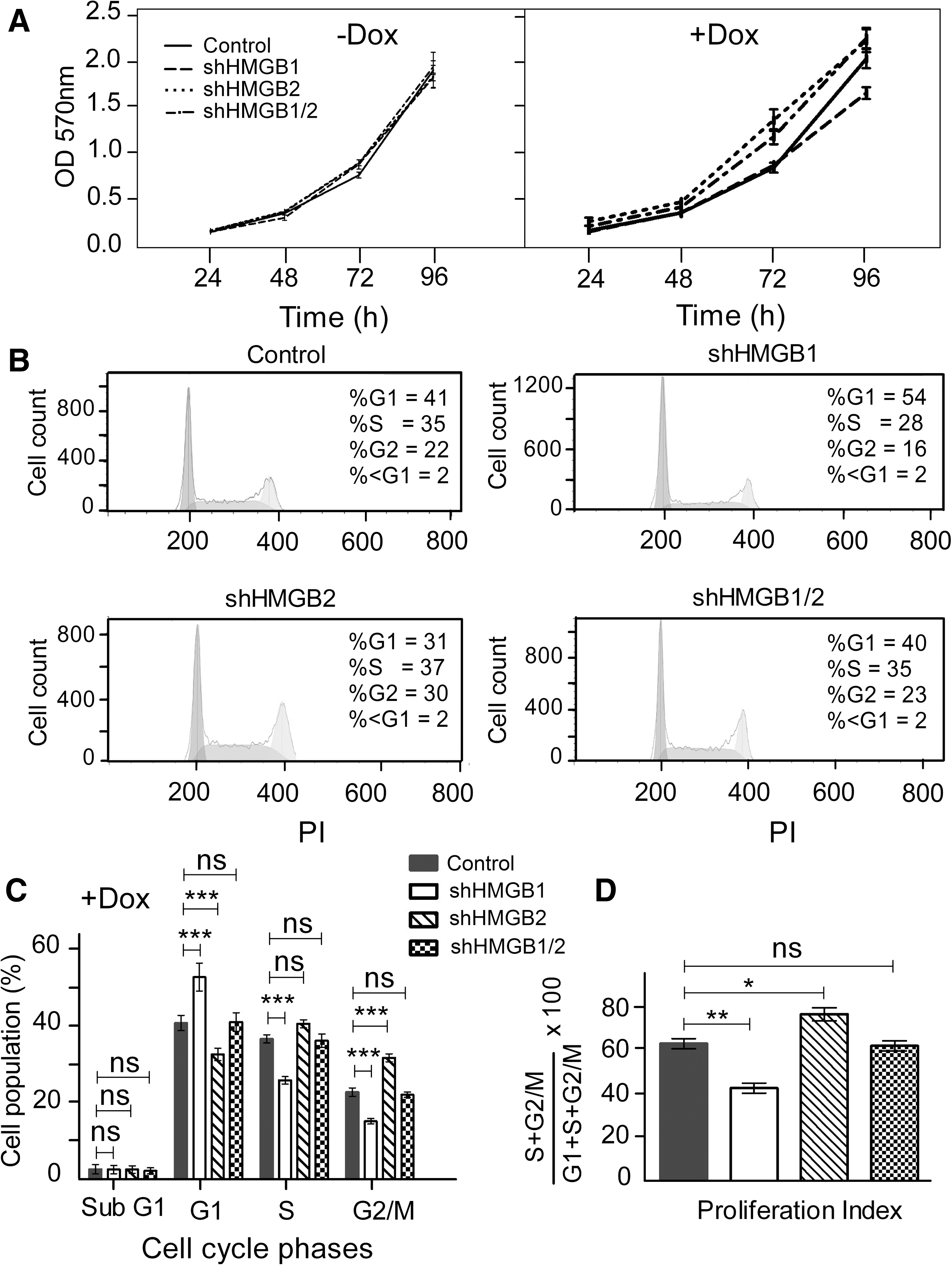
Downregulation of HMGB1 and HMGB2 induces changes to the proliferation rate and cell-cycle profile of undifferentiated hESCs.
Downregulation of HMGB1 and HMGB2 affects hESC proliferation upon induction of differentiation toward the neuroectodermal lineage
As shown in Figs. 1 and 2, the silencing of HMGB1 and/or HMGB2 had only minor, although measurable, effects on undifferentiated hESCs. It was still possible, however, that the HMGB proteins were involved in the onset and/or progression of differentiation rather than in the maintenance of the undifferentiated state. Both HMGB1 and HMGB2 have been implicated in the survival and proliferation of neural progenitor cells [13,26]. Therefore, we explored the effects of HMGB1 and HMGB2 on hESC differentiation toward the neuroectodermal lineage. We took advantage of a protocol established in our laboratory to derive human neuroectodermal cells (hNECs) from hESCs [27] by treating the hESCs with neurodifferentiation media (Supplementary Table S2), which results in the formation of neural rosettes about 8 days after the onset of differentiation. We confirmed the formation of neural rosettes by staining cell nuclei with DAPI (Supplementary Fig. S3). As demonstrated by western blotting (Fig. 3A), by day 5, the cells that were differentiating toward the neural lineage ceased to express OCT4 and instead expressed high levels of PAX6, which was not the case for hESCs cultured for the same amount of time under nondifferentiating conditions. On day 9, the OCT4/PAX6 expression continued to follow the pattern observed on day 5 (Fig. 3A). It is noteworthy that neural differentiation was accompanied by an increase in the level of HMGB2 but not in that of HMGB1 (Fig. 3A). The localization of HMGB1 and HMGB2 remained unchanged during neural differentiation, with both proteins remaining strictly in the nuclei (Fig. 4B).
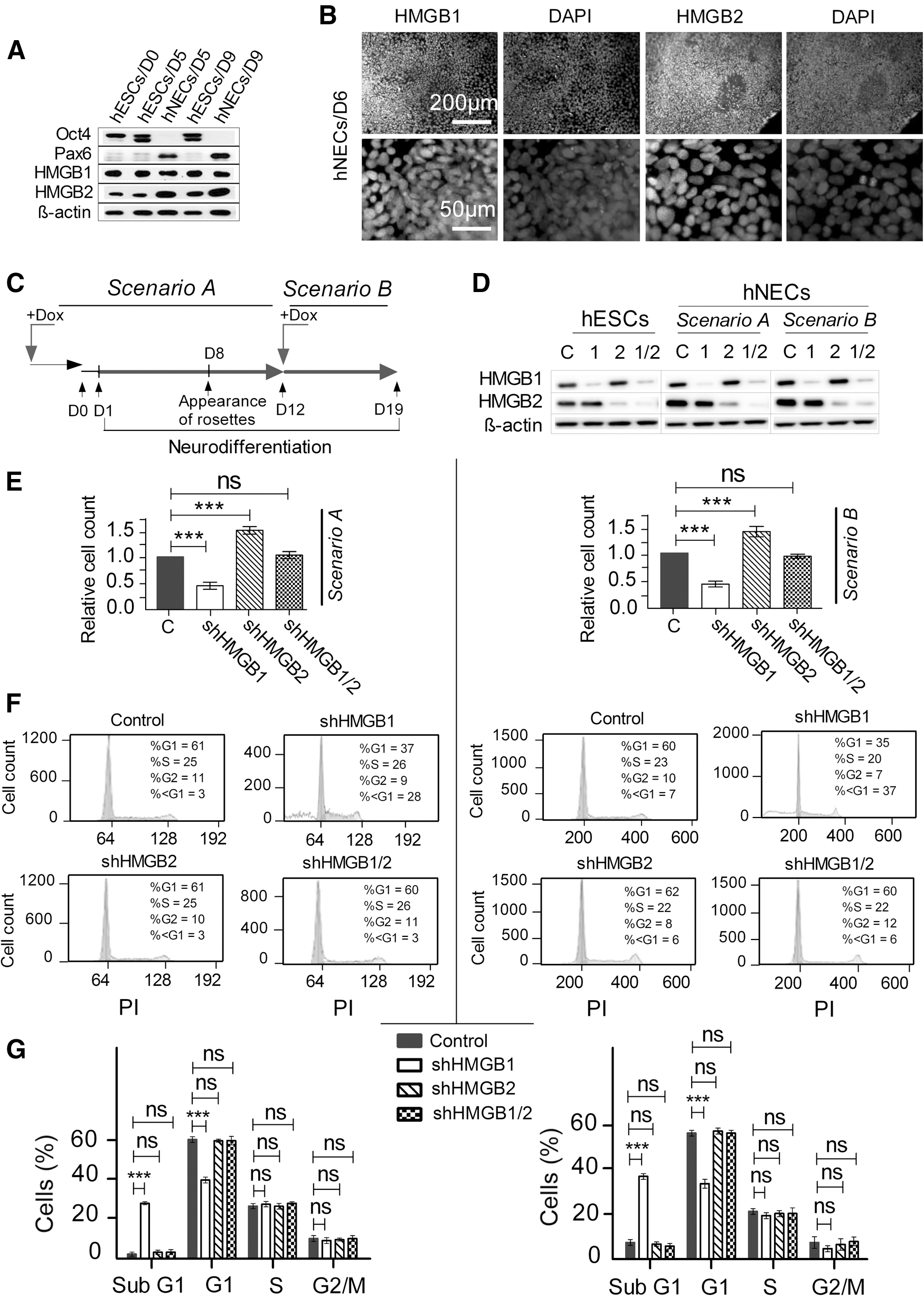
Downregulation of HMGB1 and HMGB2 affects proliferation upon induction of hESC differentiation toward the neuroectodermal lineage.
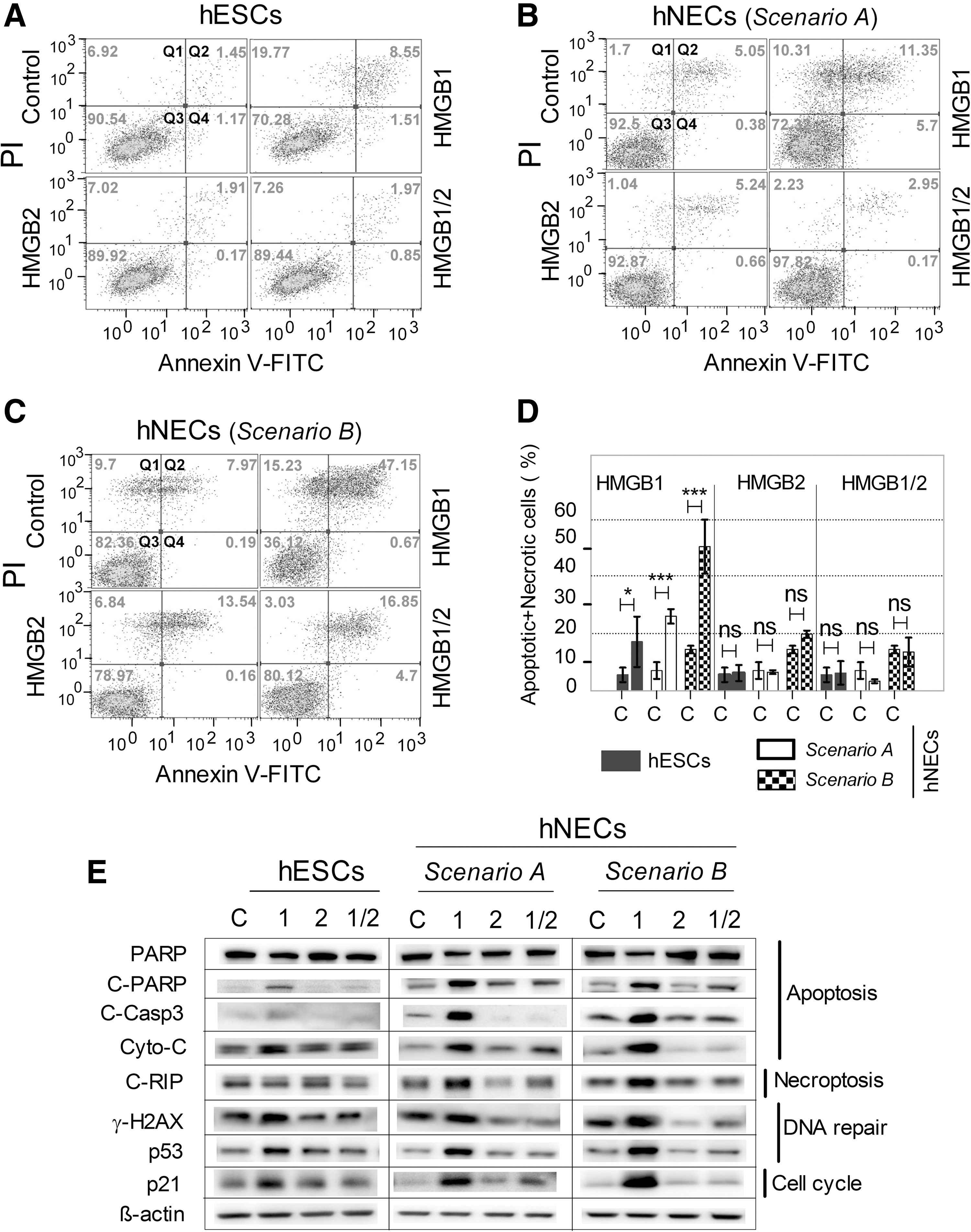
Downregulation of HMGB1 leads to increased apoptosis in hESCs and hNECs.
HMGBs might regulate various stages of neural differentiation, spanning from the initiation and progression of differentiation to the maintenance of the differentiated phenotype. We designed our experiments to evaluate at least those two distinct processes/states. We used two different scenarios for the downregulation of the HMGBs (HMGB1, HMGB2, and HMGB1/2; Fig. 3C). In Scenario A, the HMGBs were continuously downregulated from at least 72 h (3 days) before the beginning of differentiation (D1, the time at which the neurodifferentiation media were added) until day 12 of differentiation. In Scenario B, differentiation began under normal (nonsilenced) conditions, and the HMGBs were downregulated for only 7 days beginning on day 12 of differentiation (Fig. 3C). In all of the differentiation experiments, hESCs with empty vector were used as controls. The cells (hNECs) in Scenarios A and B were analyzed on days 12 and 19, respectively, after the beginning of differentiation. Western blotting and indirect immunofluorescence confirmed that the desired downregulation was achieved in both scenarios for HMGB1, HMGB2, and HMGB1/2 (Fig. 3D). We determined the growth properties of the hNECs with silenced HMGBs in a manner similar to that used for the undifferentiated hESCs. Regardless of the experimental scenario, HMGB1 silencing resulted in very prominent growth inhibition, HMGB2 silencing resulted in growth stimulation, and the combined silencing of both HMGBs had no impact on the growth rate of the hNECs (Fig. 3E). The only notable phenomenon revealed by the flow cytometric analysis of the hNEC cell-cycle phases (Fig. 4A, Scenario A and Fig. 4C, Scenario B) was a dramatic increase of the percentage of sub-G1 population (up to about 35%) at the expense of cells in G1 phase among the hNECs with downregulated HMGB1 (Fig. 4B, D); neither HMGB2 silencing nor HMGB1/2 silencing resulted in such an increase (Fig. 4B, D).
Downregulation of HMGB1 increases apoptosis in hESCs and hNECs
The appearance of the sub-G1 population indicated apoptosis among the hNECs with downregulated HMGB1. Previously, HMGB1 knockdown caused apoptosis in several different cancer cell lines independently of other types of damage (for a review see Tang et al. [28]). Therefore, we determined by flow cytometry the proportions of AnnexinV-positive cells, reflecting the early stage of apoptosis, among populations of hESCs and hNECs. HMGB1 downregulation increased the proportions of apoptosing hESCs and hNECs, whereas HMGB2 or HMGB1/2 downregulation did not (Fig. 4A–C). The proapoptotic effect of HMGB1 downregulation differed among cell categories, being minimal in undifferentiated hESCs (about 20% apoptosing cells), intermediate in hNECs produced according to Scenario A (about 30% apoptosing cells), and maximal in hNECs produced according to Scenario B (about 50% apoptosing cells; Fig. 4D). In consonance with those findings, the HMGB1-knockdown cells showed many other molecular signs that are functionally linked with cell damage and dying (Fig. 4E). Specifically, they showed (1) increased cleavage of PARP and Caspase 3, (2) increased levels of Cytochome-C and the pronecroptotic protein RIP, (3) increased phosphorylation of γ-H2AX (DNA damage marker), and (4) accumulation of p53 and its target p21 (negative regulator of the cell cycle).
Altogether, our data indicate that decreased HMGB1 expression leads to apoptosis in both hESCs and hNECs and that the effect is more profound in differentiated cells. Furthermore, the apoptotic effect develops only in cells that express HMGB2; in the absence of both HMGB1 and HMGB2, differentiated hNECs maintain the properties of control cells.
Downregulation of HMGB1 and HMGB2 affects hESC differentiation
Previously published findings suggest that HMGBs could exercise some role in differentiation processes that take place early in development. We asked more specifically whether or not HMGB1 and HMGB2 are important for the establishment and/or maintenance of the neural phenotype from hESCs. The critical step in the differentiation of hESCs toward the neural lineage is the formation of neural rosettes. We therefore used the number of neural rosettes as a measure of differentiation efficiency. As shown in Fig. 5A, HMGB1 downregulation during differentiation (Scenario A) dramatically decreased the number of neural rosettes (by about 70%), whereas HMGB2 downregulation during differentiation slightly, but still significantly, increased the number of neural rosettes (by about 25%). The simultaneous downregulation of both HMGB1 and HMGB2 during differentiation had no measurable effect on neural rosette formation. When we downregulated the HMGBs later in neural differentiation (Scenario B), the only significant outcome was a decrease in the number of neural rosettes (by about 25%) in cultures with downregulated HMGB1 only (Fig. 5B). We also determined by western blotting the expression of transcription factors associated with pluripotency and differentiation (Fig. 5C). In hNECs, the levels of OCT4 and NANOG, both markers of pluripotency, were at or below the detection limit, irrespective of the HMGB status and the downregulation scenario (A or B). The behavior of the differentiation-associated proteins SOX2 and PAX6 was somewhat peculiar. The levels of Sox2 were slightly but consistently lowered in hNECs with any HMGB downregulation in Scenario A and in those with HMGB1 downregulation in Scenario B. PAX6, which was not detectable in undifferentiated hESCs, was detected in hNECs in a pattern similar to that of Sox2.
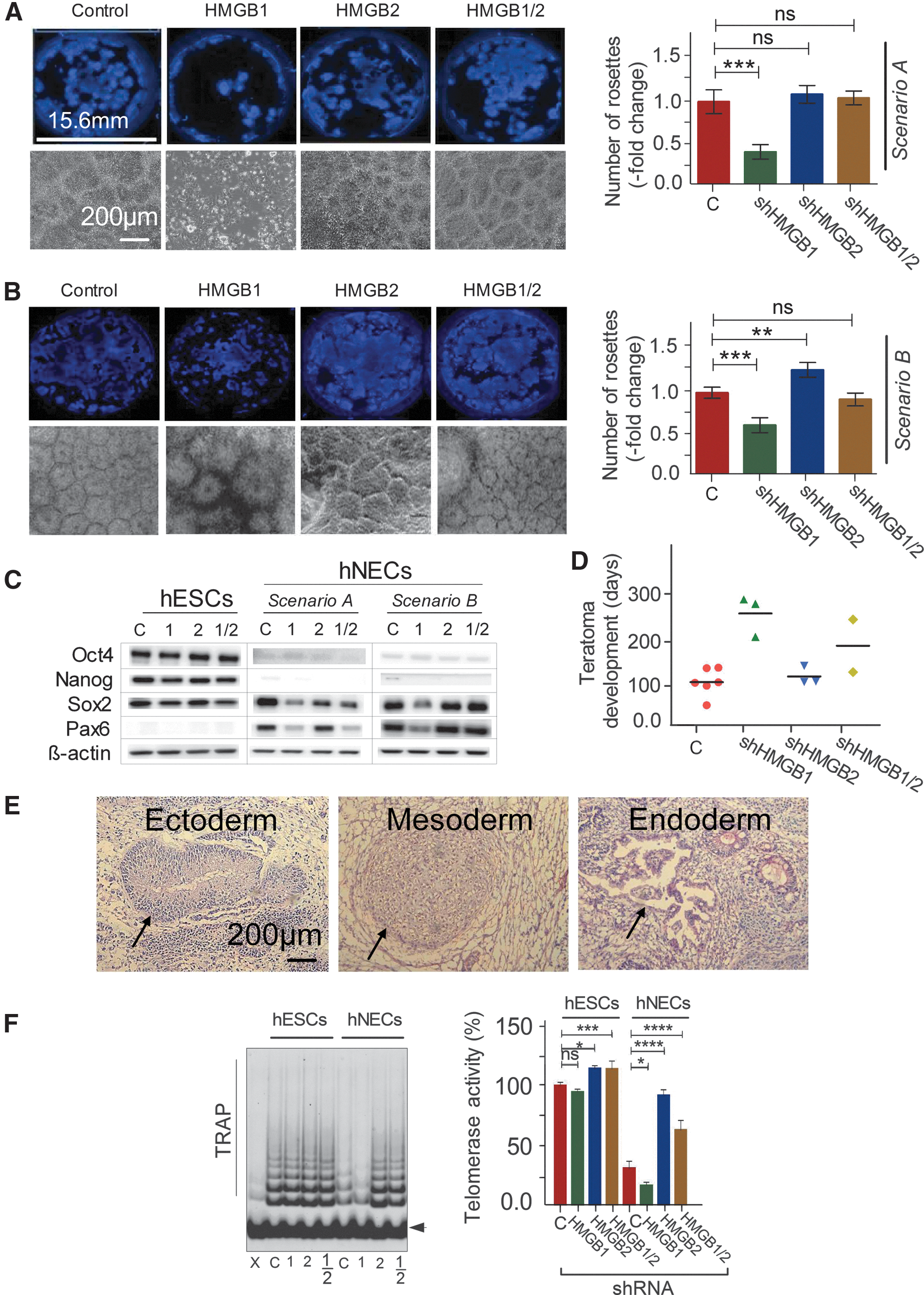
Downregulation of HMGB1 and HMGB2 affects the differentiation of hESCs and telomerase activity.
Our findings clearly indicated that HMGB1/2 expression influences some aspects of hESC differentiation. To assess those influences in more natural conditions, we subjected normal hESCs and hESCs with downregulated HMGB1 and/or HMGB2 expression to a teratoma formation assay in mice. We injected each of the following cells types into a separate group of five mice: (1) unmodified normal hESCs, (2) hESCs with empty vector, (3) hESCs with HMGB1 shRNA, (4) hESCs with HMGB2 shRNA, and (5) hESCs with HMGB1 and HMGB2 shRNA. The first two cell types collectively represented unmodified controls. Some animals died during the experiment and/or did not develop teratoma (Supplementary Table S4). We used the number of days needed for palpable (about 1-cm3) teratomas to develop in the surviving animals as a measure of teratogenesis efficiency. The control hESCs and those with downregulated HMGB2, and to some extent those with downregulated HMGB1/2, formed palpable teratomas early after injection (2–3 months; Fig. 5D). In strong contrast, the hESCs with downregulated HMGB1 took much longer to form palpable teratomas (6–8 months). Histological examination did not reveal any tangible deviation from the normally differentiated teratoma morphology in any of the teratomas. Taken together, those results indicate that although HMGBs, and HMGB1 in particular, are required for development to proceed at a normal tempo, they are not decisive for the ability to give rise to cell phenotypes belonging to the three germ lineages. Examples of derived ectoderm, mesoderm, and endoderm in the teratomas are shown in Fig. 5E.
Pluripotent stem cells have many specific aspects of genome organization and regulation, including high telomerase activity [29]. HMGB1 and HMGB2 are known to contribute to the normal regulation of telomerase in somatic cells and cancer cells [18,30]. Therefore, we assessed whether the same is true for hESCs and their differentiated derivatives using a TRAP assay analyzed either by a gel electrophoresis (Fig. 5F, left panel) or quantified by a real-time PCR (Fig. 5F, right panel). Telomerase activity was high in undifferentiated hESCs and was about 2.5-fold lower in differentiated hNECs (Fig. 5F). The effects of HMGBs downregulation differed between the undifferentiated hESCs and the differentiated hNECs. In undifferentiated hESCs, HMGB1 downregulation had no significant effect; however, HMGB2 downregulation, and HMGB1/2 downregulation, caused a small (about 30%) but significant increase in telomerase activity. In differentiated hNECs, the effects were much more dramatic. HMGB1 downregulation caused a drop in telomerase activity to about half the level in control cells. In contrast, the downregulation of HMGB2 and HMGB1/2 resulted in a dramatic increase in telomerase activity of about 2.5- and 2-fold, respectively. The downregulation of HMGB2 in hNECs brought the telomerase activity back to the level typical for undifferentiated hESCs.
Discussion
An understanding of the molecular mechanisms that regulate pluripotency has direct implications on developmental biology, regenerative medicine, epigenetics, and cancer research. Our findings directly link the expression of the nonhistone, DNA-binding proteins HMGB1 and HMGB2 to the biology of hESCs and their differentiated derivatives.
All our experiments were performed on stably transfected hESCs with Dox-inducible shRNA-inhibited HMGB expression. We have tested a number of sequences for silencing of HMGB1 or HMGB2 genes in several human cell lines and selected for our experiments only those with the highest efficiency of HMGB knockdown, and at the same time, exhibiting similar cellular outcomes (such as in apoptosis, response to anticancer drugs and telomerase activity) upon HMGB KD ([16,17] and unpublished data). We have also confirmed that the expression of housekeeping genes (such as glyceraldehyde-3-phosphate dehydrogenase, β-actin, and 18S rRNA or 28S rRNA) was not affected by HMGB silencing (unpublished data). Neither HMGB1 KD nor HMGB2 KD affected the expression of the HMGB2 or HMGB1 genes, respectively (Fig. 1). Thus, similar phenotypes observed with different shRNA sequences for the silencing of a given HMGB gene (HMGB1 or HMGB2) provided a strong evidence that the results of our shRNA experiments were genuine and did not originate from off target effects. Our data show that the morphology and self-renewing capacity of pluripotent hESCs were not impaired even after prolonged downregulation of HMGB1 and/or HMGB2. The expression of OCT4 and NANOG also remained unchanged, and we detected only negligible changes in the rate of proliferation, cell-cycle profile and the induction of apoptosis in hESCs with downregulated HMGB1. Upon induction of differentiation, the downregulation of the HMGB proteins manifests much more strongly, affecting cell proliferation, apoptosis, telomerase activity, and the efficiency of differentiation toward the neuroectodermal lineage. Furthermore, all of those effects (in most cases opposite) appear to manifest only with the downregulation of one of the two HMGB proteins, as the downregulation of both HMGB1 and HMGB2 results in a normal phenotype.
The fact that the previously reported single-knockout HMGB1−/− or HMGB2−/− pups were born viable suggested that HMGB proteins might be, to at least some extent, redundant [7,8]. However, it is also likely that other members of the HMGB superfamily (HMGB3 and HMGB4) might to some extent substitute for a lack of HMGB1 and HMGB2, although we did not see the upregulation of those two proteins upon HMGB1/2 knockout in any of the cells (hESCs or hNECs) that we analyzed (data not shown). To date, only one study has shown a direct interaction of HMGB2 with CDK2 in undifferentiated hESCs [12]. Because CDK2 is the major driver of proliferation in undifferentiated hESCs [12], further experiments are necessary to establish whether HMGB2 could act as a direct inhibitor of CDK2.
Our data indicate that HMGB1 and HMGB2 might contribute to the regulation of the onset of differentiation in hESCs rather than to the maintenance of the undifferentiated state. Previously, both HMGB1 and HMGB2 were shown to be associated with the survival and proliferation of mouse neural stem cells (NSCs) [13,26]; however, there was no information about the human analogs of those cells. Our Dox-inducible system enabled us to demonstrate that HMGB1 and HMGB2 are important for hESC differentiation toward the neuroectodermal lineage. We specifically showed that the downregulation of HMGB1 during differentiation could decrease cell proliferation and induce apoptosis, while the downregulation of HMGB2 had no effect on apoptosis and significantly increased cell proliferation. Those findings correspond very well to previously reported data from rodent models. Wang and colleagues [26] showed that HMGB1 could enhance mouse embryonic NSC proliferation by activating the MAPK signaling pathway. Li and colleagues [31] showed that HMGB1 upregulation significantly increased the proliferation of cultured primary rat NSCs. Finally, Abraham et al. [13] reported that mice lacking HMGB2 showed increased NCS proliferation and also adult neurogenesis. Our results extend that knowledge by showing that human neuroectodermal cells that differentiated in vitro from hESCs used analogous molecules to modulate their proliferation.
We found that the progression of hESC differentiation, both in vitro and in vivo, was considerably influenced by the inadequate expression of HMGB1 and HMGB2. While hESCs with downregulated HMGB1 developed markedly fewer neural rosettes in vitro and dramatically delayed teratoma development in vivo, hESCs with downregulated HMGB2 displayed significantly enhanced neural rosette formation in vitro and teratogenesis efficiency in vivo. Those properties might have a functional link with the findings published by Abraham et al. [13], who showed that HMGB2−/− mice display increased neurogenesis in vivo. It has also been reported that HMGB1 might increase neurite outgrowth and cell survival in neurons [32,33]. Therefore, its absence could result in decreased neurodifferentiation.
Finally, we showed that both HMGB1 and HMGB2 are required for a normal level of telomerase activity. Telomerase activity has been correlated with the aggressiveness of tumor development in many cancers (reviewed in Zisuh et al. [34]) and in hESC-derived teratomas [35]. We found that teratoma development is affected by HMGB expression and hypothesized that this could be linked to telomerase activity. In undifferentiated hESCs, the downregulation of HMGB1 and HMGB2 affected the telomerase activity very little. In contrast, the downregulation of HMGB1 in hNECs caused a marked decrease in telomerase activity, indirectly supporting our findings of delayed tumor formation in HMGB1-knockdown hESCs.
The downregulation of HMGB2 in hNECs induced a very dramatic upregulation of telomerase activity to levels about the same as those in undifferentiated hESCs. That finding is in consonance with our previous observation of increased telomerase activity in mouse embryonic fibroblasts derived from HMGB2−/− mice [18]. Whether HMGB1 deficiency could also induce telomere shortening and dysfunction in human NECs, as previously shown in HMGB1−/− mouse embryonic fibroblasts [18], remains to be established.
Conclusions
Results of this article directly link the expression of the chromosomal DNA-binding proteins HMGB1 and HMGB2 to the biology of hESCs and their differentiated derivatives. HMGB1 and HMGB2 proteins appear to be dispensable for hESC self-renewal but influence the onset of hESC differentiation both in vitro and in vivo. Our data reveal that HMGB1 and HMGB2 proteins play important, and likely opposing, modulatory roles during early human development.
Footnotes
Acknowledgments
The authors would like to thank Jan Verner and František Dráfi (Department of Histology and Embryology, Masaryk University) for help with the animal experiments; Bořivoj Vojtěšek (Masaryk Memorial Cancer Institute, Brno, Czech Republic) for providing the anti-p53 antibody; Klára Koudelková (Department of Histology and Embryology, Masaryk University) for technical support; and Lukáš Čajánek, Josef Jaroš, and Ondřej Urbanovský (Department of Histology and Embryology, Masaryk University) for helpful comments and suggestions. This research was supported by grants from the Grant Agency of the Czech Republic (P305/12/2475, P305-15-01354S, GJ15-18316Y, GJ16-24004Y, and GA15-23033S), by funds from the Ministry of Education, Youth, and Sport of the Czech Republic (MUNI/A/1352/2015), and by the Project No. LQ1605 (MEYES CR, NPU II).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.