
Abstract
An acidic environment is vital for the maintenance of cellular activities but can be affected tremendously during intervertebral disc degeneration (IVDD). The effect of changes in the acidity of the environment on human nucleus pulposus mesenchymal stem cells (NP-MSCs) is, however, unknown. Thus, this study aimed to observe the biological effects of acidic conditions mimicking a degenerated intervertebral disc on NP-MSCs in vitro. NP-MSCs were isolated from patients with lumbar disc herniation and were further identified by their immunophenotypes and multilineage differentiation. Then, cells were cultured at acidic pH levels (pH 6.2, pH 6.5, pH 6.8, pH 7.1, and pH 7.4) with/without amiloride, an acid-sensing ion channel (ASIC) blocker. The proliferation and apoptosis of NP-MSCs and the expression of stem cell-related genes (Oct4, Nanog, Jagged, Notch1), ASICs, and functional genes (Aggrecan, SOX-9, Collagen-I, and Collagen-II) in NP-MSCs were evaluated. Our work showed that cells obtained from human degenerated NP met the criteria of International Society for Cellular Therapy. Therefore, cells obtained from a degenerated nucleus pulposus were definitively identified as NP-MSCs. Our results also indicated that acidic conditions could significantly inhibit cell proliferation and increase cell apoptosis. Gene expression results demonstrated that acidic conditions could decrease the expression of stem cell-related genes and inhibit extracellular matrix synthesis, whereas it could increase the expression of ASICs. Our study further verified that the above-mentioned biological activities of NP-MSCs could be significantly improved by amiloride. Therefore, the results of the study indicated that the biological behavior of NP-MSCs could be inhibited by acidic conditions during IVDD, and amiloride may meliorate IVDD by improving the activities of NP-MSCs.
Introduction
I
Stem cells have shown an ability to replace nucleus pulposus (NP) cells and restore the function and structure of degenerated discs [4]. Previous studies have shown that IVD tissue is nonrenewable [5]. However, recent studies confirmed that normal and degenerated IVDs also contain nucleus pulposus mesenchymal stem cells (NP-MSCs), which could differentiate into nucleus pulposus-like cells [6,7]. Previous studies indicated that endogenous stem cells played an important role in maintaining the homeostasis when damage occurred to body tissues [8]. Thus, the administration of NP-MSCs in degenerative IVD may provide a new cell therapy for IVD degeneration. However, these NP-MSCs could not differentiate into IVD cells, thereby maintaining the stability of IVD or even reversing the degenerative process.
The IVD is the largest avascular tissue in the body and it receives all essential nutrients, such as oxygen and glucose, through the cartilage endplate [9]. During aging and degeneration of the IVD, especially calcification of the end plate, the flow of nutrients and metabolites is reduced, leading to an accumulation of lactic acid in the disc, which causes an acidic microenvironment in the IVD [10]. Previous studies showed that the pH value in a normal IVD was 7.2 to 7.4, while the pH was reduced to 6.2–6.8 in degenerated IVDs [11].
A normal pH is necessary to maintain normal cell functioning, whereas an excessively low pH induces apoptosis of cells and destruction of the ECM structure. There have been investigations regarding whether an acidic environment is able to change the properties of human NP cells and reported that detrimental effects of low pH on NP cells include a reduction in cell viability, cell proliferation, and aggrecan expression, suggesting a shift in matrix metabolism [12]. Other studies have also found that an acidic environment has a significant negative effect on the function and metabolism of IVD cells [13,14]. In addition, the expression of related genes (Aggrecan, Collagen-II, and Collagen-I) by mesenchymal cells derived from the bone marrow of rats [15,16], human adipose tissue, and rat IVD [17] cultured in an acidic environment have been investigated.
Interestingly, all studies demonstrated a decrease in the expression of matrix-associated genes following exposure to an acidic pH. Wang et al. [18] concluded that an excessively acidic state could restrict the proliferation and differentiation of stem cells. NP-MSCs were similar to bone marrow-derived stem cells (BMSCs), and they all have similar morphological characteristics, cell phenotypes, and multipotential differentiation. Thus, we hypothesized that an acidic environment in a degenerative IVD may alter the properties of NP-MSCs. Furthermore, the mechanisms by which an acidic extracellular environment regulates the properties of NP-MSCs have not been fully understood.
In recent years, there has been progress toward elucidating the cellular sensory mechanisms by which cells respond to changes in extracellular pH. Membrane proteins called acid-sensing ion channels (ASICs) are major extracellular receptors that respond to acidic pH [19]. ASICs are proton-gated ion channels that are activated by acidosis, lactate, and arachidonic acid and are located in the nervous system [20] and other cells such as bone and disc cells [21]. ASICs are members of the DEG/ENaC superfamily of amiloride-sensitive ion channels [22]. They play an important role in pathophysiological processes in many tissues [23].
Previous studies have indicated that pH regulates the activity of cells mainly through ASICs, including ASIC1, ASIC2, ASIC3, and ASIC4, in nervous and joint tissues [24,25]. Hu et al. [26] found that acidic environments inhibited proliferation and increased apoptosis of chondrocytes through ASICs, while the use of amiloride to block these channels relieved the damage to chondrocytes caused by the acidic pH. Li et al. [27] demonstrated that acid-induced apoptosis, induced through ASICs, occurred in endplate chondrocytes and blockade of ASICs by psalmotoxin 1 (PcTx1) or specific small interfering RNA (siRNA) restrained such acid-induced apoptosis in the endplate chondrocytes of IVDs. These results provided evidence that the correlation between acidic pH and ASIC functioning may contribute to IVD degeneration.
In our study, we speculated that acidic environment inhibits the biological activities of human NP-MSCs and may execute this inhibiting effect through ASICs. Thus, we isolated NP-MSCs from patients undergoing IVD surgery and, using flow cytometry and multipotential differentiation, determined whether this type of cell fulfills the criteria of MSCs established by an expert panel from the International Society for Cellular Therapy (ISCT). In addition, we examined the effects of extracellular acidic environments on the biological properties of NP-MSCs in vitro, including cell proliferation, apoptosis, and expression of stem cell-related genes and functional genes (SOX-9, Aggrecan, Collagen-I, and Collagen-II), and investigated the role of ASICs in producing these effects. Furthermore, we assessed the effect of blocking ASICs with amiloride on the biological activities of human NP-MSCs cultured at different acidic pH levels (Fig. 1). We hypothesized that ASIC activation by acidic extracellular conditions play a critical role in NP-MSC survival.
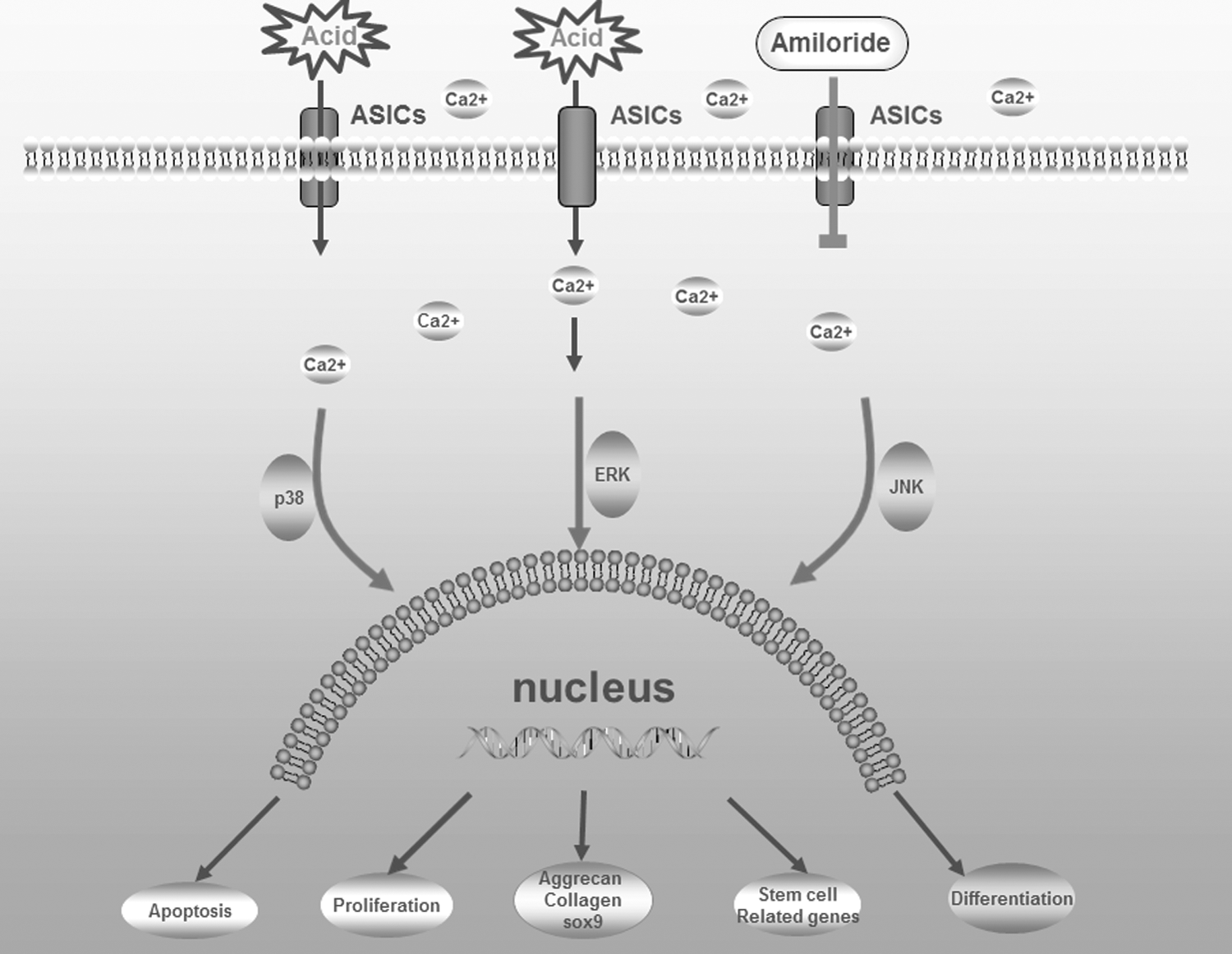
A schematic image of an overview of the experimental design. ASIC, acid-sensing ion channel.
Materials and Methods
Cell isolation and culture
Cells were isolated from patients undergoing spine fusion for degenerative spine diseases. Table 1 provides details about the subjects' ages, genders, and degrees of disc degeneration. All procedures performed for this work were approved by the local ethics committee of our institution in accordance with the Declaration of Helsinki. Specific informed consent was obtained in all cases. Extreme care was taken to prepare uncontaminated NP tissue. First, the extraneous ligaments were cleaned from intact samples using a dissecting microscope. Second, the avascular NP tissue was carefully isolated from the AF. NP tissue from degenerative IVDs was then mechanically fragmented and subjected to enzymatic digestion with 0.02 mg/mL collagenase type II (Gibco) at 37°C in 5% CO2 overnight.
The liberated cells, along with the partially digested tissue, were cultured as an explant in a standard MSC expansion medium, consisting of Dulbecco's modified Eagle's medium-low glucose (HyClone), 10% fetal calf serum (Gibco), and 1% penicillin/streptomycin (Gibco) in a humidified incubator at 37°C in 5% CO2. Suspended cells and fragments were removed from the medium after 24 h, and growth of adherent cells was encouraged by complete replacement of the medium every 2 to 3 days. When cells reached 80%–90% confluence, cultures were passaged at a ratio of 1:3. Cells in passage 2 were harvested with trypsin–EDTA (Gibco) for testing.
Immunophenotype of NP-MSCs
After the NP-MSCs were digested with 0.25% trypsin (Biosharp), they were washed and resuspended in phosphate-buffered saline (PBS; Sigma) and incubated with the following monoclonal antibodies in accordance with the recommendations of the ISCT: CD34-APC, CD73-FTTC, CD45-PE, CD90-FTTC, CD105-PE, and HLA-DR-APC (eBioscience). Each tube contained 100 μL. Isotype control (eBioscience) was used in each case. After incubating with antibodies at room temperature for 30 min, cells were washed in PBS and the supernatant discarded. Cells were resuspended in 500 μL of PBS with 1% paraformaldehyde (Sangon Biotech, China) and examined using flow cytometry (BD). Immunophenotypic analysis was performed to identify the percentage of positive cells and the fluorescence intensity.
Osteogenic differentiation
NP-MSCs in passage 2 were plated at 2 × 104 cells/cm2 in six-well plates. Once 60–70% confluence was achieved, the normal culture medium was then replaced with an osteogenic differentiation medium (Cyagen). The differentiation process lasted for 14 days, and the medium was changed every 2 to 3 days. The control group was cultured with the normal culture medium for 14 days. After differentiation, the medium was removed and cells were rinsed with PBS, followed by fixation with 4% paraformaldehyde solution (Sangon Biotech) at room temperature for 30 min. Then, after washing with PBS twice, the cells were stained with alizarin red working solution (Cyagen) for 15 min. Finally, the cultures were washed with PBS thrice and visualized under a light microscope [28].
Adipogenic differentiation
NP-MSCs in passage 2 were plated at 2 × 104 cells/cm2 in six-well plates. When cells grew to 100% confluence, the normal culture medium was replaced with an adipogenic differentiation medium A (Cyagen). After 3 days, the differentiation medium A was replaced with an adipogenic differentiation medium B (Cyagen) for 24 h. Then, the medium B was replaced with medium A again. This cycle was repeated for three to four cycles for a total of 28 days. The control group was cultured with the normal culture medium for 28 days. After differentiation, the medium was removed from the wells, and the cells were washed with PBS and fixed with 4% paraformaldehyde solution for 30 min. Then, after washing with PBS twice, the cells were stained with oil red O working solution (Cyagen) for 30 min at room temperature. The cultures were then washed with PBS thrice and visualized under a light microscope.
Chondrogenic differentiation
NP-MSCs in passage 2 were resuspended in a chondrogenic differentiation basal medium (Cyagen), then cells were centrifuged at 1500 r/min for 5 min, and the supernatant was discarded. The cells were resuspended in a chondrogenic differentiation complete medium (Cyagen) to a concentration of 5 × 105 cells/mL. Next, 500 μL of cell suspension liquid was placed in a 15-mL polypropylene culture tube containing 2.5 × 105 cells and centrifuged at speed of 1500 r/min for 5 min to form a pellet. Without being disturbed, the pellet was cultured at 37°C in 5% CO2 in the chondrogenic differentiation complete medium. After 3 days, the medium was replaced, and each tube was manipulated to ensure that the pellet was free floating. Thereafter, the differentiation medium was replaced every 2 days, and the differentiation process lasted for 28 days. The control group was cultured with the normal culture medium for 28 days.
When each pellet's diameter had increased to 3 mm, the pellets were fixed by formalin (Sangon Biotech, China) in paraffin sections. Finally, the paraffin sections were deparaffinized and hydrated with distilled water. Then, the sections were stained with Alcian blue (Cyagen) for 30 min. After that, the sections were washed with running tap water for 2 min and rinsed with distilled water several times. Then, the sections were visualized under a light microscope [29].
Preparation of culture media with different pH levels
Culture media with different pH levels were prepared by adding an appropriate amount of sterilized HCl (1 M) and NaOH (1 M) into the media and monitoring the pH using a commercial pH microelectrode (Lazarlab). Media with five pH levels, namely, 7.4, 7.1, 6.8, 6.5, and 6.2, were obtained. The culture media were kept at 37°C in 5% CO2 for 3 days to allow pH equilibrium (CO2 dependent) to be established.
Cell counting kit-8 assay
A cell counting kit-8 (CCK-8) contains a highly water-soluble tetrazolium salt called WST-8, which is reduced by dehydrogenases in cells to give an orange-colored product (formazan), which is soluble in the tissue culture medium. The amount of the formazan dye generated by dehydrogenases in cells is directly proportional to the number of living cells.
In our study, NP-MSCs in passage 2 were plated at 5 × 104 cells/mL in 96-well plates. These cells were cultured at five pH levels at 37°C in 5% CO2. To observe the effect of amiloride (Sigma) on proliferation, experimental groups also contained 100 μM amiloride. Three well subsamples were collected on days 1, 3, 5, 7, 9, and 11, and 10 μL CCK-8 (DOJINDO, Japan) was added to the cells, which were then incubated for 4 h. The isotype group did not contain cells. A spectra MAX microplate reader was used to measure each group's light absorbance.
Apoptosis assay of NP-MSCs
NP-MSCs in passage 2 were plated at 1 × 105 cells/mL in six-well plates. These cells were cultured at five pH levels at 37°C in 5% CO2 for 2 days. To observe the effect of amiloride on apoptosis, experimental groups also contained 100 μM amiloride. Then, adherent cells were collected by digestion of trypsin without EDTA (Biosharp) and were washed with PBS twice. Five microliters of Annexin V-FITC and 5 μL of PI (KeyGEN BioTECH, China) were added to each group and incubated for 5 min. Flow cytometry (BD) was used to detect the percentage of apoptotic cells.
Stem cell-related genes, SOX-9, Aggrecan, Collagen-I, Collagen-II, and ASICs
NP-MSCs in passage 2 were plated at 1 × 105 cells/mL in 6-well plates. These cells were cultured at the five aforementioned pH levels at 37°C in 5% CO2 for 2 days (stem cell-related genes and ASICs) and for 7 days and 14 days (SOX-9, Aggrecan, Collagen-I, and Collagen-II). To observe the effect of amiloride on gene expression, experimental groups also contained 100 μM amiloride. Total RNA was extracted using Trizol Reagent (Ambion) according to the product manual, and RNA samples were treated with DNase-/RNase-free water. RNA quality and quantity were examined using a Nanodrop ND-1000 spectrophotometer (Nanodrop Technologies).
Then, the cDNA of total RNA was obtained using a reverse transcribed reagent (Takara, Japan). One microliter of RNA was mixed with 2 μL of 5 × PrimeScript RT® MasterMix and 10 μL of RNase-free dH2O. The mixed solutions were then incubated at 85°C for 5 s after 15 min at 37°C and were stored at −80°C for quantitative real-time polymerase chain reaction (qRT-PCR). All genes were analyzed by qRT-PCR, and the glyceraldehyde-3-phosphate dehydrogenase housekeeping gene (GAPDH) was used as a control. A SYBR Premix Ex Taq PCR kit (Takara, Japan) and LightCycler (Roche, Switzerland) were used for the qRT-PCR analysis. Primers of all genes were designed using the software Premier 5.0, as shown in Table 2.
Statistical analysis
Experiments and analysis were performed separately at least thrice by different authors, and all data were expressed as the mean ± SD. SPSS version 16.0 software (IBM) was used for statistical analysis. One-way analysis of variance (ANOVA) and univariate general linear analysis were performed to assess the effects of pH levels. P < 0.05 was considered statistically significant.
Results
NP-MSCs were identified by immunophenotypic characterization and multipotential differentiation in vitro
NP-MSCs from degenerated IVDs were isolated. A layer of primary NP-MSCs was observed after 3–5 days of initial cell culture (Fig. 2A). The cultured cells presented the short spindle shape in the first generation. The time to reach 60%–70% confluence was approximately 2 weeks. When cultures were passaged, the cells grew significantly faster than primary cells and in a spiral formation. The morphology of the cells in passage 2 was fairly consistent, presenting the characteristic spindle shape in culture (Fig. 2B).
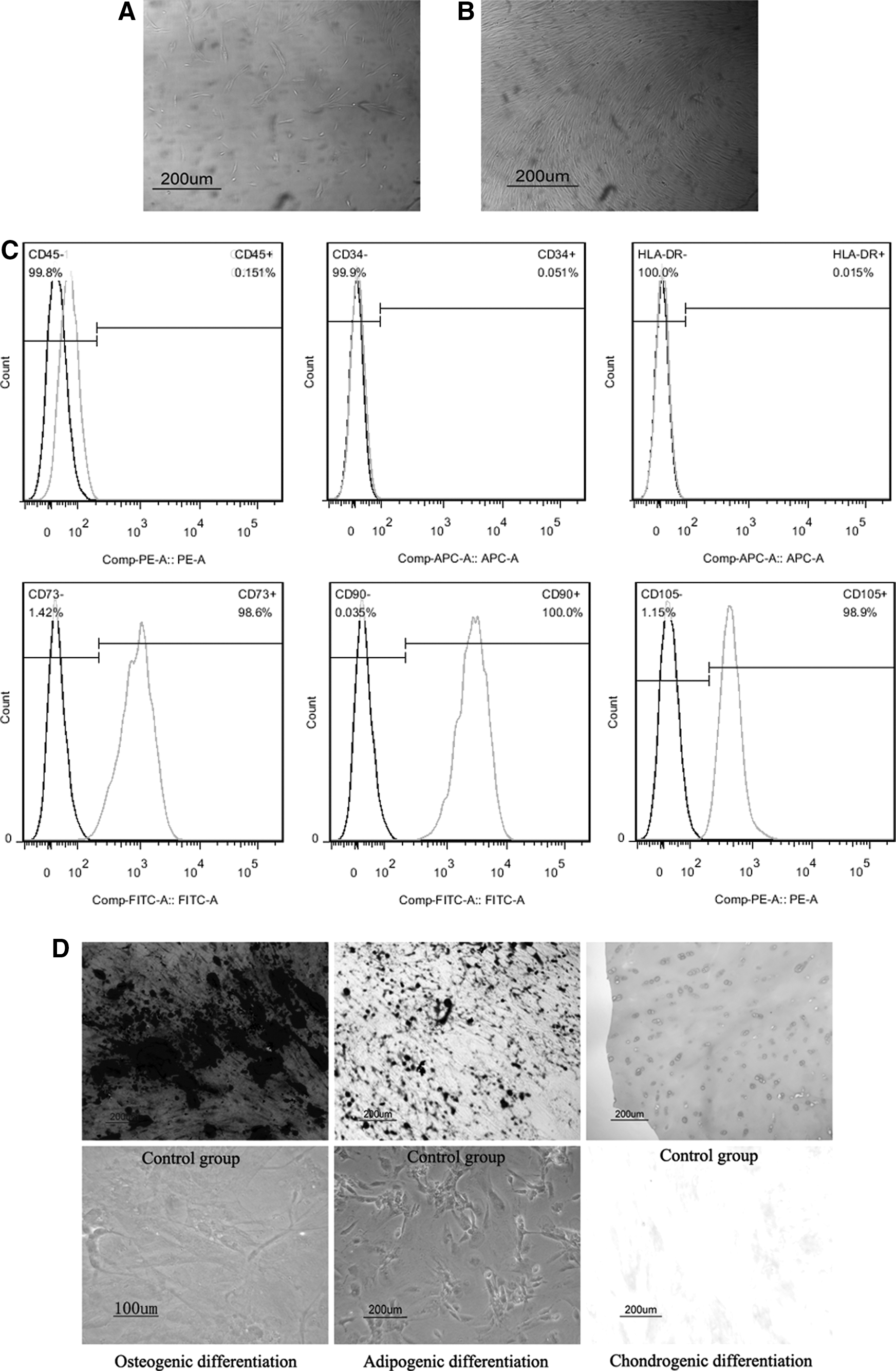
Characteristics of NP-MSCs from NP tissue.
NP-MSCs from degenerative IVD highly expressed CD73, CD90, and CD105 and were negative for CD34, CD45, and HLA-DR (Fig. 2C). For osteogenic differentiation, NP-MSCs appeared to form intensely stained mineralized nodules. NP-MSCs exhibited intracellular lipid vacuoles with oil red O staining after 28 days of adipogenic differentiation. For chondrogenic differentiation, the histological observation demonstrated that NP-MSCs exhibited intense staining of sulfated proteoglycan (Fig. 2D).
Acidic conditions inhibited the proliferation of NP-MSCs, while amiloride relieved this inhibiting effect
As observed in Fig. 3A, the proliferation of NP-MSCs significantly decreased in concordance with increased acidic pH levels from pH 7.4 to pH 6.2 (P < 0.05). Human NP-MSCs proliferated at pH 7.1 and pH 7.4, but the proliferation rate at pH 7.4 was greater than that at pH 7.1. However, at pH 6.5 and 6.2, the proliferation rate decreased with more acidic conditions resulting in a greater decrease in rate. Proliferation at pH 6.8 also decreased as a whole. Then, after adding amiloride to the culture medium, the results showed that blockade of ASICs by amiloride could significantly increase the proliferation rate (P < 0.05); at pH 7.1, this beneficial effect was slight (increased by 4% on day 11), while at pH 6.8 and pH 6.5, it was significant (increased by 21.3% and 91.7% on day 11, respectively). However, at pH 6.2, the beneficial effect of amiloride only persisted to 7 days (Fig. 3B1–B4).

The proliferation of NP-MSCs cultured in different acidic pH levels evaluated with cell-counting kit-8 (CCK-8) after 1, 3, 5, 7, 9, and 11 days before
Acidic conditions induced apoptosis of NP-MSCs, while amiloride inhibited this effect
As observed in Fig. 4, the percentage of apoptotic cells significantly increased with increased acidity from pH 7.4 to pH 6.2; at pH 6.2, the percentage of apoptosis was 3.55 times that at pH 7.4 (P < 0.05). Then, after adding amiloride to the culture medium, we observed that the percentage of apoptosis was inhibited by 13.9%, 15%, 16.4%, and 14.9% compared to the percentage at corresponding pH levels (P < 0.05) (Fig. 4).

Percentage of apoptotic cells of NP-MSCs cultured in different acidic pH levels evaluated with flow cytometry after 2 days before and after adding amiloride to the culture medium. *P < 0.05 indicated significant difference between groups.
Acidic conditions increased the expression of ASICs, while amiloride inhibited this effect
The results were expressed relative to the control groups, which included human NP-MSCs cultured under standard conditions (pH 7.4). As observed in Fig. 5, the relative gene expression of ASIC genes increased along with increased acidity from pH 7.4 to pH 6.5 (P < 0.05). Then, after adding amiloride to block ASICs, the relative gene expression of these genes decreased (P < 0.05) compared with NP-MSCs cultured at corresponding pH levels (Fig. 5).
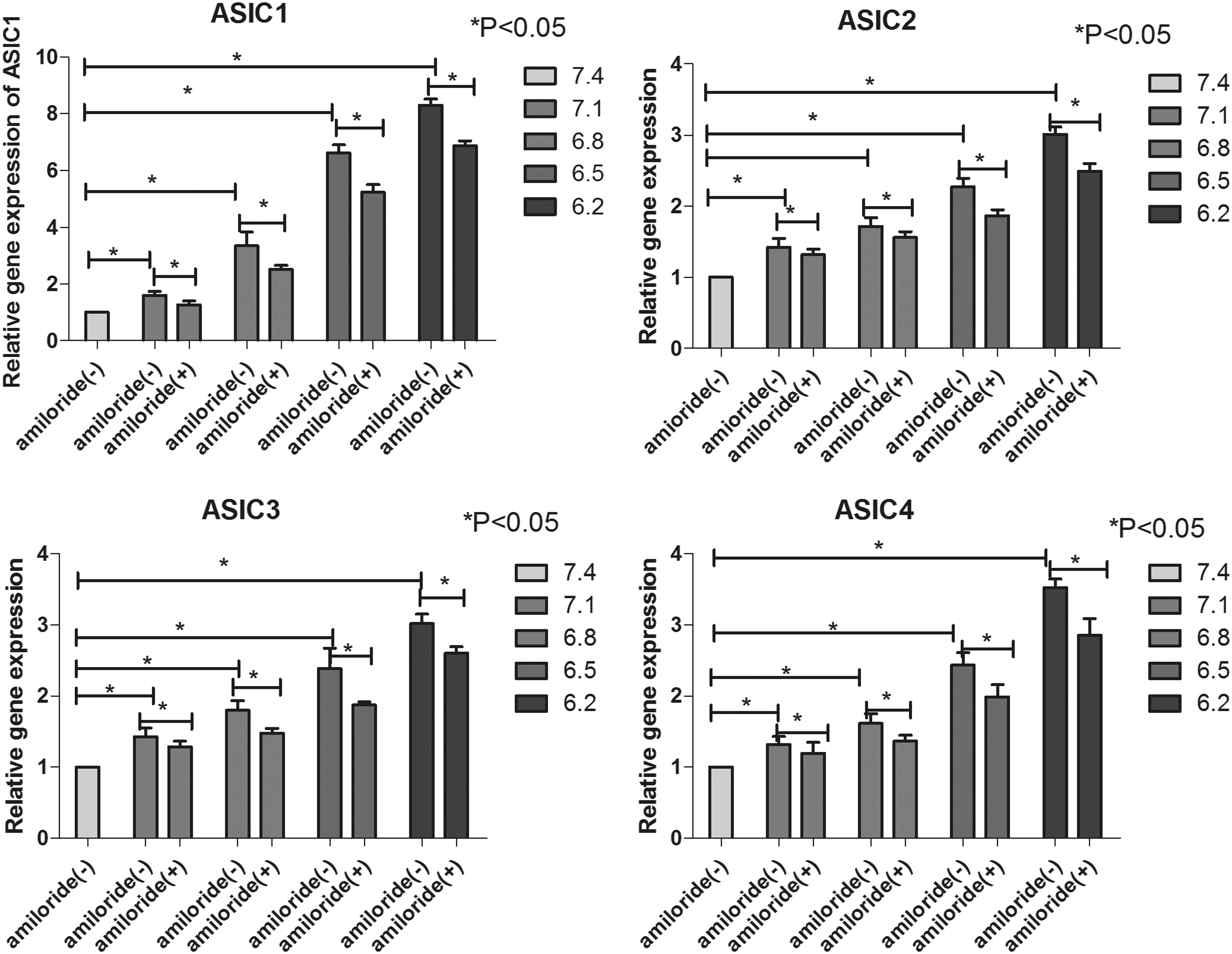
The relative gene expression of ASIC1, ASIC2, ASIC3, and ASIC4 in NP-MSCs cultured in different acidic pH levels after 2 days before and after adding amiloride to the culture medium. *P < 0.05 indicated significant difference between groups.
Acidic conditions inhibited the expression of stem cell-related genes, while amiloride relieved this inhibiting effect
The results were expressed relative to the control groups, in which the human NP-MSCs were cultured under standard conditions (pH 7.4). As observed in Fig. 6, the relative gene expression significantly decreased with increasing acidity (P < 0.05). Then, after adding amiloride to block ASICs, compared with NP-MSCs cultured at corresponding pH levels (pH 7.1, pH 6.8, pH 6.5, and pH 6.2) without amiloride, the relative expression of these genes increased (P < 0.05) (Fig. 6).
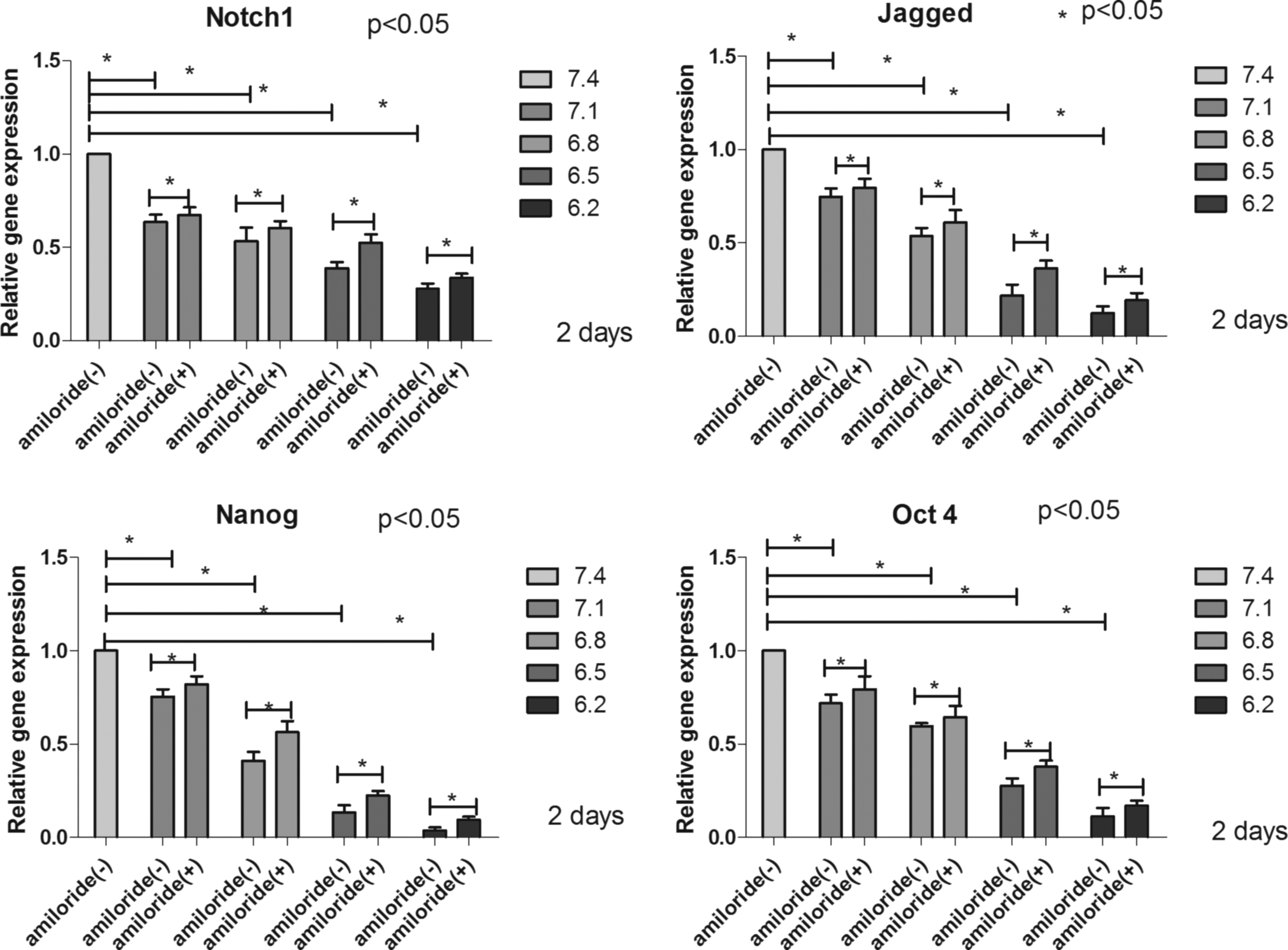
The relative gene expression of Notch1, Jagged, Nanog, and Oct4 in NP-MSCs cultured in different acidic pH levels 2 days before and after adding amiloride to the culture medium. *P < 0.05 indicated significant difference between groups.
Acidic conditions inhibited the expression of Collagen-I, Collagen-II, Aggrecan, and SOX-9, while amiloride relieved these inhibiting effects
The results were shown relative to the control groups, in which the human NP-MSCs were cultured under standard conditions (pH 7.4). As observed in Fig. 7, the relative gene expression significantly decreased with increasing acidity (P < 0.05). Then, after adding amiloride to block ASICs, compared with NP-MSCs cultured at corresponding pH levels (pH 7.1, pH 6.8, pH 6.5, and pH 6.2) without amiloride, the relative gene expression increased (P < 0.05) (Fig. 7).

The relative gene expression of Collagen-II, Aggrecan, SOX-9, and Collagen-I in NP-MSCs cultured in different acidic pH levels 7 days and 14 days before and after adding amiloride to the culture medium. *P < 0.05 indicated significant difference between groups.
Discussion
Previous studies showed that IVD was a nonrenewable tissue. However, recent studies confirmed that human NP, which also contained MSC-like cells, could be isolated and proliferated in vitro [30], producing cells similar to BMSCs, sharing morphological characteristics, cell phenotype, and multipotential differentiation. In our study, we isolated stem cells from degenerated IVDs and cultured them in the complete medium. In addition, we observed that (1) these cells were able to grow by adhering to the flask wall and showed long-term proliferation in vitro; (2) these cells highly expressed CD73, CD90, and CD105, while expressed CD34, CD45, and HLA-DR at low levels; (3) these cells could undergo osteogenic, adipogenic, and chondrogenic differentiation; and (4) these cells could express stem cell-related genes (Oct4, Nanog, Jagged, and Notch1). All the above findings indicated that these cells fulfilled the definition criteria of MSCs [31]. Our results were in agreement with Liu's study [32].
The pH within the IVD is lower than that in most tissues in the human body and decreases with age and level of degeneration [33], which causes the microenvironment in the IVD to be acidic [5]. Acidic conditions have been shown to promote matrix catabolism [12 –14], indicating that acidic conditions may link to the cellular process apparent in the IVD degeneration. Furthermore, acidic conditions promoted a decrease in the expression of matrix-associated genes observed in the stem cells [15 –17]. Blanco et al. [34] observed that human NP-MSCs were similar to BMSCs. Whether acidic conditions had similar effects on NP-MSCs has, until now, remained unknown. Thus, in this work, NP-MSCs were cultured under different acidic pH levels to observe the effects of acidic conditions on the biological properties of human NP-MSCs.
The proliferation of human NP-MSCs significantly decreased in conjunction with increased acidity from pH 7.4 to pH 6.2. NP-MSCs proliferated at pH 7.1 and pH 7.4, while the proliferation rate decreased at pH 6.2–6.8. Meanwhile, culture of NP-MSCs in acidic conditions led to a significant increase in the number of apoptotic cells. Our data suggest that acidic cellular microenvironments could inhibit the proliferation of NP-MSCs and contribute to the death of cells undergoing IVD degeneration, and this effect was most significant at pH 6.2.
The function of an IVD depends on the structure and biochemical composition of the IVD matrix, which consists of proteoglycans and collagen. In our study, aggrecan, SOX-9, and collagen type II and I were used as representative of the ECM. PCR results showed that NP-MSCs cultured in acidic conditions had decreased expression of Aggrecan, SOX-9, Collagen- I, and Collagen-II, indicating that acidic conditions may be involved in cellular and molecular changes in NP-MSCs. In addition, these cells expressed stem cell-related genes (Oct4, Nanog, Jagged, and Notch1) [35], and our study showed that the expression levels of stem cell-related genes were downregulated by acidic conditions. All the above results showed that acidic conditions could not only inhibit proliferation and ECM synthesis but also promote the apoptosis and death of human NP-MSCs. Therefore, these results may explain why these NP-MSCs could not differentiate into IVD cells, which could maintain the stability of IVD or reverse the degenerative process in some ways.
Some studies have found that pH regulates cells' biological activities through ASICs. Researchers observed [27] that acid-induced apoptosis through ASIC1 occurred in endplate chondrocytes. Uchiyama et al. found that low pH could regulate mouse-derived nucleus pulposus cells' biological activities through ASICs, especially ASIC3 [36]. ASICs, including AISC-1, -2, -3, and -4, have been discovered in both normal and degenerated IVDs, and expression of ASIC1–3 within degenerated IVDs was significantly higher than that within normal IVDs [37]. Our study is the first to show that human NP-MSCs cultured in an acidic pH increased the expression of ASICs (ASIC1, ASIC2, ASIC3, and ASIC4), and the expression of AISCs increased along with increasing acidity from pH 7.4 to pH 6.2. These results suggested that ASICs may play an important role in the pH-dependent response of NP-MSCs.
ASICs as a potential therapeutic target to relieve the underlying damage of an acidic environment were confirmed by use of the ASIC antagonist amiloride [38,39]. Amiloride has a low potency (EC50 of 10–100 μM) and binds in the pore of ASICs [40]. Thus, in this study, we further assessed the effect of blocking ASICs by amiloride on the biological activities of human NP-MSCs. Our study observed that treatment of human NP-MSCs with 100 μM amiloride prevented the acid-induced decrease in cell proliferation and ECM gene expression (Aggrecan, SOX-9, Collagen II, and Collagen I). Blocking ASICs by amiloride could also prevent human NP-MSCs from acid-induced apoptosis. In addition, treatment of human NP-MSCs with 100 μM amiloride prevented the decrease in the expression of stem cell-related genes (Oct4, Nanog, Jagged, and Notch1).
Treatment of human NP cells with APETx2 (a specific inhibitor of ASIC-3) could prevent the increase in proinflammatory cytokine and neurotrophin expression; however, it was unable to influence the proliferation, cell death, and expression of aggrecan at different pH levels [12]. The inhibitor used in this study (amiloride, a nonspecific inhibitor for ASICs) may suggest reasons for potential therapeutic effects of ASIC blockers on cell proliferation and ECM synthesis.
These results showed that blockade of ASICs by amiloride could relieve the harmful effect induced by acidic conditions, which further demonstrated that an acidic pH may regulate the biological activities of NP-MSCs through ASICs. In our study, we also observed a decrease in the expression of ASICs (AISC1, ASIC2, ASIC3, and ASIC4) after adding amiloride to the culture medium under acidic conditions, but the mechanisms behind this are unknown and need to be further investigated.
However, there are some limitations in our study. First, the effect of an acidic environment on human NP-MSCs was performed in vitro, so further studies need to be carried out in vivo. The microenvironment of degenerated IVDs is complex, characterized by acidic conditions, hypoxia, high osmotic pressure, and limited nutrition [41 –43]. It was reported that many factors in the microenvironment of the degenerated IVD significantly impair the proliferation and matrix synthesis of bone marrow mesenchymal stem cells [16,35], but the inhibiting effect of low pH was the most obvious. Thus, our work mainly explored the effects of acidic conditions on human NP-MSCs. In addition, we used fetal calf serum to culture human NP-MSCs, not human serum. Fetal calf serum may contain growth factors that are not present in adult human serum and could affect the results, but the resource of human serum was limited.
Second, our study mainly provided the expression of genes to indirectly reflect the changes that appeared in an acidic microenvironment, devoid of proteins that may directly reflect the results. Third, the study was conducted in a two-dimensional culture medium, and NP-MSCs may respond differently in a three-dimensional culture system. Finally, more work needs to be performed to investigate the effects of knockout and siRNA knockdown of ASIC genes on the biological properties of human NP-MSCs under acidic conditions. In addition, the precise mechanism of ASICs in regulating the metabolic activity of human NP-MSCs in response to acidic environments remains unclear.
In summary, our results indicated that an acidic environment is a deleterious factor that inhibits the biological activities of human NP-MSCs, and this inhibiting effect may occur through ASICs. We hypothesized that blockade of ASICs by amiloride may protect NP-MSCs from acid-induced injury.
Footnotes
Acknowledgment
This work was supported by grants from the National Nature Science Foundation of China (no. 81472088, no. 81601935).
Author Disclosure Statement
No competing financial interests exist.