
Abstract
Sepsis is the main cause of morbidity and mortality in neonates. Mesenchymal stromal cells (MSCs) are potent immune-modulatory cells. Their effect in neonatal sepsis has never been explored. We hypothesized that human umbilical cord-derived MSCs (hUC-MSCs) improve survival in experimental neonatal sepsis. Sepsis was induced in 3-day-old rats by intravenous injection of Escherichia coli (5 × 105/rat). One hour after infection, rats were treated intravenously with normal saline, hUC-MSCs, or with interferon-γ preconditioned hUC-MSCs (107 cells/kg). Eighteen hours after infection, survival, bacterial counts, lung neutrophil and macrophage influx, phagocytosis and apoptosis of splenocytes plasma, and LL-37 concentration were evaluated. Animals were observed for survival for 72 h after E. coli injection. Treatment with either hUC-MSCs or preconditioned hUC-MSCs significantly increased survival (hUC-MSCs, 81%; preconditioned hUC-MSCs, 89%; saline, 51%; P < 0.05). Both hUC-MSCs and preconditioned hUC-MSCs enhanced bacterial clearance. Lung neutrophil influx was decreased with preconditioned hUC-MSCs. The number of activated macrophages (CD206+) in the spleen was increased with hUC-MSCs and preconditioned hUC-MSCs; preconditioned hUC-MSCs increased the phagocytic activity of CD206+ macrophages. hUC-MSCs and preconditioned hUC-MSCs decreased splenocyte apoptosis in E. coli infected rats. Finally, LL-37 plasma levels were elevated in neonatal rats treated with hUC-MSCs or preconditioned hUC-MSCs. hUC-MSCs enhance survival and bacterial clearance in experimental neonatal sepsis. hUC-MSCs may be an effective adjunct therapy to reduce neonatal sepsis-related morbidity and mortality.
Introduction
S
Mesenchymal stromal cells (MSCs) are putative repair cells originally isolated from bone marrow [10], and later also identified in other tissues (eg, fat, umbilical cord blood, and placental tissues) [11,12]. MSCs are increasingly investigated as a promising cellular therapy in regenerative medicine. Encouraging preclinical studies have led to early phase clinical trials for a variety of diseases including heart failure, acute lung injury or graft versus host disease [13 –15]. Recent evidence suggests bone marrow-derived MSCs improve survival in adult mouse models of sepsis due to their main ability of immune-modulation but also through enhanced bacteria clearance and organ protection [16 –19]. This has led to a current phase I trial in adults with septic shock (NCT02421484).
Human umbilical cord-derived MSCs (hUC-MSCs) from the Wharton's jelly is of particular clinical relevance for the treatment of neonatal diseases and possess additional advantages: UC is readily available at birth; 100 million births wordwide offer a large source of stem cells that can be safely and painlessly harvested after delivery from otherwise discarded tissue and is thus devoid of ethical dilemma [20]; Importantly, hUC-MSCs appear to have higher anti-inflammatory [21], neuroprotective and proangiogenic properties in vitro [22] and superior cardiovascular healing capabilities in vivo compared to adult bone marrow-derived MSCs [23], but more extensive studies are required to provide robust evidence. These advantages make hUC-MSCs a highly attractive alternative for cell-based therapies, not only for neonates.
At the present time, only bone marrow-derived MSCs have been investigated and solely in adult models of sepsis. Because neonates display different and incompletely characterized immune responses to infection, much remains to be learned regarding the potential effect of hUC-MSCs in neonatal sepsis. In addition, preconditioning of hUC-MSC as a means to boost the MSCs' immunoregulatory function [18,24] has not been explored. We hypothesized that hUC-MSCs and interferon (IFN)-γ preconditioned hUC-MSC would improve survival and enhance bacterial clearance in E. coli induced neonatal sepsis.
Methods
All animal protocols were approved by the Animal Care Committee of the University of Ottawa.
hUC-MSC and preconditioned hUC-MSC culture
Cell isolation
hUC-MSCs were isolated from the UC of healthy, term pregnancies after parental consent. After delivery, the cord was dissected from the placenta, milked to remove remaining intravascular blood, and placed in a storage solution containing 30% [vol/vol] citrate phosphate dextrose adenine anticoagulant (CPDA-1) in phosphate-buffered saline (PBS). After disinfection with povidone-iodine (Purdue Pharma), multiple incisions were made into the epithelium of the cord. Sterile cord tape (Ethicon) was used to ligate the cord vessels before placing the tissue in Dulbecco's PBS (D-PBS) supplemented with Ca2+, Mg2+, glucose, and pyruvate (Thermo Fisher Scientific) along with 750 U/mL collagenase and 500 U/mL hyaluronidase (both from Worthington). Following digestion at 38°C for 4 h, 2,000 BAEE-U/mL trypsin and 3 mM ethylenediaminetetraacetic acid (both from Sigma-Aldrich) were added and allowed to incubate for another 30 min at 38°C. The dissociation process was stopped by placing the mixture on ice and adding fetal bovine serum (FBS) to a final concentration of 10% [v/v]. Remaining undigested tissue was removed by utilizing 40 μm cell strainers (BD Biosciences). Cells were rinsed in PBS and seeded in minimum essential medium Eagle, alpha modification (αMEM) containing 20% FBS (both from Sigma-Aldrich) and standard antibiotics (Thermo Fisher Scientific) in tissue culture treated plastic flasks (Greiner Bio-One). Culture was maintained in a 5% CO2, 5% O2 humidified atmosphere with media changes every 3rd day. Upon reaching 80%–90% confluence, cells were lifted using recombinant trypsin (Thermo Fisher Scientific) and reseeded as passage 1 (P1) MSCs at 5,000 cells per cm2.
Cell characterization
P2 MSCs displayed typical spindle-shaped morphology (Fig. 1A) and were harvested and stained with antibodies against CD11b, CD19, CD34, CD45, HLA-DR, CD73, CD90, and CD105 using a multicolor FACS kit (BD Biosciences) according to the manufacturer's directions and analyzed on a FACS Canto II (BD Biosciences) (Fig. 1B).
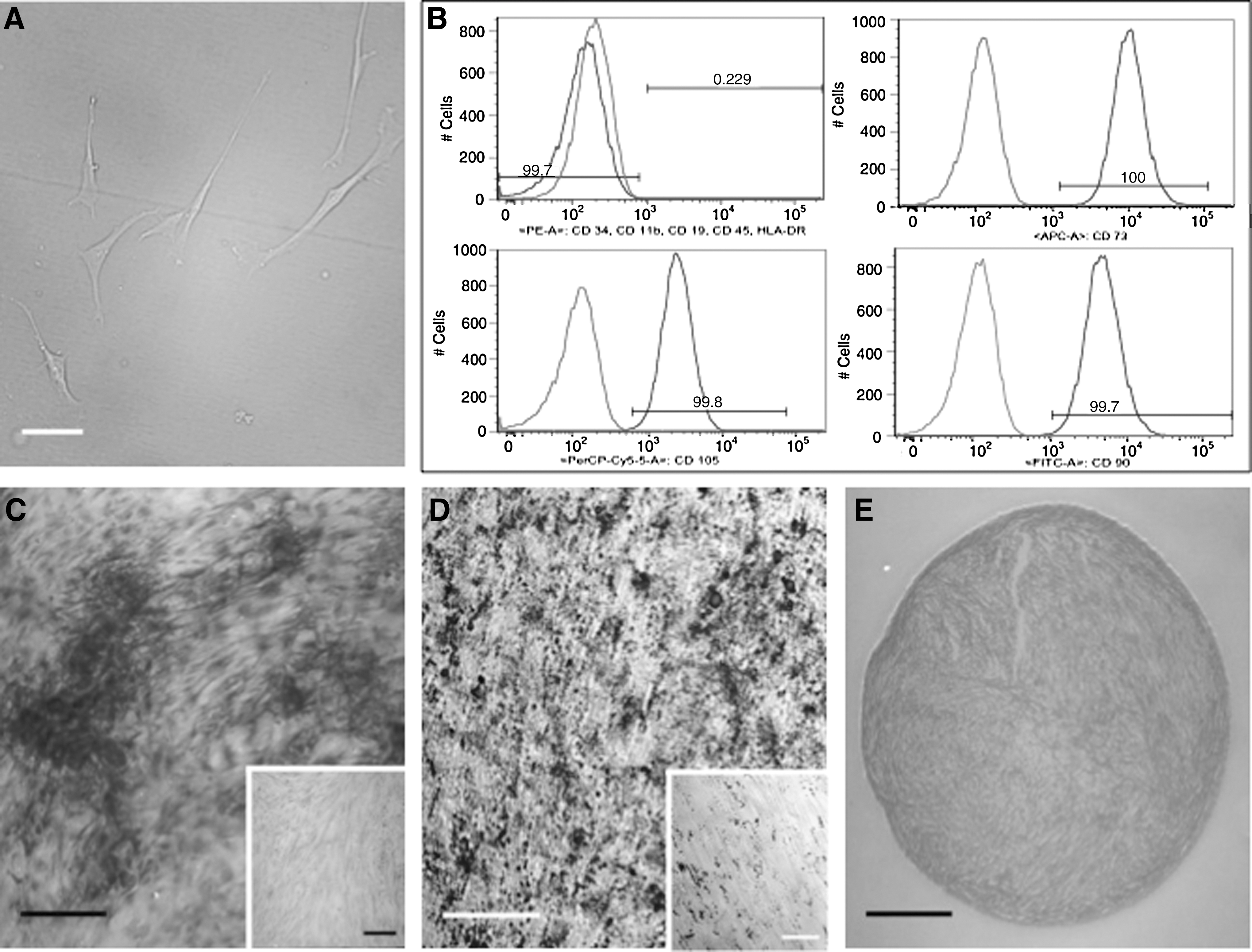
Characterization of hUC-MSCs.
To demonstrate tri-lineage differentiation potential, P2 MSCs were induced using commercially available media formulations (adipogenic and osteogenic: Lonza; chondrogenic: R&D Systems) according to the manufacturer's directions (Fig. 1C). Following fixation with 4% paraformaldehyde (PFA), adipogenic and osteogenic cultures were stained with 0.36% [w/v] Oil-red O (Sigma-Aldrich, St. Louis, MO) in isopropanol or 2% [w/v] Alizarin red (Sigma-Aldrich) respectively. Chondrogenic assay products were embedded after fixation in 4% [w/v] PFA (Sigma-Aldrich). Five micrometer sections were stained with 1% Alcian blue solution (Sigma-Aldrich) to reveal deposited glycosaminoglycans. Images were acquired using an Axiovert 200 M/ICc1 system (Carl Zeiss Microimaging) or an Eclipse TE2000-E/DXM1200C platform (Nikon). After characterization, cells were expanded and frozen in pentastarch-based cryomedium (6% HES 200/0.5, 5% dimethyl sulfoxide) until use.
Cell preparation before injection
P2 cells were thawed and expanded in tissue culture flasks. For hUC-MSC therapy, cells were cultured in 20% FBS/αMEM for 4 days until they reached 100% confluence. For preconditioned hUC-MSCs therapy, cells were treated with 50 ng/mL human IFN-γ (R&D Systems) for 24 h before collection. Before each experiment, cells were trypsinized, counted, washed, and resuspended in sterile PBS to obtain a final concentration of 4 × 106 cells/mL, such that a 25 μL dose contained 105 cells.
Bacterial culture
We used E. coli serotype K1 (generous gift from Dr. Matthay, UCSF) originally isolated from the blood of a patient with biliary sepsis [25 –27]. Frozen aliquots of E. coli were inoculated into Luria-Bertani Broth (LB, Thermo Fisher Scientific) and grown to mid-logarithmic phase by shaking culture (300 rpm) at 37°C. Bacterial counts were quantified with a spectrophotometer (Ultrospec 2000; Pharmacia Biotech). Fresh cultured E. coli were washed twice with normal saline (0.9% NaCl) and resuspended to the final concentration of 5 × 105 colony forming units (CFU) of E. coli per 25 μL normal saline.
Induction of sepsis and hUC-MSC administration after infection
Pregnant Sprague-Dawley rats were purchased from Charles River Laboratories International. Rats were at 14 days of gestation upon arrival and were housed separately. Neonatal mixed gender pups aged 3–4 days were used in the experiments. Pups were anesthetized by inhalation of isoflurane and injected via the right jugular vein with E. coli in 25 μL normal saline. In preliminary experiments, we examined survival after intravenous injection of 104–106 E. coli CFU/pup. Injection of 5 × 105 CFU of E. coli per pup resulted in survival between 30% and 60% at 72 h. This dose was used in subsequent experiments. One hour after the infection, 25 μL of resuspended hUC-MSCs or preconditioned hUC-MSCs (107cells/kg) were injected intravenously through the left jugular vein. Control animals were injected with an equal volume of normal saline. Pups were randomized to receive different treatments. In survival experiments all pups were observed every 6 h until 72 h. Based on the survival analysis, we carried out separate experiments in which pups were sacrificed at 17 h after hUC-MSC treatment (18 h after the injection of E coli). Samples were collected for further analysis at this time point.
Tissue preparation
At 18 h postinfection, pups were sacrificed with an intraperitoneal euthanyl injection followed by exsanguinization via cardiac puncture. To ensure maximal sterility, animal skin, work surfaces, and instruments were cleaned with 70% ethanol. Blood, spleen, right lobe of the lung, and brain were collected in 1.5 mL sterile tubes and placed on ice for analyses.
Bacterial load determination
Blood was serially diluted with sterile saline into 1:10 and 1:100 for bacterial culture. The spleen, lung, and brain were homogenized in 500 μL of sterile saline and prepared in multiple dilutions (spleen at neat, 1:10; lung at 1:10, 1:100; brain at neat, 1:10). Fifty microliters of diluted samples were plated onto LB agar (Thermo Fisher Scientific) and incubated at 37°C under 5% CO2 for 24 h. Bacterial CFUs were counted. One CFU per plate or higher was determined to be positive bacterial load. Bacterial culture results were expressed as the ratio of animal number with positive blood cultures over the total animal number in a given group.
LL-37 and C-reactive protein measurement
To assess systemic inflammation, we measured plasma level of C-reactive protein (CRP). To investigate antimicrobial candidates contributing to the observed effect of bacterial clearance, we analyzed the plasma level of LL-37, an antimicrobial peptide produced by phagocytic cells, epithelial cells, and MSCs [25]. Blood was taken by cardiac puncture and centrifuged to obtain plasma. Plasma was stored at −80°C until the measurements.
To assess LL-37 production by hUC-MSCs and IFN-γ preconditioned hUC-MSCs, 2.5 × 105 hUC-MSCs were plated in T75 flasks and incubated under the same conditions as described above. Three days later, hUC-MSCs were stimulated or not with 50 ng/mL IFN-γ. Eighteen hours later, the medium was changed to αMEM containing 5% FBS without antibiotics. hUC-MSCs were stimulated or not with 50 ng/mL IFN-γ and costimulated or not with E. coli (11,250 CFU/flask) for 6 h. Then, the cells were washed twice with 1 × PBS and FBS free medium (αMEM containing standard antibiotics) was added to the flasks. Twenty-four hours later, serum-free conditioned media were collected to perform the ELISA. Proteins in the conditioned media were quantified by Bradford assay and secreted LL-37 concentrations were normalized to protein concentrations measured in the conditioned media.
Plasma concentrations of CRP and LL-37 and conditioned media concentrations of LL-37 were determined using an ELISA kit (CRP: R&D Systems; LL-37: Hycult Biotech). The colorimetric reaction was quantified on a microplate reader (Bio Tek).
Immunohistochemistry for lung macrophages and neutrophils
Lungs were inflation fixed at 20 cm H2O2 in 10% formalin and embedded in paraffin as previously described [28]. Four micrometer sections were stained with CD68 (1:400 dilution; Abcam, ab125212) and MPO (1:500 dilution; Abcam, ab45977). Photomicrographs of sections were obtained using a light microscope (DM4000 B LED; Leica) and analyzed with MetaMorph software. For each staining (CD68 staining and MPO staining), four lung sections per animal were analyzed: 10 images per section were taken for a total of 40 images per animal. Total numbers of CD68+ cells (macrophages) and MPO+ cells (neutrophils) were counted in each image and averaged to total number of macrophages or neutrophils per 10 fields of views.
Phagocytosis
After collection, spleens were mashed through 40 μm cell strainers to obtain a single cell suspension in Weiss splenocyte medium on ice [29]. To assess phagocytic activity, the pHRodoEcoli BioParticles (Thermo Fisher Scientific) were added to isolated splenocytes and incubated for 1 h at 37°C. These splenocytes were fixed with 4% PFA and then incubated with rabbit-anti-rat CD206 antibody (Abcam), a marker of activated M2 macrophages [30,31]. Flow cytometry was performed using a BD LSRFortessa cell analyzer (BD Biosciences). Percentages of cells positive for phagocytosed fluorescent bacterial particles and CD206 antibody staining were analyzed with BD FACSDiva software (BD Biosciences).
Splenocyte apoptosis
Spleens were fixed in 10% formalin and embedded in paraffin. Sections (4 μm) were stained for terminal deoxynucleotidyl transferase dUTP nick end labeling(TUNEL). Tissue sections were processed with ApopTag Plus Fluorescein in Situ Apoptosis Detection Kit (Millipore). Photomicrographs of apoptotic cells were obtained with a fluorescence microscope (Zeiss Axio Imager M2 microscope). Apoptotic cells were quantified using CellProfiler cell image analysis software (Broad Institute).
All histological analyses were performed by investigators blinded to the study groups.
Statistical analysis
Comparisons of mortality are presented as Kaplan–Meier survival curves, and differences were assessed by log-rank test. Chi-square test was used to analyze the bacterial culture results. Values are expressed as mean ± SEM. Inter-group differences were compared by analysis of variance (ANOVA) followed by Tukey's multi-comparison post hoc test. The numbers of samples per groups are indicated in the figure legends. Statistical analysis and graphical presentations were performed by GraphPad Prism 7 Software (GraphPad Software). Statistical significance was accepted at P < 0.05.
Results
hUC-MSCs improve survival in E. coli-induced sepsis
Intravenous injection of 5 × 105 CFU of E. coli (Fig. 2A) resulted in severe sepsis with a survival rate of 51% at 72 h (Fig. 2B). hUC-MSCs and IFN-γ preconditioned hUC-MSCs protected neonatal rats against intravenous infection of E. coli: the survival rate 72 h after infection was significantly higher in hUC-MSC and in IFN-γ preconditioned hUC-MSC-treated rats compared to saline-treated rats (81% and 89% vs. 51% respectively, Fig. 2B). The survival rates between the hUC-MSC group and IFN-γ preconditioned hUC-MSC group were not statistically different.
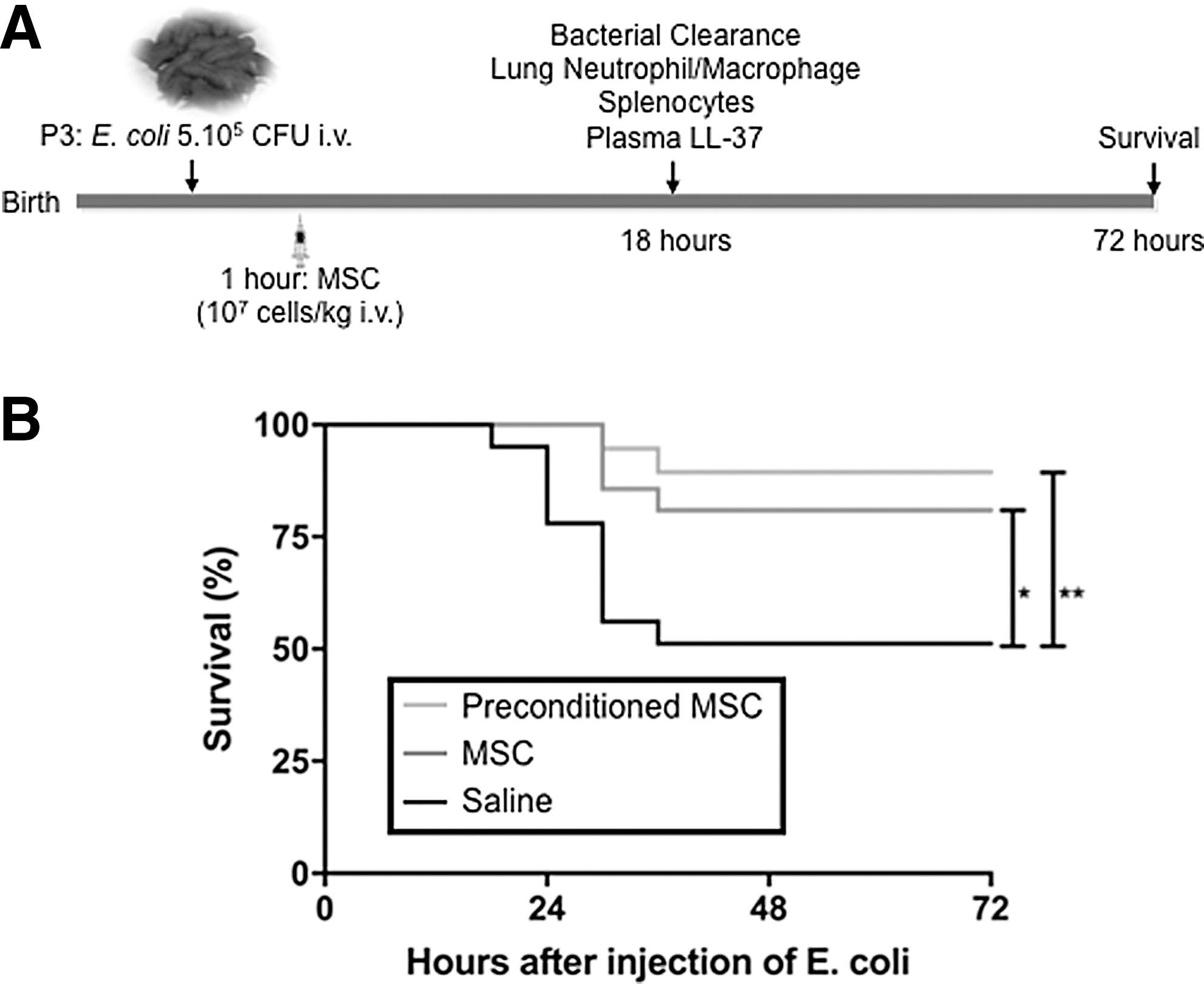
hUC-MSCs improve survival following Escherichia coli-induced sepsis in neonatal rats.
hUC-MSCs enhance bacterial clearance
Measurements of bacterial clearance were performed on samples collected from rats sacrificed 18 h after infection. Bacteria were found in the blood and major organs (lung, spleen, and brain) from a large proportion of pups after E. coli injection (Fig. 3). The ratio of positive bacterial culture in the blood, lung, spleen, and brain was significantly reduced in the hUC-MSC treatment group compared to saline treatment (20% vs. 49%, 58% vs. 75%, 5% vs. 36%, 8% vs. 46%, respectively; Fig. 3A–D). Preconditioned hUC-MSCs increased bacterial clearance in blood, lung, and brain compared to saline treatment (18% vs. 49%, 25% vs. 75%, 25% vs. 46%, Fig. 3A, B, D).
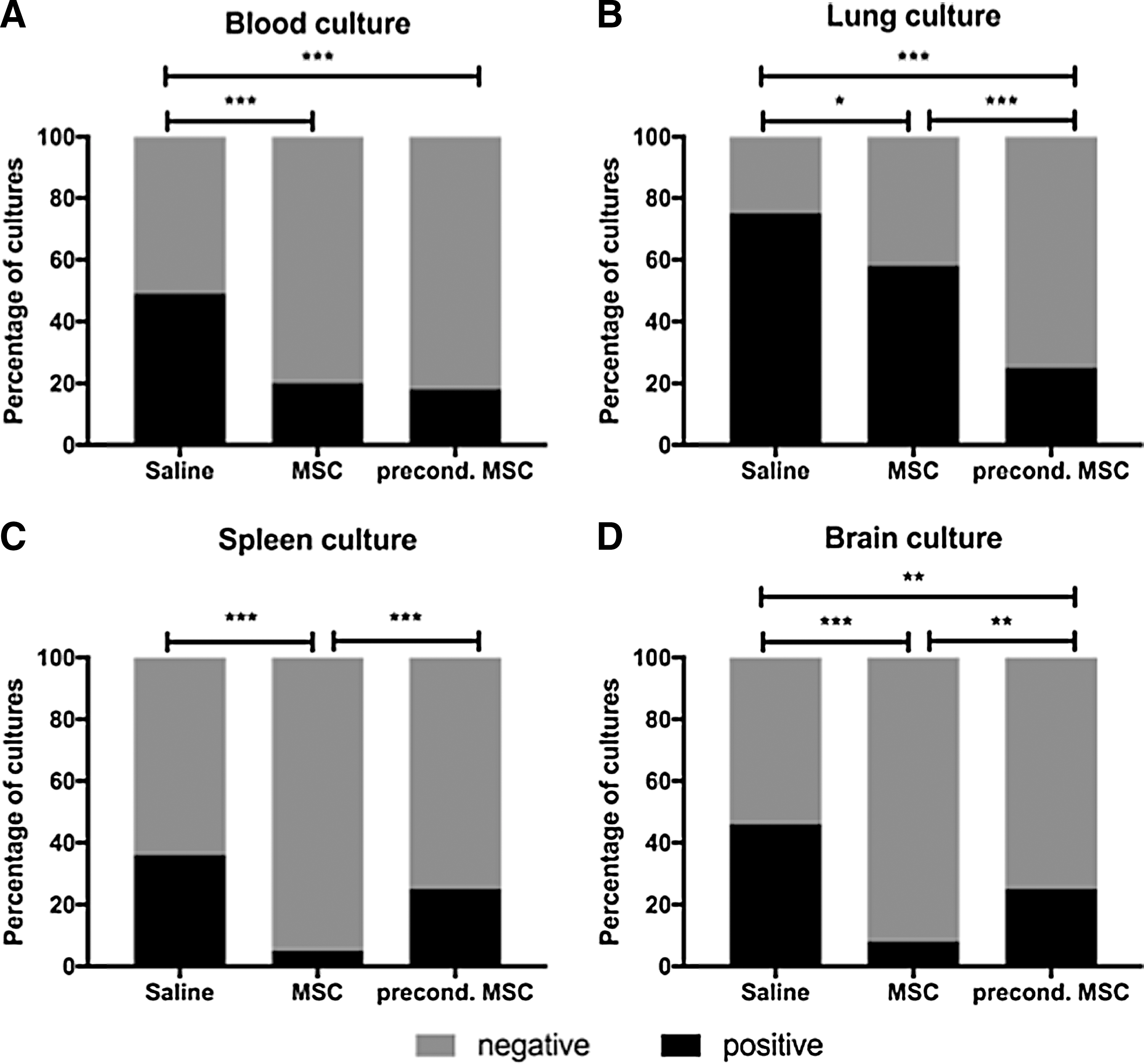
Enhanced bacterial clearance in E. coli-induced sepsis in neonatal rats treated with hUC-MSC.
hUC-MSCs attenuate inflammation, dampen lung neutrophil cell influx, increase phagocytosis, and protect the spleen
hUC-MSC and preconditioned hUC-MSC treatment decreased plasma CRP levels compared to the saline-treated group (Fig. 4A).
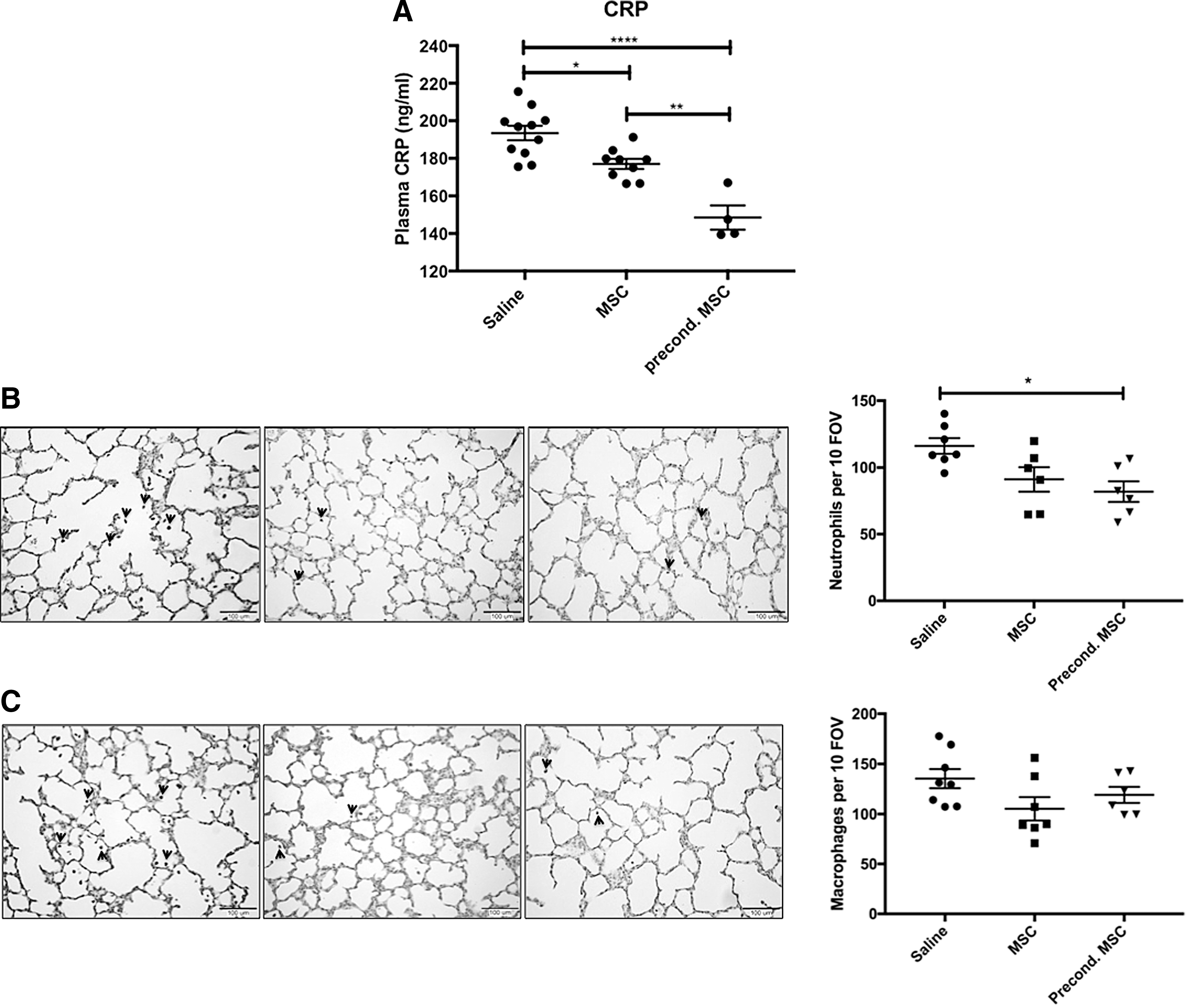
hUC-MSC effect on systemic inflammation and the number of lung neutrophils and macrophages.
In the lung, preconditioned hUC-MSC treatment decreased the number of lung neutrophils in septic rat pups compared to saline-treated pups (Fig. 4B). There was no difference in the number of lung macrophages between groups (Fig. 4C).
In the spleen, hUC-MSC and preconditioned hUC-MSC treatment increased the number of CD206+ cells compared to the saline-treated group (Fig. 5A–D). Preconditioned hUC-MSCs also increased the phagocytic capacity of CD206+ spleen macrophages compared to the saline-treated group (Fig. 5E).
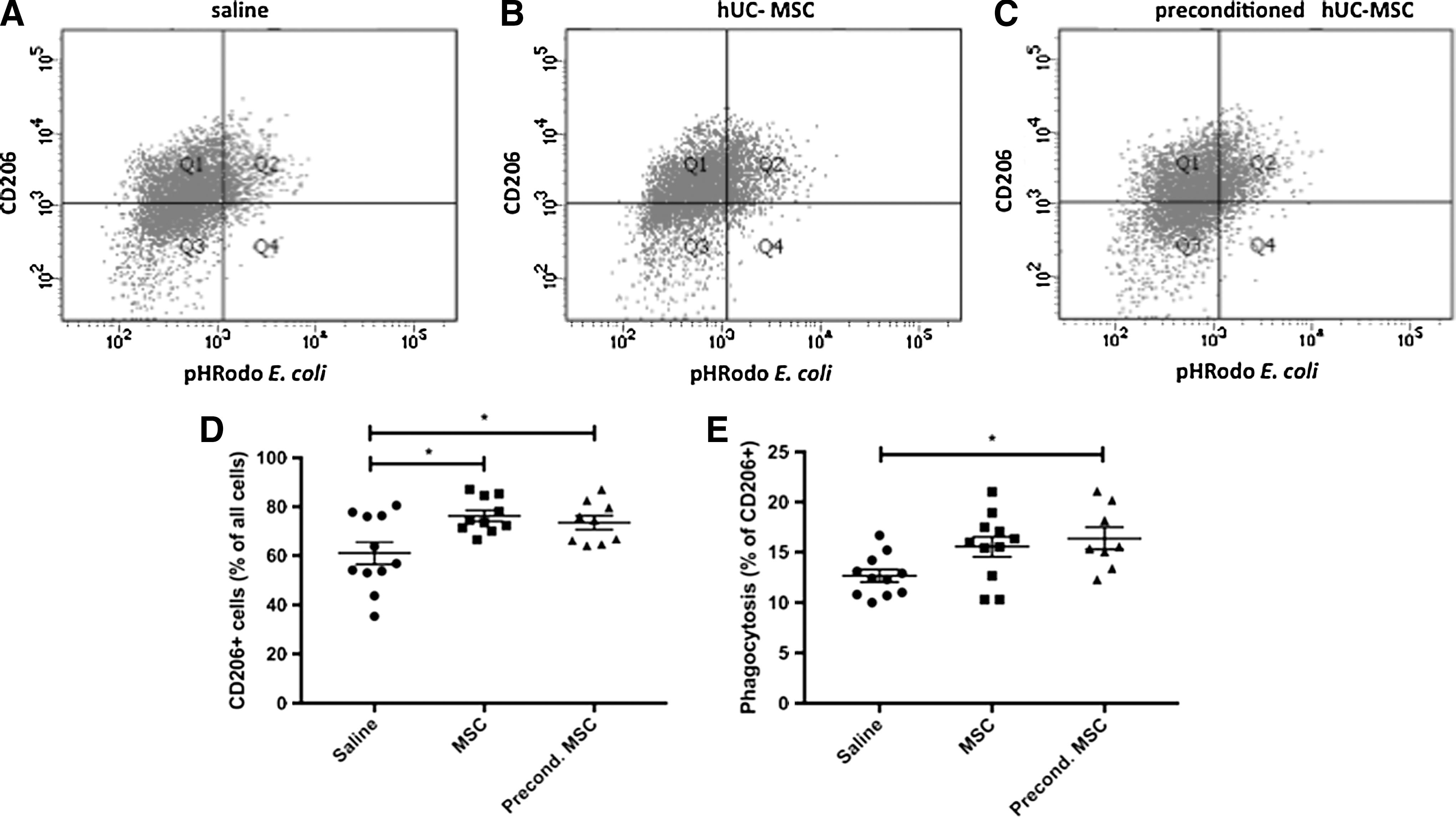
Spleen cells and phagocytic activity.
Apoptosis as indicated by positive TUNEL staining was significantly reduced in the spleens of pups treated with either hUC-MSCs or preconditioned hUC-MSCs compared to saline-treated pups (Fig. 6A, B).
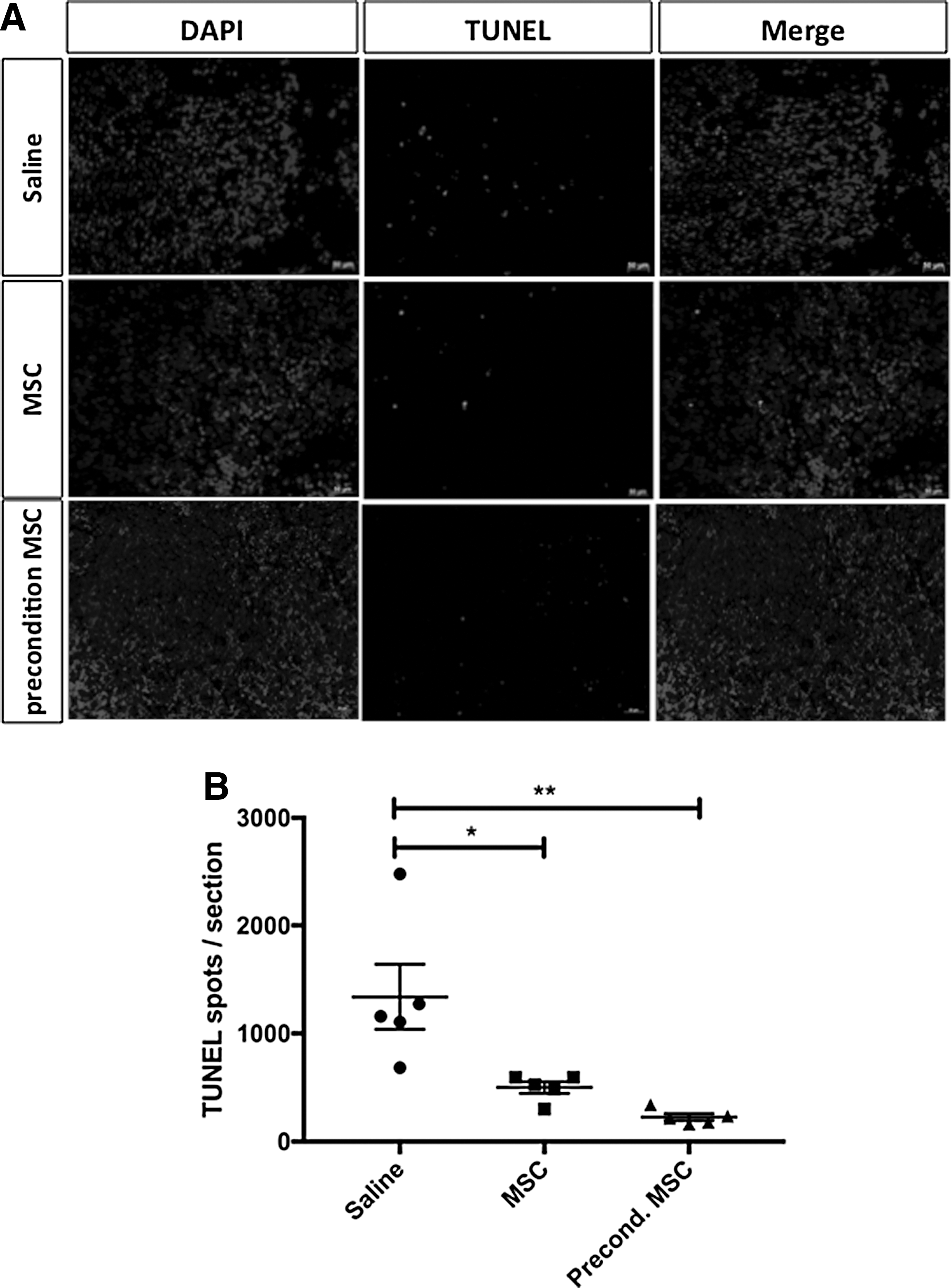
TUNEL Staining of splenocytes.
hUC-MSCs enhance antimicrobial plasma levels
To further explore the potential mechanism for the enhanced bacteria clearance, we investigated the antimicrobial activity of hUC-MSCs. Both hUC-MSC and preconditioned hUC-MSCs increased the plasma concentration of the antimicrobial peptide LL-37 in E. coli-infected animals compared to saline-treated animals, with no difference between native and preconditioned hUC-MSCs (Fig. 7A). In vitro, E. coli enhanced the production of LL-37 by hUC-MSCs and preconditioned hUC-MSCs (Fig. 7B). Preconditioning did not enhance LL-37 production compared with native MSCs.
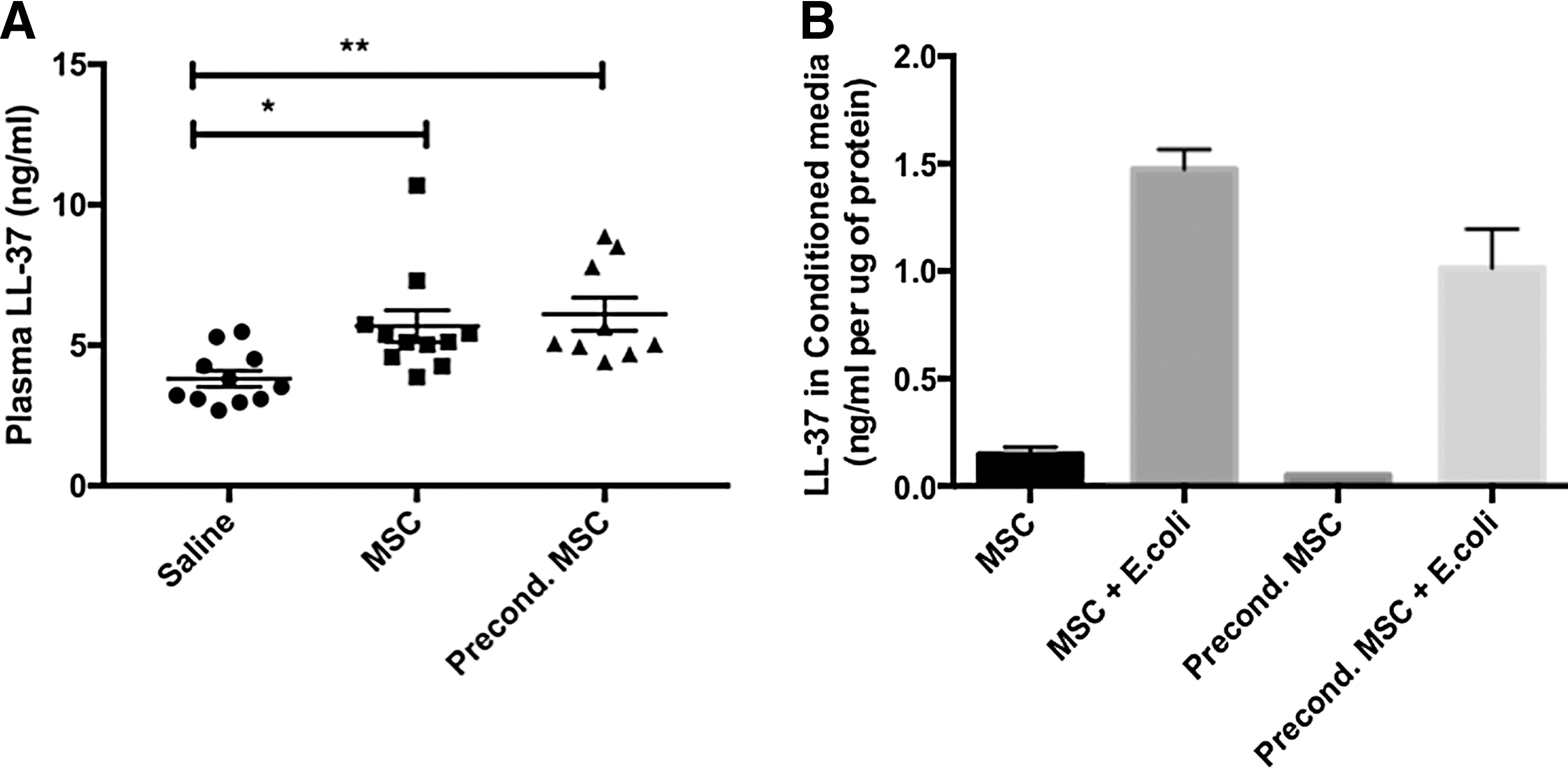
Antimicrobial peptide LL-37.
Discussion
The immature immune response of neonates and the increasing resistance to antimicrobials of bacteria highlight the current limitations of managing sepsis [32,33]. Recent studies have indicated that MSC derived from bone marrow could be beneficial to sepsis in adult animal models [18,34]. This study demonstrates for the first time that MSCs derived from human umbilical cord improve survival and bacterial clearance in E. coli-induced neonatal sepsis in rats, reduce lung neutrophil cell influx and enhance macrophage phagocytosis in the spleen.
Phagocytosis by macrophages, an essential component of innate immunity and host defense, is deficient in preterm infants [35,36]. Boosting this macrophage function by hUC-MSCs may provide a potent adjunct therapy in this specific patient population. We investigated the number of macrophages in the septic lung and phagocytic activity of the macrophages in the spleen of septic pups. Macrophages have two different polarization states, the classically M1 phenotype and the alternatively activated M2 phenotype [37]. hUC-MSC treatment increased the absolute number of CD206+ macrophages (M2) in the spleens of septic pups. In addition, preconditioned hUC-MSCs significantly increased the phagocytic activity of CD206+ macrophages compared to saline-treated septic pups. Our findings in neonatal rats are consistent with previous findings from our lab and others [38 –41] in adult animals, demonstrating that MSCs regulate the phenotypes of macrophages and enhance the phagocytosis of macrophages exposed to pathogens. These data may partly explain the enhanced bacterial clearance in infected rat pups.
Our study also found that the intravenous injection of hUC-MSCs significantly decreased the number of apoptotic splenocytes. The spleen is the largest immune organ of the body and plays an important role in hormonal and cellular immune responses in serious infection. As a consequence of sepsis, spleen cells undergo apoptosis leading to immune cell loss and immune cell dysfunction that will decrease the capability of clearance of pathogen [19,42,43]. MSCs are capable of attenuating cell apoptosis by regulating the ratio of the antiapoptotic protein (Bcl-xL) to proapoptotic protein (Bax) expression [44]. Thus, some of the salutary effects of hUC-MSC in our study may be explained by the preservation of splenocytes in septic neonatal rat pups.
Similar to bone marrow-derived MSCs in septic adult animal models [34,45], our study showed that hUC-MSCs increase survival of neonatal septic rats and inhibit bacterial growth. To investigate the mechanism of the beneficial effects of hUC-MSCs in our model, we measured the plasma levels of LL-37, an antimicrobial peptide secreted by cells in response to environmental signals. LL-37 belongs to the cathelicidin family and is produced by phagocytic leukocytes, epithelial cells, and MSCs [46,47]. LL-37 is able to kill gram-positive or gram-negative bacteria directly, and provide protection against a wide variety of pathogens [48,49]. Previous studies indicated that exposure to bacteria could enhance the secretion of LL-37 by human MSC [25,46]. LL-37 may also lower the severity of sepsis in neonates [50]. Our study demonstrates that hUC-MSCs are capable of elevating LL-37 plasma levels and this mechanism may contribute to the improvement in survival.
Various preconditioning strategies including exposure of cells to hypoxia/hyperoxia, LPS, stroke serum, pharmacological agents, cytokines, and growth factors have been proposed to enhance the therapeutic potential of MSCs in ischemic diseases, neonatal lung injury, lung fibrosis, and wound healing [51 –54]. There is no publication of the effect of preconditioned MSC on neonatal sepsis. In this study, IFN-γ preconditioned hUC-MSCs were very effective in improving survival in this neonatal E. coli sepsis model even though there was no statistical difference compared to native hUC-MSC. Likewise, IFN-γ preconditioned hUC-MSCs induced slightly higher LL-37 plasma levels than native UC-MSC. IFN-γ preconditioned hUC-MSCs also enhanced splenocyte phagocytosis. However, sepsis is a complex disease involving multiple interactions between the microbial pathogen and the host systemic inflammatory response. Thus, ex vivo preconditioned hUC-MSC with IFN-γ alone may not sufficiently mimic the infection and appropriately prime the hUC-MSCs. In addition, the therapeutic effect achieved with native hUC-MSC, such as the survival rate, was already high, making it challenging to attain significance in further improvements. These facts may explain the reason IFN-γ preconditioning could not further increase the therapeutic property of hUC-MSC in this study. Nonetheless, the preconditioning approach appears promising and warrants further exploration by varying the type, dose, and timing of the preconditioning stimulus.
In conclusion, intravenous hUC-MSC therapy improves survival in neonatal E. coli induced sepsis. The beneficial effect is associated with enhanced bacterial clearance in major organs. The elevated bacteria clearance may relate to enhanced phagocytosis by activated macrophages, decreased apoptosis of splenocytes, and increased plasma LL-37 concentrations. IFN-γ preconditioned MSCs and native MSCs appeared therapeutically equally effective in E. coli–induced sepsis, and should both be further explored as promising therapeutic strategies in neonatal sepsis.
Footnotes
Acknowledgments
We acknowledge Shanghai Municipal Education Commission, China for their support to this project. In addition, we would like to acknowledge Dr. Michael Matthay for his generous gift of the E. coli used in this study. Y.Z. was supported by a stipend from Xinhua Hospital/Shanghai Jiaotong University School of Medicine. J.J.P.C. was supported by a Canadian Institutes of Health Research (CIHR) postdoctoral fellowship. B.T. is supported by the CIHR, Canadian Thoracic Society, Stem Cell Network and the Children's Hospital of Eastern Ontario foundation.
Author Disclosure Statement
No competing financial interests exist.