
Abstract
An undesirable complication that arises during dental treatments is external apical-root resorption, which causes root-cementum and root-dentin loss. To induce de novo cementogenesis, stem cell therapy is required. Cementum-forming cells (cementoblasts) are known to be differentiated from periodontal-lineage mesenchymal stem cells (MSCs), which are derived from the dental follicle (DF) in developing tissues and the periodontal ligament (PDL) in adult tissues, but the periodontal-lineage MSC type that is optimal for inducing de novo cementogenesis remains unidentified, as does the method to isolate these cells from harvested tissues. Thus, we investigated the cementogenic potential of DF- and PDL-derived MSCs that were isolated by using two widely used cell-isolation methods: enzymatic digestion and outgrowth (OG) methods. DF- and PDL-derived cells isolated by using both methods proliferated actively, and all four isolated cell types showed MSC gene/protein expression phenotype and ability to differentiate into adipogenic and chondrogenic lineages. Furthermore, cementogenic-potential analysis revealed that all cell types produced alizarin red S-positive mineralized materials in in vitro cultures. However, PDL-OG cells presented unique cementogenic features, such as nodular formation of mineralized deposits displaying a cellular intrinsic fiber cementum-like structure, as well as a higher expression of cementoblast-specific genes than in the other cell types. Moreover, in in vivo transplantation experiments, PDL-OG cells formed cellular cementum-like hard tissue containing embedded osteocalcin-positive cells, whereas the other cells formed acellular cementum-like materials. Given that the root-cementum defect is likely regenerated through cellular cementum deposition, PDL-OG cell-based therapies might potentially facilitate the de novo cellular cementogenesis required for regenerating the root defect.
Introduction
E
For future cell-based therapies, mesenchymal stem cells (MSCs) are considered to represent a promising cell source. Human MSCs have been isolated from numerous sources, including bone marrow [6], umbilical cord [7], umbilical cord blood [8], adipose tissue [9], placenta [10], breast milk [11], and teeth [12 –16]. MSCs can differentiate into osteogenic, chondrogenic, adipogenic, myogenic, and neurogenic lineages in vitro [17 –19], and MSCs have been used in preclinical trials in therapy for severe refractory diseases [20 –23]. However, among the diverse sources of stem cells, the stem cell compartments derived from teeth are likely the most suitable for regenerating the dental root cementum. Human teeth, which can be obtained by using routine procedures, are typically discarded during dental treatments and, thus, represent a ready source of material for both basic and clinical research. In addition, in terms of embryologic importance, teeth are unique in that they are research materials that can be used for investigations (including studies on stem cells) on human tissues/organs/anatomies at both immature and completely mature stages of development.
Certain types of human MSCs are widely recognized to be isolated from dental tissues of extracted wisdom teeth: dental pulp stem cells (DPSCs) [12] and periodontal ligament stem cells (PDLSCs) [14], which are derived from the dental pulp and periodontal ligament (PDL) of mature wisdom teeth; and stem cells from apical papilla [16] and dental follicle (DF) precursor cells [15], which are derived from the apical papilla and DF of immature wisdom teeth, respectively. These dental tissue-derived MSCs possess the stem cell characteristics of clonogenicity and multilineage differentiation potential, which are comparable with those of other types of MSCs [24]. Recently, certain dental MSCs have been used in preclinical studies on large animals to assess their regenerative potential [25,26].
To date, very few studies have comparatively analyzed multiple dental MSC types isolated and grown under identical culture conditions. An enhanced understanding of the biological characteristics of these dental MSC populations is essential for investigating their potential for clinical application. We previously reported that four types of human dental MSCs, which were derived from dental pulp, apical papilla, PDL, and DF, showed greater proliferative ability than did bone marrow-derived MSCs (BMMSCs) obtained from the iliac bone [27]. Moreover, the dental MSCs presented multidifferentiation potential and gene-expression profiles that were equivalent to those of BMMSCs. However, the type of dental MSC that is the most suitable for potential application in de novo cementum formation remains unestablished. With regard to periodontium development, DF-derived stem cells give rise to three types of periodontal-lineage cells, cementoblasts, osteoblasts, and fibroblasts, which produce cementum, alveolar bone, and PDL, respectively [28]. In the adult periodontium, the PDL-derived stem cells play the central role of recruiting the aforementioned three types of periodontal functional cells. Thus, the stem cells derived from DF and PDL, which are periodontal-lineage tissues, are considered to hold the potential for differentiating into cementoblasts. However, further investigation is required to comparatively analyze the cementoblastic potentials of the specific cell types derived from DF and PDL.
In addition to the matter related to cell types, the methods used for primary cell isolation from harvested tissues distinctly influence stem cell properties [29 –32]. The two cell-isolation methods that have been the most widely used thus far for obtaining dental MSCs are the enzymatic digestion (EZ) method and the outgrowth (OG) method. The EZ treatment involves the application of collagenase and/or dispase to digest the harvested tissue for acquiring single-cell suspensions, after which the obtained cells are cultivated [12,27]. The OG procedure is based on the explant culture of harvested tissue fragments on culture dishes to facilitate the OG of the cells from the tissue fragments [33,34]. However, the isolation method that should be favored when obtaining dental MSCs remains debated. Therefore, in this study, we investigated the in vitro and in vivo cementoblastic potential of DF- and PDL-derived stem cells exhibiting MSC characteristics that were obtained by using either the EZ or the OG procedure. Our aim was to comprehensively address the clinically important question of which cell type would be the most suitable for de novo cementum formation.
Materials and Methods
Ethics statement
This study was approved by the ethics committee of Nippon Dental University School of Life Dentistry at Tokyo (Approval no. NDU-T2013-10), and it was conducted in accordance with the amended Declaration of Helsinki. Informed consent was obtained from each study participant after fully explaining the nature of the procedure and the intended use of the tissue obtained.
Tissue collection
Normal human third molars and the alveolar bone of the mandible were collected from systemically healthy patients, aged 15–29 years, undergoing tooth extraction. Tissue was collected from the extracted teeth as previously described [27]. Briefly, DF tissue was separated from the tooth crown of fully impacted immature teeth by using a disposable scalpel, and PDL tissue was scraped from the middle third of the fully grown tooth root with a sterilized razor blade. The collected tissues were rinsed, minced (into pieces ∼2 × 2 mm in size), and subjected to the EZ and OG procedures. At least five teeth were collected for obtaining each of the four cell types as described in the following subsections.
Cell isolation by EZ procedure
To obtain enzymatically digested DF and PDL cells, which were designated DF-EZ and PDL-EZ cells, respectively, the minced dental tissues were digested in a solution containing 3 mg/mL collagenase type I (Merck KGaA, Darmstadt, Germany) and 4 mg/mL dispase (Wako Pure Chemical Industries, Osaka, Japan) for 1 h at 37°C [12,27]. Single-cell suspensions were obtained by passing the cells through a 70-μm cell strainer (Merck KGaA), and these suspensions were seeded at 1 × 105 cells/dish in 100-mm dishes (Becton Dickinson, Franklin Lakes, NJ) and cultured in Dulbecco's modified Eagle's medium/Ham's nutrient mixture F12 (DMEM/F12; Thermo Fisher Scientific, Waltham, MA) that was supplemented with 15% fetal bovine serum (FBS; Lot No.: 1010399; Thermo Fisher Scientific), 100 μM glutamate (GlutaMAX I; Thermo Fisher Scientific), 0.1% MEM Non-Essential Amino Acids (Thermo Fisher Scientific), 50 U/mL penicillin, 50 μg/mL streptomycin (Thermo Fisher Scientific), and 0.25 mg/mL Fungizone (Thermo Fisher Scientific); this medium was used as the standard growth medium (GM) throughout this study. At 70%–80% confluence, the cells were detached by using 0.25% trypsin (Becton Dickinson)/0.02% ethylenediaminetetraacetic acid (EDTA; Dojindo, Kumamoto, Japan) and then subcultured at a 1:3 split ratio.
Cell isolation by OG procedure
To obtain outgrown DF and PDL cells, DF-OG and PDL-OG cells, respectively, the minced tissue fragments were placed at the center of 60-mm culture dishes (Becton Dickinson) with a minimal amount of GM. After incubation for 24 h in an incubator, 2 mL of GM was added gently, ensuring that the tissues did not float up from the culture dish bottom. The cells grew out from the tissue fragments within 1 week and then proliferated actively. When the cells reached 70%–80% confluence, the tissue fragments were removed by using forceps; then the cells were detached by using trypsin/EDTA and subcultured at a 1:3 split ratio. All cell cultures were maintained at 37°C in a humidified atmosphere containing 4.7% CO2 in air, and GM was changed every 3–4 days.
In all experiments described next, we used the aforementioned four types of cell populations at Passage 3 or 4, and for each assay, we used cells derived from at least three different study participants.
Cell growth and population doubling time calculation
Cells were seeded at 3 × 103/well in 24-well plates and cultured in GM. The cultured cells were detached by adding trypsin/EDTA and counted in triplicate by using a hemocytometer every 48 h for 12 days. For all cell types, the population doubling time (PDT) was calculated by using this formula: PDT = (t−t 0)log2/log(N−N 0), where t is time (h), N is the number of harvested cells, and N 0 is the number of cells in the inoculum.
Flow cytometric analysis
Flow cytometry was performed to examine the expression of cell-surface markers. The DF and PDL cells isolated by using the EZ and OG methods were trypsinized from culture dishes and fixed in 4% paraformaldehyde (PFA) for 15 min at room temperature (RT). The cells were then incubated for 1 h at RT with an appropriate amount of the following antibodies: fluorescein isothiocyanate (FITC)-conjugated antibodies against CD14 and CD90; phycoerythrin (PE)-conjugated antibodies against CD29, CD31, CD105, CD106, CD140b, and CD146 (Becton Dickinson); FITC-conjugated antibodies against CD34 and CD44; PE-conjugated antibody against CD166 (Beckman Coulter, Brea, CA); and anti-NG2 chondroitin sulfate proteoglycan (NG2) antibody (Merck KGaA). For anti-NG2 staining, the cells were additionally incubated with Alexa Fluor 488-conjugated goat anti-rabbit IgG (H + L) antibody (Thermo Fisher Scientific) for 30 min at RT. The expression profiles were examined and analyzed by using a Guava™ flow cytometer and Guava TM Express Plus (version 5.3) software (Merck KGaA). Data were obtained from three independent experiments.
Reverse transcription-polymerase chain reaction and reverse transcription-quantitative polymerase chain reaction analyses
When the cultured cells reached confluence, total RNA was isolated by using an RNeasy Mini kit (Qiagen, Hilden, Germany), and cDNA was synthesized from 1 μg of total RNA by using a High-Capacity cDNA synthesis kit (Thermo Fisher Scientific). Polymerase chain reaction (PCR) amplifications were performed in a 20-μL reaction volume by using a PCR Supermix Platinum kit (Thermo Fisher Scientific). All assay kits were used according to the manufacturer's instructions. The gene-specific primers used here are listed in Table 1. The PCR conditions used were 94°C for 2 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 1 min; PCR was performed in a Veriti™ 96-well Thermal Cycler (Thermo Fisher Scientific). The obtained PCR products were electrophoretically separated on 1.8% (w/v) agarose gels, which were stained with 0.5 μg/mL ethidium bromide, and all PCR products were sequenced to confirm the identity of the amplified products (data not shown). PCR reactions were optimized by using at least three different cycles, and only the results from the linear phase of PCR amplification were used as the representative data. Real-time quantitative PCR (qPCR) was performed by using TaqMan® gene-expression assays and the StepOnePlus™ Real-Time PCR System (Thermo Fisher Scientific), according to the manufacturer's instructions. The qPCR condition was as follows: preincubation at 95°C for 10 min, and then 40 cycles of denaturation at 95°C for 15 s, primer annealing at 60°C for 1 min, and extension at 95°C for 15 s. The primers used for the qPCR targeted the genes encoding these human proteins: cementum attachment protein (CAP; assay ID, Hs00171965_m1; GenBank accession number, NM_014241.3; amplicon length, 74 bp; Thermo Fisher Scientific) and cementum protein 1 (CEMP1; assay ID, Hs04185363_s1; GenBank accession number, NM_001048212.3; amplicon length, 72 bp; Thermo Fisher Scientific). The values obtained for CAP and CEMP1 were normalized to the expression level of the gene encoding β-actin (assay ID, Hs99999903_m1; GenBank accession number, NM_001101.3; amplicon length, 171 bp; Thermo Fisher Scientific), which was used as a housekeeping gene. The relative expression levels of the tested genes were calculated by using the ΔΔCt method. The reactions were run in quadruplicate, and three independent experiments were performed. Total RNA was also extracted from human DPSCs, which we previously reported [27], and the RNA was used as the reference control sample for reverse transcription (RT)-PCR and RT-qPCR.
CAP, cementum attachment protein; CEMP1, cementum protein 1; Collagen I, collagen type I alpha 1 chain; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; GDF5, growth differentiation factor 5; Nanog, nanog homeobox; Oct3/4, POU class 5 homeobox 1; Runx2, runt-related protein 2; Sox2, SRY-box 2.
Immunocytofluorescence
Cultured cells were plated at 1 × 105 cells/well in four-well chamber slides and fixed with cold methanol for 10 min at −30°C. After washing with PBS, cells were incubated in Blocking One Histo (Nacalai Tesque, Kyoto, Japan) for 10 min at RT. Cultures were incubated with the following primary antibodies at 4°C overnight: rabbit polyclonal anti-CAP (1:100; LifeSpan Biosciences, Seattle, WA), rabbit polyclonal anti-CEMP1 (1:100; Abcam, Cambridge, United Kingdom), and mouse monoclonal anti-human-specific vimentin (1:1,000; Merck KGaA). After washing with PBS, the samples were incubated with the following secondary antibodies for 30 min at RT in the dark: Alexa Fluor 488-conjugated donkey anti-mouse IgG and Alexa Fluor 488-conjugated goat anti-rabbit IgG (both diluted 1:1,000; Thermo Fisher Scientific). The samples were washed with PBS thrice for 5 min each and mounted with Vectashield mounting medium containing 4′,6-diamidino-2-phenylindole (Vector Laboratories, Burlingame, CA). For negative controls, the primary antibody was omitted during immunostaining. Images were obtained by using a confocal laser-scanning microscope (LSM-510; Carl Zeiss, Oberkochen, Germany).
In vitro multilineage differentiation
To determine the differentiation capabilities of the putative stem cells, all four cell types were subjected to culture conditions that were designed to induce adipogenic-, chondrogenic-, and cementogenic-lineage differentiation.
Adipogenic differentiation
Cells were seeded at 1 × 105 cells/well in six-well plates and maintained in GM until they reached confluence, at which point the cells were transferred to α-modified minimal essential medium (α-MEM; Wako Pure Chemical) containing 10% FBS, 0.5 mM 3-isobutyl-l-methylxanthine (Merck KGaA), 0.5 μM hydrocortisone (Wako Pure Chemical), and 60 μM indomethacin (Merck KGaA) and cultured for 3 weeks. As the control, cells were cultured in α-MEM containing 10% FBS but not the adipogenic-differentiation stimuli. The differentiation medium was changed every 3–4 days. After 3 weeks of differentiation, intracellular accumulation of lipid droplets was visualized by means of oil red O staining.
Chondrogenic differentiation
Cells were subjected to pellet culture as previously described [35], with minor modifications. Briefly, ∼1 × 106 cells in GM were suspended in a 15-mL centrifuge tube (Becton Dickinson) and centrifuged at 430 g for 5 min to generate a pellet. Next, 1 mL of chondrogenic medium consisting of DMEM/F12 containing 10% FBS, 10 ng/mL transforming growth factor-β1 (Peprotech, Oak Park, CA), 1% ITS + 1 supplement (Merck KGaA), and 50 mM
Cementogenic differentiation
Cells were seeded at 1 × 105 cells/well in six-well plates and maintained in GM until confluence, after which the cells were cultured in α-MEM containing 10% FBS plus the mineral-inducing supplements 10 nM dexamethasone (Dex; Merck KGaA), 10 mM β-glycerophosphate (β-GP; Merck KGaA), and 100 μM AP. As the control, cells were cultured in α-MEM containing 10% FBS but not the mineral-inducing supplements. The differentiation medium was changed every 3–4 days, and after 3 weeks of differentiation, the mineralized deposits were visualized through alizarin red S staining. Moreover, the expression profile of CAP and CEMP1 before and after the cementogenic differentiation was analyzed by using RT-PCR and RT-qPCR, as described earlier in this section.
In vivo transplantation
All animal experiments were performed in accordance with the “Regulations for Animal Experimentation and Laboratory Animal Facility” in the Nippon Dental University School of Life Dentistry at Tokyo. The protocol was approved by the Animal Experiments Committee of the Nippon Dental University School of Life Dentistry at Tokyo. All surgery was performed under sodium pentobarbital anesthesia, and all efforts were made to minimize suffering. Approximately 1 × 106 cells expanded in GM (undifferentiated cells) were combined with 40 mg of hydroxyapatite particles (HA; Calcitite, Zimmer Dental, Warsaw, IN), and the cell/HA mixture was centrifuged at 430g for 5 min. The pelleted cell/HA mixture was further mixed with collagen gel (Nitta Gelatin, Osaka, Japan). The four cell/HA constructs, containing DF-EZ, DF-OG, PDL-EZ, or PDL-OG cells, and the HA/collagen-gel scaffold alone were transplanted into subcutaneous pockets formed in the dorsal surface of 6-week-old female C.B-17/lcr-scid/scidJcl (SCID) mice (n = 5 for all groups; Nihon Crea, Tokyo, Japan). All transplants were harvested for histological analysis at 16 weeks after the transplantation.
Histological examination
The collected specimens of the cell/HA transplants, the extracted teeth, and the alveolar bone tissue from the study participants were fixed in 10% neutral buffered formalin, decalcified, embedded in paraffin, and cut into 5-μm serial sections as previously described [34]. The deparaffinized sections were subjected to hematoxylin and eosin (HE) and Masson's trichrome (MT) staining. Immunohistochemical staining was performed by using the following primary antibodies: the aforementioned antibodies used in immunocytofluorescence, against vimentin (1:10,000), CAP (1:100), and CEMP1 (1:100), rabbit polyclonal anti-osteopontin (OPN; 1:1,000, Abcam), and mouse monoclonal anti-osteocalcin (OCN; 1:500, Abcam). Endogenous peroxidase activity was quenched by adding Peroxidase-Blocking Solution Dako REAL™ (Dako, Glostrup, Denmark), and some of the sections were pretreated with 0.1% trypsin/PBS at 37°C for 30 min for antigen retrieval, depending on the primary antibodies used. Sections were incubated for 10 min at RT in Blocking One Histo to block nonspecific binding of antibodies. After overnight incubation at 4°C with primary antibodies, sections were incubated with Dako Envision™ + Dual Link System-HRP for 30 min at RT. Lastly, for visualization, sections were treated with 3,3′-diaminobenzidine by using an ImmPACT™ DAB Peroxidase substrate kit (Vector Laboratories) according to the manufacturer's instructions, and the sections were counterstained with hematoxylin. Control sections were treated in the same manner, except that they were not incubated with primary antibodies.
Statistical analysis
Statistical analysis was performed by using the IBM SPSS Statistics software (version 23.0; IBM Japan, Tokyo, Japan). All experiments were repeated under at least three independent conditions, with three replicates each. Statistical differences were determined by using two-way ANOVA with Bonferroni's correction. P < 0.05 was considered statistically significant.
Results
Morphological appearance and proliferation ability of DF and PDL cells isolated using EZ or OG procedure
When cells were isolated by using EZ and OG cell-isolation methods, spindle-shaped adherent cells were observed in both DF- and PDL-derived cultures. After 1–2 weeks, both DF-EZ and PDL-EZ cells formed adherent clonogenic cell clusters (Fig. 1A). The cells obtained by using the OG procedure started to grow out from the plated DF or PDL tissue fragments at approximately days 2–4 and then proliferated actively (Fig. 1B). The results of cell-proliferation tests showed that DF-derived cells (DF-EZ or DF-OG) exhibited significantly higher growth than did the respective PDL-derived cells (PDL-EZ or PDL-OG) after 8 days of culture (P < 0.01; Fig. 1C). However, growth did not differ significantly between DF-EZ and DF-OG cells or between PDL-EZ and PDL-OG cells during the cultivation period (Fig. 1C). The PDTs of all four cell types were ∼27 h, which agrees with our previous report [27].
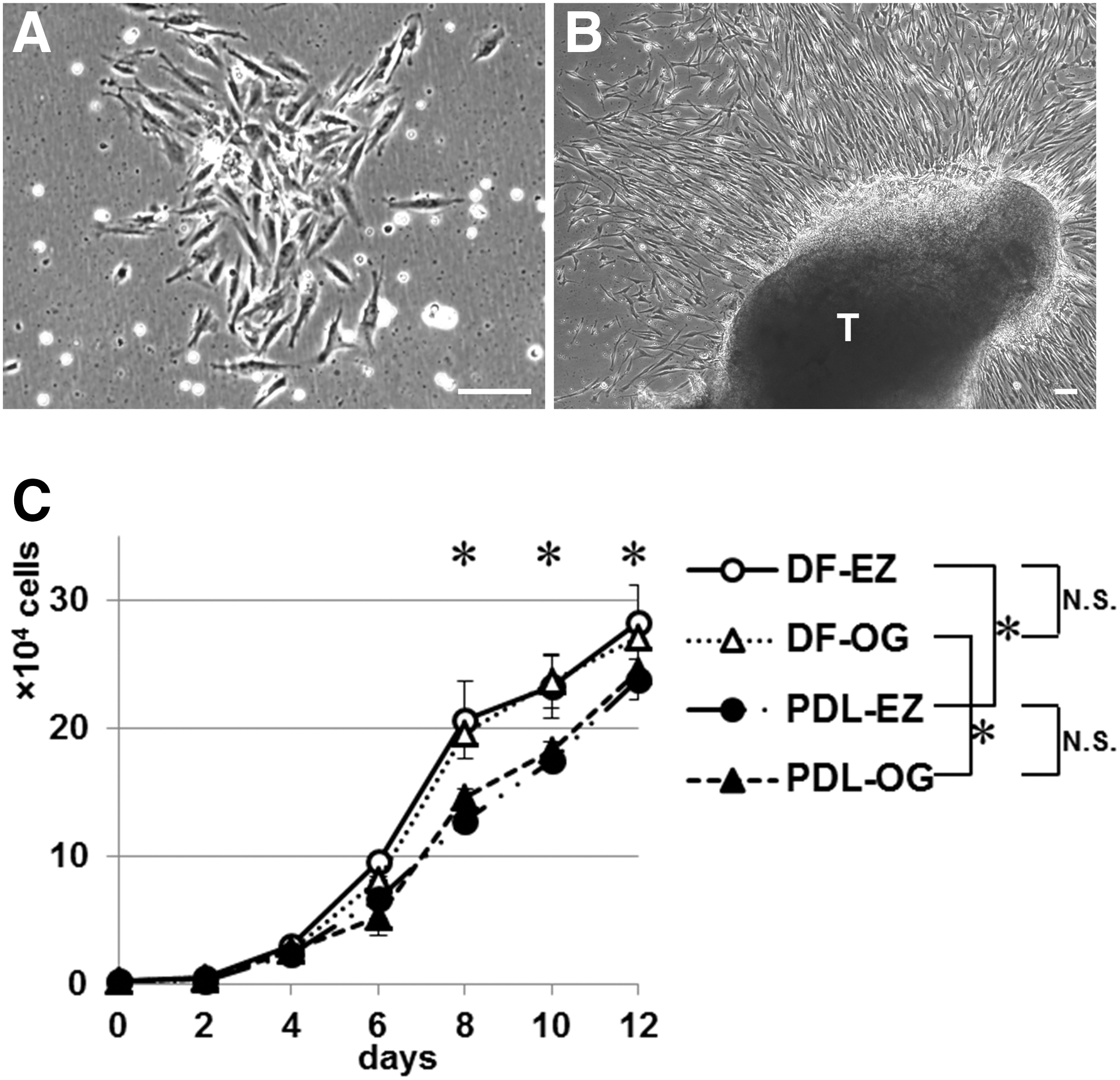
Morphological appearance and proliferation abilities of DF-EZ, DF-OG, PDL-EZ, and PDL-OG cells.
Cell-surface marker expression
We used flow cytometry to examine cell-surface markers. All four cell types were predominantly positive for CD29 (86.9%–99.9%), CD44 (90.9%–99.9%), and CD90 (86.1%–100%), which are typical mesenchymal stromal-cell markers. By contrast, the four cell types showed almost no expression (0%–2.7%) of CD14, CD31, and CD34, which are hematopoietic-cell markers. Furthermore, all four types of cells expressed cell-surface antigens such as CD105, CD106, CD146, and CD166, which are associated with the MSC phenotype, but these markers were all expressed at higher levels in the EZ-isolated cells than in the OG-isolated cells (Fig. 2). Similarly, the expression of CD140b, CD146, and NG2, which are typical pericyte markers, was markedly higher in the EZ-isolated cells than in the OG-isolated cells (Fig. 2).

Flow cytometric analysis of cell-surface antigens on DF-EZ, DF-OG, PDL-EZ, and PDL-OG cells. All cell types expressed CD105, CD106, CD146, and CD166, which are associated with the MSC phenotype, but the expression in OG-isolated cells (DF-OG and PDL-OG) was lower than that in EZ-isolated cells (DF-EZ and PDL-EZ). CD140b, CD146, and NG2, which are typical pericyte markers, were expressed at markedly higher levels in the EZ-isolated cells than in the OG-isolated cells. Representative results from three independent experiments are shown. MSC, mesenchymal stem cell.
Gene-expression profiles obtained using RT-PCR
Global gene-expression profiles in DF-EZ, DF-OG, PDL-EZ, and PDL-OG cells were analyzed by using conventional RT-PCR (Fig. 3). All four cell types expressed typical osteogenic markers (Runx2 and Osterix), periodontium-related markers (Type I collagen, Periostin, GDF5, Scleraxis, and Tenascin-N), and pluripotent stem cell markers (Nanog, Oct3/4, and Sox2). Notably, Scleraxis and Tenascin-N, which are specifically expressed in mature PDL tissue, were expressed at higher levels in PDL-derived cells than in DF-derived cells.
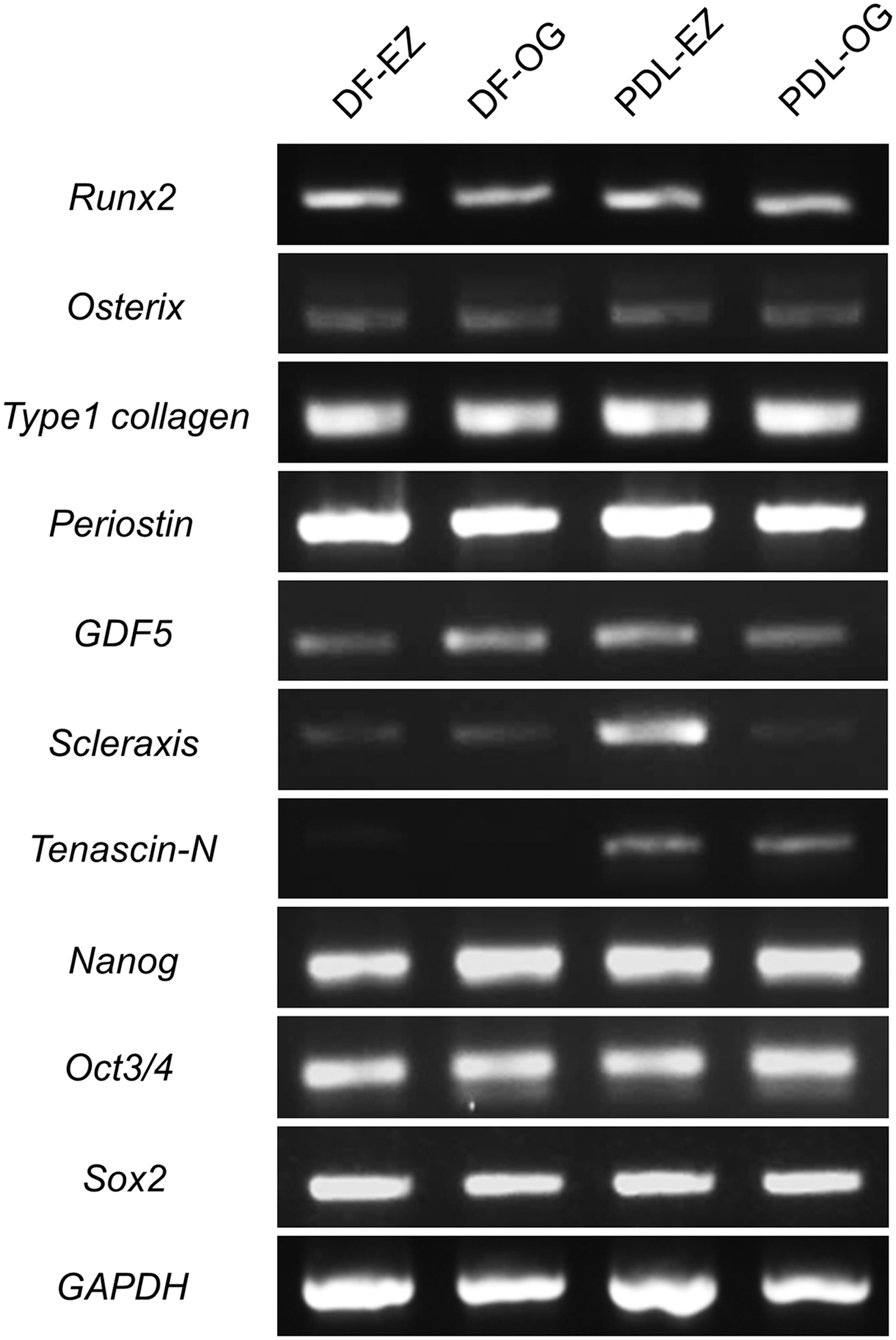
Gene-expression profiles in DF-EZ, DF-OG, PDL-EZ, and PDL-OG cells, determined by using reverse transcription-polymerase chain reaction. All cell populations express typical markers of osteogenic cells (Runx2 and Osterix), periodontal-lineage mesenchymal cells (Type I collagen, Periostin, GDF5, Scleraxis, and Tenascin-N), and pluripotent stem cells (Nanog, Oct3/4, and Sox2). Moreover, PDL-derived cells (PDL-EZ and PDL-OG) express Scleraxis and Tenascin-N, which are expressed in mature PDL tissue, at a higher level as compared with DF-derived cells (DF-EZ and DF-OG).
In vitro multidifferentiation into adipocytes and chondrocytes
After adipogenic induction for 3 weeks, oil red O-positive lipid vacuoles were observed in all four cell types (Fig. 4A–D), but not in control cells (Fig. 4A–D, insets). After chondrogenic induction for 4 weeks in pellet cultures, all four cell populations formed alcian blue- and safranin-O-positive cell spheres (Fig. 4E–H and I–L). Moreover, immunohistochemical analysis revealed that all four cell types were stained with anti-type II collagen antibody (Fig. 4M–P).

In vitro multilineage differentiation of DF-EZ, DF-OG, PDL-EZ, and PDL-OG cells. Oil red O-staining showing lipid clusters in DF-EZ
In vitro cementoblast characteristics under mineral-inducing or GM culture condition
In addition to multidifferentiation into adipogenic and chondrogenic cells, cementogenic differentiation of the four cell types was induced by a widely used mineral-inducing (osteogenic/cementogenic) medium, which was supplemented with Dex, β-GP, and AP. After a 3-week cementogenic-differentiation culture, alizarin red S-positive mineralized materials were observed in all cell types (Fig. 5A), whereas no positive staining was observed in control cultures (Fig. 5A, insets). Intriguingly, the PDL-OG cells showed a distinct pattern of deposition of the mineralized material: Nodular deposition exhibiting variable size and density was evident (Fig. 5A, PDL-OG, left). This nodular formation of mineralized materials by PDL-OG cells differed from the mineralized deposits detected in the other three cell populations (Fig. 5A, DF-EZ, DF-OG, and PDL-EZ). Moreover, in the PDL-OG cultures, a large mineralized nodule was frequently observed, and, notably, within this large nodule, an alternating lamellar pattern and incremental lines, resembling the structure of the cellular intrinsic fiber cementum (CIFC), could be observed (Fig. 5A, PDL-OG, right).
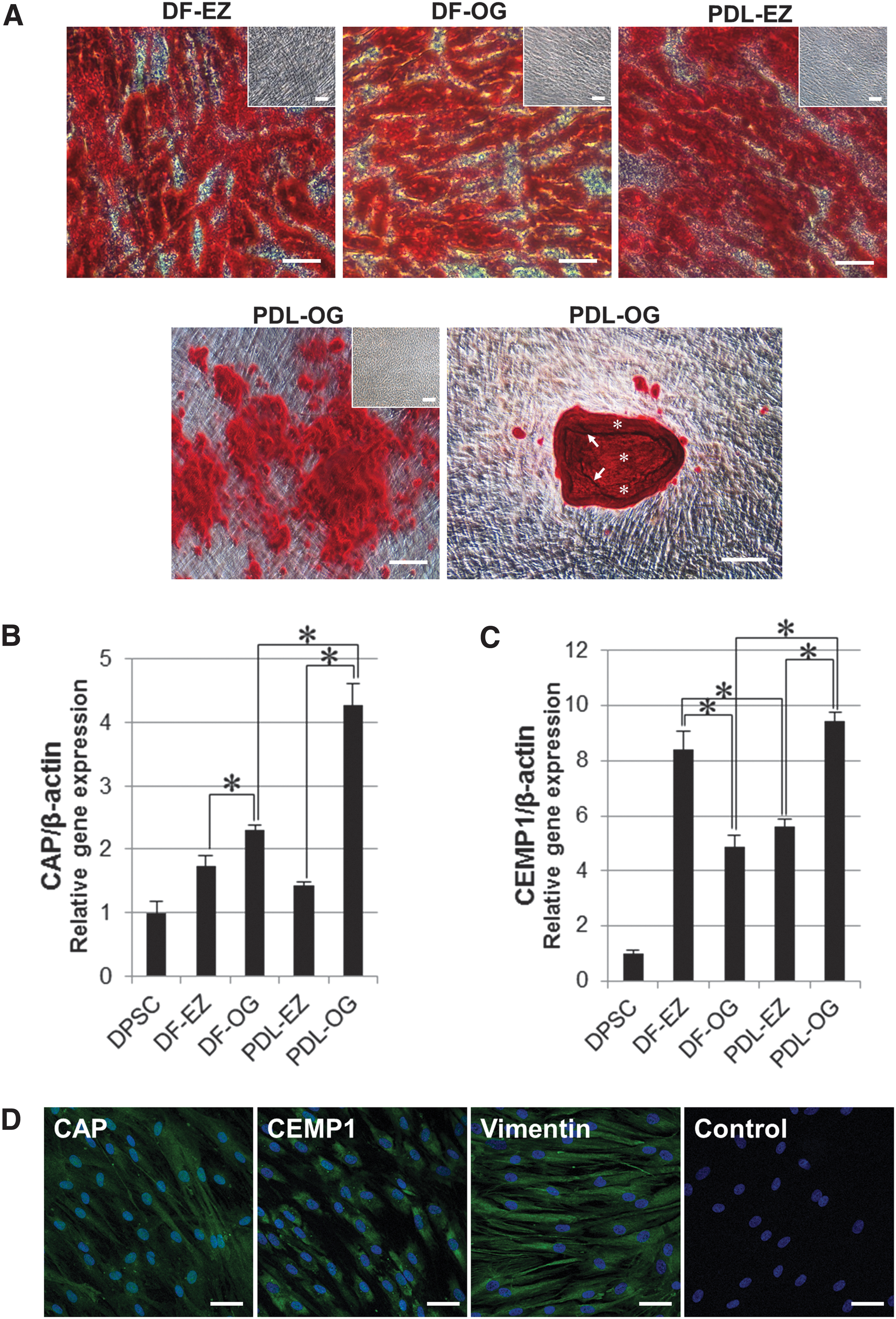
In vitro cementoblast characteristics under mineral-inducing culture conditions in DF-EZ, DF-OG, PDL-EZ, and PDL-OG cells.
To further examine the biological features of cementoblasts in the four cell types, we performed RT-PCR to detect the expression of cementoblast-specific marker genes under the GM culture condition. The results indicated that all four cell types endogenously expressed CAP and CEMP1 (Supplementary Fig. S1; Supplementary Data are available online at
In vivo cementogenesis evaluation
The capacity of the four cell types to generate the cementum hard tissue in vivo was examined by transplanting cell/HA constructs into the dorsal subcutaneous tissue of immunodeficient mice. In histological evaluation, the success of de novo cementogenesis was established in all cell transplants by staining with HE (Fig. 6B–E) and MT (Fig. 6G–J); by contrast, no hard-tissue formation was observed in the case of the control scaffold material alone (Fig. 6A, F). Immunohistochemical analysis revealed that the newly formed cementum hard tissue in cell transplants was entirely positive for OPN (Fig. 6Q–T), CAP (Fig. 6V–Y), and CEMP1 (Fig. 6a–d), in addition to being positive for human-specific vimentin in the surrounding fibroblasts (Fig. 6L–O). However, none of the antibodies stained the fibrous tissue and fibroblasts in the control transplants (Fig. 6K, P, U, Z, and e). The immunoreactivity for OPN, CAP, CEMP1, and vimentin was confirmed in the native PDL tissue (Supplementary Fig. S2): Vimentin was immunostained in cementoblasts and fibroblasts, which were located in the PDL connective tissue; OPN was predominantly immunostained within the cementum matrix and the connective tissue; CAP was detected weakly in cementoblasts and fibroblasts, in addition to within perpendicularly inserted PDL fibers, called Sharpey's fibers, in the cementum matrix; and CEMP1 was strongly immunostained in cementoblasts at the surface of the root cementum. This immunoreaction pattern agrees with the findings obtained for the cell/HA transplants (Fig. 6L–O, Q–T, V–Y and a–d), indicating that the four cell types derived from DF and PDL tissue possess the capacity of de novo cementogenesis in vivo.
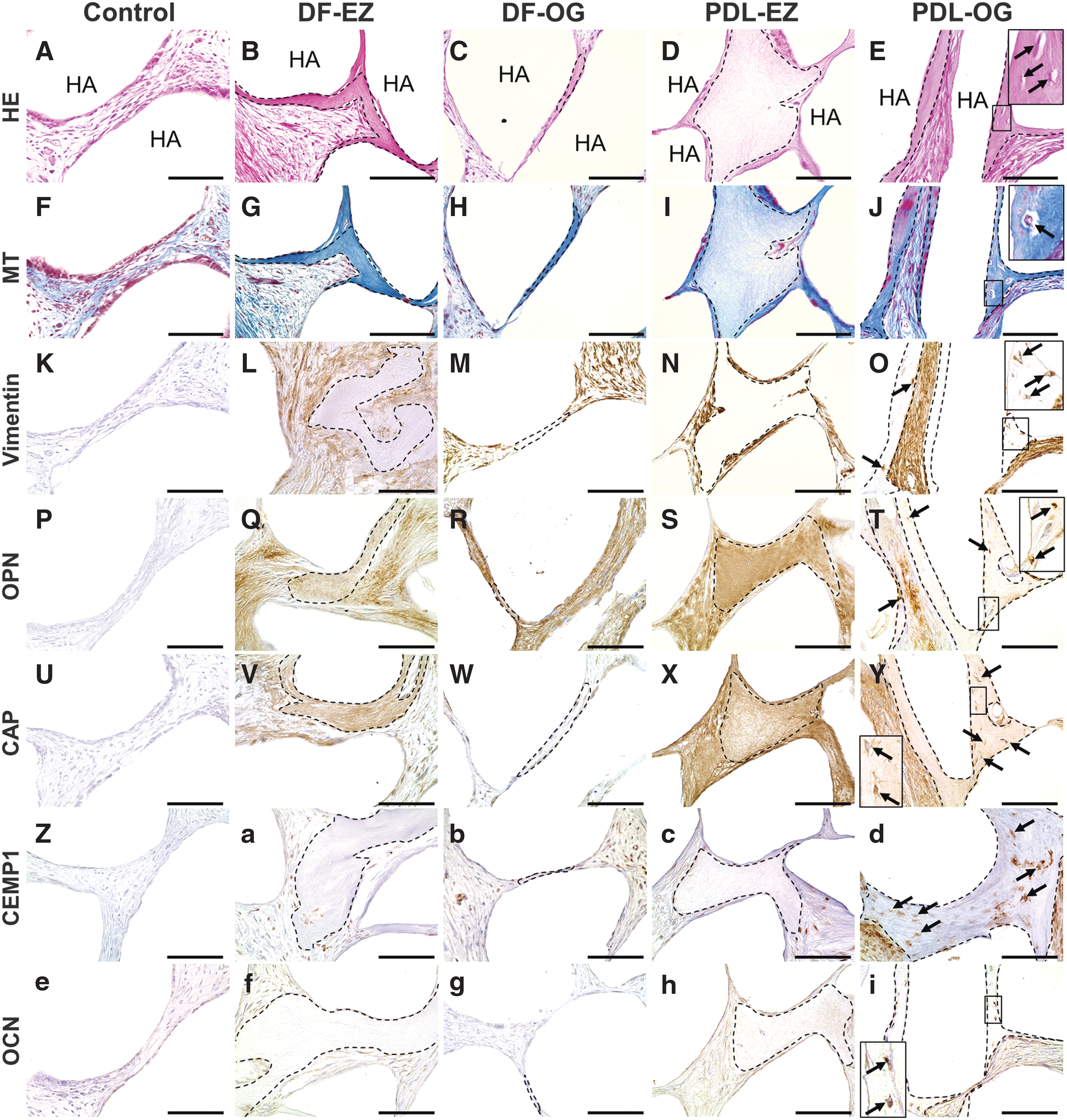
Histological and immunohistochemical evaluation of cell/HA transplants from SCID mice after implantation with EZ/OG-isolated cells. HE (
With regard to the histological structure of the newly formed cementum, the DF-EZ, DF-OG, and PDL-EZ transplants predominantly exhibited newly formed cementum hard tissue that lacked embedded cells, which is considered to be the acellular cementum (Fig. 6B–D and G–I). Notably, new cementum tissue containing embedded cells within the matrix, which is regarded as the cellular cementum, was observed only in PDL-OG transplants (Fig. 6E, J). Immunohistochemical analysis further revealed that staining for OCN (a late marker of cementogenesis) was present in the embedded cells only in the PDL-OG transplants (Fig. 6i). This anti-OCN antibody staining is consistent with the observation that mesenchymal cells are located in normal PDL tissue and alveolar bone: Mature cementoblasts and osteocytes embedded within the bone matrix were strongly stained by the anti-OCN antibody (Supplementary Fig. S2M, N). Although de novo cementogenesis was evident in transplants with all four cell types, the amount of new cementum formed by DF-OG cells was considerably lower than that formed by the other three cell types (Fig. 6C, H).
Discussion
Human extracted teeth are valuable cell/tissue samples for performing life science research, such as investigations in stem cell biology and developmental biology, as well as for developing novel cell-based therapies. This is because human teeth can be obtained by using simple procedures and they provide tissues at a variety of developmental stages, including developing tissues (deciduous teeth or immature wisdom teeth together with the surrounding DF tissue) and developed tissues (mature wisdom teeth). For the aforementioned types of research, no other human tissue/organ can be obtained at both incompletely developed (immature) and completely developed (mature) stages. In the case of dental research focused on cell-based therapies, a major aim is to identify the source of cells that is the most useful for repairing the root defect caused by external apical-root resorption. However, for inducing the de novo cementum formation that is required for root-defect repair/regeneration, the specific periodontal-lineage (DF and PDL) cells and the cell-isolation methods that are the most suitable have remained unknown. In this study, we isolated four types of adherent mesenchymal stromal cells derived from human DF and PDL tissues by using both EZ and OG isolation procedures in primary culture. Next, we characterized the stem cell characteristics of these cells by using “gold-standard” tests to evaluate their proliferative ability, gene/protein expression phenotype, and multidifferentiation potential. Lastly, we examined both the in vitro and the in vivo cementogenic potential of the periodontal-lineage MSCs by culturing them under GM and mineral-inducing conditions and by performing transplantation of the cells in SCID mice.
We found that DF-EZ, DF-OG, PDL-EZ, and PDL-OG cells could not be readily distinguished morphologically: they all exhibited the typical spindle-shaped fibroblast-like morphology (Fig. 1A, B). PDT calculation and growth-curve evaluation revealed that the PDT of all four cell types was short (∼27 h) and that the cells grew very well, with the DF-derived cells exhibiting higher proliferation than the PDL-derived cells (Fig. 1C). These findings agree with previous reports [27,36] and suggest that the DF-derived cells, which were obtained from teenagers' developing teeth containing immature roots, are more active and vital than the PDL-derived cells, which were collected from the mature teeth of adults. Moreover, the distinct cell-isolation methods (EZ and OG procedures) did not affect the growth of DF- or PDL-derived cells (Fig. 1C); similar findings have been reported regarding the growth of dental MSCs obtained from deciduous teeth [32].
To examine the phenotype of cell-surface markers, we performed flow cytometric analysis. All four cell types were positive for typical MSC markers such as CD105, CD106, CD146, and CD166, as well as for CD29, CD44, and CD90, which are also expressed in mesenchymal stromal cells; this agrees with the results of previous studies [37,38]. By contrast, all cell types were negative for the hematopoietic-cell markers CD14, CD31, and CD34, which suggests that our isolated cells represented MSC populations and were not contaminated with blood cells. PDLSCs isolated by using the EZ method were reported to express pericyte-marker antigens such as CD146, NG2, and CD140b, and PDLSCs were found to adhere to endothelial-cell capillary networks in a co-culture system and, thus, mimic pericyte-like perivascular localization [39]; these findings suggest that EZ-isolated PDLSCs possess pericyte-like properties. Therefore, we examined the four cell types for the pericyte-marker antigens CD140b, CD146, and NG2. The results indicated that the expression of these antigens in EZ-isolated cells (DF-EZ and PDL-EZ) was markedly higher than that in OG-isolated cells (DF-OG and PDL-OG) (Fig. 2). Given that we compared two cell-isolation methods for obtaining cells derived from two different dental tissues, our results strongly suggest that the pericyte-like cell population can be homogenously enriched by using the EZ-isolation method rather than the OG-isolation method when preparing primary cultures. By contrast, the OG-isolated cells are likely to present a heterogeneous cell population that includes a certain number of pericyte-like cells, the stem cell compartment, and the progenitor cells and committed cells of cementoblast lineage; the cementoblastic lineage-committed/progenitor cells would contribute to the unique cementogenic property of PDL-OG cells, as discussed later in this section.
The global gene-expression profiles of the isolated periodontal-lineage cells were revealed by using conventional RT-PCR (Fig. 3). All cell types endogenously expressed Runx2, Osterix, Type I collagen, Periostin, and GDF5, which are specific markers of osteogenic/periodontal stromal cells. Furthermore, Scleraxis and Tenascin-N, which are genes that are more specifically expressed in mature PDL tissue [40], were more highly expressed in PDL-EZ and PDL-OG cells than in DF-EZ and DF-OG cells. The gene-expression pattern suggests that, based on the developmental origin of the harvested tissues, a larger fraction of the PDL-derived cells than of the DF-derived cells can be considered to be strongly committed to the PDL lineage, regardless of the cell-isolation method used. Moreover, all four cell types expressed Nanog, Oct3/4, and Sox2, which are widely recognized transcription factors that are involved in maintaining pluripotency in cells such as embryonic stem cells and induced pluripotent stem cells [41,42]. These findings suggest that the isolated cell populations contain a small proportion of MSCs exhibiting high multipotency and plasticity. The aforementioned phenotype contributed to the multidifferentiation potential of the cells and enabled them to differentiate into adipogenic and chondrogenic lineages after the corresponding cytodifferentiation-induction cultures (Fig. 4).
In addition to the ability of the isolated cells to “multidifferentiate” into adipogenic and chondrogenic lineages, the cementogenic potential of the four types of cells was examined by preparing the widely used mineral-inducing culture. After a 3-week cytodifferentiation culture, all cell types produced alizarin red S-positive mineralized materials (Fig. 5A). Notably, our results demonstrated for the first time that PDL-OG cells showed nodular accumulation of mineralized materials, particularly of large mineralized nodules, which were frequently observed in PDL-OG differentiation cultures and displayed an alternating lamellar pattern and incremental lines within the deposits (Fig. 5A); this resembles the unique structure of the CIFC observed in vivo [43,44]. These findings indicate that PDL-OG cells, but not the other three cell types, exhibit unique cementogenic properties under in vitro cementogenic-differentiation culture conditions.
To further investigate the innate cementogenic potential of DF- and PDL-derived cells isolated by using the two distinct methods, we used conventional RT-PCR to analyze the endogenous expression of two cementoblast-related genes, CAP and CEMP1, under GM culture conditions. CAP is a collagenous protein that promotes the attachment of fibroblasts and cementoblastic progenitor cells in the PDL and plays a critical role in cementogenesis [45,46]. CEMP1, also named cementum protein-23, has been identified as a novel cementum-specific protein and is strongly expressed on cementoblasts and their progenitors [46,47]. All four periodontal-lineage cell types expressed CAP and CEMP1, and this expression of the two genes was considerably higher than that in dental papilla-derived DPSCs (Supplementary Fig. S1). Next, we used qPCR for quantitative gene-expression analysis after inducing cementogenic differentiation in culture for 3 weeks by using mineral-inducing culture medium. CAP and CEMP1 expression in all DF- and PDL-derived cells was higher than that in DPSCs (Fig. 5B, C), and the expression of these cementoblast-specific genes in PDL-OG cells was markedly higher than that in the other three cell types (Fig. 5B, C). Moreover, the results of immunofluorescent labeling indicated—at the protein-expression level—the successful cementogenic differentiation of all four cell types (Fig. 5D). These results support the strong cementogenic property of PDL-OG cells, which agrees with the unique ability of these cells to form nodular mineralized materials (Fig. 5A).
With regard to the cementogenic differentiation of periodontal-lineage MSCs, the underlying mechanisms that lead to the stem cell commitment and differentiation into either the cementoblast or osteoblast lineage remain unknown. Recently, CEMP1 was reported to preferentially regulate PDLSC commitment and differentiation into cementoblast lineage, but not osteoblast lineage [48]. CEMP1 was also reported to be capable of inducing the differentiation of nonosteogenic cells (gingival fibroblasts) into cementogenic/osteogenic cells featuring the respective phenotypes and the ability to deposit calcium [49]. Our results here indicated that the four types of periodontal-lineage MSCs endogenously expressed CEMP1 (Supplementary Fig. S1) and that this gene expression was highly upregulated after cementogenic differentiation (Fig. 5C). Notably, among the periodontal-lineage MSCs examined, the differentiated PDL-OG cells expressed CEMP1 at the highest level (Fig. 5C). These results suggest that CEMP1 contributes to the cementoblast commitment and differentiation into periodontal-lineage MSCs, and further that the highest level of CEMP1 expression confers the unique cementogenic property in PDL-OG cells, which comprise a heterogeneous PDL-committed cell population. In line with the results of in vitro gene/protein expression analyses, the findings of our in vivo experiments demonstrated the definitive cementogenic property of the cells, as described next.
To directly evaluate the in vivo ability of the isolated cells to generate hard tissue, the human DF- and PDL-derived cells were subcutaneously transplanted into immunodeficient mice. Our results indicated that all four types of cell transplants produced cementum-like hard tissue (Fig. 6B–E and G–J); the cells used in the transplants were unambiguously detected as cells that were positive for human-specific vimentin in the connective tissue (Fig. 6L–O). By contrast, the control transplants, which lacked the human cells, did not produce any hard tissue (Fig. 6A, F, and K). In terms of the histological observation of newly formed hard tissue, BMMSCs are widely recognized to form bone-like tissues featuring a lamellar pattern and including a bone marrow containing a considerable amount of blood cells [50,51]. The hard-tissue materials formed here by the DF- and PDL-derived cells lacked the lamellar pattern of the matrix itself and the hematopoietic marrows within the tissue, and this was clearly distinct histologically from what was observed with the native alveolar bone (Supplementary Fig. S2B, D). Conversely, the DF- and PDL-derived cell transplants exhibited a non-lamellar structure, very poor vascularity, and low cellularity, which are considered to represent a cementum-like appearance (Fig. 6B–E and G–J), and this is consistent with the findings of previous studies in which PDLSCs or the cementum-derived cells were transplanted in vivo [14,51]. Collectively, our results clearly indicate that cementum tissue was predominantly formed by the DF- and PDL-derived (periodontal-lineage) cells.
In our animal model study, the results of histological evaluation further demonstrated that the PDL-OG transplant produced new cementum containing cells embedded within the matrix itself, which appears similar to cellular cementum (Fig. 6E, J). Notably, cellular cementum-like tissue was not detected in any of the transplants with the other three cell types, all of which showed acellular cementum-like structures (Fig. 6B–D and G–I). To identify the properties of the cementum materials that were newly formed in vivo after transplantation, we performed immunohistological analysis by using antibodies against bone/cementum-specific proteins such as CAP, CEMP1, OPN, and OCN. The cementum materials formed by all cell transplants were entirely immunopositive for CAP (Fig. 6V–Y) and CEMP1 (a–d), the immunoreactivity for which was restricted to the cementum matrix and to other PDL-derived cells (Supplementary Fig. S2I–L). OPN is one of the noncollagenous proteins present in mineralized tissues, including the dental root cementum, and acellular cementum, in particular, contains a higher concentration of OPN than of other noncollagenous proteins [52,53]. In agreement with these features, OPN was intensely immunopositive in the acellular cementum-like materials in DF- and PDL-derived cell transplants (Fig. 6Q–S), except in the case of the PDL-OG transplant (Fig. 6T). OCN, which is a widely recognized terminal-differentiation marker of osteogenic/cementogenic cells [54], was immunopositive in the embedded cells within the cementum material of the PDL-OG transplant (Fig. 6i), and OCN immunoreactivity was detected in the mature cementoblasts and osteocytes present in the native PDL and alveolar bone, respectively (Supplementary Fig. S2M, N). By contrast, no OCN-positive cells were observed in the other three cell transplants (Fig. 6f–h). Collectively, the histological and immunohistochemical analyses revealed that the PDL-OG cells possess the unique de novo cementogenesis capacity for producing cellular cementum material, which is a clinically significant finding. In accord with this finding, in a previous study conducted by using a dog model, we demonstrated the potential of PDL-OG cells for future application in the regeneration of periodontal tissue defects through de novo cellular cementogenesis [55]. Clinically, dental-root defect, which is caused by external apical-root resorption, is recognized to be likely regenerated through the deposition of the cellular cementum [56]. Thus, the present study strongly suggests the feasibility of using PDL-OG cells for developing novel cell-based therapies. We expect that in future studies, transplantation of PDL-OG cells into the root defect will potentially be found to induce dental-root repair/regeneration through de novo cellular cementum formation.
In conclusion, although all four cell types exhibit active growth and MSC characteristics, the PDL-derived cells isolated by using the OG method present unique cementogenic properties and form cellular cementum-like hard tissue in vivo. Clinically, the development of regenerative medicine, that is, of PDL-OG cell-based therapy, might hold promise for root-defect repair/regeneration that is induced by mimicking the innate de novo formation of the cellular cementum.
Footnotes
Acknowledgments
The authors thank Dr. Daisuke Torii for technical assistance with RT-qPCR analysis. This work was supported by a Grant-in-Aid for Young Scientists (A) (no. 24689073 to T.N.) and Grants-in-Aid for Scientific Research (B) (nos. 15H05046 to T.N. and 25293422 to N.S., T.N.) from the Japan Society for the Promotion of Science (JSPS KAKENHI), and it was supported in part by a research grant (2013–2014 to T.N.) from Nippon Dental University. This study is based on a thesis submitted by RSO to the Graduate School of Dentistry, Meikai University, in partial fulfillment of the requirements for the Doctor of Dental Surgery degree.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.