
Abstract
Mesenchymal stem cells (MSCs) are effective in treating several pathologies. We and others have demonstrated that hypoxia or hypoxia-inducible factor 1 alpha (HIF-1α) stabilization improves several MSC functions, including cell adhesion, migration, and proliferation, thereby increasing their therapeutic potential. To further explore the mechanisms induced by HIF-1α in MSCs, we studied its relationship with Notch signaling and observed that overexpression of HIF-1α in MSCs increased protein levels of the Notch ligands Jagged 1–2 and Delta-like (Dll)1, Dll3, and Dll4 and potentiated Notch signaling only when this pathway was activated. Crosstalk between HIF and Notch resulted in Notch-dependent migration and spreading of MSCs, which was abolished by γ-secretase inhibition. However, the HIF-1-induced increase in MSC proliferation was independent of Notch signaling. The ubiquitin family member, small ubiquitin-like modifier (SUMO), has important functions in many cellular processes and increased SUMO1 protein levels have been reported in hypoxia. To investigate the potential involvement of SUMOylation in HIF/Notch crosstalk, we measured general SUMOylation levels and observed increased SUMOylation in HIF-1-expressing MSCs. Moreover, proliferation and migration of MSCs were reduced in the presence of a SUMOylation inhibitor, and this effect was particularly robust in HIF-MSCs. Immunoprecipitation studies demonstrated SUMOylation of the intracellular domain of Notch1 (N1ICD) in HIF-1-expressing MSCs, which contributed to Notch pathway activation and resulted in increased levels of N1ICD nuclear translocation as assessed by subcellular fractionation. SUMOylation of N1ICD was also observed in HEK293T cells with stabilized HIF-1α expression, suggesting that this is a common mechanism in eukaryotic cells. In summary, we describe, for the first time, SUMOylation of N1ICD, which is potentiated by HIF signaling. These phenomena could be relevant for the therapeutic effects of MSCs in hypoxia or under conditions of HIF stabilization.
Introduction
H
Recently, an interaction between the Notch1 receptor and the SUMOylation cascade has been described in breast cancer [23]. Small ubiquitin-like modifier (SUMO), a small (11 kDa) ubiquitin-like protein, can covalently attach to proteins as a post-translational modification and influence their stability, localization, or activity [24]. Notch1 signaling can deplete the unconjugated SUMO1 pool, which selectively impairs the growth of Notch1-activated cells. However, whether crosstalk between HIF-1, Notch1, and SUMO signaling pathways is involved in the control of MSC behavior is unknown. The aim of the present study was to examine the regulation of proliferation, migration, and spreading in MSCs through crosstalk between HIF-1α, Notch, and SUMO signaling pathways.
Materials and Methods
All experiments were carried out in accordance with the approved guidelines and approved by the Instituto de Salud Carlos III and institutional ethics and animal care committees.
Cell lines and lentiviral labeling
Human MSCs obtained from dental pulp were purchased from Inbiomed (Inbiobank, San Sebastian, Guipuzcoa, Spain). Cells were cultured in Dulbecco's modified Eagle's medium (DMEM)-low glucose (Gibco, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS; Lonza, Basel, Switzerland) and 1% penicillin/streptomycin (Gibco) in a humidified atmosphere of 95% air and 5% CO2 at 37°C. Lentiviral overexpression of HIF-1α was performed as described [25]. Briefly, cells were transduced with pWPI-green fluorescent protein (GFP) or pWPI-HIF-1α-GFP expression vectors (
Transient transfection of HEK293T cells
HEK293T cells (6 × 106) were cultured in 10-cm2 dishes with 10 mL of DMEM-high glucose supplemented with 10% FBS and 1% penicillin/streptomycin. Twenty-four hours after seeding and 1 h before transfection, the medium was replenished and supplemented with 25 μM chloroquine (Sigma-Aldrich, Madrid, Spain). Cells were transduced with a mixture of 20 μg of EF.hICN1.Ubc.GFP, 62 μL of 2 M CaCl2, and 418 μL of H2O added gently dropwise to 500 μL 2 × HBS (10 mM glucose, 40 mM HEPES, 10 mM KCl, 270 mM NaCl, 1.5 mM Na2HPO4, pH 7.1). After incubation for 6–8 h, the medium was replenished without chloroquine, and 2–3 days later, cells were analyzed by flow cytometry and western blotting [anticleaved Notch1 (A-8,1:1,000); Santa Cruz Biotechnology, Santa Cruz, CA) and real time quantitative PCR (RT-qPCR) (Hes1, Hey1, and Hey2].
Induction of hypoxia in HEK293T cells
Cells were cultured under hypoxia (1% O2) in an Invivo2 400 workstation (Ruskinn Technology Ltd., Bridgend, United Kingdom) for 4 h.
Activation/inhibition of Notch signaling pathway
To activate the Notch pathway, 60-cm2 culture plates were coated with anti-human IgG-Fc (stock at 1.3 mg/mL, 7.4 μL/1.5 mL phosphate-buffered saline [PBS]; Jackson ImmunoResearch Laboratories, Inc., West Grove, PA) for 30 min at 37°C, washed with PBS twice, and blocked with DMEM plus 10% FBS for 1 h at 37°C. Plates were washed again twice with PBS and coated with Jag1-Fc for 2 h at 37°C (stock at 200 μg/mL, 8.1 μL from stock/1.5 mL PBS; R&D Systems, Abingdon, United Kingdom). Then, control or HIF-MSCs were seeded onto the coated plates for 24 h. In some experiments, the gamma-secretase inhibitor RO4929097 (Xcess Biosciences, Inc., San Diego, CA) was used to inhibit the Notch pathway (10 μM for 48 h; 24 h before seeding cells onto Jagged1-coated plates and repeated after seeding).
Inhibition of SUMOylation
To inhibit protein SUMOylation, cells were treated with 50 μM anacardic acid (AA; cat. #172050 Calbiochem, Merck Millipore, Darmstadt, Germany) for 4 h. AA is a cell-permeable ginkgolic acid analog that inhibits protein SUMO modification by selectively targeting SUMO-activating enzyme E1 and interfering with E1-SUMO intermediate formation. With the inhibition of this first step, subsequent reactions of conjugation and ligation of SUMO to the target proteins are abolished [26].
Migration assay
Cells were seeded in basal medium (DMEM plus 0.5% FBS) at 10,000 cells/cm2 in the top chamber of an 8-μm-pore Transwell chamber (BD Falcon, Bedford, MA). After overnight culture, 25 ng/mL of IL-1β was added to the bottom chamber for 8 h. DMEM plus 0.5% and 10% FBS were used as negative and positive controls, respectively. Nonmigrated cells were removed from the upper side of the membrane with a cotton bud and migrated cells were fixed with 70% cold ethanol for 10 min. The membrane was cut and placed in a glass slide with the bottom side upward, stained with DAPI, and migrated cells were counted.
Proliferation assay
Proliferation of MSCs was assessed by incorporation of 5′-bromo-2′-deoxyuridine (BrdU; Life Technologies, Madrid, Spain). Cells were cultured at 50–60% confluence on 11-mm diameter cover slides coated with Jagged 1 and BrdU was added at 10 μg/mL for 8 h. After this time, cells were washed with PBS and fixed with 70% cold ethanol for 10 min at 4°C. Slides were then washed three times with PBS and 1 N HCl was added for 30 min at 37°C. Subsequently, the acid was neutralized with borate buffer for 10 min at room temperature and, after five washes in PBS, slides were treated with blocking buffer (PBS, 10% normal goat serum, 0.01% Triton X-100) for 1 h at 37°C, followed by addition of an anti-BrdU antibody and incubation overnight at 4°C (1:100 rat anti-BrdU; Abcam, Cambridge, United Kingdom). After five washes, an Alexa Fluor 555 (Invitrogen, Carlsbad, CA) secondary antibody was added for 2 h at room temperature and positive nuclei were then counted.
Scratch-wound assay
To analyze spreading and invasiveness, cells were seeded in 6-well culture plates and, once confluent, a scratch was made on the monolayer with a p10 pipette tip. Debris was removed with 1 mL of growth medium. Invasiveness of the scratch was tracked using time-lapse microscopy and image analysis was performed with ImageJ software (NIH).
Real time quantitative PCR
RNA was extracted using TRIzol reagent (Invitrogen) and purified with the RNeasy Plus Mini Kit (Qiagen, Dusseldorf, Germany). RNA was quantified spectrophotometrically using a NanoDrop ND-1000 (NanoDrop Technologies, Wilmington, DE). cDNA was obtained by reverse transcription using M-MLV reverse transcriptase (Invitrogen). RT-qPCR was performed with the following human-specific sense and antisense primers: Hes1 5′-AAGAAAGATAGCTCGCGGCA-3′ (forward) and 5′-TACTTCCCCAGCACACTTGG-3′ (reverse); Hey1 5′-AGGTAATGGAGCAAGGATCTGC-3′ (forward) and 5′-CCCGAAATCCCAAACTCCGA-3′ (reverse); Hey2 5′-GGATTATAGAGAAAAGGCGTC-3′ (forward) and 5′-GTTTTTCAAAAGCAGTTGGC-3′ (reverse); and ACTB 5′-AGAGCCTCGCCTTTGCCGATCC-3′ (forward) and 5′-CATGCCGGAGCCGTTGTCGAC-3′ (reverse) and SYBR Green I 1 × LightCycler 480 SYBR Green I Master Mix (Roche Molecular Biochemical, Mannheim, Germany). Multiwell plates of 96 wells were run on a LightCycler 480 Instrument (Roche Diagnostics, Mannheim, Germany).
Subcellular fractionation
Cells were seeded onto Jagged1-coated plates to activate Notch signaling for 24 h in DMEM-low glucose supplemented with 10% FBS and 1% penicillin/streptomycin. Cells were treated (+) or not (
Immunoprecipitation assay
Cells were seeded onto Jagged1-coated plates to activate Notch signaling for 24 h in DMEM-low glucose supplemented with 10% FBS and 1% penicillin/streptomycin. Cells were lysed with 1 mL of ionic buffer (50 mM Tris-HCl pH 8, 150 mM NaCl, 0.5% NP40, 0.5% sodium deoxycholate, and 0.05% SDS) supplemented with protease inhibitor cocktail, phosphatase inhibitor, and 10 μM MG132 (all from Roche). Protein A/G Plus-agarose (Santa Cruz Biotechnology, Santa Cruz, CA) and cleaved Notch1 (Val 1744; Cell Signaling Technology) were used for immunoprecipitation. Agarose beads were washed several times with cold Tris buffer containing 1 mM Tris, pH 7.4, and 15 mM NaCl (TBS) and blocked with 3% bovine serum albumin in TBS. Whole lysates were precleared with agarose beads and then incubated overnight (rocking at 4°C) with cleaved Notch1 antibody. Whole cell lysates were added onto new pellets of agarose beads (previously blocked with bovine serum albumin) and incubated for 3 h in a rocking wheel at 4°C. Finally, the supernatant was removed and beads were washed several times with cold 1 × TBS-T (TBS supplemented with 0.05% Tween). Proteins were eluted with 2 × Laemmli buffer supplemented with dithiothreitol and denatured for 5 min at 99°C.
Electrophoresis and western blotting
Cells were washed with PBS and lysed in RIPA buffer (1.5 M NaCl, 10% Nonidet P-40, 500 mM Tris-HCl, 5% deoxycholate, and 1% SDS, pH 8) supplemented with protease inhibitors, PMSF and leupeptin (Roche). Lysis was completed by five freeze–thaw cycles, followed by centrifugation at 12,000 g for 10 min 4°C. Protein concentration determination and electrophoresis were as described above. Membranes were incubated overnight at 4°C with the following primary antibodies (1:1,000 dilution): anti-Jagged1, anti-Jagged2, anti-Dll1, anti-Dll3, and anti-Dll4 (all from Cell Signaling Technology Europe, Schuttersveld, Leiden, The Netherlands), anticleaved Notch1 (A-8; Santa Cruz), and anti-HIF-1α (ab1; Abcam). An antibody to tubulin (T5158; Sigma) was used as a loading control. Exposure to secondary antibodies and immunoreactivity were performed as described above.
Immunocytochemistry and confocal microscopy
Cells in DMEM-low glucose medium were seeded onto 11-mm diameter slides. After 2 days, the slides were washed twice with PBS and cells were fixed with cold 70% ethanol for 10 min at 4°C. After three washes with PBS, the slides were incubated for 1 h at 37°C with blocking buffer consisting of PBS with 10% newborn goat serum (Gibco) and 0.01% Triton X-100 (Pronadisa, Basel, Switzerland). Cells were overlaid with the primary antibodies anti-HIF-1-α (H1alpha67, 1:200; Abcam) or antiactivated Notch1 (1:200; Abcam) diluted in blocking buffer and PBS (1:1) and incubated overnight at 4°C. The slides were then rinsed five times with PBS and overlaid with the secondary antibodies, Alexa Fluor 555 goat anti-rabbit and Alexa Fluor 647 goat anti-mouse (both 1:1,000; Invitrogen). Immunocytochemistry of negative controls was performed to discriminate between autofluorescence and specific signals. Slides were analyzed by vertical microscopy with a × 63 immersion objective (Leica 0.3). Quantification of fluorescence was performed using ImageJ.
InCell Analyzer quantification
N1ICD nuclear translocation was measured by immunolabeling. Cells were cultured in 24-well plates in triplicate in DMEM with 10% FBS and 1% penicillin/streptomycin and then fixed with 70% cold ethanol for 10 min at 4°C. Cells were labeled with antiactivated Notch1 antibody (ab8925; Abcam). The secondary antibody used was Alexa Fluor-555 (Invitrogen) and DAPI was used for nuclear labeling. Images were acquired with an InCell Analyzer 1000 epifluorescence microscope (GE Healthcare, Cardiff, United Kingdom). A 20 × objective was used to collect the fluorescence signals, and a combination of two excitation (EX) and emission (EM) filters was applied to detect DAPI (EX 405 nm/EM 450 nm) and red fluorescence from antiactivated Notch1 (EX 530 nm/EM 620 nm). Fifteen fields were acquired for each well. Analysis was performed with the InCell Analyzer 1000 Workstation software. Cells were first defined using the nuclear segmentation based on DAPI. The red signal was used to define N1ICD localization in the cell.
Statistical analysis
Data are expressed as mean ± standard error of mean or standard deviation as indicated. Comparisons were performed using unpaired Student's t-test. Statistical values were calculated using GraphPad Prism 6 software. Differences were considered statistically significant at P < 0.05 with a 95% confidence interval.
Results
HIF-1α overexpression induces protein and gene expression changes in MSCs
MSCs were transduced with pWPI-GFP (hereafter named MSC) or with pWPI-HIF-1α-GFP (hereafter named HIF-MSC) lentiviral vectors (Fig. 1A) as described [25] and cultured in normoxia. After infection, >95% of cultured cells were GFP positive (Fig. 1B). HIF-1α overexpression was confirmed by qPCR analysis (Fig. 1C) and western blotting (Fig. 1D), and confocal microscopy analysis showed nuclear localization of HIF-1α in HIF-MSCs (Fig. 1E).

HIF-1α overexpression in MSCs.
HIF-1α overexpression in MSCs increases Notch signaling activation
Crosstalk between HIF and Notch signaling has been recently described in some cancer cell lines [16,27], in which Notch relays the hypoxia signal into increased metastatic potential. To address whether a similar system was operating in MSCs, we assessed Notch signaling components in MSC overexpressing HIF-1α and cultured on Jagged1-coated plates at low confluence to prevent Notch activation by cell–cell contact. The mRNA expression levels of the Notch downstream targets, Hes1, Hey1, and Hey2, were significantly increased by Jagged1 stimulation both in MSCs (black bars) and in HIF-MSCs (gray bars), and this increase was higher in HIF-MSCs than in MSCs (Fig. 2A), suggesting a cooperative effect between HIF-1α and Notch signaling. Notch signaling was abolished by treatment with RO4929097, a selective gamma-secretase inhibitor that impairs Notch receptor processing and generation of NICD (Fig. 2A). To evaluate whether MSCs expressed Notch receptor ligands on the cell surface and whether these ligands were functional, we performed western blotting on MSCs and HIF-MSCs. Results showed that the Notch ligands Jagged1, Dll1, Dll3, and Dll4, but not Jagged2, were all expressed in MSCs and all were significantly increased in cells expressing HIF (Fig. 2B). To test if the ligands were functional and could activate Notch, we seeded HIF-MSCs and MSCs at confluence onto plates in the absence of Jagged1 and analyzed the expression of Notch downstream genes. Under these conditions, the expression of Hes1 and Hey2, but not Hey1, was significantly higher in HIF-MSCs than in MSCs (Fig. 2C). These results indicate that Notch signaling is activated in confluent HIF-MSCs and that more than one Notch ligand could be implicated in the process.

HIF-1 enhances Notch signaling in MSCs.
Notch mediates migration and spreading in HIF-MSCs
We have previously shown that HIF overexpression in bone marrow-derived MSCs stimulates their migration and invasion [25]. Given the well-documented role of Notch in migration and invasion in homeostasis and cancer [20,28], we investigated whether it participated in HIF-mediated spreading of MSCs. Overexpression of HIF-1α in MSCs significantly increased invasion, as measured by the scratch-wound assay, and promoted the development of lamellipodia and filopodia (Fig. 3A). The addition of RO4929097 to cultures blocked HIF-α-mediated invasion, which was similar to that of control MSCs (Fig. 3A). Likewise, the HIF-mediated increase in migration of HIF-MSCs through a gradient of IL-1β in the Transwell assay was abrogated by treatment with RO4929097, pointing to the participation of NICD in this process (Fig. 3B).
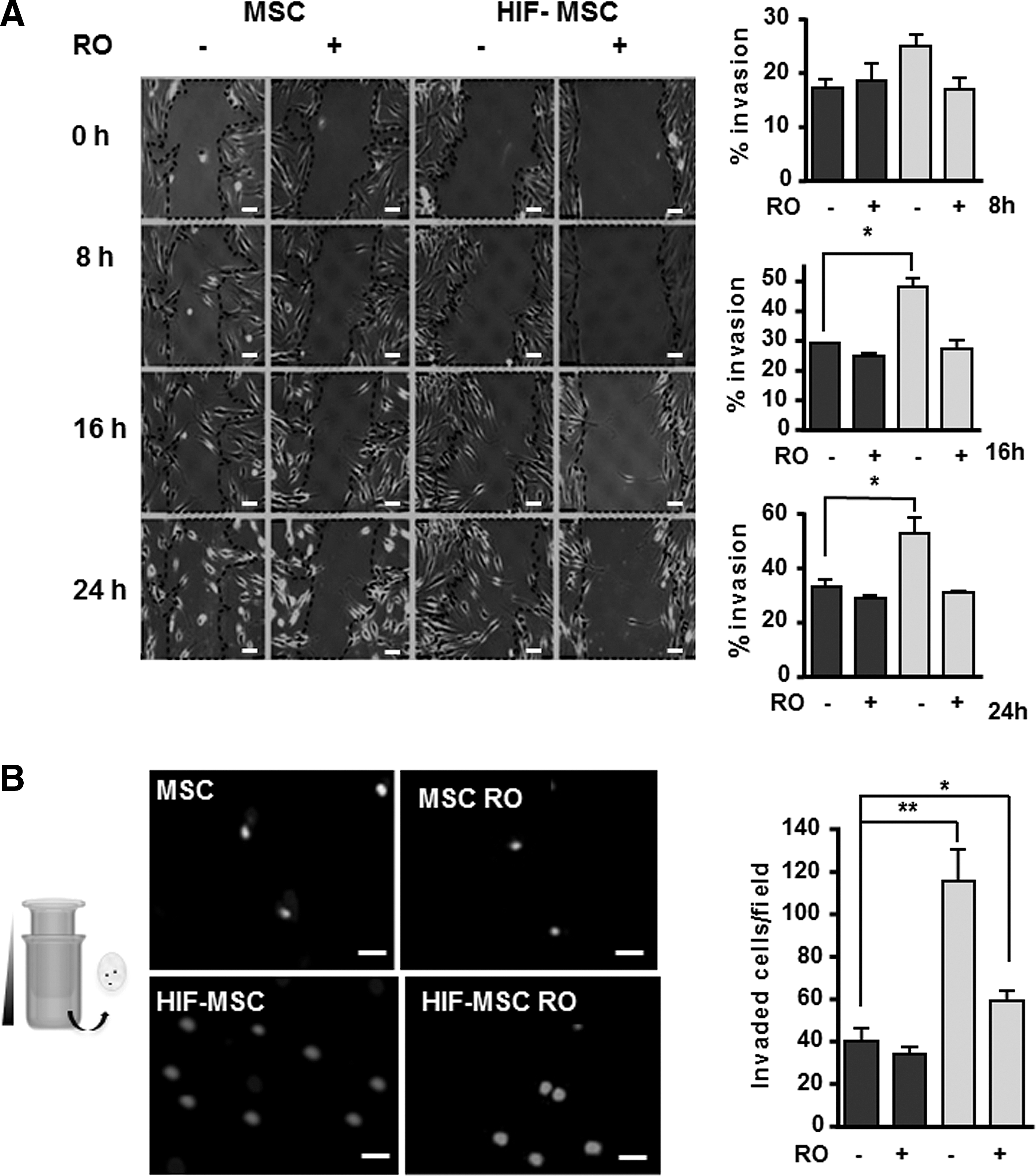
Increased migration of HIF-MSCs is dependent on Notch pathway activation.
HIF-mediated cell proliferation is dependent on SUMO, but not Notch
We next sought to evaluate the influence of Notch signaling for MSC proliferation. HIF-MSC and MSC cultures were seeded onto Jagged 1-coated plates to ensure Notch pathway activation and were incubated with BrdU for 8 h in the presence or absence of RO4929097. Subsequently, fixed cells were immunostained with anti-BrdU antibodies. Consistent with our previous results using bone marrow-derived MSCs [25], HIF-1α overexpression increased MSC proliferation as shown by a significant increase in BrdU incorporation; however, this effect was not abolished by RO4929097, indicating that HIF-induced cell proliferation is independent of Notch (Fig. 4A).

Increased proliferation of HIF-MSCs is Notch independent, but SUMO1 dependent. MSCs and HIF-MSCs were treated or not with 10 μM RO4929097 (RO) for 48 h
It has been reported that a fine balance between SUMOylation/deSUMOylation is required for activation of the hypoxia-signaling cascade, possibly through post-translation modifications to HIF-1α [29]. To question the potential role of SUMO in HIF-1α-mediated proliferation in MSCs, we repeated the BrdU incorporation study using 50 μM AA, an inhibitor of SUMO ligase. As expected, cell proliferation was significantly higher in HIF-MSCs than in MSCs; however, the HIF-1α-mediated increase in cell proliferation was completely abolished by AA treatment, demonstrating the relevance of the SUMO pathway for HIF-1α-induced MSC proliferation (Fig. 4B).
HIF and Notch pathways are linked by SUMOylation
The above results suggested possible interplay between HIF, Notch, and SUMO pathways. To investigate these potential interactions, we first analyzed the influence of SUMO1 for the expression of Notch downstream genes. Treatment of MSCs and HIF-MSCs with AA, under conditions of Notch pathway activation, led to a decrease in the levels of expression of Hes1, Hey1, and Hey2 in both groups, but the decrease was significant only for HIF-MSCs (Fig. 5A). Moreover, global levels of SUMO were higher in HIF-MSCs than in MSCs as measured by western blotting (Fig. 5B), suggesting that HIF increases SUMO protein levels. Consistent with previous studies, these results suggested an association between HIF and SUMO pathways. To explore this functional relationship, we first analyzed the protein sequence of the Notch1 receptor with SUMOplot™ Analysis software, a program that predicts and scores SUMOylation sites in protein sequences. The results of this analysis showed six potential motifs with high probability of representing the SUMO binding site on the N1ICD (not shown). We next immunoprecipitated N1ICD in MSCs and HIF-MSCs with an anti-N1ICD antibody and performed western blotting for SUMO1. We observed that N1ICD was susceptible to SUMOylation and, moreover, levels of SUMOylated N1ICD were higher in HIF-MSCs than in MSCs (Fig. 5C). Additional analysis using an InCell Analyzer to quantify N1ICD nuclear translocation [30] confirmed that levels of N1ICD were significantly higher in the nuclei of Notch-activated HIF-MSCs than in equivalent MSCs and also showed that AA abolished N1ICD nuclear translocation (Fig. 5D). Overall, these results indicate a functional relationship between SUMO and Notch1, whereby cellular SUMO levels can affect N1ICD nuclear translocation. To further corroborate SUMOylation of N1ICD, we performed subcellular fractionation experiments and western blotting using MSCs and HIF-MSCs cultured on Jagged 1-coated plates and treated or not with AA to modulate SUMOylation. Analysis of N1ICD expression in nuclear fractions revealed a reduction in nuclear expression of the protein in the presence of the SUMO inhibitor (Fig. 5E).
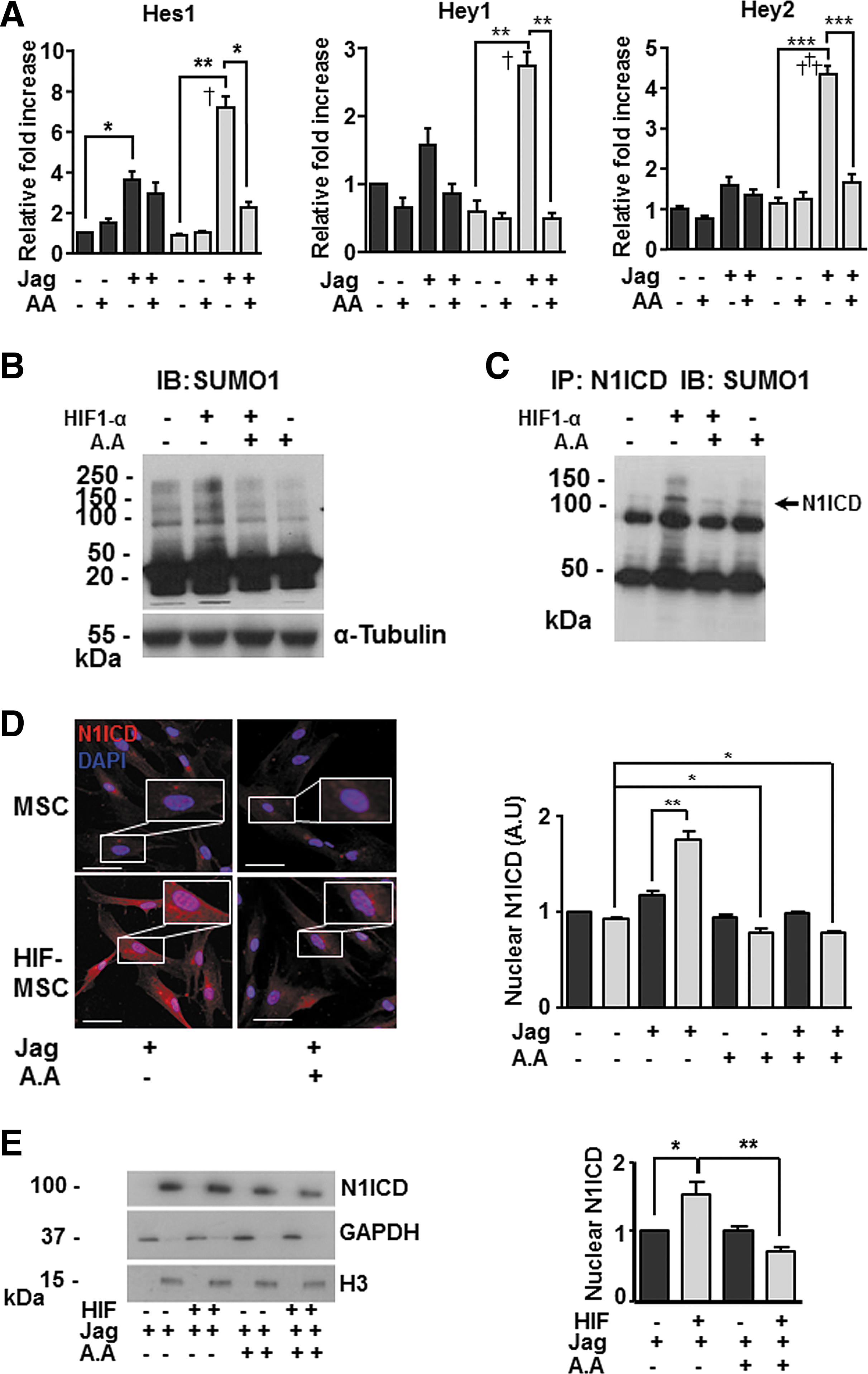
HIF-1α and SUMO1 increase nuclear translocation of N1ICD.
SUMOylation mediates HIF-induced Notch migration and invasiveness
To test the functional effect of SUMOylation for cell invasion, we repeated the scratch-wound assays in HIF-MSC and MSC cultures using AA to inhibit SUMOylation. Results showed that AA treatment completely abolished the increase in migration of HIF-MSC cultures (Fig. 6A, B). These results establish a close relationship between the three pathways.

HIF-1α-enhanced cell migration is mediated by SUMO signaling.
SUMOylation of N1ICD is increased by hypoxia in HEK293T cells
To validate the proposed model of N1ICD SUMOylation in other cell lines, we transiently transfected HEK293T cells with EF.hICN1.Ubc.GFP, a lentiviral vector encoding N1ICD (N1ICD-293T). Flow cytometry analysis showed that 95% of cells were infected with the vector. Subsequently, N1ICD expression was confirmed by western blotting using N1ICD antibodies and quantified by densitometry (Fig. 7A). In addition, genes downstream the Notch signaling pathway (Hes1, Hey1, and Hey2) were analyzed by RT-qPCR (Fig. 7B) and, as expected, were overexpressed in N1ICD-293T cells. Cells were then cultured under hypoxic conditions (1% O2) for 4 h in the presence or absence of AA (same conditions as used for MSCs) and expression of HIF-1α and N1ICD was evaluated by western blotting using anti-HIF-1α and antiactivated Notch1 antibodies and quantified by densitometry. As anticipated, hypoxia increased the steady-state levels of HIF-1α (Fig. 7C). Next, Hes1, Hey1, and Hey2 genes were analyzed by RT-qPCR and an increase in gene expression was observed in N1ICD-293T cells exposed to hypoxia. Moreover, when hypoxic N1ICD-293T cells were treated with AA, Hes1, Hey1, and Hey2 expression decreased significantly in comparison with the levels in N1ICD-293T cells in normoxia (treated or not with AA) (Fig. 7D). These results correlate with previous findings observed in MSCs, suggesting that the intracellular domain of the Notch1 receptor is SUMOylated, which increases its nuclear translocation, and that this SUMOylation is enhanced by hypoxia or by HIF-1α. Finally, to confirm SUMOylation of N1ICD, we performed immunoprecipitation of N1ICD in N1ICD-293T cells under normoxia/hypoxia and treated or not with AA. Protein extracts were immunoprecipitated with an anti-N1ICD antibody and SUMOylated proteins were detected using an anti-SUMO1 antibody. We observed that N1ICD was SUMOylated both in normoxia and hypoxia and that treatment with AA decreased this post-translational modification, especially under hypoxia (Fig. 7E).
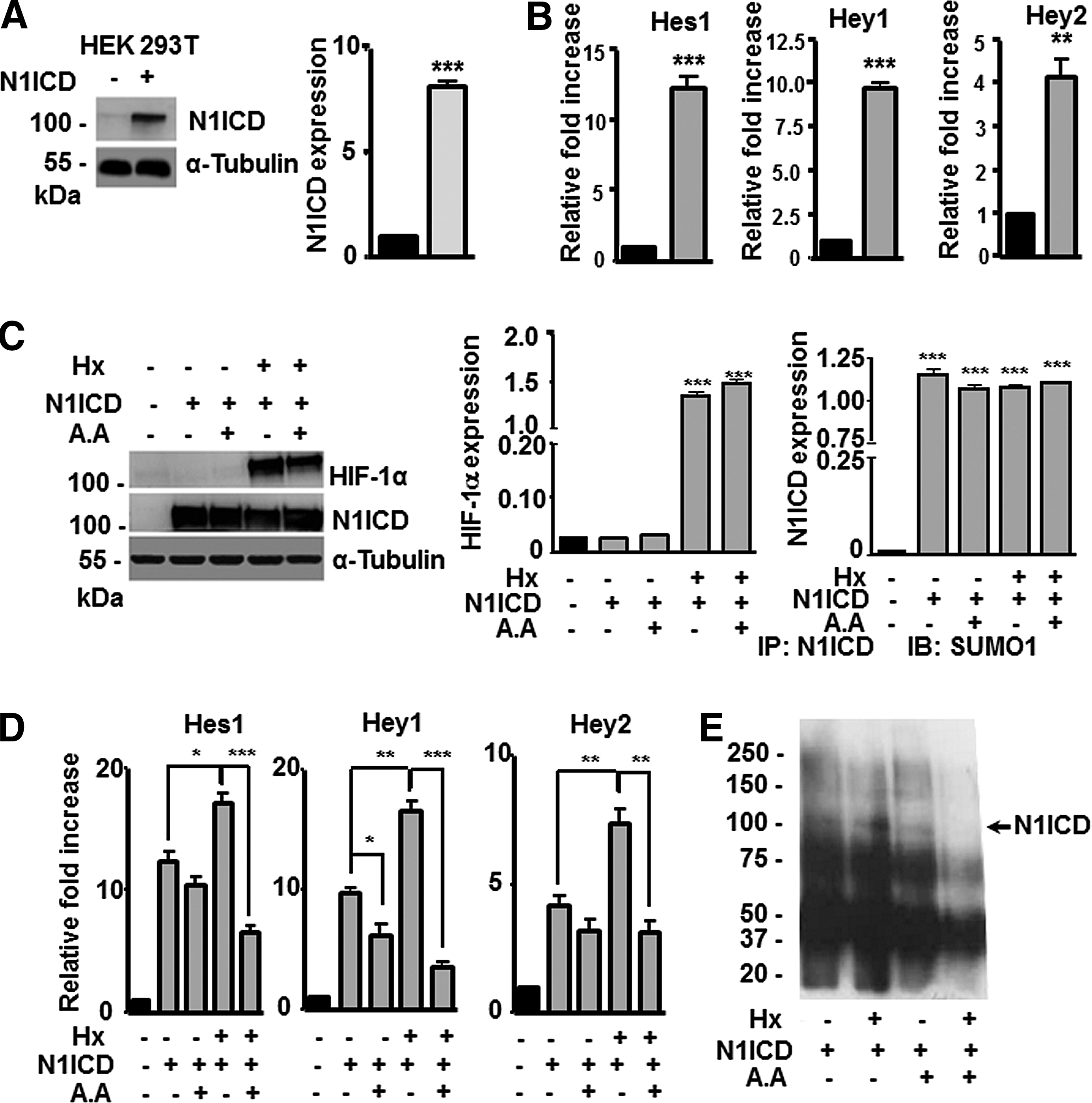
HIF-1α and SUMO1 increase nuclear translocation of N1ICD in HEK293T cells.
Discussion
MSCs can repair damaged tissues and we have previously demonstrated that infusion of MSC overexpressing HIF-1α promotes myocardial healing in an experimental rat model of myocardial infarction. As described by us and others, signaling pathways related to several paracrine factors and interleukins are upregulated in HIF-MSCs. For example, expression levels of p38 mitogen-activated protein kinase (p38/MAPK) [25,31], JNK/SAP [25], AKT [25], and Notch [32,33], together with fibronectin [34,35], fibroblast growth factor [25,36], angiopoietin-1 (ANGPT1) [37], NGF [38], and insulin growth factor [25,39] signaling pathways, are all induced by HIF-1α. We examined Notch signaling since hypoxia can promote Notch pathway activation and HIF-1α is a key mediator in the adaptation of eukaryotic cells to hypoxia. We first compared Notch downstream target gene expression in HIF-MSCs and MSCs. Activation of Notch signaling in HIF-MSCs was observed when HIF-MSCs were cultured at confluence or seeded at low density on Jagged1-coated plates, indicating that HIF-1α enhanced Notch signaling only after activation of the Notch receptor. This is in accordance with previous studies showing that hypoxia increases activation levels of Notch1 in different tumor cell lines only after coculture with a Jagged1-expressing cell line [27].
We next analyzed the contribution of the Notch pathway to HIF-MSC migration and proliferation since Notch is a major participant in these processes [16,27,40]. We found that migration and invasiveness, but not proliferation, were dependent on Notch signaling. The contribution of HIF-1α to enhanced Notch-dependent migration and invasiveness has been previously described in choriocarcinoma cell lines, suggesting that this is a general mechanism in normal and tumor cells [16]. However, the observation that increased cell proliferation in HIF-MSCs was Notch independent pointed to the participation of other mechanisms and led us to explore alternative pathways. In this regard, a relationship has been described between HIF-1α and the ubiquitin family member, SUMO-1 [41,42]. In contrast to ubiquitination, SUMO-1 modifications enhance the stability of nuclear proteins [43]. Previous studies have shown that HIF-1α undergoes post-translational modification by the three isoforms of SUMO (SUMO-1, -2, and -3) in vitro [44]. Increased SUMO-1 mRNA and protein level have been reported after hypoxic stimulation in various cell lines [45] and in vivo [42]. Moreover, SUMOylation of HIF-1α results in its stabilization and increases its transcriptional activity [41]. To investigate whether SUMOylation was involved in HIF/Notch crosstalk in MSCs, we cultured cells in the presence or absence of AA, a SUMOylation inhibitor. Under these conditions, we observed that both proliferation and migration were abolished and this effect was stronger in HIF-MSCs. Western blotting showed an increase in global SUMO levels in HIF-MSCs. Moreover, immunoprecipitation studies showed higher amounts of SUMOylated N1ICD in HIF-MSCs. Immunofluorescence labeling, followed by InCell quantification, showed decreased nuclear translocation of NICD in HIF-MSCs in the presence of AA, indicating that SUMOylation of N1ICD favors the nuclear translocation of this transcriptional factor and thus confirming that enhanced SUMO pathway by HIF-1α overexpression might contribute to the differential activation of the Notch pathway in MSC-HIF with respect to MSCs.
Several mechanisms may explain the increased SUMOylation of N1ICD in HIF-MSCs. First, it has been described that HIF-1α can increase γ-secretase activity, which would increase N1ICD cleavage leading to higher cytosolic concentrations [46]. Second, the global increase in cytosolic SUMO levels in HIF-MSCs could favor N1ICD SUMOylation. Third, SUMOylated N1ICD could be more easily translocated to the nucleus since it is known that protein SUMOylation promotes nuclear localization [30,47].
The effect of HIF-1α stabilization on MSCs is summarized in the model proposed in Fig. 8. Overexpression of HIF-1α leads to an increase in N1ICD levels by stabilization of the transcription factor, increasing its half-life, or by enhancing gamma-secretase enzyme activity [46,48,49]. Concurrently, HIF-1α increases the cellular amounts of SUMO1, which is reflected by an increase in the SUMOylation of N1ICD, a phenomenon described for the first time here. This process together with N1ICD nuclear translocation is important for cellular proliferation and migration, as well as for tumoral processes where the independent roles of SUMO, HIF, and Notch are well described. With this model, we are able to explain how HIF-1α acts as a potent enhancer of Notch transcriptional activity, enhancing the therapeutic properties of hMSCs through SUMO1. Moreover, we establish a link between these two highly conserved pathways and the key role of post-translational SUMO modification, which is implicated in tissue renewal, regeneration, proliferation, migration, invasiveness, and tumorigenesis.

Proposed model for HIF-1α, Notch, and SUMO crosstalk in hMSCs. Under normoxic conditions, HIF-1α undergoes ubiquitin–proteasomal-mediated degradation, while N1ICD is SUMOylated at a low level. Under hypoxic conditions or after HIF stabilization, levels of SUMO1 are higher and SUMOylation of proteins, including N1ICD, is elevated, which results in increased N1ICD translocation and increased Notch transcriptional activity, leading to enhanced cell migration and proliferation. SUMO1-increased levels by HIF-1α stabilization increased the amount of SUMOylated proteins, including N1ICD. RBPJ, recombining binding protein suppressor of hairless; MAML1: mastermind-like protein-1.
In summary, we demonstrate at the molecular and cellular levels the mechanism underlying HIF-SUMO-Notch crosstalk in hMSCs and how this interaction results in (i) autoactivation of Notch signaling pathway, (ii) increased Notch ligand expression, (iii) increased levels of SUMO and SUMOylation of N1ICD, (iv) increased N1ICD nuclear translocation, and (v) increased migration, invasiveness, and proliferation of HIF-MSCs. The relationship between the Notch pathway and cancer is well recognized [50]. Thus, this novel level of regulation for N1ICD could have potential implications in cancer pathogenesis. Further studies are needed to identify the residues SUMOylated in N1ICD and to elucidate whether higher concentrations of N1ICD by increased γ-secretase activity or higher SUMO-mediated nuclear translocation of N1ICD account for the enhanced Notch activity in HIF-MSCs.
Footnotes
Acknowledgments
This work was supported, in part, by grants from the Instituto de Salud Carlos III PI13/00414, PI16/0107, RETICS RD12/0019/0025 to P.S. and RETICS RD12/0019/0003 (TERCEL) to J.L.D.L.P cofunded by FEDER “una manera de hacer Europa.” It was also supported by the Regenerative Medicine Program of Instituto de Salud Carlos III and Valencian Community to Centro de Investigación Principe Felipe. The authors are grateful to Dr. A. Dorronsoro for critical review of the work and Dr. K McCreath for manuscript editing.
Author Disclosure Statement
No competing financial interests exist.