
Abstract
Neural stem cells have been considered as a source of stem cells that can be used for cell replacement therapies in neurodegenerative diseases, as they can be isolated and expanded in vitro and can be used for autologous grafting. However, due to low percentages of survival and varying patterns of differentiation, strategies that will enhance the efficacy of transplantation are under scrutiny. In this article, we have examined whether alterations in Geminin's expression, a protein that coordinates the balance between self-renewal and differentiation, can improve the properties of stem cells transplanted in 6-OHDA hemiparkinsonian mouse model. Our results indicate that, in the absence of Geminin, grafted cells differentiating into dopaminergic neurons were decreased, while an increased number of oligodendrocytes were detected. The number of proliferating multipotent cells was not modified by the absence of Geminin. These findings encourage research related to the impact of Geminin on transplantations for neurodegenerative disorders, as an important molecule in influencing differentiation decisions of the cells composing the graft.
Introduction
I
The use of fetal ventral mesencephalic neurons in clinical trials has resulted in long-lasting improvement of motor symptoms, nevertheless, it was followed by side effects in many patients, and it has also triggered ethical issues [6 –8]. To circumvent these obstacles, research has been focused on the use of alternative stem cell sources in PD transplantations.
Adult neural stem cells (aNSCs) are multipotent precursor cells that reside in specific areas of the adult brain, namely the subependymal zone (SEZ) of the lateral ventricles [9 –13]. The use of aNSCs in transplantations can bypass ethical concerns and complications, such as teratoma formation, which has been observed in embryonic stem cells (ESCs) transplantations. In addition, use of aNSCs offers the advantage of a possible autologous transplantation. Several studies have been carried out for the transplantation of aNSCs derived from the SEZ either into the intact or the parkinsonian rodent striatum. Survival rates were consistently low in all studies and differentiation into dopaminergic neurons differed widely between them [14 –20].
Geminin is a small protein that has been shown to regulate self-renewal and commitment decisions in several stem cell populations [21 –24]. Our previous work has shown that Geminin is expressed in cortical progenitor cells during embryogenesis, while its expression persists in aNSCs of the SEZ. Its expression is diminished upon initiation of differentiation [25]. Moreover, we have shown that loss of Geminin's expression in the developing cortex leads to the expansion of neural progenitor cells (NPCs) located in the ventricular zone and subventricular zone, while its overexpression promotes cell cycle exit and subsequent differentiation [26].
Alteration of gene expression in aNSCs could be advantageous for the survival of transplanted cells and their differentiation pattern. The aim of the present study was to elucidate whether differences in expression levels of Geminin could influence cell division and differentiation decisions of aNSCs transplantations in the 6-OHDA hemiparkinsonian mouse model. Our findings suggest that lack of Geminin promotes aNSCs commitment toward the oligodendrocytic lineage at the expense of the neuronal differentiation, without altering their ability to self-renew.
Materials and Methods
Experimental animals
Transplantation experiments were carried out in C57 BL/6 adult male mice (3–5 months old, 25–30 g), which derived from a colony maintained in Biomedical Sciences Research Center “Alexander Fleming.”
Mice that permit the conditional inactivation of Geminin were generated by targeting exons 3 and 4 of the mouse Geminin gene (GemininFl/Fl) [27]. For the spatiotemporal inactivation of Geminin in the aNSCs, GLASTCreERT2 transgenic mice were used [28]. To label the cells in which recombination has occurred, GLASTCreERT2 mice were crossed with mice in which a Fl-STOP-Fl-YFP cassette was introduced in the ROSA26 locus [29]. Mice were genotyped for the different Geminin alleles and the GLASTCreERT2 transgene by polymerase chain reaction using specific primers [27,28,30]. CreERT2 activity was induced by intraperitoneal injection of 1 mg of tamoxifen (Sigma, T-5648) twice a day for five consequent days. Tamoxifen was dissolved in corn oil at 37°C overnight at a concentration of 20 mg/mL.
Mice were sacrificed 21 days after the last injection. aNSCs were isolated from GemininFL/KO;GLASTCreERT2;Rosa26StopYFP and GemininFL/WT;GLASTCreERT2;Rosa26StopYFP mice, respectively. Animals were kept in a light–dark cycle 12:12 with lights on at 08:00 h and had free access to food and water.
All animal-related procedures were approved by the Veterinary Administrations of the Prefectures of Achaia, Greece and were conducted in strict accordance with the European Communities Council Directive (86/609/EEC) guidelines concerning the protection of laboratory animals.
6-OHDA injections
To establish the 6-OHDA PD mouse model, stereotactic injections of 6-OHDA were carried out unilaterally in the right ascending nigrostriatal pathway (2.1 mm caudal, 1.1 mm lateral to bregma, 4.7 and 4.9 mm to the skull surface) of experimental animals. Animals were anesthetized by intraperitoneal injections of ketamine/xylazine (100 mg/mL/20 mg/mL). They were subsequently placed in stereotactic device (Stoelting, Germany), where they were injected with 1.2 μL of 6-OHDA (divided equally between the two stereotactic coordinates), using a Hamilton syringe. 6-OHDA was prepared fresh for each surgery at a concentration of 4 μg/μL in 0.2% ascorbate saline. Twenty minutes earlier, animals were treated with desipramine (20 mg/mL, 25 mg/kg; Sigma), which protects noradrenergic neurons, and Rimadyl (50 mg/mL, 16 mg/kg), a nonsteroidal anti-inflammatory drug.
During the stereotactic injection, 6-OHDA solution was released at 0.2 μL/min, and at the end of the injection, the needle remained in place for 5 min before withdrawal. Postsurgically, experimental animals were treated with intraperitoneal injections of 5% glucose in saline, and placed in cage to recover at 25°C room temperature.
Apomorphine-induced rotational testing
Apomorphine-induced rotational behavior was assessed 2 weeks postsurgically to check whether the 6-OHDA lesion triggered successfully the neurodegeneration of A9 dopaminergic neurons of the right ascending nigrostriatal pathway. Experimental animals were placed in individual plastic cages, and 10 min after habituation, they were intraperitoneally injected with a subcutaneous dose of apomorphine (0.6 mg/mL, 2 mg/kg; Sigma). Subsequently, the number of rotations contralateral to the lesion was counted for the following 30 min (rotations were recorded for three consecutive minutes every 10 min). A lesion was considered successful when mice performed at least three contralateral rotations per minute. The aforementioned experimental animals were selected for transplantation.
Genetically engineered aNSCs' isolation and culture
Genetically engineered aNSCs were isolated 21 days following the induction of Cre activity, from the SEZ of 2–3 months old GemininFL/KO;GLASTCreERT2;Rosa26StopYFP and GemininFL/WT;GLASTCreERT2;Rosa26StopYFP transgenic mice. Transgenic mice were euthanized with CO2, and their brains were dissected to isolate the layer of tissue surrounding the lateral ventricles that contains the SEZ. Isolated SEZ was digested in a solution containing trypsin and hyaluronidase at 37°C for 30 min. The trypsin activity was terminated with trypsin inhibitor. Mechanical trituration and cell straining were followed to dissociate the SEZ to single cells and remove cell aggregates (see details in ref. 31).
Primary cells were cultured in T25 flasks at 37°C in 5% CO2 atmosphere. The basal growth medium was DMEM-F12/Glutamax, supplemented with basic fibroblast growth factor (bFGF) (10 μg/mL; Peprotech) and epidermal growth factor (EGF) (10 μg/mL; Peprotech), B27 supplement (50 × ; Gibco), Penicillin/Streptomycin (10 kU/mL; Gibco), and HEPES (1 M; Gibco). After culturing for 5–6 days, free-floating neurospheres were formed, which were subsequently redissociated and allowed to reform neurospheres, at least two times, to enhance cells population. Single cells were plated into T75 flasks at a concentration of 8,000 cells/mL to reform neurospheres.
Transplantation of genetically modified aNSCs suspensions
Three weeks after the 6-OHDA lesions, suspensions of genetically engineered aNSCs were transplanted in the right lesioned striatum of experimental animals (1 mm rostral, 1.9 mm lateral to bregma, 2.7 and 3 mm to the skull surface). Specifically, experimental animals were divided into three groups: mice that received intrastriatal grafts of GemininFL/KO;GLASTCreERT2; Rosa26StopYFP aNSCs lacking expression of Geminin (n = 3, will be mentioned as “FL/KO;GLASTCre;R26”) at a concentration of 100,000 cells/2 μL in 1× phosphate-buffered saline (PBS), a group of mice that received intrastriatal grafts of GemininFL/WT;GLASTCreERT2; Rosa26StopYFP aNSCs exhibiting a normal expression of Geminin (n = 4, will be mentioned as “FL/WT;GLASTCre;R26”) at a concentration of 100,000 cells/2 μL PBS 1 × , and a Control group of animals that received 2 μL of PBS 1× instead of being grafted with aNSCs (n = 3, will be mentioned as “Control”).
For aNSCs transplantations, neurospheres were enzymatically and mechanically triturated to create single-cell suspensions. To verify viability and lack of aggregates, trypan blue exclusion was used. Cell suspensions were transplanted to experimental animals in the aforementioned coordinates using a Hamilton syringe and following the procedure of 6-OHDA injections. Apomorphine-induced rotational behavior was assessed 4 weeks after transplantations to evaluate grafting effectiveness.
Immunofluorescence experiments and data analysis
Five weeks after transplantation, mice were anesthetized with Ketamine/Xylazine (100 mg/mL/20 mg/mL) and transcardially perfused with 4% paraformaldehyde (PFA) in PBS 1 × . Dissected brains were postfixed in 4% PFA for 16 h, cryoprotected in 30% sucrose solution for 48 h, and embedded into gelatin cubes soaked into chilled Isopentan (−20°C). Subsequently, serial brain sections of 12 μm were obtained with a Leica cryostat and used for immunofluorescence experiments.
For immunofluorescence experiments, coronal brain sections of adult mouse brains were fixed in 4% PFA (Sigma) for 10 min, washed in PBS 1× and permeabilized with 0.3% Triton X-100 in PBS 1× . Sections were treated in blocking buffer (3% bovine serum albumin, 10% fetal bovine serum in PBS-Tween 0.1%) for at least 1 h and incubated with primary antibodies overnight. Subsequently, sections were washed with 0.1% Tween in PBS 1× and incubated for 1 h with fluorescently labeled secondary antibodies (see details in Ref. [25,32]).
The following primary antibodies were used: chicken anti-green fluorescent protein (GFP) (1:1,000; Aves Labs), rabbit anti-tyrosine hydroxylase (1:200, Merck Millipore), mouse anti-Tuj1 (1:1,000; Sigma), rabbit anti-Ki67 (1:1,000; Vector Laboratories), rabbit anti-Sox2 (1:1,000; Chemicon), rabbit anti-Olig2 (1:1,000; Chemicon), and mouse anti-O4 (1:1,000; Merck Millipore). Concerning secondary antibodies, Alexa Fluor anti-chicken 488, Alexa Fluor anti-rabbit 568, and Alexa Fluor anti-mouse 647 (Molecular Probes) were used at a dilution of 1:1,000. In all immunofluorescence experiments, Draq5 (Biostatus) or Hoechst (Sigma) was used for nuclear staining.
Sections were analyzed by confocal fluorescence microscopy (Leica TCS SP5 with a Leica DMI600B microscope, which can excite molecules at four wavelengths: 488, 568, and 647 nm and ultraviolet spectrum). Image analysis was performed in Adobe Photoshop CS6.
Data analysis
Coronal brain sections of Control animals both at the level of striatum and of the substantia nigra pars compacta (SNC) were photographically captured by 20× and 63× objectives, to evaluate the effectiveness of 6-OHDA in the lesioned striatum. To characterize grafts' morphology and size in FL/WT;GLASTCre;R26 and FL/KO;GLASTCre;R26 experimental animals, all striatal sections containing grafted cells were photographically captured with a 20× objective. Evaluation of graft survival was performed by immunofluorescence experiments using anti-GFP and Hoechst or Draq5, and cells were counted every third 12 μm coronal tissue section of the transplanted striatum. Grafts were captured with a 63× objective, and photographs were covering the whole length of each graft. All GFP+ cells containing a Hoechst + nucleus were counted, and the total number of double-positive GFP+Hoechst+ cells was multiplied by three to compensate for the sampling frequency.
To detect and compare grafts' cell fate in FL/WT;GLASTCre;R26 and FL/KO;GLASTCre;R26 experimental animals, data presented for each marker were obtained by quantifying grafted cells expressing this marker in at least two different sections per animal containing the graft and captured by a 63× objective in as many slices as needed to cover the whole length of the graft. All photographs were obtained at four different levels of each 12 μm-section using 3.46 μm confocal optical sections. Data are presented as mean ± standard deviation (SD). A statistical analysis was performed using unpaired two-tailed Student's t-test. Differences were considered statistically significant at P < 0.05.
Results
Effectiveness of 6-OHDA injections
Parkinsonian mouse model was generated following stereotactic unilateral injections of 6-OHDA in the right ascending nigrostriatal pathway. Left hemisphere remained intact, to be used as a control. Efficiency of 6-OHDA toxicity was evaluated in mice by apomorphine challenges. The extent of 6-OHDA lesion was verified by immunofluorescence experiments on brain sections of mice both at the level of striatum and at the SNC site (Supplementary Fig. S1; Supplementary Data are available online at
Derivation of aNSCs used for transplantation
aNSCs were isolated from the SEZ of mice permitting the conditional inactivation of the Geminin gene (GemininFl/Fl) [23], which were crossed with mice carrying transgene GLASTCre and the fluorescent lineage marker Rosa26StopYFP. Experimental animals grafted with aNSCs derived from GemininFL/KO;GLASTCreERT2;Rosa26StopYFP transgenic mice were referred to as FL/KO;GLASTCre;R26, while animals transplanted with aNSCs derived from GemininFL/WT;GLASTCreERT2;Rosa26StopYFP transgenic mice were referred to as FL/WT;GLASTCre;R26.
Before transplantation, fluorescence microscopy was used to verify that transplanted cells were GFP positive, suggesting that successful recombination has occurred (Supplementary Fig. S2). Cells derived from FL/WT;GLASTCre;R26 and FL/KO;GLASTCre;R26 animals were transplanted into n = 4 and n = 3 parkinsonian mice, respectively. These mice successfully received intrastriatal grafts (1 mm rostral, 1.9 mm lateral to bregma, 2.7 and 3 mm to the skull surface), composed of 100,000 cells/2 μL in 1× PBS. Control animals (n = 3) received 2 μL of phosphate-buffered saline at the same coordinates.
Rotational behavior of grafted mice
Apomorphine-induced rotational behavior test was conducted 1 week pretransplantation and 4 weeks posttransplantation, to detect whether grafted cells altered rotational behavior. All animals exhibited contralateral to the side of lesion rotations, when injected with apomorphine before transplantation. Mice receiving cells derived from FL/WT;GLASTCre;R26 mice (animals 4–7, Fig. 1) and three mice receiving FL/KO;GLASTCre;R26 aNSCs (animals 8–10, Fig. 1) presented statistically significant decrease of rotations/3 min. No statistically significant difference was detected between grafted groups. The group of mice that received only phosphate-buffered saline into their denervated striatum (animals 1–3, Fig. 1) did not show any improvement in rotational scores.

Apomorphine-induced rotational behavior improved after adult NSCs striatal transplantation derived from FL/WT;GLASTCre;R26 and FL/KO;GLASTCre;R26 mice. Contralateral rotational behavior was tested 2 weeks following 6-OHDA injections (Apomorphine I) and 3–4 weeks postgrafting (Apomorphine II). All seven transplanted animals demonstrated significant decrease in the number of rotations counted for 3 min, while Control animals did not (*P < 0.05, **P < 0.005). Data are presented as mean ± SD. SD, standard deviation.
Survival and incorporation of grafted cells into the lesioned striatum
To comparatively study grafts' survival and integration into the lesioned striatum between the two transplanted groups, 5 weeks posttransplantation animals were sacrificed and postfixed. Immunofluorescence experiments were performed on 12 μm coronal brain sections of mice receiving aNSCs derived from FL/WT;GLASTCre;R26 or FL/KO;GLASTCre;R26. To track grafted cells, sections at the level of striatum were immunostained using anti-GFP antibody. In all animals examined, GFP+ cells were detected in the denervated striatum 5 weeks following transplantation.
Grafted cells of both groups formed a vertical elongated structure in the dorsal part of striatum (Fig. 2). The rostro–caudal dimension of the graft was estimated to be at least 288 μm in both groups, presenting no statistically significant difference. On the contrary, the dorsal–ventral extent was on average 397.5 ± 7.41 μm in FL/WT;GLASTCre;R26-grafted cells, while FL/KO;GLASTCre;R26-grafted cells presented an increased value of 512.3 ± 88.15 μm.
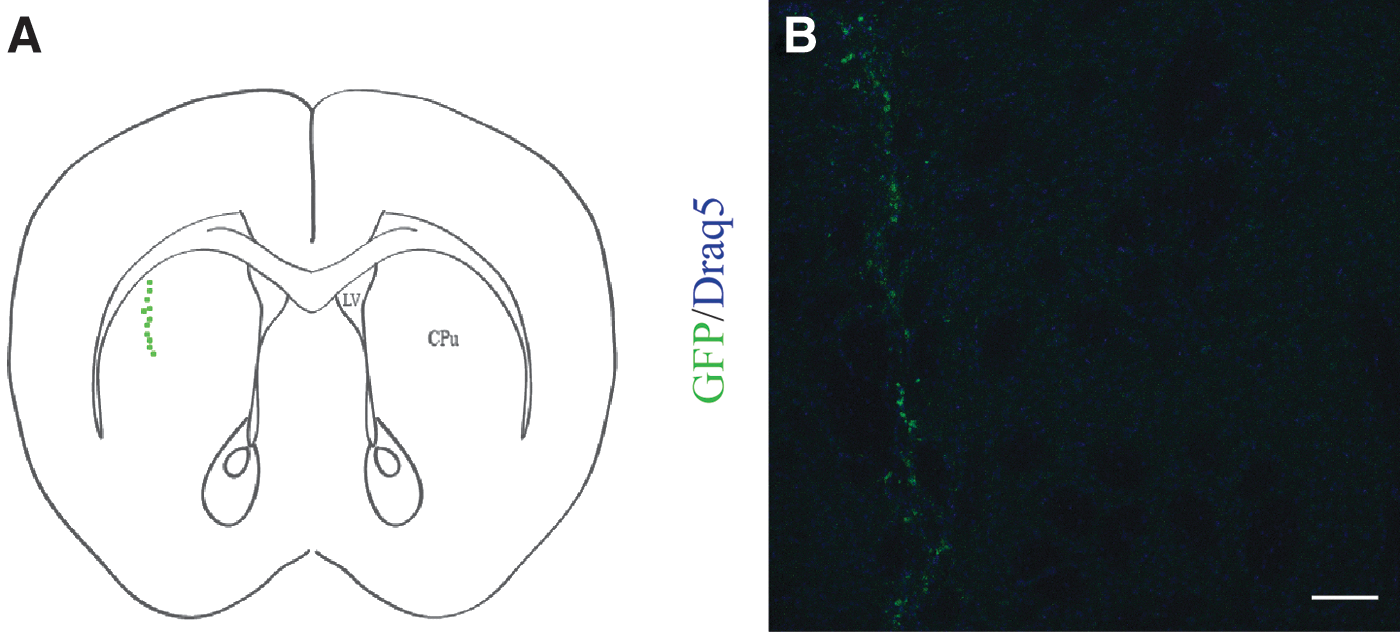
Identification of the graft in the right denervated striatum.
Concerning cell survival, the total number of GFP-positive cells in striatal sections of each graft was measured, and the percentage of survival was calculated relative to the initially transplanted number of cells. The percentage of survival ranged between 0.32% and 0.58% with a mean of 0.38 ± 0.06 for FL/WT;GLASTCre;R26 and 0.19%–0.66% with a mean of 0.46 ± 0.14 for FL/KO;GLASTCre;R26-grafted cells presenting no statistically significant differences.
Impact of Geminin's ablation on the differentiation pattern of grafts
To shed light on how the absence of Geminin impacts on the differentiation pattern of the grafted cells in the 6-OHDA mouse model of PD, immunofluorescence experiments were conducted using neuronal, glial, and stem/progenitor cell markers. Triple immunofluorescence experiments were conducted on coronal brain sections of mice transplanted with aNSCs using anti-GFP, to mark grafted cells, anti-TH, to label dopaminergic neurons, as TH is a limiting enzyme for DA synthesis, and anti-beta III Tubulin (anti-Tuj1), as a marker of early postmitotic and differentiated neurons [33].
The percentage of TH+ cells was 54.74% ± 3.08% and 42.05% ± 1.44% in FL/WT;GLASTCre;R26 (n = 4) and FL/KO;GLASTCre;R26 (n = 3) grafted cells, respectively (P < 0.05, Fig. 3). All Tuj1-positive cells in the grafted area were expressing the TH enzyme, indicating that aNSCs that differentiate into neurons were dopaminergic (Fig. 3).
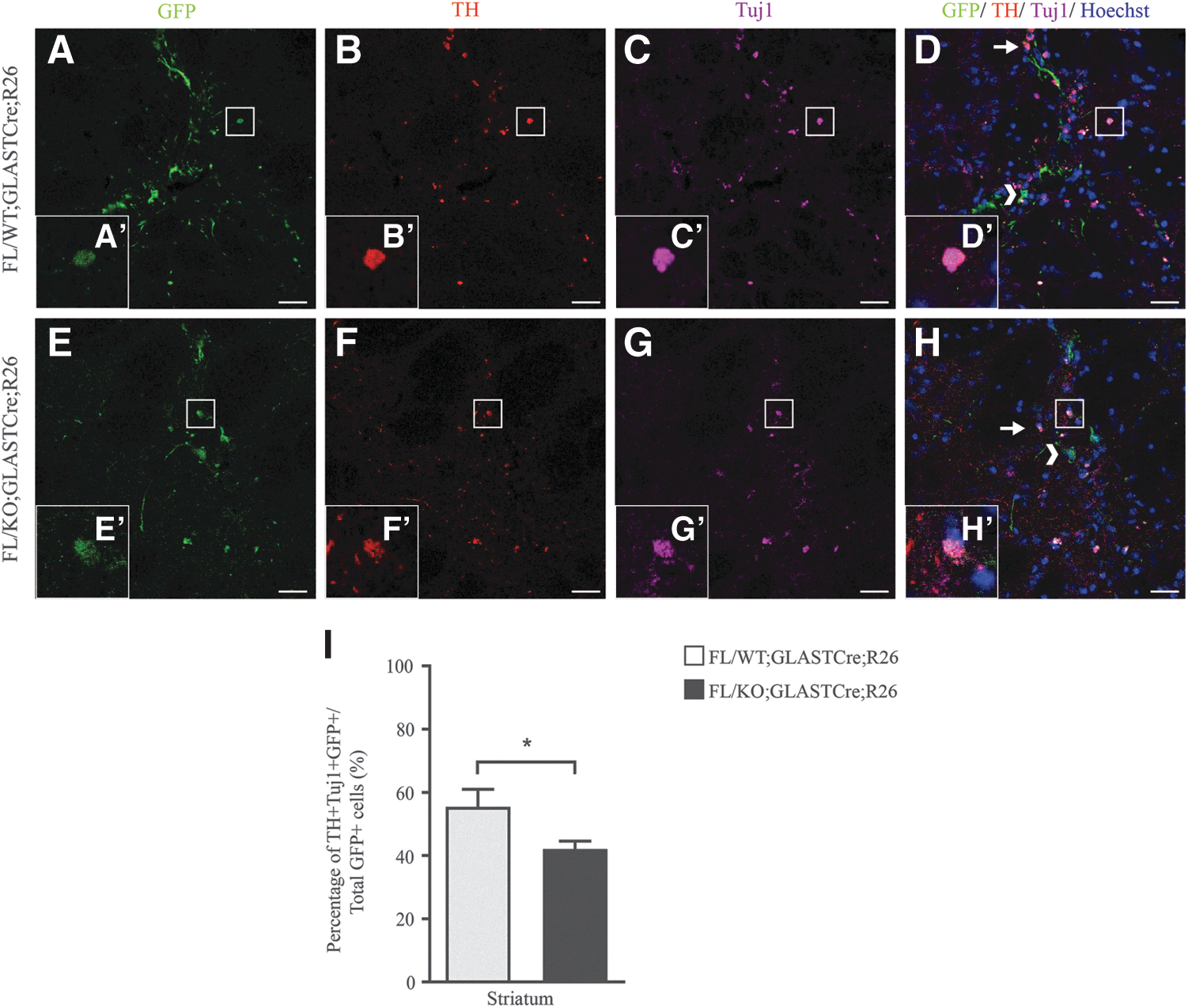
Grafted cells that differentiate into dopaminergic neurons are decreased upon deletion of Geminin. Triple-immunofluorescence staining with antibodies against TH
Grafted cells were immunostained with an antibody against Ki67, a protein detected in proliferating cells, and an antibody against Sox2, which consists a transcriptional factor, expressed in NPCs. The percentages of proliferating cells in the grafted area were 65.17% ± 3.53% and 58.98% ± 1.85% for FL/WT;GLASTCre;R26 (n = 3) and FL/KO;GLASTCre;R26 (n = 3), respectively, without presenting a statistically significant difference between the two groups (Fig. 4). Multipotency capacity was not affected by the ablation of Geminin, since the percentage of Sox2-positive cells was 51.70.18% ± 2.59% in FL/KO;GLASTCre;R26 (n = 3) and 57.00% ± 3.16% in FL/WT;GLASTCre;R26 (n = 4)-grafted cells (Fig. 5).
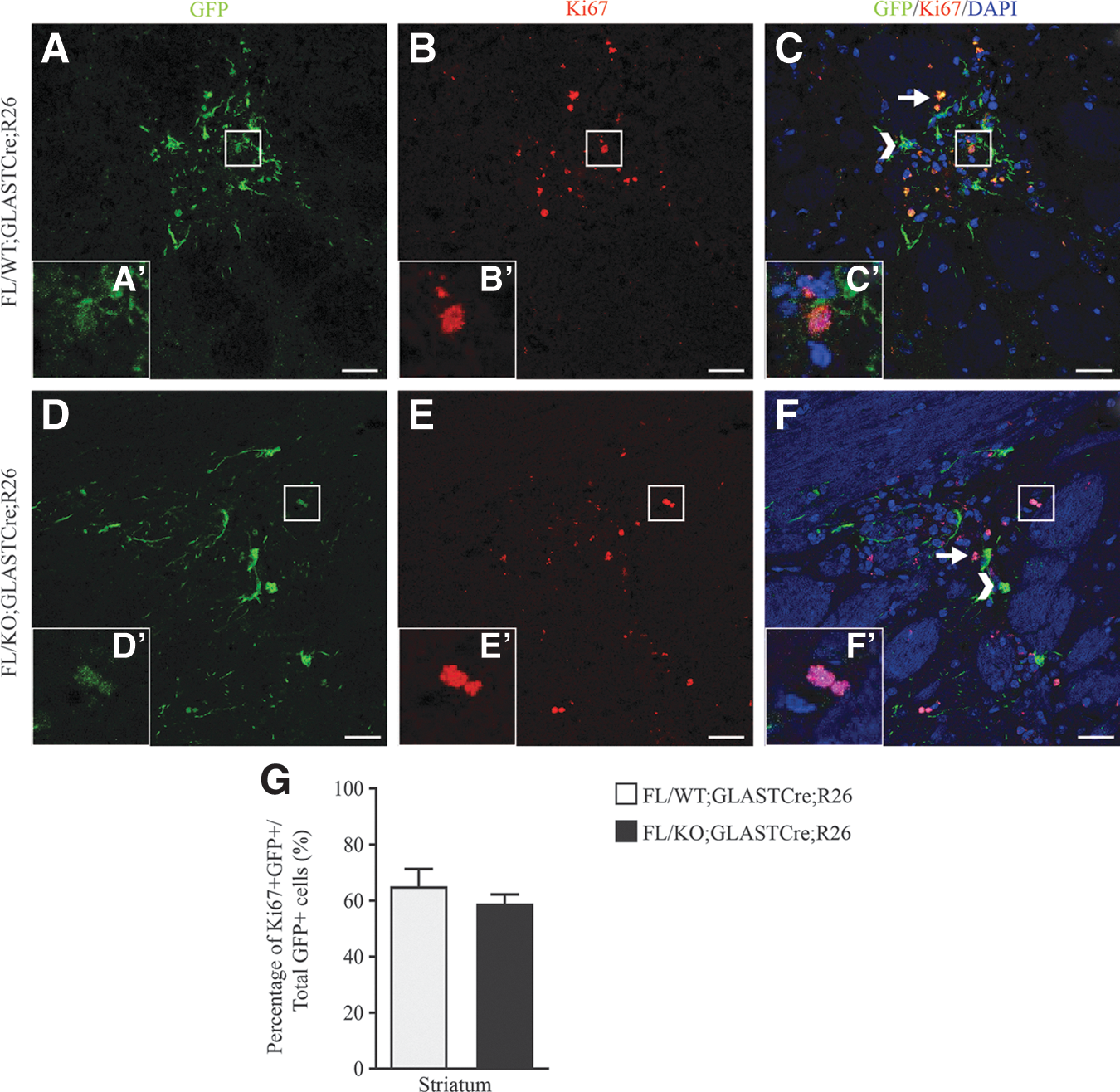
Proliferation rate of adult NSCs transplanted into the lesioned striatum is not altered by Geminin's absence. Coronal brain sections of FL/WT;GLASTCre;R26
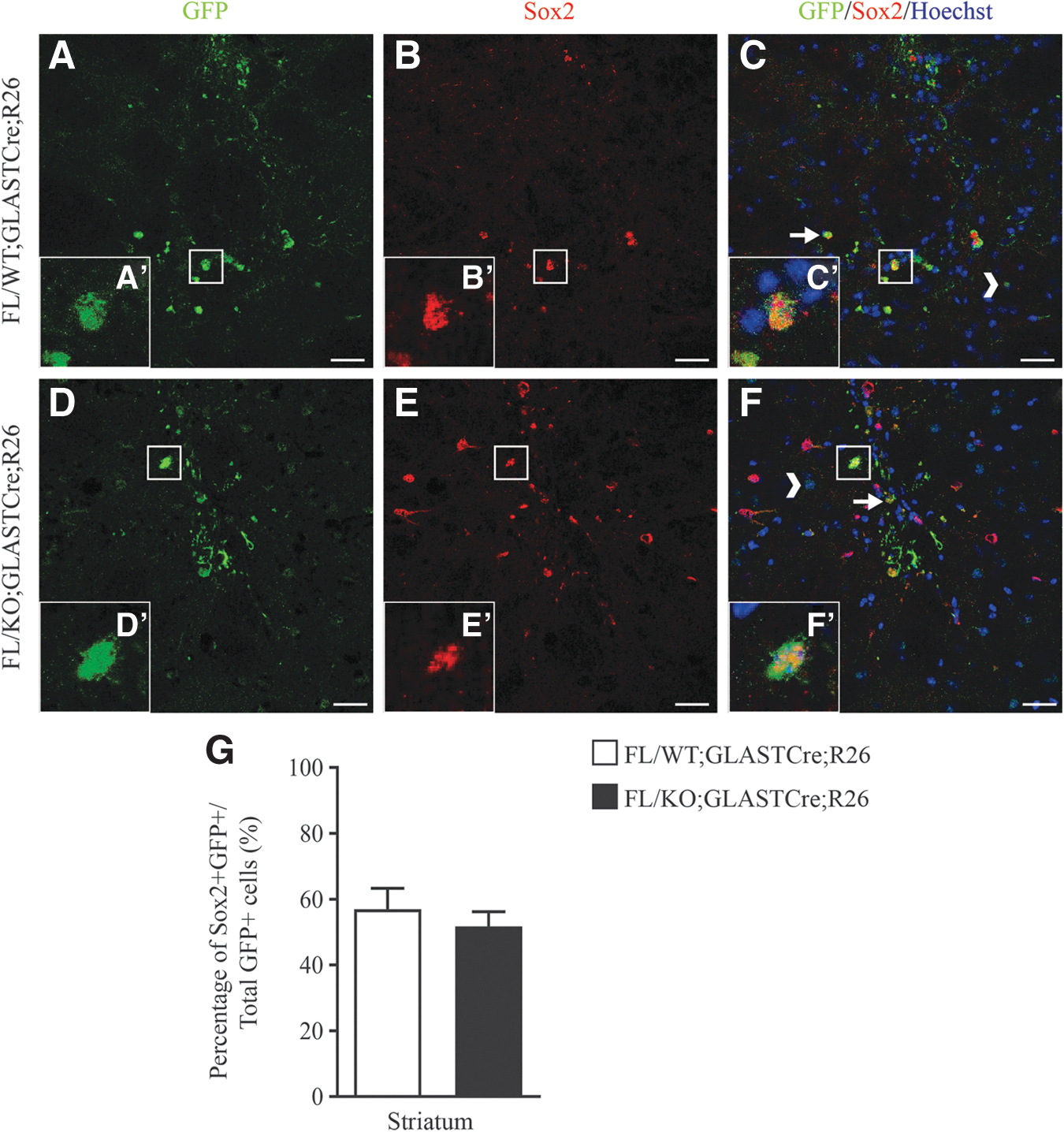
Lack of Geminin's expression does not affect grafted cells multipotency. Striatal sections of mice transplanted with FL/WT;GLASTCre;R26
Finally, the inclination of grafted cells to follow the oligodendrogenesis pathway was assessed. Immunofluorescence experiments were carried out using an antibody against Olig2, a transcription factor expressed both in oligodendrocytes and in NPCs committed to the oligodendrocytic lineage. The percentage of Olig2-positive cells compared with the total number of GFP-positive cells was 30.31% ± 3.69% and 55.28% ± 2.25% in FL/WT;GLASTCre;R26 (n = 3) and FL/KO;GLASTCre;R26 (n = 3)-grafted animals, when compared with wild-type grafted cells (P < 0.005, Fig. 6).

Hemiparkinsonian mice transplanted with recombined adult neural stem cells lacking Geminin present increased levels of oligodendrogenesis in the graft. Coronal brain sections of 6-OHDA mice transplanted with genetically modified wild-type
Immunofluorescence experiments were further carried out using an O4 antibody, which labels fully differentiated oligodendrocytes. The percentage of grafted aNSCs differentiated into O4+ mature oligodendrocytes, compared with the total number of GFP-positive cells, was 12.73% ± 0.437% and 39.59% ± 8.83% in cells derived from FL/WT;GLASTCre;R26 (n = 3) compared with grafted cells from FL/KO;GLASTCre;R26 (n = 2), respectively (P < 0.05, Supplementary Fig. S3).
Discussion
Transplantations of aNSCs in different brain regions have shown that aNSCs survive into the intact rodent striatum, cortex, hippocampus, and olfactory bulbs [16]. Grafted aNSCs generate mostly astrocytes, and only at the olfactory bulbs, neuronal differentiation has been observed [16,17]. For that reason, culturing of aNSCs in bFGF revealed a broader capability of migration and differentiation into neurons [20]. Grafting aNSCs into PD models resulted in improving motor behavior [34]. Immunohistochemistry experiments exhibited migrating behavior of the grafted cells, but their survival rate was low and the percentage of cells differentiating into dopaminergic neurons varied a lot (2%–70%) between the studies [18,34]. To ameliorate the above characteristics, several studies were focused on Nurr1 and Oct-4, which induce the dopaminergic phenotype [15,19].
Previous work from our laboratory showed that Geminin plays a critical role on regulating self-renewal and commitment/differentiation decisions in neural and hematopoietic stem cells [22,26,35].
In the present study, we have evaluated the potential impact of Geminin, in transplantations of aNSCs derived from SEZ. Our results show that aNSCs isolated from the SEZ of genetically modified mice, either lacking or retaining the expression of Geminin, were integrated efficiently into the lesioned striatum of 6-OHDA-induced hemiparkinsonian mouse model. Transplanted cells led to a statistically significant decrease of their rotational behavior that was not observed in mice that received PBS solution, similar to what we have previously shown [36]. Proliferation, survival rate, and multipotency of aNSCs were not altered by the absence of Geminin. Differentiation of transplanted cells into neurons was over 50%, possibly due to the presence of bFGF and EGF in the culture media of aNSCs, which are considered to enhance neuronal lineage differentiation [37].
Our findings show that absence of Geminin leads to decreased generation of dopaminergic neurons composing the graft. These findings are in line with previous studies of our laboratory, indicating that lack of Geminin expression decreases the ability of NPCs of the developing cortex to generate early born neurons [26]. Quite interestingly, lack of Geminin expression favors the oligodendrocytic lineage at the expense of neuronal differentiation. It was previously shown that deletion of Brg1 in the SEZ leads aNSCs to acquire oligodendrocyte lineage at the expense of the neuronal fate [38]. Furthermore, Geminin has been supported to interact with Brg1 regulating ESC self-renewal and neuronal differentiation [26,39 –41]. It is therefore possible that Geminin and Brg1 act in the same pathway, regulating fate commitment of aNSCs.
Our data suggest that altering the expression levels of Geminin may be used to modulate the fate of aNSCs transplanted in a mouse model of PD. This is quite interesting as previous experiments from our laboratory suggest that overexpression of Geminin, in NPCs of mouse developing cortex, was favoring neuronal differentiation [26]. Amplification of oligodendrocytes followed by Geminin's ablation, could act supportively to the remaining and the newborn DA neurons of the host striatum, as oligodendrocytes are responsible for the long-distance transmission of action potentials, through the production of myelin enfolding the adjacent axons, but have also other myelin-independent supportive to neuron functions, affecting neuronal survival and protection [42].
This outcome could be beneficial in the preclinical studies of several neurodegenerative diseases, where the dysfunction of oligodendrocytes has been reported, such as multiple system atrophy, amyotrophic lateral sclerosis, and Alzheimer's disease [43].
Footnotes
Acknowledgments
This work was supported by Aristeia II, GEMCCTR “Self-renewal and differentiation decisions in neural stem cells: Geminin, cell cycle control and transcriptional regulation” to S.T. We thank Prof. Magdalena Goetz for providing GlastCreERT2 mice.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.