
Abstract
The aim of stem cell therapy after cardiac injury is to replace damaged cardiac tissue. Human cardiac progenitor cells (CPCs) represent an interesting cell population for clinical strategies to treat cardiac disease and human CPC-specific antibodies would aid in the clinical implementation of cardiac progenitor-based cell therapy. However, the field of CPC biology suffers from the lack of human CPC-specific markers. Therefore, we raised a panel of monoclonal antibodies (mAb) against CPCs. Of this panel of antibodies, we show that mAb C1096 recognizes a progenitor-like population in the fetal and adult human heart and partially colocalize with reported CPC populations in vitro. Furthermore, mAb C1096 can be used to isolate a multipotent progenitor population from human heart tissue. Interestingly, the two lead candidates, mAb C1096 and mAb C19, recognize glycosylated residues on PECAM1 (platelet and endothelial cell adhesion molecule 1) and GRP78, respectively, and de-N-glycosylation significantly abolishes their binding. Thereby, this report describes new clinically applicable antibodies against human CPCs, and for the first time demonstrates the importance of glycosylated residues as CPCs specific markers.
Introduction
P
The human protein repertoire lacks a Sca-1 homologue [9], which makes it difficult to translate this isolation method into a clinically relevant protocol. Since other used markers for CPCs lack specificity, we decided to generate new human CPC-specific monoclonal antibodies (mAb) specifically raised against Sca-1+ CPCs [10]. We have defined a panel of antibodies, of which mAb C19 and mAb C1096 are the top hits, demonstrating specificity to human embryonic stem cell-derived CPCs while they do not bind to fibroblasts, endothelial cells, or undifferentiated human embryonic stem cells [10]. Interestingly, when analyzing the binding epitope of mAb C19 in more detail, we found that the antibody is directed against a glycosylated residue on the progenitor cell's surface [10].
Glycosylation is an enzymatic post-/co-translational modification during which carbohydrate structures or glycans are attached to a protein or lipid. It plays a critical role in stem cell maintenance and cell fate decisions and changes in the glycosylation profile can lead to altered biological behavior [11]. Furthermore, glycans on the cell surface are important during development [12] and signal transduction. The glycome changes upon differentiation and many of the pluripotency-associated markers are glycoproteins, such as stage-specific embryonic antigen 3 (SSEA3) and SSEA4 [11,13]. Therefore, the complexity of carbohydrate motifs on the cell surface can function as a hallmark of the progenitor state.
Unfortunately, the field of CPC biology has progressed very slowly, suffering from the lack of human CPC-specific markers. Therefore, the goal of this study was to identify the epitope of mAb C1096 and determine whether mAb C1096 labels progenitor cells in the human heart. Thereby, we questioned whether mAb C1096 can be used as a tool to isolate a progenitor population from human heart tissue. Since our data reveal that mAb C1096 recognizes N-glycosylated platelet and endothelial cell adhesion molecule 1 (PECAM1/CD31), we also investigated the importance of glycan motifs for the recognition of human CPCs.
Materials and Methods
Ethics statement and human cardiac tissue collection
This study was conducted according to the ethical statements expressed in the Declaration of Helsinki. Human fetal hearts, aged between 14 and 22 weeks postgestation, were obtained after elective abortion, based on individual informed consent. Adult human atrial samples (auricles) were collected as surgical waste under general informed consent. Handling of human heart tissues was carried out according to the official guidelines of the Leiden University Medical Center and approved by the institutional Medical Ethics Committee.
Cell isolation, culture, and cardiomyocyte differentiation
Cells were isolated, cultured, and cardiomyocyte differentiation was induced as previously described for Sca-1+ CPCs [5]. For CPC isolation, fetal (whole hearts or ventricles) heart tissue was digested to a single cell suspension with collagenase A (Roche) overnight (O/N) at 4°C followed by magnetic-activated cell sorting (MACS) according to the manufacturer's recommendations (Miltenyi Biotec). Primary antibodies that were used include anti-Sca-1 (FITC-conjugated; Miltenyi Biotec), mAb C19 (mouse IgM), and mAb C1096 (mouse IgG1). Secondary antibodies, coupled to magnetic beads: anti-FITC, anti-mouse IgM, and anti-mouse IgG (Miltenyi Biotec).
Cells were cultured on 0.1% gelatin (Sigma)-coated dishes and maintained in growth medium, which consists of a 1:4 mixture of EGM-2 complete (Lonza) and Medium 199 (M199; Invitrogen) supplemented with 10% fetal calf serum (FCS; Gibco), 1× nonessential amino acids (Gibco), and 100 U/mL penicillin/streptomycin (pen/strep; Gibco).
To induce differentiation toward cardiomyocytes [5], cells were cultured in a medium containing 1:1 Iscove's modified Dulbecco's medium and Hams F12 (Invitrogen), supplemented with 2% horse serum (Gibco), 1× nonessential amino acids (Gibco), insulin–transferrin–selenium (Invitrogen), and 100 U/mL pen/strep (Gibco). Differentiation was induced by addition of 5 μM 5-azacytidine (Sigma) for three consecutive days. Subsequently, cells were treated with 1 ng/mL transforming growth factor beta 3 (TGFβ3) twice weekly for a period of 3 weeks. During the differentiation experiment, ascorbic acid (0.1 mM; Sigma) was added every 2 days.
Heart tissue and cardiosphere processing for immunohistochemistry
Human heart tissue (fetal whole hearts or ventricles and adult auricles) was isolated in phosphate-buffered saline (PBS), fixated O/N at 4°C in 4% paraformaldehyde. The hearts were processed through series of PBS, 50% ethanol (EtOH), and 70% EtOH for 1 h each. Human cardiospheres were handled as previously described [14]. Briefly, the spheres were fixed in 4% paraformaldehyde, immersed in increasing concentrations of EtOH and washed in butanol for 15 min each. Heart tissue and cardiospheres were embedded in Paraclean II® (Klinipath). Sections of 6 μm (heart tissue) or 5 μm (cardiospheres) thickness were mounted onto coated glass slides (SuperFrost® Plus slides from Klinipath).
Immunofluorescent staining
Cultured cells were fixed in 4% paraformaldehyde for 15 min and permeabilized using PBS/0.25% Triton X-100. Samples were blocked in 5% FCS/PBS or 10% FCS/0.1% Triton X-100/PBS and incubated O/N at 4°C with primary antibody.
Paraffin-embedded heart sections or cardiospheres were deparaffinized and dehydrated in EtOH. Antigen retrieval was performed by boiling for 20 min in Vector® Antigen Unmasking Solution (Vector) in a pressure cooker. The sections were washed in Milli-Q (MQ) and PBS, blocked with 1% bovine serum albumin in 0.1% Tween-PBS for 30 min, and incubated with primary antibody O/N at 4°C.
Primary antibodies used in this study: mAb C19 (mouse IgM, 1:100), mAb C1096 (mouse IgG1, 100% hybridoma supernatant), cTnI (goat, 1:1,000; HyTest) PECAM1 (Clone M20, goat or rabbit, 1:800; Santa Cruz), C-Kit (rabbit, 1:50; Dako), NKX2.5 (rabbit, 1:100; Santa Cruz), GATA4 (rabbit, 1:100; Santa Cruz), alpha smooth muscle actin (αSMA) (mouse IgG2a, 1:100; Sigma), Vimentin (rabbit, 1:100; Cell Signaling), NANOG (rabbit, 1:100; Abcam), Survivin (rabbit, 1:100; Santa Cruz), and Endoglin (goat, 1:100; R&D Systems). Alexa Fluor-conjugated secondary antibodies (Invitrogen) were incubated for 1 h at a 1:250 dilution, followed by 0.1% Tween-PBS and PBS washes. The slides were mounted with ProLong Gold-DAPI Antifade (Invitrogen) reagent.
For all immunofluorescence stainings, the high-magnification views correspond to the outlined areas. Number of mAb C1096+ and mAb C19+ CPC-like cells were quantified in at least three longitudinal sections per heart and represented as the number of cells per mm2 of cardiac tissue.
De-N-glycosylation of heart tissue sections
Ultrapure water was used for all preparations and washes and was generated from an 18.2 MΩ-cm PURELAB Ultra System from Elga (Ede, The Netherlands). Glass slides were incubated for 1 h on a 60°C heating plate to enhance the adherence of the sections, followed by deparaffinization through sequential washes in xylene (1 × 5 min, 1 × 10 min; Merck), 100% EtOH (2 × 5 min; Merck), 70% EtOH (1 × 5 min; Merck), and water (2 × 5 min). Heat-induced antigen retrieval (HIAR) was performed in a retriever (Electron Microscopy Sciences, Hatfield, PA) in 10 mM citric acid buffer adjusted to pH 6 with 1 M NaOH at high temperature (>120°C) and high pressure. After HIAR and cooling down, tissues were washed with water (2 × 5 min) and dried in a vacuum desiccator (10 min). N-Glycosidase F (PNGase F) solution (0.1 μg/μL in water (recombinant PNGase F was obtained from Flavobacterium meningosepticum and a kind gift from Anand S. Mehta and Richard R. Drake, Medical University South Carolina, SC) was sprayed homogenously on the tissue using a SunCollect pneumatic sprayer (10 layers at 10 μL/min; Sunchrom, Friedrichsdorf, Germany), followed by O/N incubation at 37°C in a humid environment for the release of N-glycans. Released N-glycans were removed with repeated wash steps in water and subsequent immunofluorescence staining for mAb C1096 and mAb C19 were performed as described above.
Deep sequencing and cluster analysis
Human Sca-1+ CPC were differentiated into cardiomyocytes following the protocol as described above. Total RNA was isolated using QIAzol reagent, ribosomal RNA was removed, and cDNA prepared using the Illumina Kits (RiboZero and ScriptSeq, respectively), and sequenced on an Illumina Hi-Seq instrument. Single 50-nucleotide reads were aligned to hg19 genome assembly by TopHat and analyzed by SAMtools and BAMtools. Analysis of gene expression during human CPC to cardiomyocyte differentiation was performed as described in Anders and Huber [15]. Cluster analysis was subsequently performed using the cluster analysis function in MATLAB (Version 2016a).
Statistics
Graphs are represented as mean ± standard error of the mean. Samples were compared using an unpaired Student's t-test, one-way ANOVA with Bonferroni correction, and Kruskal–Wallis one-way ANOVA with Dunn's post hoc test. Significance was assumed when P < 0.05. GraphPad Prism (Version 6) was used for statistical analysis. The remaining materials and methods can be found in the Supplementary Data; Supplementary Data are available online at
Results
MAb C1096 recognizes a CPC-like cell in fetal and adult human heart tissue
To investigate which cell types are recognized by mAb C1096 in the human heart, we performed co-immunofluorescent stainings of human fetal heart tissues for mAb C1096 reactivity from week 14 until 22 postgestation, in combination with antibodies recognizing specific cell populations, PECAM1 for endothelium, cardiac troponin I (cTnI) for cardiomyocytes, and Wilms' tumor 1 (WT1) for epicardium. As can be appreciated in Fig. 1A, mAb C1096 binds to a population of single cells scattered throughout the myocardium, not colocalizing with the cardiomyocyte marker cTnI. This pattern is similar to the expression pattern observed when we use mAb C19 (Fig. 1A) and was also observed in adult human heart tissue (atrium in Fig. 1C) Mostly, mAb C1096+ or mAb C19+ cells were located as single cells in between cardiomyocytes and outside blood vessels (Fig. 1D), a pattern which has also been reported for Sca-1+ CPCs [16]. When counting the number of cells stained in different heart sections, we observed that the total number of mAb C1096+ cells per fetal heart section was significantly lower compared with mAb C19+ cells (Fig. 1B). The epicardium, which is the outer-most layer of the heart, has been suggested to harbor a progenitor population (reviewed in Smits and Riley [17]). Interestingly, mAb C1096 also stained cells within the fetal epicardial layer. Colabeling for WT1, an epicardial marker, suggests that these cells are of epicardial origin (Fig. 1E).
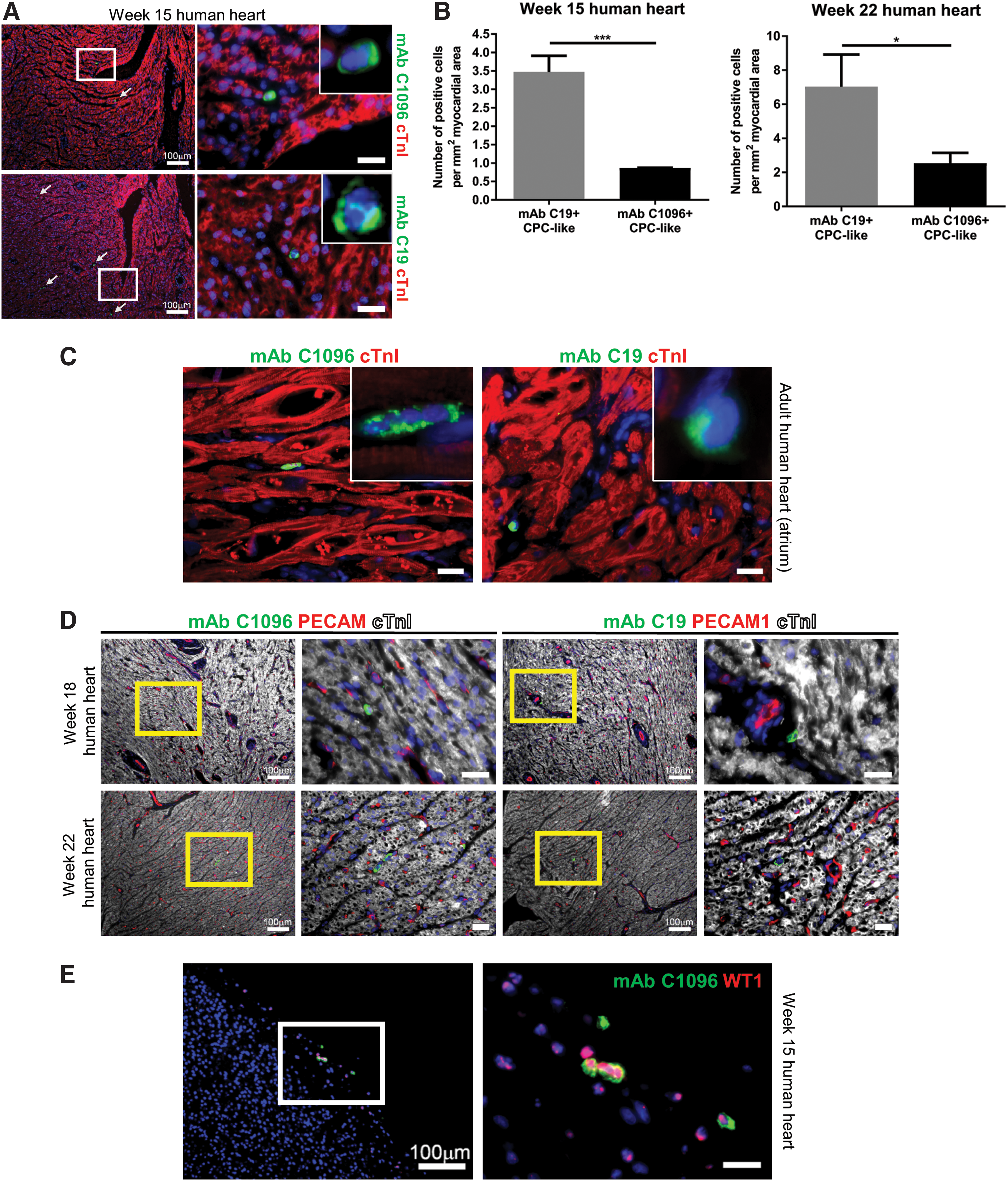
MAb C1096 and mAb C19 recognize a progenitor-like population in the fetal and adult human heart.
Since fluorescent colocalization analysis using multiple cell type-specific antibodies revealed that mAb C1096 expression does not colocalize with PECAM1 or cTnI, we next investigated whether these may represent a CPC population. To date, the expression of C-Kit is mostly used to identify human CPCs, although debated. Therefore, we analyzed whether the expression of mAb C1096 and mAb C19 colocalizes with an anti-C-Kit antibody. In fetal and adult human cardiac tissue, we observed hardly any colocalization between mAb C1096 and C-Kit (±0.23% of C-Kit+ cells is mAb C1096+) (Fig. 2A, B, and week 22 heart in Supplementary Fig. S1A). In contrast, there was partial overlap between mAb C19 and C-Kit in both the fetal and the adult human heart (Fig. 2C, D, and week 22 heart in Supplementary Fig. S1B), suggesting that mAb C1096 and mAb C19 recognize a different population of cells in vivo.
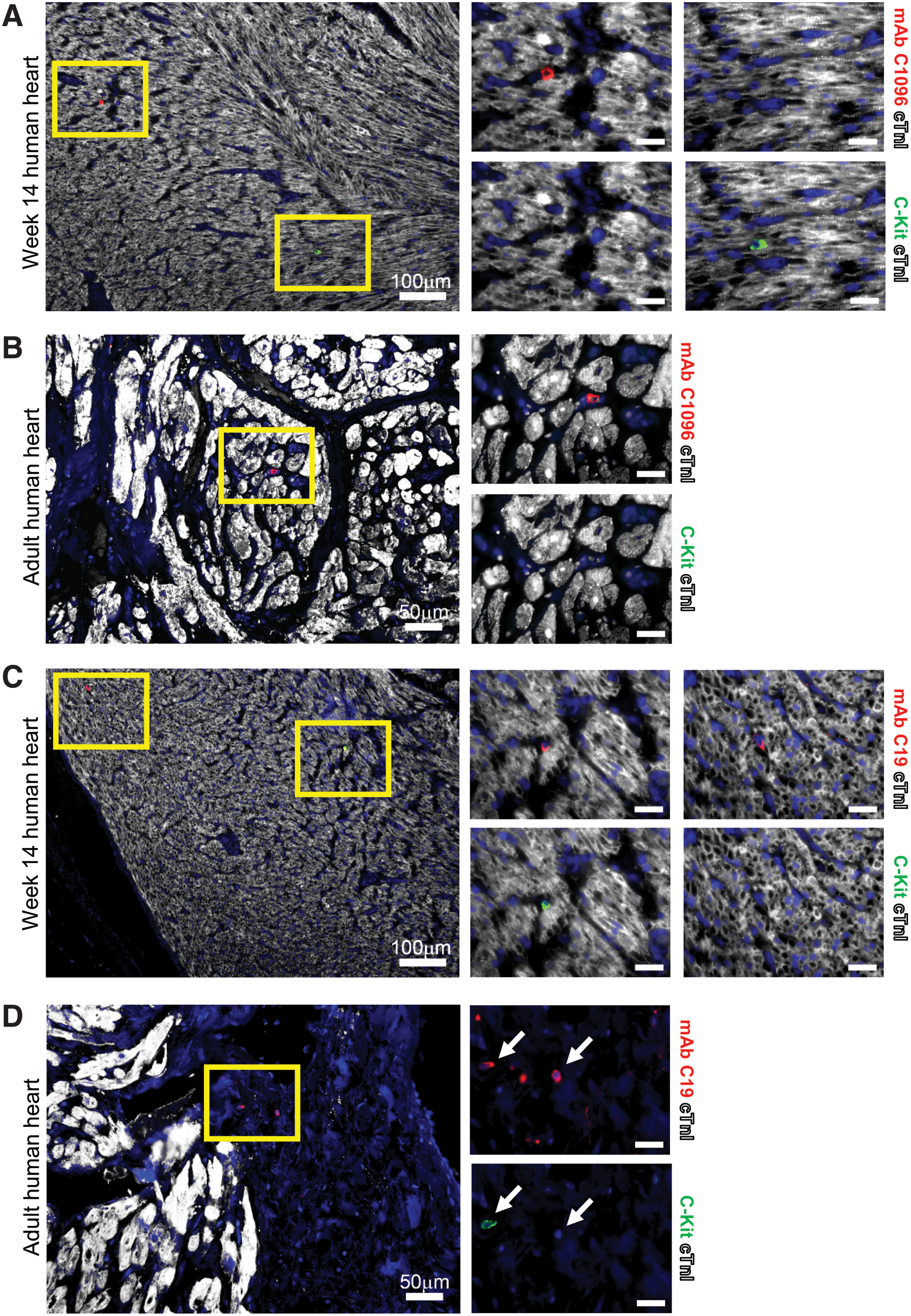
MAb C19 partially colocalize with C-Kit in fetal and adult human heart tissue. Immunostaining shows almost no overlap between C-Kit+ and mAb C1096+ cells in
Since the number of cells within one section is very low, we analyzed if mAb C1096 recognizes progenitor cells in vitro. We first analyzed MACS isolated Sca-1+ and mAb C19+ CPCs and found that in vitro, mAb C1096 binds to both CPC populations (Fig. 3A). We then analyzed cardiospheres as they have been reported to contain cells that express the progenitor markers C-Kit and Sca-1 [18]. Similar to C-Kit, mAb C1096 and mAb C19 bind to cells within human cardiospheres (Fig. 3B, C); however, they do stain significantly less cells compared with C-Kit (Fig. 3D). Furthermore, we analyzed colocalization between C-Kit, and mAb C1096 or mAb C19 within cardiospheres. While only ∼20% of the mAb C1096+ cells is also positive for C-Kit, C-Kit expression was observed in half of the mAb C19+ population (Fig. 3E). Of the C-Kit+ cells, 10%–15% is positive for mAb C19 or mAb C1096 (Fig. 3F).

MAb C1096 recognizes in vitro human CPC populations.
In summary, we show that mAb C1096 identifies a cell type in the human heart that belongs to the non-myocyte population and partially colocalizes with reported human CPC populations in vitro.
MAb C1096 isolates a progenitor cell population from the human heart
To gain more insight into the cell type recognized by mAb C1096, we performed MACS with mAb C1096 and the anti-Sca-1 antibody from the same fetal heart tissue sample, thereby enabling a direct comparison between the two sorted cell populations. MACS yielded an equal number of cells, 6 days after isolation (Fig. 4A, left). MAb C1096+ cells did grow using the same culture conditions as for Sca-1+ CPCs and maintained their mAb C1096 reactivity during in vitro maintenance (Fig. 4B). Furthermore, the morphology of mAb C1096+ cells is similar to Sca-1+ CPCs (Fig. 4A, right).
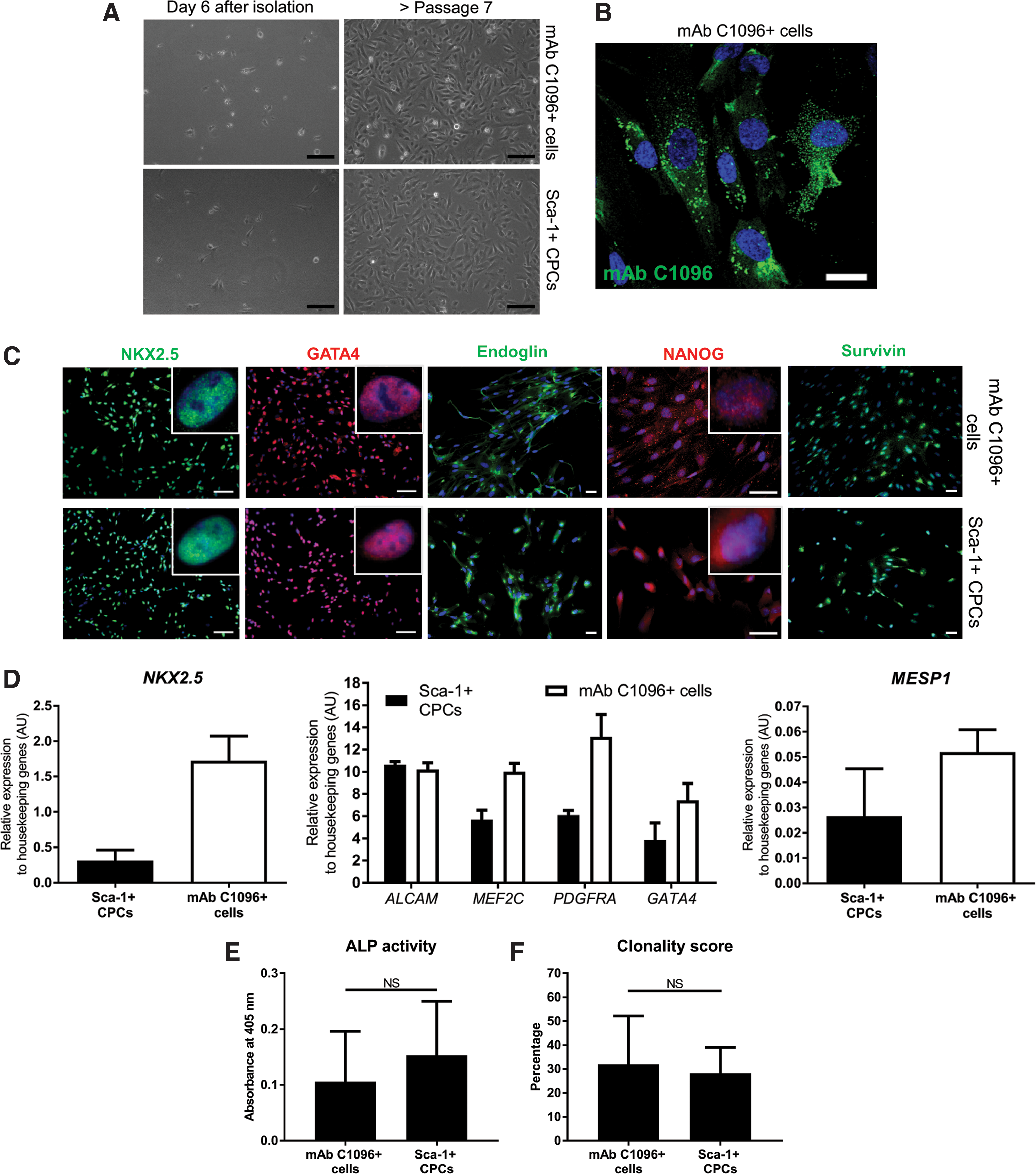
MAb C1096 isolates a progenitor population from the fetal human heart.
One of the features of CPCs is that they express early cardiac transcription factors [10]. To determine if mAb C1096+ cells are committed to the cardiac lineage, the expression of early cardiac markers in mAb C1096+ cells was analyzed. The transcription factors, NKX2.5 and GATA4, were present in the nucleus of mAb C1096+ cells to a similar level as Sca-1+ CPCs (Fig. 4C, D). In addition, mAb C1096+ cells were positive for the stem cell markers Endoglin and NANOG (Fig. 4C), indicative of a progenitor cell profile. We further validated the CPC character of mAb C1096+ cells analyzing the expression of MEF2C, ALCAM, and PDGFRA (Fig. 4D) [10] and show that all genes, including the mesodermal transcription factor MESP1, were equally expressed in mAb C1096+ cells and Sca-1+ CPCs (Fig. 4D) [19]. The mature cardiac markers, cardiac troponin T (cTnT), beta myosin heavy chain, and cardiac actin, were not expressed by mAb C1096+ cells (data not shown). Furthermore, by determining the alkaline phosphatase activity we found that mAb C1096+ cells, Sca-1+ CPCs, and mAb C19+ CPCs have equal mesenchymal properties (Fig. 4E, and data not shown). Finally, we performed a colony-forming assay and observed that mAb C1096+ cells have the ability to form a colony when plated as a single cell (Fig. 4F), again suggesting that the mAb C1096-sorted cells have a progenitor cell phenotype.
MAb C1096+ cells have multipotent capabilities
One of the features of CPCs is that they can form multiple cell types of the mesodermal lineage. To determine if mAb C1096+ cells fulfill this criterion, we performed cardiac and vascular differentiations and a calcification assay. We have previously shown that Sca-1+ and mAb C19+ CPCs differentiate toward cardiomyocytes when treated with 5-azacytidine, followed by TGFβ stimulation [4,5,10]. Using the same protocol, we did see an upregulation of cTnT in mAb C1096+ cells after 3 weeks of treatment (Fig. 5A), but cTnT levels were lower compared with Sca-1+ CPCs. This suggests that either mAb C1096+ cells need a different cardiac differentiation protocol or represent a progenitor population with a different differentiation preference compared with Sca-1+ CPCs and mAb C19+ CPCs.
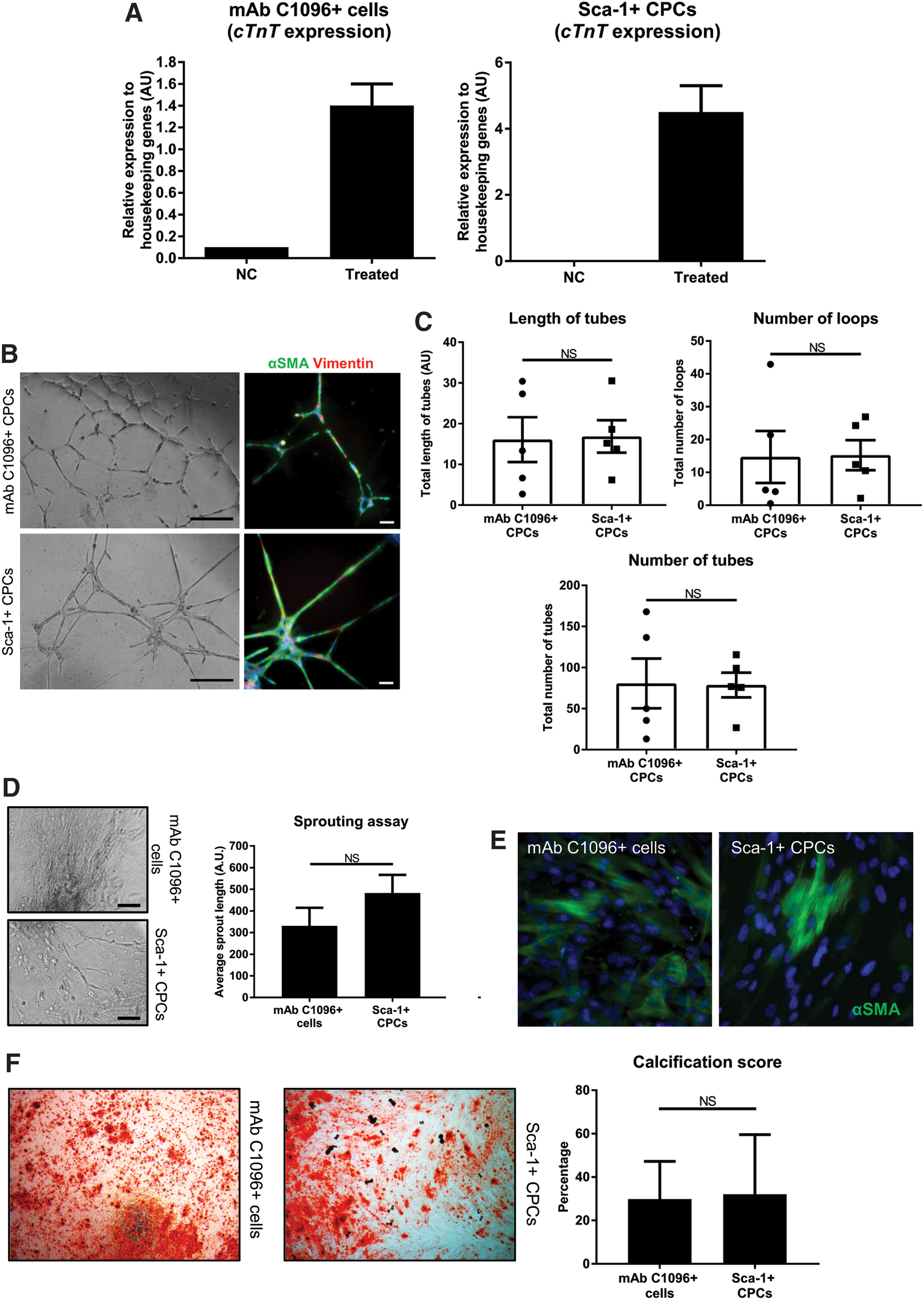
MAb C1096+ cells have multipotent capabilities.
To determine if mAb C1096+ cells can form vascular cells we performed two angiogenesis assays. mAb C1906+ cells were seeded on a thin layer of Matrigel, where they formed a network of capillary-like structures comparable to Sca-1+ CPCs (Fig. 5B) [4]. The tubes contained αSMA- and Vimentin-expressing cells suggesting differentiation toward a smooth muscle cell phenotype (Fig. 5B, right). The length of tubes, number of tubes, and number of closed loops were not significantly different between the two populations (Fig. 5C). In addition to their ability to form tubes, mAb C1096+ cells were able to migrate away from a collagen-embedded spheroid (Fig. 5D) and differentiate toward smooth muscle cells in 2D cultures (Fig. 5E). Finally, mAb C1096+ cells underwent calcification when present in osteogenic medium (Fig. 5F). In conclusion, mAb C1096+ cells, such as Sca-1+ CPCs, are multipotent in their differentiation ability.
MAb C1096 recognizes PECAM1
To understand the role of mAb C1096 as a novel marker for CPCs we conducted further characterization of its antigen. Western blot analysis of CPC whole cell lysates showed that mAb C1096 identified a protein of ∼115 kDa, of which antibody binding was sensitive to reduction (Fig. 6A). This indicates that disulfide linkages played a key role in maintaining conformation of the antigen recognized by mAb C1096. Enrichment of the antigen by immunoprecipitation was successful, and the antigen band was excised, and sent for protein identification by mass spectrometry. The top hit from the mass spectrometry data was PECAM1. This putative antigen identity was validated by using a commercial anti-PECAM1 antibody to probe against the immunoprecipitation product of mAb C1096, which showed that they recognized the same protein (Fig. 6B). Next, we verified that Sca-1+ and mAb C19+ CPCs in vitro also express PECAM1 at the mRNA as well as at the protein level (Fig. 6C, D).

MAb C1096 recognizes PECAM1 and binds the endothelium with low affinity.
MAb C1096 binds with high affinity to human CPCs
Given that the endothelial marker PECAM1 is the target of mAb C1096, we investigated mAb C1096 localization to the endothelium in vivo. As expected, mAb C1096 recognizes the vasculature of the human heart, such as the endothelial lining of blood vessels (indicated by arrows in Fig. 6E) and cardiac valve (Supplementary Fig. S2A). Interestingly, mAb C1096 staining intensity is much lower at the endothelium compared with CPC-like cells (Fig. 6E) indicating that mAb C1096 recognizes the endothelium with low affinity and has a high affinity for human CPC-like cells.
The subcellular localization of mAb C1096 compared with a commercial PECAM1 antibody (clone M20) seems to be different which was observed for both Sca-1+ CPCs (Fig. 6F) and human umbilical vein endothelial cells (in Supplementary Fig. S2B). While mAb C1096 showed localization to the membrane, commercial anti-PECAM1 was mainly found intracellular. PECAM1 is known to localize to the cell–cell borders in a confluent culture, but becomes uniformly distributed in subconfluent monolayers [20]. Therefore, we speculate that mAb C1096 recognizes a subset of total PECAM1 protein present on the surface of the cell.
De-N-glycosylation of human heart tissue abolishes mAb C1096 and mAb C19 binding
To identify the differences between mAb C1096 and commercial anti-PECAM1, we investigated whether mAb C1096 binding is dependent on glycosylation. Upon treatment of CPC whole cell lysates with periodate, which specifically oxidizes glycan ring structures on glycopeptides, the antigen band at 115 kDa was abolished, whereas anti-actin binding was unaffected (Fig. 7A, left). Similarly, PNGaseF treatment, which cleaves off N-linked glycans specifically, also abolished binding to the 115 kDa antigen band (Fig. 7A, right). This demonstrates that mAb C1096 binds to an N-glycosylated form of PECAM1, of which the glycan is an important feature in its recognition site.

Glycosylation as CPC-specific marker.
We previously showed that mAb C19 recognizes glycosylated GRP78 [10]. Therefore, we questioned whether mAb C1096 and mAb C19 recognition of CPC-like cells in the human heart requires glycosylation. We treated consecutive serial heart sections with the glycosidase PNGase F to remove N-glycans or with water as control. Subsequently, we performed immunofluorescence staining for mAb C1096 or mAb C19, together with the commercially available PECAM1 and cTnI antibodies. The PECAM1 antibody (clone M20) was raised against a peptide at the C-terminus and therefore, binding of commercial PECAM1 should not be affected by de-N-glycosylation. Strikingly, mAb C1096 binding was completely abolished upon de-N-glycosylation (Fig. 7B). While mAb C1096 recognizes progenitor-like cells in control hearts, no positive cells were found upon PNGase F treatment. For mAb C19, de-N-glycosylation significantly reduced the amount of positive progenitor-like cells (Fig. 7C). Importantly, de-N-glycosylation of the tissue sections did not interfere with binding of commercial PECAM1 and cTnI suggesting that enzymatic treatment does not hamper general antibody recognition. Altogether, these data reveal that mAb C1096 binding to progenitor-like cells in human heart tissue is dependent on N-glycosylation of its PECAM1 antigen and suggest that also mAb C19 recognition of GRP78 is (partially) dependent on N-glycosylation.
The glycan repertoire on a cell is defined by the combinatorial action of glycosyltransferase and glycosidase enzymes at a given time. Therefore, we performed cluster analysis for glycosylation-related genes on Sca-1+ CPCs to cardiomyocyte differentiation, which suggests a shift in glycosylation from nondifferentiated to differentiated cells (Fig. 7D). Enzymes upregulated in differentiated cells (day 30) are enriched for sialyltransferases (eg, ST3GAL6) and mannosidases (eg, MANBA). In addition, we separated the glycosylation-related transcripts into the two major types of protein glycosylation, N-linked (Fig. 7E) and O-linked (Fig. 7F) glycosylation. The clustergrams suggest an increase in N-linked glycosylation and a shift in O-linked glycosylation upon cardiomyocyte differentiation. This shows that a glycan motif can be an indication of the corresponding progenitor/differentiation status.
Discussion
Cardiac death is the result of the inability of the remaining cardiomyocytes to compensate for the loss of contractile units upon injury. As potential therapeutic application, CPCs have been extensively studied as source of new cardiac tissue. However, there is no consensus on the marker profile that defines CPCs. Therefore, we have generated antibodies that can be used to isolate a multipotent progenitor population from human heart tissue.
In this study, we show that mAb C1096 can be used to identify and isolate a human cell population with CPC properties and multipotent differentiation abilities and identified PECAM1 as the target for mAb C1096. PECAM1 is often used as endothelial marker, however, it has also been found on various stem cell populations, including Sca-1+ CPCs, embryonic stem cells, and progenitors from the hematopoietic lineage [4,21,22]. In addition, Bollini et al. reported that a subfraction of mouse WT1+ epicardial cells express PECAM1 [23] showing that PECAM1 is not specific for the endothelium.
Our data suggests that there are different populations of CPC within the human heart. When analyzing human heart sections, we observed hardly any overlap between mAb C1096 and C-Kit, which has previously also been reported for Sca-1 [24]. Interestingly, overlap was observed in cardiospheres, which may be due to the selection of a certain cell type during in vitro culture. Given that our data suggest that mAb C1096 is able to isolate a CPC population from the human heart, our immunofluorescence stainings indicate that mAb C19 and mAb C1096 recognize a distinct population of CPCs. These may be developmentally related subpopulations of a common mesodermal precursor. This concept has been shown for C-Kit+ CPCs of which the C-Kit+/KDR− (kinase domain receptor) population was assumed to be myocyte progenitor cells, whereas the C-Kit+/KDR+ cells have a vascular commitment [25,26]. Furthermore, Pfister et al. showed that within the pool of side population cells [27], the PECAM1−/Sca-1+ cells have the greatest potential to mature into cardiomyocytes [28]. On the other hand, it has been reported that two lineages give rise to different parts of the heart [29]. Therefore, different CPC populations may also represent a distinct origin or cardiogenic lineage. Whether mAb C19 and mAb C1096 recognize CPCs at different developmental stages derived from a common mesodermal precursor or rather reflect a distinct lineage specification will be an important question for future research.
Interestingly, both mAb C1096 and mAb C19 recognize glycosylated residues, and binding to their antigens is dependent on this post-/co-translational modification. The glycan complex at the cell surface possesses a lineage-specific nature [11]. Glycan motifs can be used to discriminate a stem cell from its differentiating progeny and thereby reflect the differentiation stage of a cell. For example, Nash et al. identified an O-glycan motif, which was present on mouse embryonic stem cells and decreased when the cells adopted a primitive ectoderm or epiblast fate [30]. Therefore, given the various subpopulations of CPCs, glycans are interesting CPC-specific markers which can improve our CPC isolation strategies.
We showed that upon de-N-glycosylation, mAb C1096 binding to CPC-like cells was completely abolished, which confirmed that mAb C1096 recognizes an N-glycan motif. The glycan structure recognized by mAb C19 remains to be identified. In contrast to mAb C1096, the number of mAb C19+ CPC-like cells was reduced after de-N-glycosylation. This may suggest that mAb C19 binding is not completely N-glycosylation dependent or subpopulations exist within the mAb C19+ CPC-pool, which were either responsive or nonresponsive to de-N-glycosylation.
In this study, we revealed a shift in glycosylation-related enzymes upon cardiomyocyte differentiation. Glycosylation is known to play a fundamental role in the maintenance of pluripotency and embarking the path toward differentiation [11]. For example, enzymatic removal of surface sialic acids induced ectoderm differentiation of induced pluripotent stem cells (iPSCs) [31]. Downregulation of the glycosyltransferase ST6GAL1 in human pluripotent stem cells results in upregulation of developmental genes and downregulation of genes involved in pluripotency [32]. Based on our cluster analysis, ST6GAL1 was also expressed by CPCs and downregulated upon the induction of differentiation (day 2 vs. CPCs). Upon cardiomyocyte differentiation, we reported an upregulation of sialyltransferases (day 30 vs. CPCs). Consistent with this observation, the sialyltransferase ST3GAL3 has been previously shown to be significantly increased in iPSC-derived cardiomyocytes [33]. In addition, when human embryonic stem cells transition toward the three germ layers, endoderm, mesoderm, and ectoderm, they acquire polysialic acid conjugations. The glycotransferases involved in this process are expressed in a lineage-specific manner; knockdown of the endoderm glycotransferase interferes with endoderm differentiation [34]. In the context of O-linked glycosylation, our data suggest an upregulation of the N-acetylglucosamine transferase (OGT) upon initiation of cardiomyocyte differentiation. OGT plays an important role during cardiac development and heart failure (reviewed in Medford and Marsh [35]). Cardiomyocyte-specific deletion of OGT results in dysfunctional hearts during heart development and exacerbate cardiac function following MI compared with wild type [36,37]. Altogether, this suggests that glycosylation plays an active role during CPC to cardiomyocyte differentiation.
In summary, we describe new human-specific and clinically applicable antibodies against human CPCs, and for the first time stress the importance of glycosylated residues as CPC-specific markers. We show that glycan motifs are important for the recognition of human CPCs, which makes them highly interesting as therapeutic targets and suggest that already well-known markers (eg, PECAM1) can be reevaluated for progenitor-specific glycosylation marks, and could represent the missing link in the isolation of human CPCs. Our glycan-binding antibodies could be used to identify and fully characterize the human CPC population(s) and their developmental stages, and also to unravel the role of glycosylation in CPC biology. More importantly, these antibodies are a step forward in the isolation of a population of human CPCs to eventually treat a patient with heart failure. As such, glycans can be considered the next generation of CPC markers.
Footnotes
Acknowledgments
The authors are grateful to the members of their group for valuable discussion and W.C.R. Sloos, J.C.A.G. Wiegant, and A.M.A van der Laan for technical assistance. They thank Roberto Gaetani and Joost Sluijter (UMC Utrecht, the Netherlands) for providing human cardiospheres and Serguei Denissov and Mark Musters (Lead Pharma Medicine B.V., Nijmegen) for deep sequencing analysis. Furthermore, they thank the department of Cardiothoracic Surgery (LUMC, Leiden, the Netherlands) and CASA Leiden for collecting and providing human adult and fetal heart tissue. This work was supported by the Netherlands Institute for Regenerative Medicine (grant no. FES0908), the Dutch Heart Foundation project DHF-2009B063 (M.-J.G.) and project DHF-2014T013 (N.A.M.B.), the Netherlands Organization for Scientific Research (NWO) VENI (016.146.079) (A.M.S.), and a LUMC Research fellowship (A.M.S.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.