
Abstract
Pluripotent stem cells have demonstrated the potential to generate large numbers of functional cardiomyocytes (CMs) from different cell sources. Besides Wnt signaling, additional pathways are involved in early cardiac development and function. To date however, no study exists showing the effects of perturbing the canonical Wnt pathway using nonhuman primate embryonic stem (ES) cells. In this study, we investigated the effect of canonical Wnt inhibition during differentiation of nonhuman primate ES cell-derived CMs under defined, growth factor conditions. Rhesus monkey ES (rES) cells were differentiated into spontaneously beating CMs in the absence (control) or presence (treated) of Wnt inhibitor Dickkopf1 (DKK1), vascular endothelial growth factor, and basic fibroblast growth factor combined or added in a sequential manner during differentiation. Quantification and functional characterization of CMs were assessed by molecular and electrophysiological techniques. Analysis revealed no difference in average ratio of spontaneously beating clusters in both control and treated groups. However, the percentage of CMs was significantly reduced and the expressions of specific cardiac markers tested were also decreased in the treated group. Interestingly, we found that in CMs obtained from treated group, β-adrenergic receptors (β-ARs) were less expressed, their function was altered and electrophysiological studies revealed differences in action potential responsiveness to β-AR stimulation. We demonstrated that the Wnt/β-catenin pathway inhibitor, DKK1 associated with other growth factors repressed functional expression of β-ARs in rES cell-derived CMs. Thus, control of this pathway in each cell line and source is important for proper basic research and further cell therapy applications.
Introduction
C
Cardiac development is a dynamic process that is tightly orchestrated by the sequential expression of multiple signal transduction proteins, transcription factors, and signaling molecules working in a combinatory manner [6], and even subtle perturbations of these events can have catastrophic consequences in the form of congenital disease ultimately leading to heart failure. Several methods have been proposed for differentiating PSCs into CMs, ranging from the classical embryoid bodies (EBs)’ aggregation approach to chemically defined protocols. Besides Wnt/beta-catenin signaling, controlling pathways, such as fibroblast growth factors (FGF), members of the transforming growth factor β superfamily (TGFβ), bone morphogenic proteins (BMPs), activin, and nodal have been shown to improve the cardiac differentiation of PSCs [7]. Previous studies indicate that Wnt/beta-catenin pathway negatively regulates cardiac differentiation, whereas studies with pluripotent embryonal carcinoma cells suggest opposite results. Moreover, early treatment of differentiating mouse embryonic stem (ES) cells with Wnt-3A stimulates mesoderm induction, activates a feedback loop that subsequently represses the Wnt pathway, and increases cardiac differentiation [8].
Combining different small molecules and/or growth factors in a concentration and temporal-dependent manner, PSCs are guided to efficient CM differentiation. However, the generation of a homogeneous and intact functional population of CMs remains an important limitation of the current methodologies. The existing CM differentiation protocols not only generate a heterogeneous cell population but mainly focus on ES cells derived from several sources in mice and humans.
Originally, our aim was to establish a differentiation protocol that could drive the efficient differentiation of nonhuman primate rhesus monkey ES (rES) cells toward functionally intact CMs. In this study, we show that rES cells differentiated using modified previously published protocols [8 –11] with defined growth factors known to support cardiomyogenesis markedly reduce cardiac differentiation, and significantly affect electrophysiological properties of the remaining CMs. Interestedly, remaining CMs have decreased numbers of beta-adrenergic receptors (β-ARs) in comparison to control CMs. However, despite this decrease in cell and β-ARs numbers, these CMs expressed most cardiac-specific markers and retained their sarcomeric structure with an intact nucleus, thereby providing a complementary model system for comparing differentiation protocols and understanding the effect of signaling factors on cardiomyogenesis in other cell models.
Materials and Methods
Chemicals and media
Basic culture media were purchased from Gibco/Invitrogen; supplements and chemicals were from Sigma-Aldrich, unless otherwise specified. All drugs were prepared according to the manufacturer's instructions.
Cell culture and in vitro differentiation
rES cells as described previously [12] were kindly provided by Florence Wianny and Colette Dehay (Institut cellule souche et cerveau INSERM: U846, Université Claude Bernard - Lyon I, Centre de recherche Inserm 18). Cells were maintained in culture as described previously [12]. Briefly, cells were maintained in an undifferentiated state in irradiated mouse embryonic fibroblasts in 80% Dulbecco's modified Eagle's medium (DMEM) with nutrient mixture F12 and GlutaMAX (DMEM/F12- GlutaMAX) containing 20% Knockout serum replacer, 1 × nonessential amino acids, 2 mM
For cardiac differentiation of rES cells, undifferentiated cell colonies were manually cut into clumps and cultivated in a suspension culture for 6 days in Ko-DMEM (Gibco®; Invitrogen, Darmstadt, Germany) with 20% fetal calf serum (FCS), 200 μM β-mercaptoethanol (all from Life Technologies), 0.1 mM nonessential amino acids, 2 mM
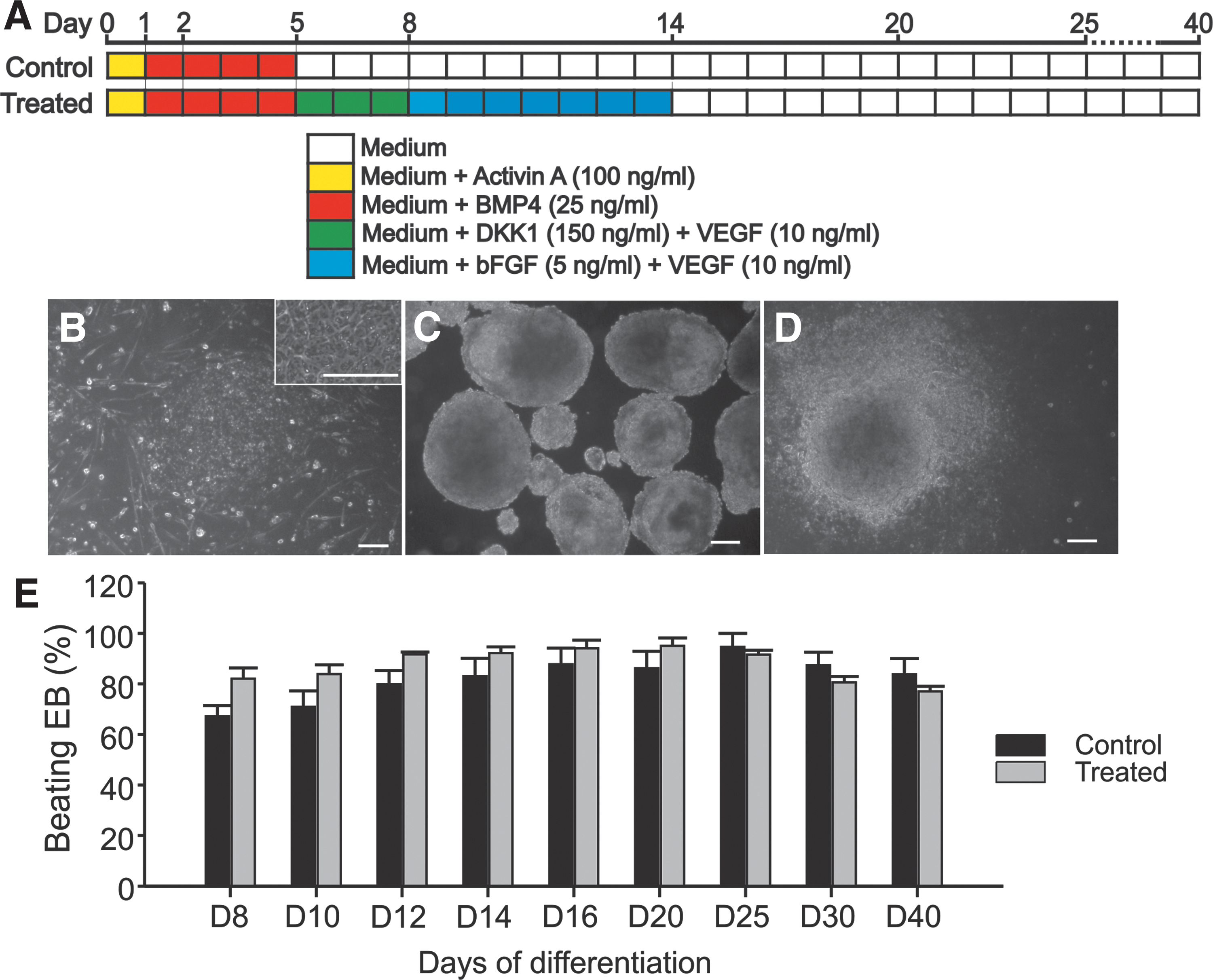
Differentiation of rES cells into CMs.
Single CM preparation
CMs from control and treated conditions were dissociated from day 23 contracting EB using 0.05%Trypsin-ethylenediaminetetraacetic acid (EDTA). The isolated cells were plated on sterile 0.1% gelatin-coated glass coverslips and cultured in DMEM (Gibco) containing 20% fetal bovine serum (FBS). Isolated cells were further used for immunocytochemical characterizations and electrophysiological experiments after 2–3 days of plating.
Immunofluorescence
Dissociated single CMs were fixed with 4% paraformaldehyde, permeabilized with solution containing 0.25% Triton X-100 (Sigma-Aldrich) and 0.5 M ammonium chloride in 0.25 M Tris buffered saline (TBS) (pH 7.4) for 10 min and blocked with Roti-Block solution (1:10) for 1 h. For sarcomeric structure evaluation of CMs, samples were further incubated overnight at 4°C with primary antibodies against α-actinin (1:800; Sigma-Aldrich) or cardiac Troponin T (cTnT, 1:100; Thermo Scientific), diluted using 1:10 Roti-block solution. On the next day, the preparations were incubated with the secondary antibodies: anti-mouse-IgG1 (1:1,000; Alexa Fluor 555) or anti-rabbit-IgG1 (Alexa Fluor 647). Nuclei were counterstained with Hoechst 33342 (1:500; Sigma-Aldrich). After washing, stainings were embedded in ProLong Gold Antifade Reagent and evaluated using a Zeiss Axiovert 200 epifluorescence microscope equipped with an ApoTome unit. The total number of nuclei and number of positive cells were counted manually and used to calculate the percentage of α-Actinin and cTnT-positive cells. A minimum of 400 cells per experiment were analyzed.
For cardiac β1-AR and β2-AR detection, double staining β1-AR/cTnT, and β2-AR/cTnT were performed. CMs fixed and blocked as described above were incubated for 60 min at room temperature (RT) with anti-β1-AR (1:100; Thermo Scientific) or anti-β2-AR (1:100; Thermo Scientific) antibodies to detect and characterize β-ARs. After, all samples were incubated with goat anti-rabbit IgG secondary antibody AlexaFluor-555 (1:100; Invitrogen) for 60 min at RT. This was followed by incubation for 60 min at RT with antibody to cardiac Troponin T (cTnT, 1:1,000; Thermo Fisher) to detect CMs. Next, cells were immunolabeled with fluorophore-conjugated secondary antibody AlexaFluor-488 goat anti-mouse (1:1,000; Invitrogen) for 1 h. Nuclei were stained by using bisBenzimide H 33258 dye (Hoechst, 1:1,000; Sigma) followed by embedding in ProLong Gold (Invitrogen, Darmstadt, Germany) as described above. Fluorescence was analyzed using Carl Zeiss software AxioVision LE 4.6 and Leica DMRB (Leica, Wetzlar, Germany) equipped with Diskus software (Diskus, Königswinter, Germany).
Quantification of β-ARs in rES
The quantification of β-ARs in rES cell-derived CMs from control and treated conditions was carried out by measuring the fluorescent signals of specifically labeled cells. Only CMs coexpressing cTNT and β1-AR or β2-AR were included in the analysis. The specificity of the fluorescent signals was assessed according to our preceding experience and all autofluorescence-associated pitfalls were avoided as previously published [14,15]. Stained cells were visualized in high-power fields (400 × ) and captured by Diskus program (Königswinter, Germany) using Leica DMRB fluorescence microscope coupled to 3CCD JVC KY-F70B camera at conditions constant for all groups (halogen lamp output, exposure time, beam aperture, etc.). In each group, at least 10 most accurate representative high-power fields containing a total of minimum 50 cardiac cells with consistent morphological phenotypes were chosen for the evaluation. The quantification of the conventional and fluorescent cell signals is feasible in different ways, as the cell number, size, and background signals have to be taken into consideration [16 –19]. We used ImageJ freeware (Bethesda) to determine the level of fluorescence signals of a given cell region in contrast to background, thus reflecting the density of the expressed β-ARs [20,21]. First, the cell margin, was precisely selected by one of the drawing/selection tools followed by setting the measurements criteria (area, integrated density, and mean gray value selected) and activating the measure function from the analyze menu. Afterward, the regions next to the cell, containing only the background fluorescence, were measured in triplicates. Corrected total cell fluorescence (CTCF) ( = integrated density of single cell − (area of selected cell × mean fluorescence of background readings)) was determined and analyzed as described in the Statistical Analysis section. Both mean values as well as the total fluorescence of the entire inspected cell population/group are presented in the Result section.
Real-time polymerase chain reaction analysis
For quantitative real-time-polymerase chain reaction (RT-PCR), total RNA from undifferentiated rES cells and contracting EB outgrowths (∼50 EBs from each condition) was extracted at indicated time points using TRIzol reagent (Invitrogen) according to the manufacturer's instructions (Prod No. 15596-026; Life Technologies, Ambion®, Germany) and stored at −80°C until further use. cDNA was synthesized from 1 μg of total RNA by using the SuperScript® Vilo™ cDNA Synthesis Kit (Prod No. 11754050; Invitrogen, Karlsruhe, Germany) according to the manufacturer's instructions.
Real-time PCR analysis was performed using ABI-7500 Fast PCR system (Applied Biosystems, Weiterstadt, Germany). The 20 μL PCR with SYBR® Advantage® qPCR Premix (Prod No. 639676; Clontech, Mountain View, CA) was used according to the manufacturer's guidelines. The standard conditions for PCR were used, 95°C/10 min, then 40 cycles of 95°C/15 s for denaturation, 58°C/30 s for annealing, and 60°C/45 s for extension. The total RNA expression levels were normalized against the housekeeping β-actin gene levels and calculated using a relative quantitation method by 7500 Fast System SDS software 1.4.0. (Applied Biosystems, Carlsbad). The PCR primers and reaction conditions are depicted in Table 1.
Flow cytometry
For flow cytometric (FC) analysis, EBs from control and treated conditions from day 23 of differentiation were dissociated to a single cell suspension by trypsin treatment (120 μL of trypsin/EDTA solution) for 2–3 min. Cells were permeabilized in 0.5% saponin solution in 5% bovine serum albumin (BSA) for 1 h at RT and labeled with cTnT and Alexa Fluor 647 for fluorescence-activated cell sorting (FACS) analysis. Measurements were performed on a FACSCalibur flow cytometer with CellQuest Pro 6 software (both BD).
Multielectrode array measurements
To characterize the functional properties of rES cell-derived CMs generated under DKK1, VEGF, and bFGF as mentioned above, extracellular recordings of field potentials (FPs) were performed using a microelectrode array (MEA) data acquisition system (Multi Channel Systems, Reutlingen, Germany) as described previously [22]. For this purpose, day 23 spontaneously beating EBs containing CMs were mechanically dissected and plated on fibronectin-coated (2 μg/mL) MEA culture plates in DMEM supplemented with 20% FBS, 1% NAA, and 0.1 mM βME and measured 24–48 h after attachment. Standard measurements were performed at 5 kHz in differentiation culture medium. Isoproterenol (Prod No. I-2760; Sigma-Aldrich, Germany), a standard stimulator of the β-adrenergic signaling cascade, and carbachol (Prod No. C-4382; Sigma-Aldrich, Germany), a synthetic acetylcholine analog, were dissolved in serum-free medium. During recordings, the temperature was kept at 37°C. MEA data were analyzed using the Spike 2 version 7.12 Software (Cambridge Electronic Design, Cambridge, England) with in-house written specialized macros as previously described [23,24]. These tools allow for analysis of MEA signals for FP frequencies, FP duration, and amplitude.
Patch clamp recordings
Single rES cell-derived CMs generated under DKK1, VEGF, and bFGF were dissociated as described above, plated on 0.1% gelatin-coated coverslips and cultured 24–48 h before measurement. The coverslips were placed in a temperature-controlled (37°C ± 1°C) recording chamber and put on the stage of Axiovert 135TV inverted microscope (Zeiss, Germany). Action potential (APs) of each single spontaneously beating cell was recorded by the whole cell patch clamp method in current clamp mode using the EPC-9 amplifier (HEKA Electronics, Lambrecht, Germany). The PULSE/PULSEFIT program (HEKA) was used for data acquisition and analysis. The resistances of the glass electrodes used were 2–6 MΩ when filled with the pipette solution containing (mM) 50 KCl, 80 K-Aspartate, 1 MgCl2, 3 MgATP, 10 EGTA, and 10 HEPES (pH 7.2 adjusted with KOH). The glass coverslips containing the cells were placed onto a temperature-controlled (37°C) recording chamber and perfused continuously with extracellular bathing solution containing (mM) 140 NaCl, 5.4 KCl, 1.8 CaCl2, 1 MgCl2, 10 glucose, and 10 HEPES (pH 7.4, adjusted with NaOH). The response of CMs to pharmacological intervention was assessed by administering isoproterenol (ISO, 1 μM) as β-adrenoreceptor agonist, the muscarinic receptor agonist carbachol (CCh, 10 μM) as the synthetic analog of acetylcholine, direct adenyl cyclase activator Forskolin (1 μM; F6886), and isobutyl methyl xanthine (IBMX, 100 μM; I5879) as a nonspecific inhibitor of phosphodiesterase (PDE). APs were analyzed offline with custom-made AP analysis software (provided by Prof. Philipp Sasse, University of Bonn, Germany).
Statistical analysis
Results were expressed as mean ± SEM. Student's t-test was used for statistical analysis. Values with P < 0.05 were considered as statistically significant. In case of CTCF evaluation, data were first transferred from ImageJ to Microsoft Excel and later into SPSS for Windows (SPSS Inc., Surrey, United Kingdom). Kolmogorov–Smirnov test was performed to test normal distribution of the data sets. The numbers stated in the figures indicate the amount of measured cells or EBs, respectively, if not mentioned elsewhere in the legends. For MEA and Patch-clamp recordings, only cells with change in beating rate of >10% after drug application were considered responding cells and included into the statistics to avoid artificial influences.
Results
Cardiac differentiation of rES cells
To investigate the maturation process toward the formation of typical growing cell clusters called EBs, we induced cell differentiation of rES cell toward CMs by using protocol based on growth factors and DKK1 as described in Fig. 1A. During the differentiation, the EBs generated from treated condition exhibited similar morphology and growth properties to control. At day 8 of differentiation, 67.2% ± 8.4% (n = 3 differentiations) EBs of the control started beating, and increased up to 94.6% ± 9.4% (n = 3 differentiations) at day 25 of differentiation, whereas within the treated group, 82.0% ± 8.6% (n = 3 differentiations) of EBs at day 8 and 91.6% ± 3% (n = 3 differentiations) at day 25, respectively, were beating (Fig. 1E). The spontaneously beating EBs generated under treated condition (in the presence of DKK1, VEGF, and bFGF) did not show significant differences in beating rates as compared with control EBs at day 25 of differentiation (62.4 ± 28.7 beats/min (n = 11) versus 67.4 ± 28.7 beats/min (n = 12); respectively, P > 0.05). However, the derived CMs from beating EBs obtained from treated conditions at day 25 of differentiation expressed appropriate, but less cardiac mesodermal markers, GATA4 (1.5-folds less) and NKX2.5 (2.5-folds less), as well as less CM-specific markers, α-MHC (twofolds less) and cTNT (1.3-folds less) (Fig. 2A). In beating EBs derived from treated condition, immunostaining for cTNT and α-actinin demonstrated a decrease in the total number of CMs as compared with control condition (Fig. 2B and C). This result was confirmed by the FC analysis (Fig. 2E, F), which showed a significant decrease of cardiac cells from 8.6% (control) to 2.4% (treated). Moreover, higher magnification of single CMs revealed the presence of well-defined striated sarcomeric structures representing typical cardiac cross-striated patterns as stained by sarcomeric α-actinin and cTNT (Fig. 2D). The results suggest that CMs generated under DKK1, VEGF, and bFGF conserve at least their structure and/or morphology despite substantial reduction in their number.
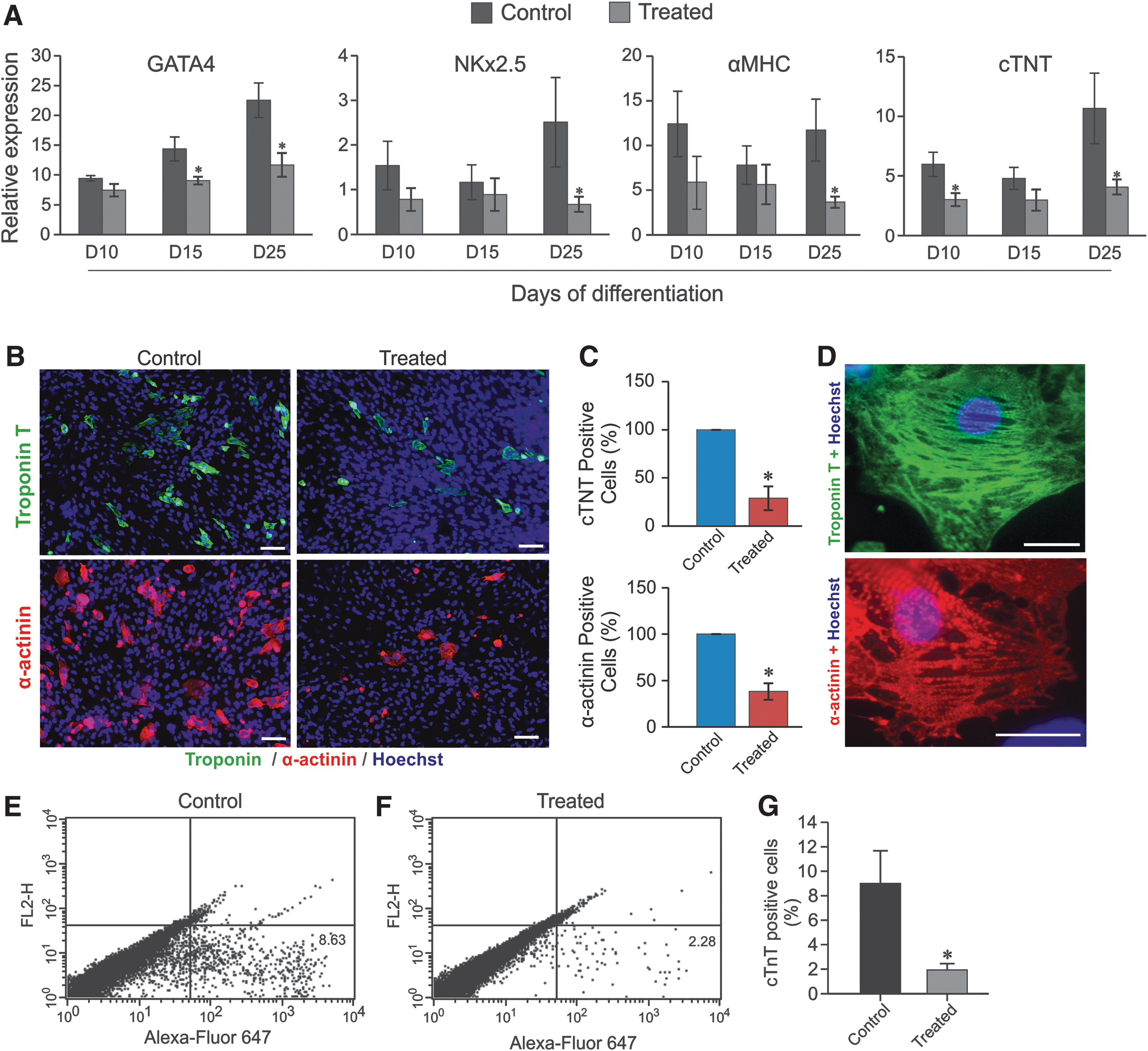
Characterization of cardiac marker genes and proteins.
Hormonal regulation in rES cell-derived CMs
Since β-adrenergic and muscarinic signaling cascade systems play major roles in regulating cardiovascular functions and to understand how the cyclic AMP-dependent protein kinase or A kinase (cAMP/PKA) signal could affect physiological function of CMs, we first investigated whether both systems are functionally expressed and active in spontaneously contracting EBs generated under DKK1, VEGF, and bFGF by examining the response of CM contraction rates to stimulation with isoproterenol. For these experiments, contracting EBs (day 23) were plated onto MEAs and cultured for up to 48 h for good attachment and growth (Fig. 3A). The analysis of FPs (Fig. 3B, C) revealed reduction of beating frequencies at baseline conditions in treated EBs (0.90 ± 0.11 Hz, n = 12) as compared with control EBs (1.26 ± 0.13 Hz, n = 11) (P < 0.05). Application of isoproterenol (ISO, 1 μM) on spontaneously beating EBs from control condition led to a significant increase in baseline FP frequency of about 63% (P < 0.05), whereas subsequent application of the muscarinic receptor antagonists carbachol (CCh, 10 μM) during the same measurements was able to inhibit the effect of Iso, and induced a decrease of FPs frequency of about 15% as compared with baseline, suggesting functional expression of both β-adrenergic and muscarinic pathways. It was however, surprising that ISO (1 μM) treatment of EBs generated under DKK1, VEGF, and bFGF did not reveal any significant changes in beating frequency of FPs, but application of CCh significantly decreased the FP frequencies (Fig. 3B, C) induced after ISO exposure. Statistical analysis confirmed the significant positive chronotropic effect of ISO in control EBs as compared with treated ones (Fig. 3C, P < 0.05). This result suggests that the β-adrenergic pathway stimulation, which normally causes an increase in heart rate seems to be nonfunctional in spontaneously beating EBs containing rES cell-derived CMs generated under DKK1, VEGF, and bFGF, whereas the muscarinic pathway remains functional.
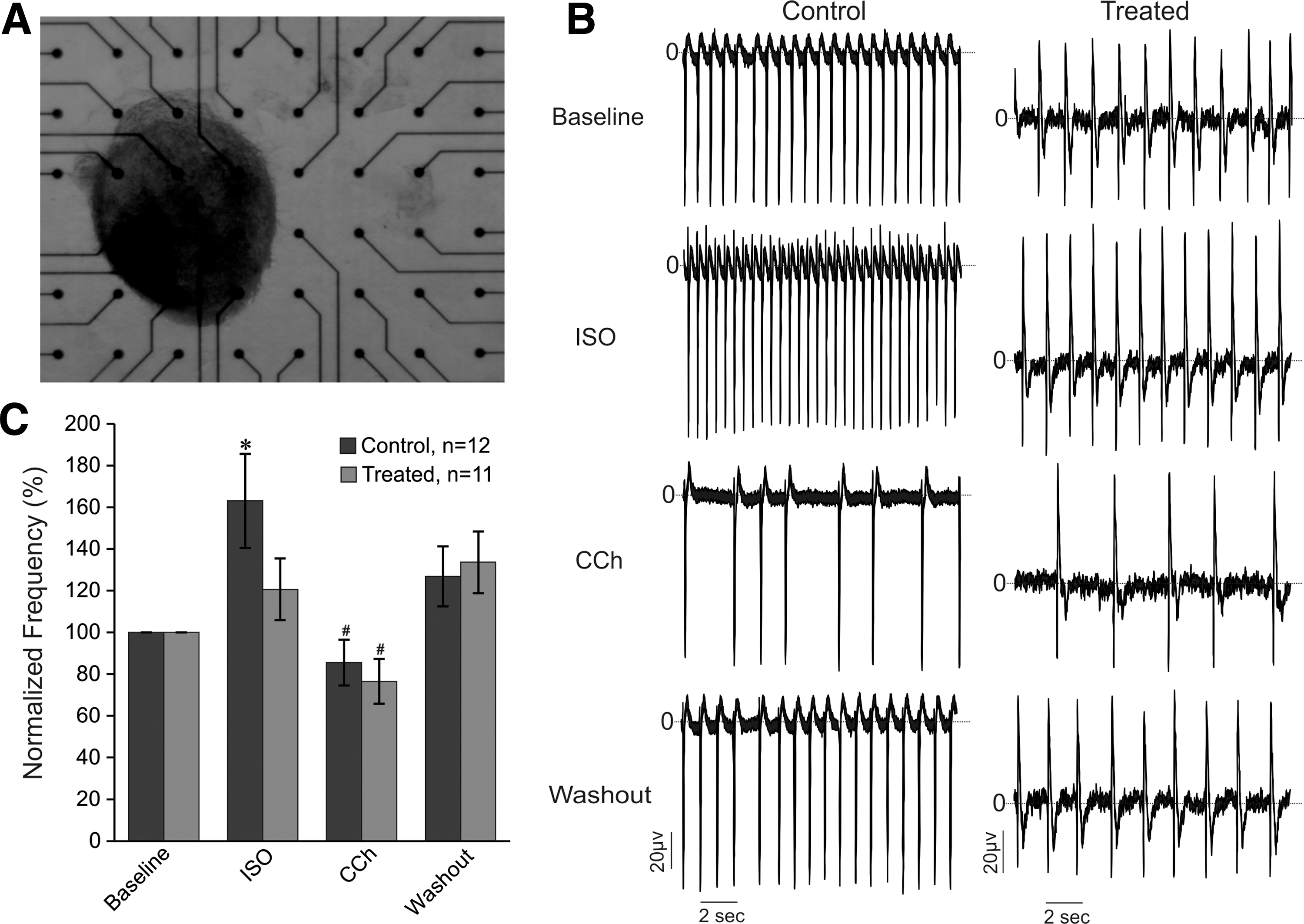
Electrophysiological analysis by MEAs.
Although we did not observe a significant change in ISO-mediated response in treated EBs as compared with control, it is possible that at the cellular level, β-AR responses are altered. We further evaluated the β-adrenergic pathway function in single beating myocytes exposed to ISO (1 μM) using patch-clamp measurement to record spontaneous APs. In control CMs (n = 9) and in CMs derived from the treated group (n = 8), we could measure APs of all three major CM types: nodal-like, atrial-like, and ventricular-like APs (data not shown), respectively, based on their shape and properties as described previously [25]. In single rES cell-derived CMs generated under control condition, the application of β-AR agonist ISO (1 μM) induced significant and partially reversible positive chronotropic effect on APs (increase of beating frequency to about 56%, n = 9, P < 0.05) (Fig. 4A, upper and Fig. 1B), whereas, in CMs generated under treated condition, the application of ISO (1 μM) did not induce any changes in AP beating frequency (Fig. 4A, bottom) as confirmed by the statistical analysis (Fig. 1B) (induced change of ∼3%, n = 8, P > 0.05), confirming at the single cellular level that the beta adrenergic pathway is also significantly affected or may be absent, whereas the muscarinic pathway remains unaffected in rES cell-derived CMs generated under DKK1, VEGF, and bFGF.

Effects of isoproterenol, forskolin, and IBMX on APs in rES cell-derived CM.
As β-ARs induce cAMP/PKA activation to regulate cardiac contraction, we therefore evaluated the functional expression of adenylate cyclase and phosphodiesterases (PDEs) as well as their association upon adrenergic stimulation on rES cell-derived CMs. As shown in Fig. 4C and D, the direct adenylate cyclase activator forskolin and the phosphodiesterase inhibitor IBMX increased in reversible manner the spontaneous beating frequency of the APs. In the presence of Forskolin (Fig. 4C), the AP frequency of rES cell-derived CMs from control and treated groups was increased to 151.72% ± 12.8% (n = 8; P < 0.05) and 136.73% ± 5.9% (n = 7; P < 0.05) of their initial value, respectively. On application of IBMX (Fig. 4D), the frequency of APs were also considerably increased to 152.90% ± 20.7% (n = 5; P < 0.05) and 151.35% ± 12.0% (n = 7; P < 0.05) of their baseline value, respectively. Taken together, this result suggests that PDEs and adenylate cyclase (AC) are constitutively active in rES cell-derived CMs generated under DKK1, VEGF, and bFGF.
Quantification of the β-ARs subtypes in rES cell-derived CMs
It was important to determine whether the unresponsiveness of rES cell-derived CMs generated under DKK1, VEGF, and bFGF to a β-AR agonist could be the decline in β-AR mRNA levels. We first determined β-AR mRNA levels by quantitative reverse transcription–polymerase chain reaction and found twofold higher levels (n = 3, P < 0.05) of β1-AR in the CMs from control compared with CMs from treated conditions (Fig. 5A, left), whereas no significant changes in β2-AR expression levels were observed (Fig. 5A, right).
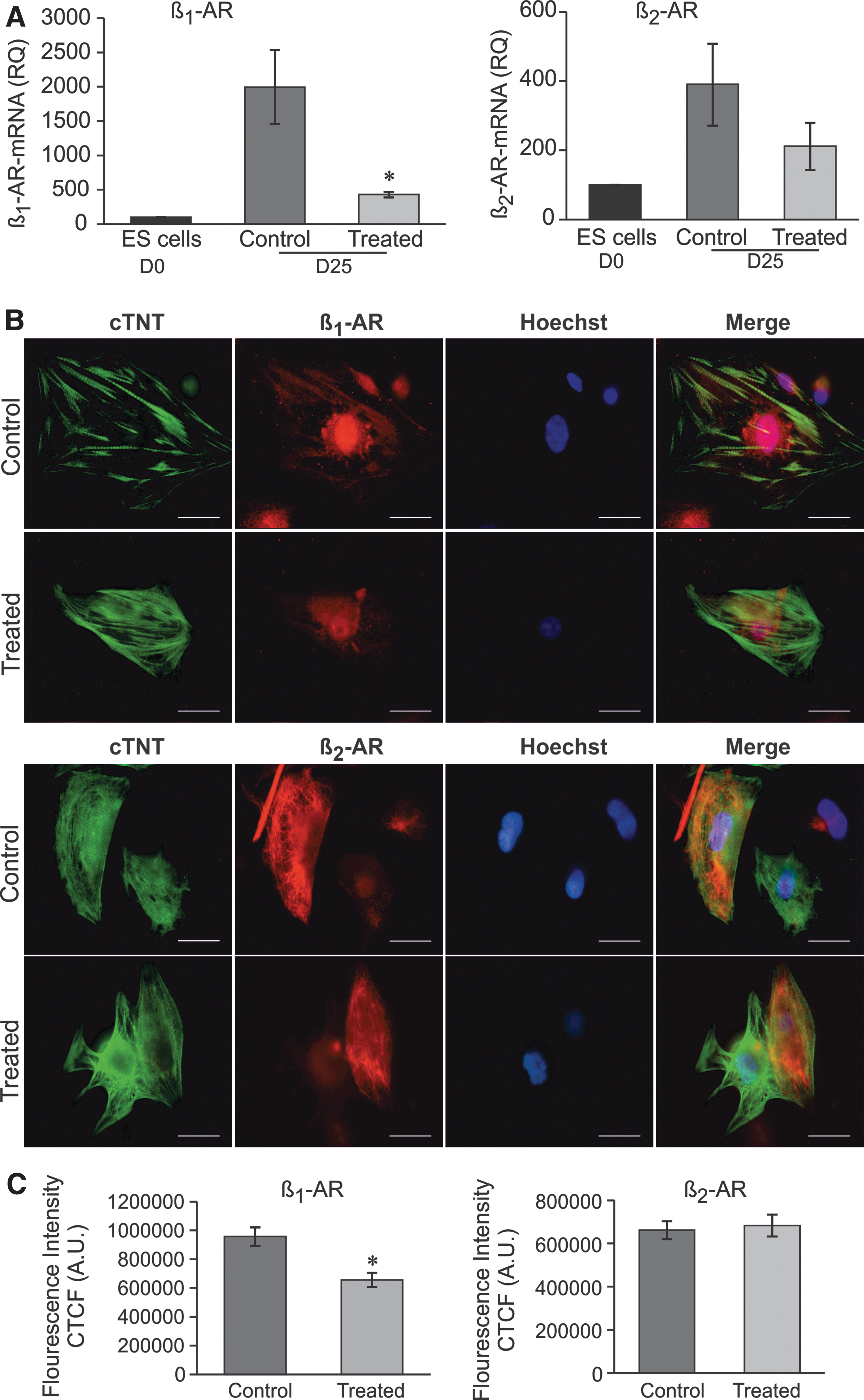
Assessment of β-AR expression in rES cell-derived CMs.
Next, to specify the discriminate expression of β-AR on the rES cell-derived CMs, rather than in the heterogeneous cell populations found in the EBs and to further investigate the quantification of β-AR subtypes, we performed double immunostaining with antibodies against cTNT and β1-AR and β2-AR. As shown in Fig. 5B (upper) and C (left), the expression of β1-AR in the differentiated CMs (average CTFC) of the control group was significantly higher than that of the treated group (n = 3, P < 0.05). In contrast, there was no difference between β2-AR expressions in CMs of control and treated groups (Fig. 5B, bottom and C, right), confirming the mRNA protein result obtained from quantitative PCR analysis.
Taken together, these results indicated that application of Wnt/β-catenin pathway inhibitor, DKK1, associated to VEGF and bFGF at certain time points during differentiation of rES cells into CMs, induce abnormal function and expression of the β-AR system.
Discussion
PSCs provide a chance to obtain a renewable source of healthy cells and tissues, such as CMs, because they are easily accessible and expandable in culture, have broad developmental potential and high capacity to reproducibly differentiate into CMs in in vitro condition. The main challenge of many investigators in recent years has been to develop in vitro methods for efficient cardiogenesis of PSC-CMs. Differentiation protocols currently used require optimization and CM yield shows high variability between different cell lines. Moreover, each individual stem cell line has its own differentiation capacity in culture depending on its endogenous and exogenous regulating signaling pathways [26]. In this study, using the protocol described in Materials and Methods section and developed previously [13], we show that Wnt/β-catenin pathway inhibitor, DKK1, associated to VEGF and bFGF, applied at certain time points during rES cell differentiation into CMs: (1) reduce the percentage of derived CMs, (2) leads to downregulation of β-AR of remaining CMs, and (3) impairs/desensitizes their β-AR signaling pathways. Several reports indicate that Wnt family members can act to enhance or inhibit cardiac differentiation [8,27]. For example, it has been show that Wnt/β-catenin signaling promotes cardiac differentiation at early developmental stages, and inhibits it later [8]. In addition, the presence of activin A and BMP4 at the early stage of differentiation (from day 0 to day 3), followed by the addition of Wnt inhibitor DKK1 and VEGF (from day 4 to day 7), and by the addition of a combination of bFGF and VEGF (from day 8 to day 14) significantly promote the differentiation of undifferentiated cells into CMs [9,28]. The presence of DKK1 in human ES cells culture medium during differentiation between day 5 and day 11 was previously shown to increase the efficiency of CMs [9]. Other studies showed that continuous application of DKK1 gave conflicting results on different cell lines. For example, the continued presence of DKK1 during the differentiation of CGR8 mouse ES cells enhances the percentage of CMs [29], but the same DKK1 treatment on A2lox and MC50 mouse ES cells suppressed the cardiac differentiation [30]. Therefore, stage-specific inhibition of Wnt/β-catenin pathway and activation of BMP and FGF signaling significantly improve cardiac lineage specification and subsequently increase of cardiac differentiation [8,11,13,31]. Signals initiated by BMPs, Wnts, and FGFs are first necessary to generate cardiac mesoderm-like cells that express specific markers [32]. Consequently, basic inhibition of the Wnt/β-catenin pathway by DKK1 is afterward required for the expression of genes affiliated with mesoderm and mesodermal derivatives [30,33]. As indirect proof for the importance of Wnt/β-catenin-mediated gene expression, we found that inhibition of the Wnt/β-catenin pathway by DKK1 significantly decreased the expression level of important mesodermal markers, GATA4 and NKX2.5, and late cardiac markers, α-MHC and TnT, suggesting a partial impairment in mesodermal specification of rES cells. The consequence of the reduced gene expression level listed above is the significant decrease in both the number and the function of CMs within the EBs, confirming the possibility of direct or indirect interaction of DKK1 with regulators involved in cardiac lineage either by blocking the activities of Wnt/β-catenin and possibly through other unknown mechanism. Most of these genes have been shown to work synergistically to coordinate the complex system and network leading to the function of CMs through mechanisms known to initiate, activate, and maintain the spontaneous beating activity as well as to modulate the maturity of CMs [6,34].
As critical determinant of normal CM function is the intact response to hormones and transmitters of the central nervous system [6], we further assessed the functional integrity of the remaining rES cell-CMs generated under DKK1. Surprisingly, the application of β-adrenergic agonist ISO (1 μM) did not affect the beating rate in CMs generated under DKK1, whereas the subsequent addition of CCh (1 μM), an acetylcholine analog receptor stimulation induced a significant negative chronotropic effect. This result would suggest at least in part a defective and/or uncoupling of β-adrenergic signaling cascade in remaining CMs. There are many lines of evidence that lead to the conclusion that β-AR systems play an important role in cardiac development and function [35 –37]. Further investigations in ES cells demonstrated that β-ARs play a role in the differentiation process of CMs from PSCs through ERK and p38 activation [38,39]. β1-ARs and β2-ARs are expressed at different stages of cardiac differentiation both at mRNA and protein levels. It is assumed that β2-AR and β1-AR subtypes are predominantly expression subtypes during the early and late stages of cardiac differentiation, respectively [39]. Although both subtypes share some signaling pathways, each has been shown to have specific signaling and regulatory properties [40]. In normal conditions, stimulation of CMs with β-AR agonist induces a signaling cascade that results in a rapid increase of cAMP levels in the cells [36]. This activates cAMP/PKA, which in turn not only modifies the cardiac excitation and contraction, but also other important cellular mechanism and functions, such as gene and protein expressions, growth, and can also lead to cell death [35,41]. We therefore conclude that the inhibition of cardiomyogenesis of rES cells observed with DKK1 in our study is most likely dependent on reduced β-AR expression and signaling. Besides this inhibition, we further provide evidence for alterations in the expression of β-ARs in CMs generated under DKK1, with downregulation of β1-AR than β2-AR as compared with control CMs. In line with these findings as also previously described [42], IBMX and forskolin, which increase the level of cAMP by inhibiting and stimulating of phosphodiesterase and AC, respectively, were found to increase the beating activity of CMs from treated and control conditions, suggesting that this signaling pathways is still functionally intact in remaining CMs generated under DKK1. Chronic human heart failure has been shown to be characterized by marked abnormalities in β-AR signaling that result mostly from a number of alterations, such as a 50% reduction in β1-ARs without change in β2-AR density as well as significant increased levels of the inhibitory G protein [43,44]. PSCs deprived of early Wnt/β-catenin signaling failed to generate cardiomyogenic mesoderm in long-term assays [30], which would again support our finding.
Taken together, we assume that downregulation of β-adrenoreceptors that we observed during cardiomyogenesis may at least in part be responsible for the lower response of CMs to isoproterenol exposure. One could also describe this phenomenon as β-adrenoreceptor desensitization, which is mainly characterized by β-AR molecular changes that induce the reduction of receptor density and their internalization as in the case of heart failure [45]. However, further investigation will help clarify this motion. Thus, our data not only reflect, but also confirm a crucial dependence on Wnt/β-catenin signaling for the initiation of a complete program of cardiac differentiation in vitro as previously revealed [29,30]. Our finding also confirms the importance of β-AR in cardiomyogenesis as well as in arrhythmogenesis. These findings could enhance future exploitation of ES cells and induced pluripotent stem cells to study the molecular and cellular processes of heart diseases and to develop possible strategies for cardiac regeneration.
Footnotes
Acknowledgments
The authors thank Florence Wianny and Colette Dehay (Institut cellule souche et cerveau INSERM: U846, Université Claude Bernard—Lyon I, Centre de recherche Inserm 18) for providing us with rhesus monkey ES cells. They acknowledge the skillful technical assistance of Annette Köster and Susan Rohani. They also thank Matthias Matzkies for the constructive discussions. This research work was conducted under the support of the German Academic exchange service (DAAD) through the scholarship offered to Mrs. Nermeen Eldabah.
Author Disclosure Statement
No competing financial interests exist.