
Abstract
Immune thrombocytopenia (ITP) is an autoimmune disease in which dendritic cells (DCs) play a crucial role in the breakdown of self-tolerance. Studies have identified the function of mesenchymal stem cells (MSCs) in promoting the development of regulatory DCs (regDCs). Our previous work revealed that MSCs in ITP exerted senescence, apoptosis, and impaired immunosuppressive effects on T and B cells. However, it is unclear whether the effects of MSCs on regDC induction are altered in ITP. Our data demonstrated that MSCs in ITP were impaired in inhibiting CD1a+ DC and CD14+ DC differentiation from CD34+ hematopoietic progenitor cells (CD34+ HPCs). DCs differentiated with MSCs in ITP exhibited an increased expression of costimulatory molecules CD80/CD86 and secretion of proinflammatory interleukin-12 (IL-12). Accordingly, the tolerogenic characteristics were deficient in DCs induced by MSCs in ITP. DCs differentiated with MSCs in ITP exhibited an impaired ability to inhibit CD3+ T cell proliferation, to suppress T helper (Th)1 cell differentiation, and to induce anergic and regulatory T cells (Tregs). The expression of Notch signaling components was measured in MSCs in ITP. Reduced expression of the ligand Jagged-1, the receptor Notch-1 intracellular domain (NICD-1), and the target gene Hes-1 was identified in MSCs in ITP. The addition of biologically active Jagged-1 to CD34+ HPCs was observed to promote regDC differentiation. When cultured on Jagged-1-coated plates, MSCs in ITP showed an enhancement of the Notch-1 pathway activation, Jagged-1 expression, and the function in inducing regDCs. Pretreatment with all-trans retinoic acid (ATRA) was found to partially restore the capacity of MSCs in both ITP patients and healthy controls in inducing CD34+-derived regDCs. Our data elucidated that MSCs in ITP were impaired in inducing CD34+-regDCs, associated with the Notch-1/Jagged-1 signaling pathway. ATRA could partially correct the impairment of MSCs, suggesting that ATRA could serve as a potential therapeutic alternative for ITP.
Introduction
I
Dendritic cells (DCs), the most potent antigen-presenting cells (APCs) [6], play an important role in initiating and controlling the magnitude and quality of adaptive immune responses [7]. In steady state, DCs induce peripheral tolerance through mechanisms such as the secretion of soluble factors, the clonal deletion of autoreactive T cells, and feedback control of Tregs [8]. However, DCs in ITP have been shown to have an increased capacity to stimulate autologous T cell proliferation and a reduced ability to convert T cells into Tregs [9 –11]. Additional studies have reported that DCs from ITP patients increased B cell proliferation and antibody production through B lymphocyte stimulator (BlyS) [12,13]. These reports indicated that DCs were functionally deficient in maintaining self-tolerance and may play a pathogenic role in ITP.
Mesenchymal stem cells (MSCs) are multipotent cells that play an important role in immune tolerance [14]. Despite their direct immunosuppressive effects on immune cells of the adaptive immune system [15], they are capable of modulating the T cell responses indirectly by promoting the tolerogenic properties of DCs. In particular, MSCs have been found to inhibit the differentiation, maturation, and function of DCs derived from CD34+ hematopoietic progenitor cells (HPCs) [16,17] or monocytes [18 –20]. In addition, it has been shown that MSCs induced mature DCs (mDCs) into a regulatory DC (regDC) population [21,22]. In our previous study, we found that MSCs in ITP displayed enhanced senescence and apoptosis, along with an impaired ability to inhibit T cell proliferation, induce Tregs, and suppress anti-GPIIb-IIIa antibody production [23]. However, less information is available on the immunosuppressive effects of senescent and apoptotic MSCs on DCs in ITP. Following our previous work identifying a pathogenic role of MSCs in ITP [23], we have continued the study and further investigated the immunomodulatory property of MSCs on DCs.
Our data revealed the impaired function of MSCs from ITP patients in inducing regDCs. The impairment may be correlated with the downregulation of the Notch-1/Jagged-1 signaling pathway and possibly be corrected with all-trans retinoic acid (ATRA). The results will help us better understand the pathogenesis of ITP and the potential therapeutic efficacy of ATRA in ITP patients.
Materials and Methods
Patients
Thirty-two newly diagnosed primary ITP patients were enrolled in this study. The diagnosis of ITP patients was based on previously reported criteria [24]. Twenty age- and sex-matched healthy donors were recruited as normal controls. This study was approved by the Medical Ethics Committee of Peking University People's Hospital. Informed consent was obtained from all patients in accordance with the Declaration of Helsinki.
Isolation and culture of MSCs
MSCs from ITP patients (ITP-MSCs) and healthy controls (control-MSCs) were isolated and cultured as previously described [22], with minor modifications. Briefly, bone marrow mononuclear cells (MNCs) were isolated by Ficoll (specific gravity 1.077 g/mL; Sigma, St. Louis, MO) gradient centrifugation and cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA), supplemented with 10% defined fetal calf serum (FCS; Gibco) and 100 U/mL penicillin/streptomycin. The cultures were maintained at 37°C in a 5% CO2 incubator, and the medium containing nonadherent cells was replaced every 3–5 days of culture. Once the cells reached 80% confluence, they were detached with 0.25% trypsin–EDTA (Sigma). MSCs at passage 3 were used for coculture experiments. The control-MSCs and ITP-MCSs were of a similar confluence and passage in each experiment. MSCs were identified by immunophenotypic criteria based on the expression of CD73, CD90, and CD105 and the absence of CD45, CD34, CD14, CD19, and HLA-DR expression [25]. Adipogenic, osteogenic, and chondrogenic differentiation was induced as described previously [25]. Culture supernatants were collected and stored at −20°C for future use.
Generation of human CD34-DC
CD34+ HPCs from healthy donors were purified from bone marrow MNCs using magnetic-activated cell sorting (MACS) and microbead-conjugated CD34 Abs according to the manufacturer's recommendations (Miltenyi Biotec, Bergisch Gladbach, Germany). CD34+ cells were plated in 24-well culture plates (Corning, Cambridge, MA) in RPMI 1640 culture medium (Gibco) supplemented with 10% FCS, 100 ng/mL granulocyte-macrophage colony-stimulating factor, 20 ng/mL tumor necrosis factor-alpha, and 200 ng/mL Flt3-L (Gibco). Cultures were incubated at 37°C in a 5% CO2 atmosphere. At days 3 and 5, fresh medium and cytokines were added to the cultures. At day 8, the cells were induced to mature by exposure to 100 ng/mL lipopolysaccharide (LPS) for 48 h. In some experiments, recombinant human Jagged-1 (R&D Systems, Minneapolis, MN) was added to CD34+ HPCs to a final concentration of 2 μg/mL to investigate its effects on DC differentiation.
Coculture experiment
Cocultures were initiated by seeding MSCs in 24-well plates 24 h before the coculture at a density of 5 × 104 cells per well in RPMI 1640 medium as described previously [16]. CD34+ HPCs (2.5 × 105 cells/well) were then seeded on top of the MSC layer at a 5:1 monocyte/MSC ratio. The following day, differentiation of DCs was achieved using the method described in the Generation of Human CD34-DC section. Cytokines were added to the cultures at days 0, 3, and 5. At day 8, DCs were separated from MSCs by centrifugation and washing. CD1a+ DCs and CD14+ DCs were purified by flow cytometry-based cell sorting with a fluorescence-activated cell sorter (FACS) Aria cytometer (BD Biosciences) to a purity of >95%. DCs were induced to mature with LPS in the absence of MSCs.
FACS analysis
For immunophenotype analysis, cultured cells were washed with phosphate-buffered saline (PBS) and incubated with primary antibodies for 30 min in the dark at room temperature. We used same-species, same-isotype irrelevant antibody as negative control. Cell analysis was performed using an FACS Calibur system (BD Bioscience) with Flowing software. For the analysis of Th1/Th2 or Tc1/Tc2 differentiation, the harvested CD3+ T cells were washed and stained with monoclonal antibodies (mAbs) specific for CD3 plus CD4 or CD3 plus CD8 for 30 min in the dark at room temperature. After washing with PBS, the cells were fixed and permeabilized for 15 min at room temperature. Following further washing, the cells were stained with mAbs against interferon-γ (IFN-γ) and interleukin-4 (IL-4) for 30 min in the dark at room temperature. For Treg differentiation analysis, the harvested CD3+ T cells were membrane stained with CD3 or CD4 and CD25 antibodies. Intracellular staining of FoxP3 and transforming growth factor-β (TGF-β) was performed as described for IFN-γ staining. Abs used for the FACS analyses included anti-CD1a-PE, anti-CD14-FITC, anti-CD80-PE, anti-CD86-PE, anti-CD83-APC, anti-HLA-DR-FITC, anti-CD3-FITC, anti-CD4-PE, anti-CD8a-PE, anti-IFN-γ-PerCP-Cyanine5.5, anti-IL-4-APC, anti-Foxp3-APC, and anti-Notch-3-APC antibodies, all purchased from eBioscience. In addition, anti-TGF-β1 [2Ar2] (Abcam, Cambridge, MA), Alexa Fluor® 647 AffiniPure donkey anti-rabbit IgG (H+L) (Jackson, West Grove, PA), anti-Jagged-1-FITC (LifeSpan, Seattle, WA), anti-DLL-1-APC (R&D Systems), anti-Notch-1-PE (BD, San Diego, CA), anti-Notch-2-PE (BD), and anti-Notch-4-Alexa Fluor 647 (BD) antibodies were also used in the FACS analyses.
Endocytosis assay
Endocytosis was measured as the cellular uptake of fluorescein isothiocyanate (FITC)-CM-dextran and was quantified by flow cytometry. Approximately 5 × 105 cells per sample were incubated in medium containing FITC-CM-dextran (10 μg/mL; Sigma) at 37°C for 30 min. After incubation, the cells were washed twice with cold PBS to stop endocytosis and remove excess dextran. The quantitative uptake of FITC-CM-dextran by the cells was determined by FACS. At least 10,000 cells per sample were analyzed.
Cytokine analysis
Cytokines produced in the culture supernatants were measured using enzyme-linked immunosorbent assay kits (R&D Systems) for IL-10, IL-12, IL-2, IL-6, TGF-β, CCL-2, and PGE-2.
Isolation and labeling of CD3 + T cells
Human peripheral blood mononuclear cells (hPBMCs) from healthy donors were isolated by Ficoll-Paque (1.077 g/mL; Sigma) density gradient centrifugation. CD3+ T lymphocytes were isolated from hPBMCs using CD3 micromagnetic beads (Miltenyi Biotec) according to the manufacturer's instructions. For the production of carboxyfluorescein diacetate succinimidyl ester (CFSE; Sigma)-labeled CD3+ T cells, CD3+ T cells were labeled with CFSE at a concentration of 5 μM for 10 min at 37°C. The labeling reaction was quenched by adding cold FCS and incubating for 5 min on ice. The cells were washed three times with RPMI 1640 culture medium with 10% FCS before being added to the cocultures.
CD3 + T cell proliferation assay
In the assay testing, the immunogenicity of DCs, CFSE-labeled CD3+ T cells (2 × 106/well) were cocultured with allogeneic mDCs at a 1:10 DC/T cell ratio in 96-well plates for 5 days. The lymphocytes were harvested, and lymphocyte proliferation was assessed by the CFSE dilution method.
In the assay testing, the immunomodulatory potential of MSCs induced DCs (MSC-DCs), CFSE-labeled CD3+ T cells (2 × 106/well) were cocultured with mDCs+ control-MSCs induced DCs (control-MSC-DCs) and mDCs+ ITP-MSCs induced DCs (ITP-MSC-DCs). The cocultured cells were in a 10:1:1 ratio (T cell:mDC:MSC-DC). All cells were cultured in flat-bottomed 96-well plates for 5 days. Subsequently, T cells were collected and assessed for proliferation by FACS.
T cell differentiation assay
CFSE-labeled CD3+ T cells were maintained in the absence of stimulation or cocultured with allogeneic DCs at a 1:10 DC/T cell ratio for 5 days in RPMI 1640 supplemented with 10% FCS. On day 5, lymphocytes were MACS purified and analyzed for Th1/Th2, Tc1/Tc2, and Treg differentiation by FACS.
T cell anergy assay
CFSE-labeled CD3+ T cells were cultured alone or cocultured with allogeneic mDCs, ITP-MSC-DCs, or control-MSC-DCs at a 1:10 DC/T cell ratio for 5 days in RPMI 1640 supplemented with 10% FCS. On day 5, lymphocytes were MACS purified and restimulated with immunostimulatory allogeneic mDCs from the same donor as in the primary culture at a 1:10 DC/T cell ratio for 5 days in 96-well plates. On day 5, lymphocytes were harvested and analyzed for proliferation by FACS.
Recombinant Jagged-1-coated culture plates
The human Jagged-1 recombinant protein (R&D Systems) contains the signal peptide and extracellular domain of Jagged-1 fused at the C terminus to the Fc portion of human IgG. Briefly, culture plates were coated with anti-human IgG (10 μg/mL; Sigma) in PBS at 4°C overnight and incubated in a solution containing recombinant Jagged-1 protein (10 μg/mL) at 4°C overnight. The same concentrations of human IgG (Sigma) were used to coat plates as controls. ITP-MSCs were cultured on Jagged-1- or IgG-coated plates for 48 h before coculture with CD34+ HPCs.
ATRA treatment of MSCs
To determine the optimal concentration of ATRA in promoting the immunomodulatory function of MSCs, ITP-MSCs were seeded on six-well culture plates (Corning) in DMEM supplemented with 10% FCS to 70% confluence, and ATRA (Sigma) at concentrations of 10−9, 10−8, 10−7, and 10−6 M was added to the medium. After the optimal concentration of ATRA was identified, ITP-MSCs and control-MSCs were preincubated in DMEM containing 10% FCS plus 10−7 M ATRA (ATRA-ITP-MSCs and ATRA-control-MSCs) in humidified air in 5% CO2 at 37°C for 24 h, and then, cells were washed to remove ATRA. ATRA-treated MSCs were harvested and cocultured with CD34+ HPCs to induce DC differentiation (ATRA-ITP-MSC-DCs and ATRA-control-MSC-DCs).
Quantitative real-time polymerase chain reaction
Total RNA from cells was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA). RNA was transcribed into cDNA using the PrimeScript™ RT reagent kit (Takara, China) according to the manufacturer's instructions. An amount of 500 ng DNase-treated RNA was amplified with gene-specific primers using the one-step Power SYBR Green RNA-to-Ct kit (Applied Biosystems, Foster City, CA) on a LightCycler 480 II (Roche, Shen Chang Road, Shanghai). Gene expression was normalized to a housekeeping gene (18s rRNA) for relative quantification. Gene-specific primers are listed in Table 1.
qRT-PCR, quantitative real-time polymerase chain reaction.
Western blotting
For western blotting, equivalent amounts of protein lysates, obtained from induced cells, were loaded per lane. After sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), proteins were electrophoretically transferred onto nitrocellulose membrane (Millipore, Billerica, MA). After blocking, blots were incubated with rabbit mAbs against Notch-1 intracellular domain (NICD-1) and Hes-1 (CST, Danvers, MA) at a dilution of 1:500. Expression of the glyceraldehyde phosphate dehydrogenase (GAPDH) was used as an internal control. Immunodetection using the enhanced chemiluminescence method (ECL kit) was performed according to the manufacturer's instructions.
Statistical analysis
Data are expressed as mean ± standard deviation (SD) or median ± interquartile range and statistically analyzed using the SPSS version 20.0 statistical package (SPSS, Inc., Chicago, IL). Statistical significance among three or more groups was determined by ANOVA (analysis of variance), and the difference between two groups was determined by Sidak's multiple comparison correction test, unless the data were not normally distributed, in which case the Kruskal–Wallis test and Mann–Whitney U test were used. Differences between pre- and post-treatment groups were determined by paired Student's t-test, and the difference between two independent groups was compared using the nonpaired t-test, unless the data were not normally distributed, in which case the Wilcoxon matched-pairs test and Mann–Whitney test were used. P < 0.05 was considered statistically significant.
Results
ITP-MSC and CD34-DC differentiation
CD34+ HPCs were induced to differentiate into immature DCs (imDCs) in the presence of control-MSCs or ITP-MSCs. At the end of the culture, CD34+ HPCs were observed to expand 15.2 ± 1.7-fold (mean ± SD, n = 12), 3.6 ± 1.0-fold (mean ± SD, n = 14), or 14.6 ± 2.8-fold (mean ± SD, n = 14) when cultured alone, with control-MSCs, or with ITP-MSCs, respectively (Fig. 1A). Phenotypic analyses showed that control-MSCs strongly inhibited the differentiation of CD34+ HPCs into CD1a+ DC and CD14+ DC, whereas the ITP-MSC failed to suppress DC differentiation (Fig. 1B, C). The endocytic activity was further evaluated, and only control-MSC-DCs exhibited increased endocytosis (Fig. 1D, E).
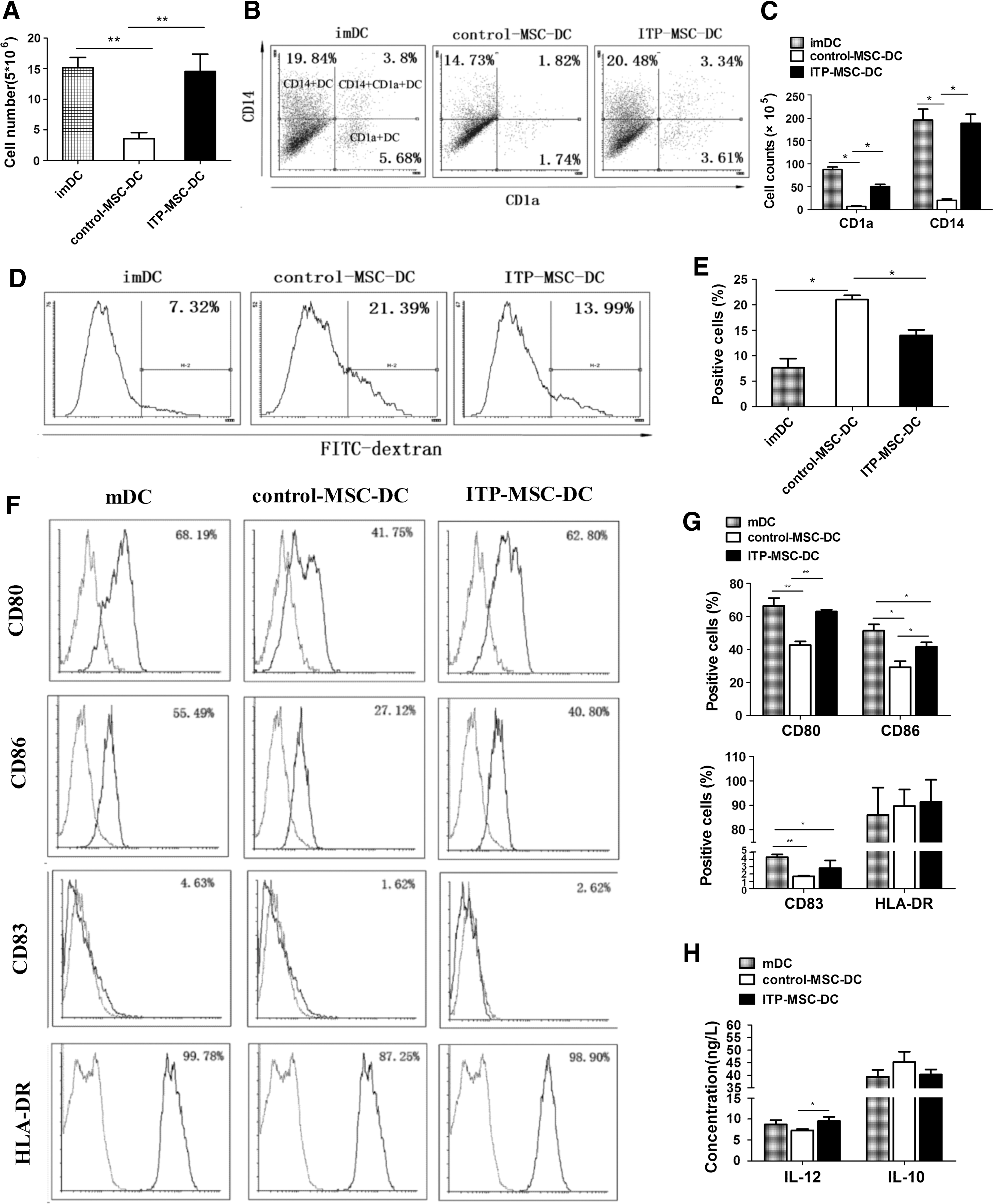
ITP-MSCs show an impaired ability in inducing CD34+ HPC-derived DC differentiation and maturation. CD34+ HPCs were induced to differentiate into DCs in the presence of control-MSCs (n = 16) or ITP-MSCs (n = 16). On day 8, imDCs were harvested and induced to maturation by LPS for two additional days without MSCs.
mDCs differentiated in the presence of ITP-MSCs were characterized. As shown in Fig. 1F and G, control-MSCs significantly inhibited the expression of costimulatory molecules CD80/CD86 on mDCs, whereas the inhibitory capacity of ITP-MSCs was dramatically impaired. The suppressive function of ITP-MSCs in the expression of maturation marker CD83 on mDCs was comparable to that of control-MSCs (Fig. 1F, G). No difference was found between control-MSC-DCs and ITP-MSC-DCs in the expression of HLA-DR (Fig. 1F, G). The cytokine secretion of ITP-MSC-DCs was also analyzed (Fig. 1H). ITP-MSC-DCs were found to exhibit significantly increased production of proinflammatory IL-12 compared with control-MSC-DCs. The secretion of anti-inflammatory IL-10 was unaffected by control-MSCs or ITP-MSCs. These results demonstrated that ITP-MSCs showed deficient suppressive effects on CD34+ HPC-derived DC differentiation and maturation.
ITP-MSC-DC immunomodulatory effects on CD3+ T cells
In addition to the impaired effects on DC differentiation, whether ITP-MSCs induced a DC subset with tolerogenic function remained to be determined. As shown in Fig. 2A, control-MSC-DCs could hardly stimulate allogeneic CD3+ T cell proliferation, whereas ITP-MSC-DCs induced CD3+ T cell proliferation significantly. While control-MSC-DCs distinctly suppressed CD3+ T cell proliferation stimulated by mDCs, ITP-MSC-DCs could only slightly inhibit mDC-activated CD3+ T cell proliferation (Fig. 2A). We then sought to investigate the effects of ITP-MSC-DCs on T cell differentiation (Fig. 2B, C). Control-MSC-DCs were observed to decrease IFN-γ expression levels in CD4+ T cells, whereas ITP-MSC-DCs increased IFN-γ expression levels, implying an impaired ability of ITP-MSC-DCs to inhibit Th1 differentiation. Moreover, it was found that ITP-MSC-DCs lost the ability to promote Th2 differentiation. Similarly, ITP-MSC-DCs displayed a remarkably suppressed capacity to instruct CD8+ T cell differentiation toward Tc2 compared with control-MSC-DCs.

ITP-MSC-DCs were deficient in maintaining the tolerance of CD3+ T cells.
To determine whether ITP-MSC-DCs induced anergy in CD3+ T cells, the proliferative response and IL-2 secretion of the CD3+ lymphocytes were detected in a two-step culture system (Fig. 2D, E). Compared with mDC-primed CD3+ T cells, CD3+ T cells primed with control-MSC-DCs exhibited a significantly weaker response to further stimulation with immunostimulatory mDCs from the same donor as the primed cells, as evidenced by a lower level of proliferative response and IL-2 production. In contrast, CD3+ T cells in primary culture with ITP-MSC-DCs responded vigorously to restimulation with the same allogeneic antigen. For the induction of Tregs, the data showed that control-MSC-DCs induced a dramatically higher percentage of CD4+CD25+FoxP3+ (Fig. 2F, G) and TGF-β-producing Tregs (Fig. 2H, I) compared with mDCs. The proportion of CD4+CD25+FoxP3+ and TGF-β-producing Tregs induced by ITP-MSC-DCs was markedly reduced when compared with control-MSC-DCs (Fig. 2F–I). The data elucidated that ITP-MSC-DCs were deficient in maintaining CD3+ T cell tolerance, indicating ITP-MSCs may be impaired in inducing a regDC population.
Jagged-1 regulation of regDC differentiation
To clarify the mechanism(s) involved in the impaired function of ITP-MSCs in inducing regDCs, the expression of several factors involved in MSC-mediated immunomodulation was assessed. We first evaluated the expression levels of Jagged-1 and DLL-1 on the cell surface as previously described [21]. The mRNA level of Jagged-1 was dramatically reduced in ITP-MSCs, but no difference was observed in the expression of DLL-1 (Fig. 3A). There was also an obvious decrease in Jagged-1 expression in ITP-MSCs, as determined by FACS analysis (Fig. 3B). FACS analysis also revealed no obvious difference in the expression of DLL-1.
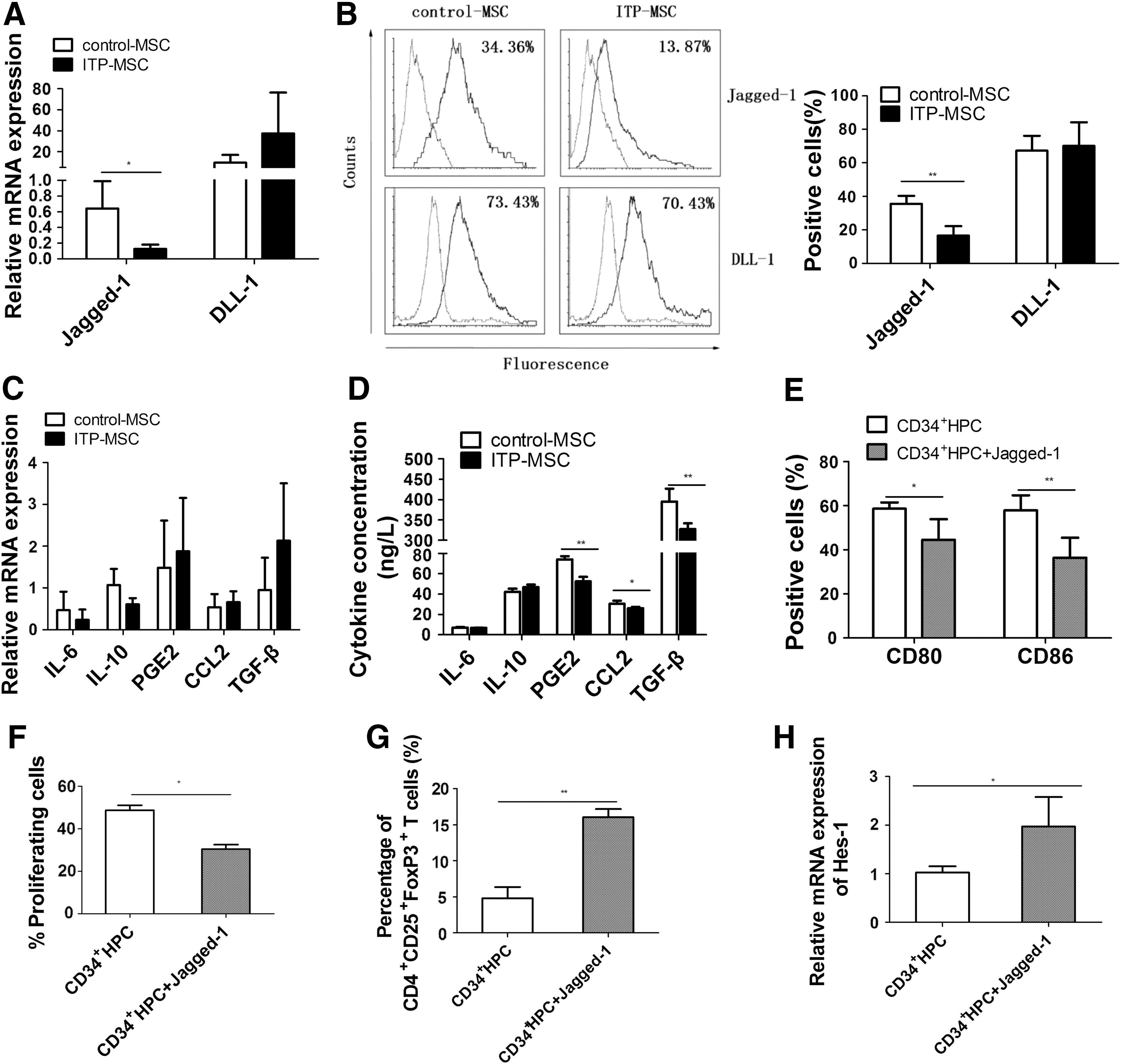
Jagged-1 may be associated with the impaired function of ITP-MSCs in inducing regDCs.
Although no significant difference of IL-6, IL-10, PGE2, CCL-2, and TGF-β expression was found between control-MSCs and ITP-MSCs (Fig. 3C), the data revealed a substantially decreased level of PGE2, CCL-2, and TGF-β secretion in ITP-MSC supernatants (Fig. 3D). Considering the dramatically decreased expression of Jagged-1 in ITP-MSCs both at the mRNA and protein levels, the function of Jagged-1 was examined in subsequent experiments. Biologically active Jagged-1 was then added when CD34+ HPCs were induced to differentiate into DCs. It was observed that Jagged-1 treatment significantly decreased the expression of CD80/CD86 on DCs (Fig. 3E). The added Jagged-1 substantially suppressed the allostimulatory potential of DCs on CD3+ T cells (Fig. 3F). The percentage of CD4+CD25+FoxP3+ Tregs induced by DCs was dramatically increased with the addition of Jagged-1 during DC differentiation (Fig. 3G). Moreover, the expression of Hes-1, the target gene of Notch signaling pathway, was substantially increased in DCs differentiated in the presence of Jagged-1 (Fig. 3H). These results indicated that Jagged-1 may activate the Notch signaling pathway in CD34+ HPCs to promote regDC differentiation.
Notch-1/Jagged-1 signaling pathway in inducing regDCs
To determine whether the Notch signaling pathway was impaired in ITP-MSCs, the expression of Notch pathway components was analyzed. There was no significant difference among Notch-1, Notch-2, Notch-3, and Notch-4 in ITP-MSCs and control-MSCs at the mRNA and protein levels (Fig. 4A, B). The expression of Hes-1, Hes-5, and Hey-1, the target genes of Notch signaling pathway, was further investigated. The results revealed a distinct decrease of Hes-1 expression level in ITP-MSCs (Fig. 4C). Considering the critical role of Notch-1 signaling pathway in the immunomodulatory capacity of MSCs, the expression of NICD-1 and Hes-1 was further tested by western blot. The data showed that the protein expression of Hes-1 and NICD-1 decreased substantially in ITP-MSCs (Fig. 4D, E). The potential role of Notch-1 signaling pathway in mediating MSC induction of regDCs was further investigated. It was observed that Notch-1 signaling pathway was activated in ITP-MSCs cultured on Jagged-1-coated plates, as manifested by a distinctly increased expression of NICD-1 (Fig. 5A) and Hes-1 (Fig. 5B, C) in ITP-MSCs. When ITP-MSCs previously cultured on Jagged-1-coated plates were cocultured with CD34+ HPCs, the differentiated DCs exerted a markedly reduced expression of costimulatory molecules CD80 and CD86 (Fig. 5D). Accordingly, the immunostimulatory potential of these DCs on allogeneic CD3+ T cells declined dramatically (Fig. 5E), while the capacity of inducing CD4+CD25+FoxP3+ Tregs increased substantially (Fig. 5F). In addition, Jagged-1 expression was found to enhance appreciably both at the mRNA and protein levels when ITP-MSCs were cultured on Jagged-1-coated plates (Fig. 5G), indicating that the Jagged-1-induced Notch-1 signaling pathway may, in turn, modulate Jagged-1 expression. The data implied that the Notch-1/Jagged-1 signaling pathway may be associated with the impaired function of ITP-MSCs in the induction of regDCs.
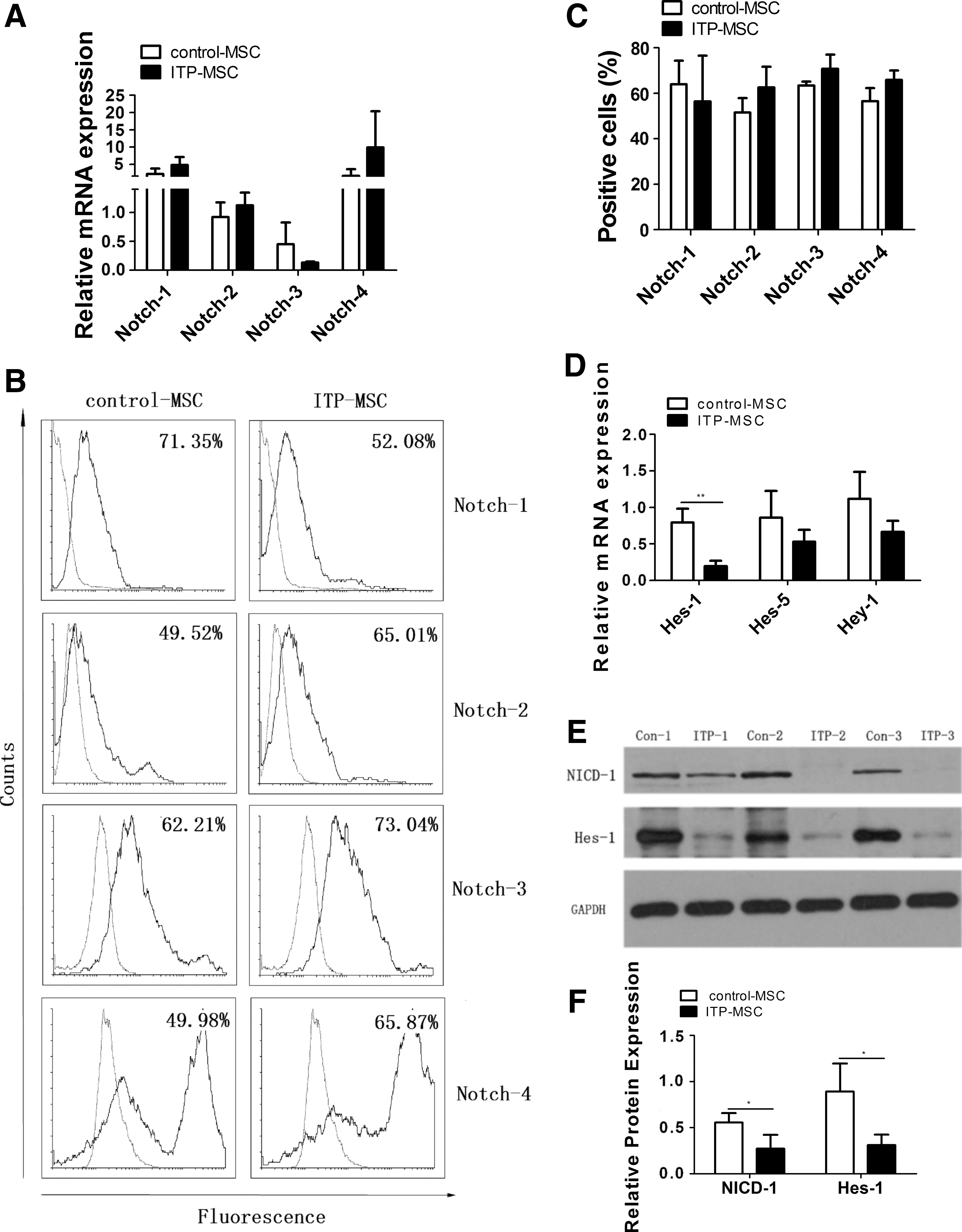
Downregulation of Notch-1 signaling pathway in ITP-MSCs.
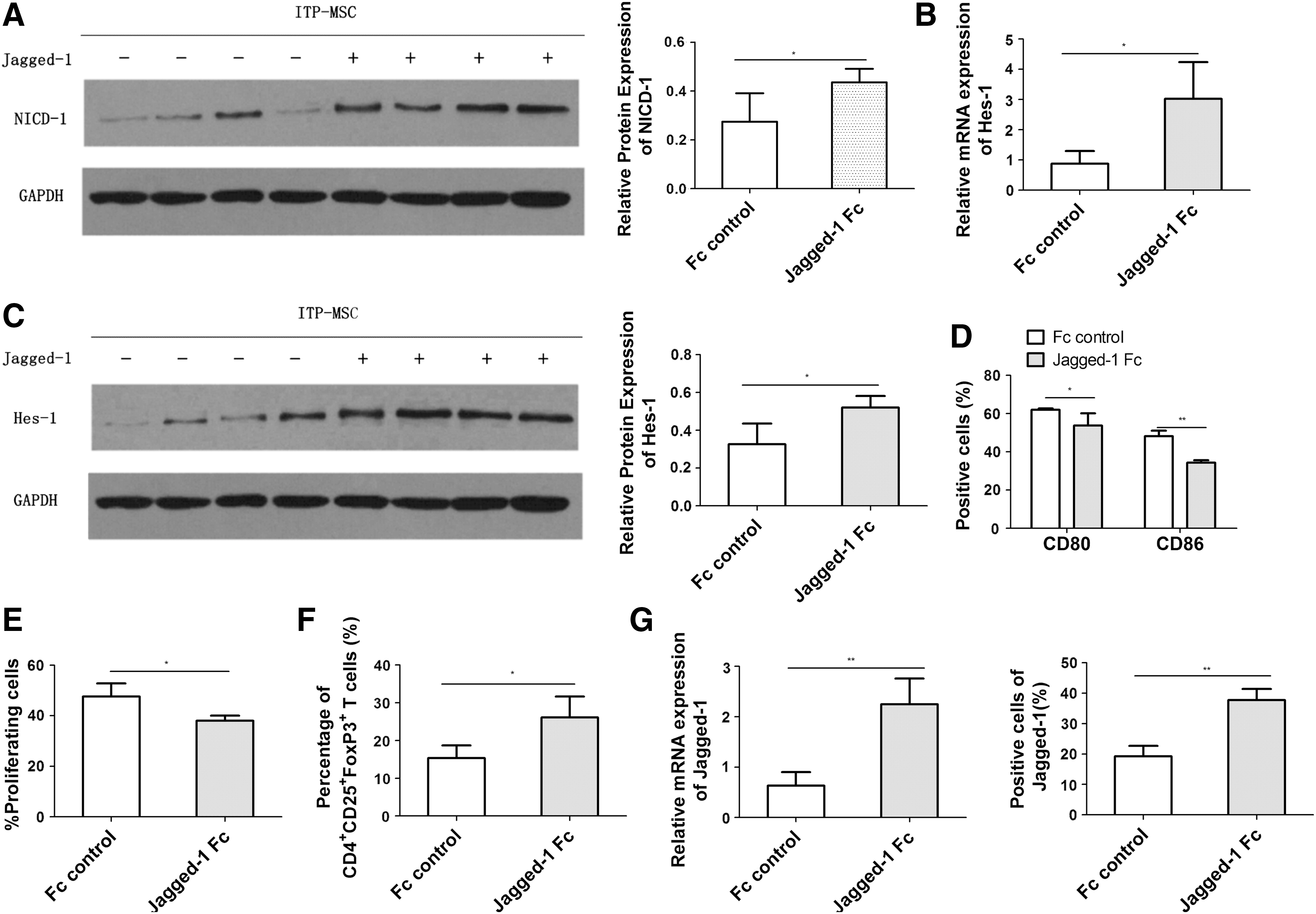
The Notch-1/Jagged-1 signaling pathway is possibly correlated with the impaired induction of regDCs by ITP-MSCs.
ATRA function of ITP-MSC induction of regDCs
ITP-MSCs were pretreated with different doses of ATRA (10−9 to 10−6 M) before culture with CD34+ HPCs. As shown in Fig. 6A, concentration gradients of ATRA from 10−9 to 10−7 M could significantly promote ITP-MSC-mediated inhibition of CD80 expression on DCs in a dose-dependent manner. Thus, a concentration of 10−7 M was adopted to treat MSCs in our work. With ATRA modulation, both control-MSCs and ITP-MSCs exerted increased capacity to inhibit CD80/CD86 expression on DCs (Fig. 6B). ATRA pretreatment enhanced the capacity of control-MSC-DCs to inhibit allogeneic CD3+ T cell proliferation, while it exerted no effects on ITP-MSC-DC-mediated inhibition of CD3+ T cell proliferation (Fig. 6C). There was a higher percentage of CD4+CD25+FoxP3+ Tregs in the ATRA-ITP-MSC-DC and ATRA-control-MSC-DC groups compared with that in ITP-MSC-DCs and control-MSC-DCs, respectively (Fig. 6D). The expression of NICD-1, Hes-1, and Jagged-1 was then determined. As shown in Fig. 6E, both ITP-MSCs and control-MSCs treated with ATRA demonstrated an increase in NICD-1 expression. The expression of the target gene Hes-1 in ITP-MSCs increased significantly after ATRA modulation, whereas control-MSCs exhibited no obvious change in Hes-1 expression with ATRA modulation (Fig. 6F). However, ATRA had no obvious effects on Hes-1 protein expression in control-MSCs and ITP-MSCs (Fig. 6G). The expression of Jagged-1 was found to be upregulated in ATRA-ITP-MSCs both at the mRNA and protein levels, whereas no obvious change was found in ATRA-treated control-MSCs (Fig. 6H). Therefore, ATRA modulation may be effective in correcting the impaired immunomodulatory capacity of ITP-MSCs in inducing regDC differentiation via the Notch-1/Jagged-1 pathway.
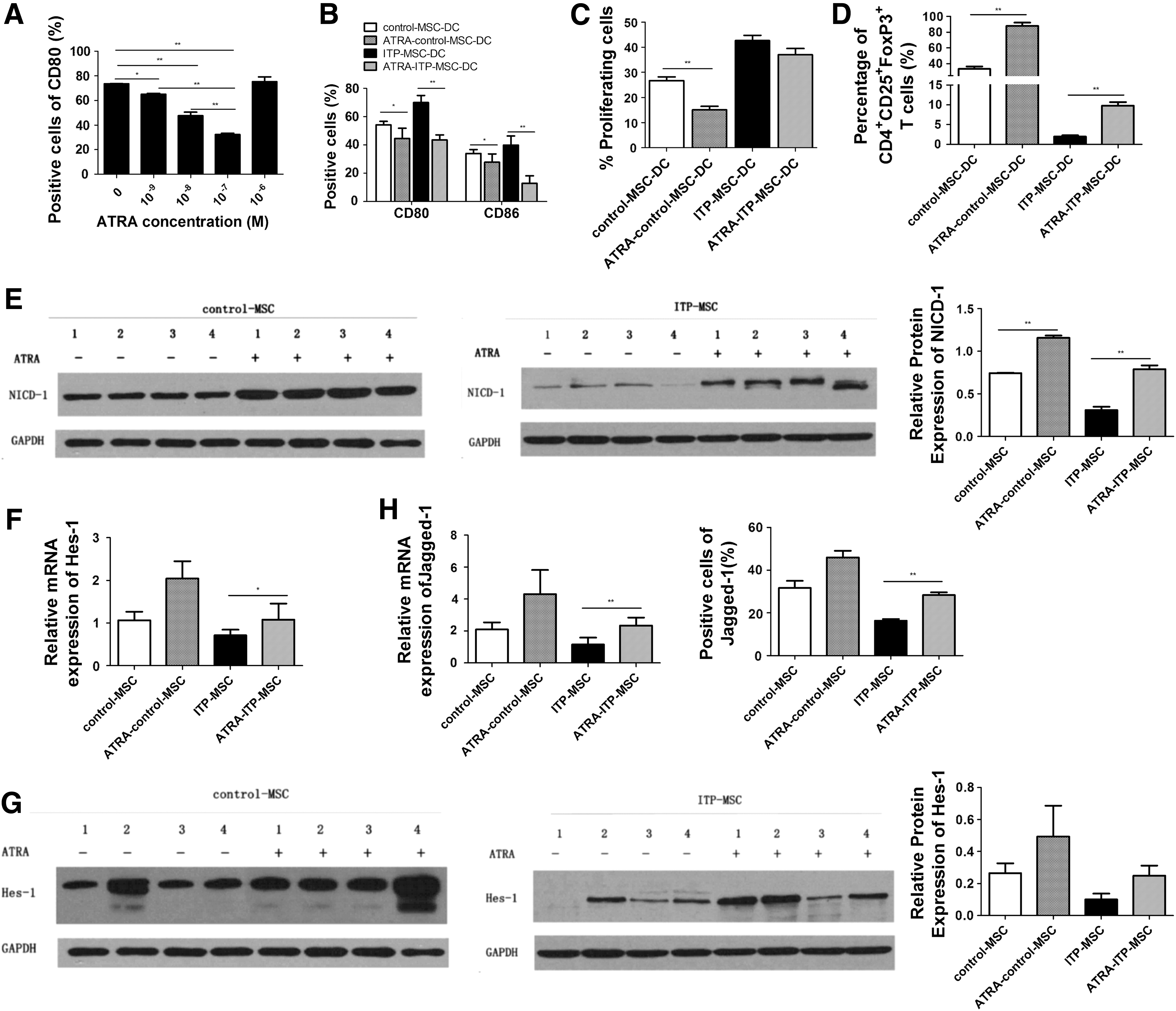
ATRA may improve the function of ITP-MSCs in inducing regDCs through activation of the Notch-1/Jagged-1 signaling pathway.
Discussion
In this study, we demonstrated the deficiency of MSCs in ITP in inducing regDC differentiation from CD34+ HPCs, which may be associated with the suppressed Notch-1/Jagged-1 signaling pathway. ATRA was found to play a role in restoring the insufficiency of MSCs in ITP via the Notch-1/Jagged-1 pathway.
ITP is an acquired autoimmune disorder in which both autoantibodies promoted by enhanced Th cell activity [26] and Tc cells [3] are potentially involved. DCs link innate and adaptive immunity and are crucial in inducing immunity and tolerance [6]. There has been increasing evidence implicating the central role of the imbalance between DC-mediated immune activation and tolerance in the pathogenesis of autoimmune diseases [27]. MSCs are multipotent cells [14] that exert important immunosuppressive effects on immune cells of the innate and adaptive immune systems, such as T lymphocytes [28,29], B lymphocytes [30,31], DCs [19], and macrophages [32,33]. The immunomodulatory effects of MSCs from ITP patients on DCs differentiated from the primitive CD34+ HPCs were examined in our study.
Our data elucidated that MSCs in ITP were impaired in inducing regDC differentiation from CD34+ HPCs. This finding is consistent with a previous report demonstrating the impaired ability of MSCs in ITP to induce tolerogenicity in mDCs [34]. MSCs in ITP failed to inhibit Langerhans CD1a+ DC and interstitial CD14+ DC differentiation. mDCs differentiated in the presence of MSCs in ITP exhibited a significant increase in the expression of CD80/CD86 and secretion of proinflammatory IL-12. The costimulatory molecules CD80/CD86 and cytokine IL-12 are the second and third signals utilized by DCs to regulate T cell responses [35]. RegDCs induced by different agents were observed to display a low expression of costimulatory molecules and a reduced production of IL-12 [36]. There are numerous, discrete populations of naturally occurring regDCs. Several kinds of regDCs have been reported that have similar functions but different phenotypes. It has been suggested that regDCs are a functional state rather than a unique subpopulation defined by phenotypical markers. In this study, coculture assays of CD34+ HPCs with control-MSCs, ITP-MSCs, or no MSCs were used to obtain regDCs as described previously [16,21]. These MSC-DCs can be considered regDCs because they expressed high IL-10 but low IL-12 mRNA levels, they inhibited the proliferation and function of reactive T cells, and they induced the generation of alloantigen-specific regulatory T cells expressing both FoxP3 and TGF-β mRNA. The function of DCs in maintaining CD3+ T cell immune tolerance was determined. RegDCs play an important role in the maintenance of immunological tolerance. Mechanisms through which regDCs promote tolerance include the induction of anergic and/or regulatory T cells, inhibition of T cell responses, induction of T cell apoptosis, and alteration of Th1/Th2 balance [37]. Anergic T cells are intrinsically functionally inactivated and remain hyporesponsive on the long term [38], whereas Tregs actively suppress the activation of immune cells [39]. The results revealed that DCs differentiated with MSCs in ITP vigorously stimulated CD3+ T cell proliferation in the absence or presence of mDCs, indicating an impaired ability to maintain low immunogenicity and high immunoregulatory potential. In addition, these DCs failed to induce anergic and regulatory T cells. In ITP, an increased Th1/Th2 ratio has been found to play a role in the initiation and/or perpetuation of the disease [40]. IL-12 is critical for directing T cell differentiation toward Th1 [41]. Consistent with the cytokine profiles, we found that DCs differentiated with MSCs in ITP displayed a deficient ability to instruct Th2 and Tc2 differentiation. In the endocytosis assay, we found that cells cocultured with ITP-MSCs have a decreased endocytic capacity compared with normal-MSCs. Considering that the imDC- or MSC-induced regDCs have strong endocytic capacity [21], decreased endocytosis in DCs cocultured with ITP-MSCs agreed with our results demonstrating that ITP-MSCs are deficient to inhibit DC mutation and induce regDC differentiation. Taken together, our findings are in agreement with recent reports illuminating the proliferative deficiency and immunomodulatory impairments of MSCs in ITP [42,43].
Notch signaling pathway is evolutionarily conserved and plays a crucial role in regulating many cellular processes during development and renewal of adult tissues [44,45]. The mammalian Notch system consists of four Notch receptors (Notch-1, -2, -3, and -4) and five ligands (Jagged-1, -2 and DLL-1, -3, and -4). Both Notch ligands and receptors are transmembrane proteins expressed on the cell surface [46]. The Notch receptor is activated and cleaved to release and translocate the NICD into the nucleus after ligand binding. NICD regulates its downstream target genes via both CBF-1/RBP-J, Su(H), Lag-1-dependent and CSL-independent pathways [47]. The genes of the Hes family and Hey family are important target genes in the Notch pathway [48]. In addition to the pivotal role of Notch signaling pathway in proliferation [49], multilineage differentiation [50], and multipotency maintenance [51] of MSCs, it has been reported to be involved in the immunomodulatory function of MSCs. Particularly, the Notch signaling pathway was shown to be required by MSCs to alleviate acute graft versus host disease in mice in vivo [52]. Studies also revealed a role for the Notch signaling pathway in regulating the immunosuppressive effects of MSCs on T cells and DCs in vitro [52,53]. Therefore, our results implicated the association of the Notch-1 pathway with MSC-mediated induction of regDCs in ITP, further broadening our understanding of the contributions of the Notch pathway to the immunomodulatory function of MSCs.
Although it has been reported that human MSCs enable CD34+ HPCs to differentiate into regDCs through activation of the Notch pathway [16], the details of the ligands expressed on MSCs to mediate regDC differentiation remain unknown. Cheng et al. have shown that bone marrow stroma expressed a substantially higher level of Jagged-1 and prevented the terminal differentiation of DCs via Jagged-1 [54]. Many studies utilizing cell lines transfected with Notch ligands have demonstrated that DLL-1 promoted the differentiation of myeloid DCs [55] and Langerhans cells [56], whereas the ligand Jagged-1 inhibited the differentiation of myeloid DCs and caused accumulation of immature myeloid cells [57]. In agreement with these reports demonstrating the inhibitory effect of Jagged-1 on DC differentiation, Jagged-1 was observed to be involved in inducing CD34+-regDCs by MSCs in our study. Several studies also found a crucial role of Jagged-1 in inducing tolerogenic DC population, in suppressing T cell proliferation, and in expanding CD4+CD25+FoxP3+ Tregs [58 –60], suggesting that Jagged-1 is closely related to the immunomodulatory function of MSCs. Recent studies have reported that Jagged-1 could be the target gene of the Notch pathway [61,62]. Similarly, the Notch pathway was observed to modulate Jagged-1 expression in MSCs in ITP. Hence, there may be a positive feedback function of the Notch signaling pathway in MSCs to mediate regDC differentiation, which needs to be studied further.
ATRA, the biologically active, key metabolite of vitamin A, has been shown to be central to tolerance and immunity [63]. In particular, ATRA has been reported to play an important role in the inhibition of Th1 and Th17 development, as well as in the promotion of Foxp3+ Tregs and tolerogenic DC development [64 –66]. ATRA has also been used for the treatment of acute promyelocytic leukemia [67] and in animal models of autoimmune diseases such as systemic lupus erythematosus [68], rheumatoid arthritis [69], and type 1 diabetes [70]. Recently, Dai et al. reported the therapeutic efficacy of ATRA in patients with ITP via the induction of Tregs [71]. In this study, we demonstrated a novel mechanism of ATRA treatment in ITP through the correction of MSC deficiency. It was found that the pretreatment of ITP-MSCs with ATRA significantly increased their ability in inducing regDCs via the Notch-1/Jagged-1 signaling pathway, further supporting the potential therapeutic application of ATRA for the treatment of ITP. Additional investigations will be needed to fully understand the effects of ATRA therapy on the immune system in ITP.
Taken together, the deficiency of MSCs in inducing regDC differentiation may be involved in the pathogenesis of ITP. The downregulation of Notch-1/Jagged-1 signaling pathway may be associated with the impaired function of MSCs in ITP. Our study provides novel information on immune therapeutic avenues targeting MSCs in the management of ITP and suggests that ATRA could serve as a novel potential therapeutic agent for ITP.
Footnotes
Acknowledgments
This work was supported by National Natural Science Foundation of China (no. 81470343 and no. 81670116), Key Program of National Natural Science Foundation of China (no. 81730004), Beijing Natural Science Foundation (no. 7171013), Beijing Municipal Science and Technology Commission (no. Z171100001017084) and the National Key Research and Development Program of China (no. 2017YFA0105503). This work was partly supported by grants from the National Natural Science Foundation of China (no. 81300439) and the Specialized Research Fund for the Doctoral Program of Higher Education (no. 20130001120078).
Author Disclosure Statement
No competing financial interests exist.