
Abstract
Liver transplantation is the only effective treatment for end-stage liver disease, but absolute donor shortage remains a limiting factor. Recent advances in tissue engineering focus on generation of native extracellular matrix (ECM) by decellularized complete livers in animal models. Although proof of concept has been reported for human livers, this study aims to perform whole liver decellularization in a clinically relevant series using controlled machine perfusion. In this study, we describe a mild nondestructive decellularization protocol, effective in 11 discarded human whole liver grafts to generate constructs that reliably maintain hepatic architecture and ECM components using machine perfusion, while completely removing cellular DNA and RNA. The decellularization process preserved the ultrastructural ECM components confirmed by histology, electron microscopy, and proteomic analysis. Anatomical characteristics of the native microvascular network and biliary drainage of the liver were confirmed by contrast computed tomography scanning. Decellularized vascular matrix remained suitable for normal suturing and no major histocompatibility complex molecules were detected, suggesting absence of allo-reactivity when used for transplantation. After extensive washing, decellularized scaffolds were nontoxic for cells after reseeding human mesenchymal stromal or umbilical vein endothelial endothelium cells. Indeed, evidence of effective recellularization of the vascular lining was obtained. In conclusion, we established an effective method to generate clinically applicable liver scaffolds from human discarded whole liver grafts and show proof of concept that reseeding of normal human cells in the scaffold is feasible. This supports new opportunities for bioengineering of transplantable grafts in the future.
Introduction
T
Although these are powerful tools worth exploring, it remains difficult to design and create artificial, yet functional liver tissue with functional vascular and biliary trees for clinical use. Alternatively, removal of cells from an existing organ, leaving a complex mixture of structural and functional proteins that constitute the extracellular matrix (ECM), may provide a natural habitat for reseeding with an appropriate population of cells [5 –7], and connected to the blood stream and biliary system.
The role of the ECM in guiding liver repair and regeneration was recently established [8], underlining the importance of naturally structured biomatrices, instead of using uncontrolled artificial compounds.
Ideally, ECM is cell free, but remains the interlocking mesh of fibrous proteins (collagen, elastin, fibronectin, and laminin) and glycosaminoglycans (GAGs) [9,10]. Evidence from rodent models shows the feasibility of decellularization of whole liver organs that provides an excellent scaffold for reseeding liver (stem) cells for graft engineering [11 –13]. Also, porcine [14,15] and sheep liver [16] have been successfully decellularized to obtain ECM for transplantation. However, so far, there is very limited experience with decellularization of whole livers from humans.
Recently, Mazza et al., showed efficient decellularization of a whole liver (n = 1) and partial livers (n = 2) to generate small cubes of human liver scaffold [5]. Different decellularization methods have been described among which are physical force (freeze/thaw, sonication, and mechanical agitation), enzymatic agents (trypsin, endonucleases, and exonucleases), and/or chemical agents (ionic, nonionic, and zwitterionic detergents) [6]. Usually, combinations of these methods are used. In larger organs, such as human or porcine liver, perfusion through the intrinsic vascular beds is the favorable route to be able to reach all cells [16,17]. So far, most experimental decellularization protocols include the use of sodium dodecyl sulfate (SDS) to generate full freedom of cells and translucency, but this also progressively destroys the ECM and hampers clinical translation [16].
In this study, we report a universal effective protocol for establishing decellularized human whole liver scaffolds (DHLS) using machine perfusion and show proof of concept of recellularization with human umbilical vein endothelial cells (HUVEC) in vitro.
Experimental Procedures
Donor characteristics and liver preparation
Adult human livers (n = 11, age 52–71 years), declined for transplantation by transplant centers across the EuroTransplant area, were included in this study. Ten livers were from donation after circulatory death and one was from donation after brain death. All were discarded because of age beyond Dutch protocol criteria, severe steatosis, extended cold ischemia period, or too long first warm ischemia time. The study was approved by the medical ethics committee of the Erasmus MC University Hospital. Informed consent for use of declined livers for research purposes by the next of kin of the donors was documented in the donor data application of EuroTransplant.
All liver procurement procedures were performed by regional organ procurement teams using a standard technique of in situ cooling and flushed with 6–10 L of cold preservation fluid [Belzer University of Wisconsin (UW) solution; Bridge to Life Ltd., London, United Kingdom], containing 300 IU/kg of heparin (Leo Pharma, Ballerup, Denmark). The livers were stored and transported on melting ice. Upon arrival, the portal vein and the hepatic artery were cannulated and the liver was additionally flushed with 1 L of cold heparinized saline (0.9% NaCl containing 5,000 IU of heparin), to remove remaining blood and thrombi. The cystic duct was ligated to prevent bile to flow into the common bile duct.
Liver decellularization
Whole human livers were decellularized by dual machine perfusion through the portal vein and hepatic artery (Fig. 1A, B). For this, a perfusion setup was custom-made to enable decellularization through the portal vein and the hepatic artery using controlled flow rates of perfusion fluids consisting of 4% Triton X-100 (Brunschwig Chemie, Amsterdam, Netherlands) with 1% ammonium hydroxide (Sigma, St Louis, MO). The initial flow rate of 60 mL/min was increased to 350 mL/min at day 3 and onward, and was refreshed every 4 h, until the liver became pale and semitransparent (duration from 4 to 6 days). Subsequently, the remainder of cellular debris was washed out by perfusion with 0.9% NaCl (1 L, 30 mL/min). DNAse-I solution (1 L, 2 mg/L; Roche, Mannheim, Germany) was recirculated for 8 h at 37°C to remove all DNA fragments sticking to the matrix, followed by 0.9% NaCl (1 L, 30 mL/min). Biopsies were taken from left and right core and periphery of the liver before and after the procedure. The DHLS were stored at −20°C until further use.
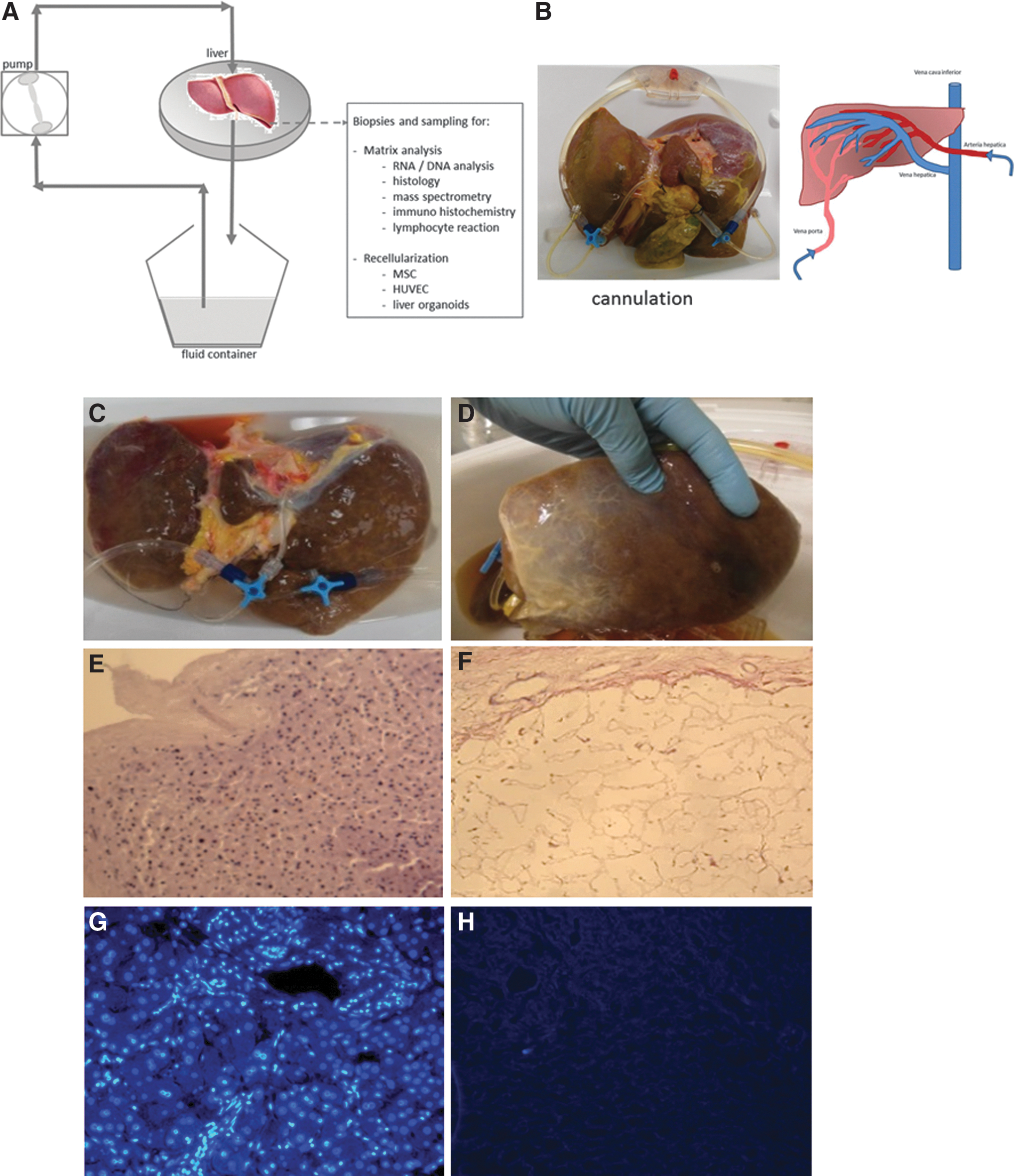
Perfusion-driven decellularization of whole human livers. Schematic representation of the machine perfusion set up
Histological analysis
Biopsies were formalin fixed (4%) for 24 h, paraffin embedded, and sectioned (5 μm) for histological examination. To analyze the general morphology and the efficiency of decellularization, sections were stained with hematoxylin and eosin (H&E) and analyzed using a bright field microscope. Additional sections were processed and stained with Picrosirius Red (collagen type I and III) and Weigert's resorcin fuchsin (collagen and elastin) [18 –20] to analyze the most abundant matrix proteins. Sudan Black B staining was performed on paraffin-embedded sections according to standard procedure, to demonstrate the presence of the aging-related pigment lipofuscin.
DNA-RNA detection
Biopsies from DHLS for RNA and DNA isolation were weighted and kept in RNA-later (500 μL; Qiagen, Venlo, Netherlands) and stored at −80°C until further analysis. After thawing, the biopsies were homogenized in 700 μL Qiazol (Qiagen) using the TissueRuptor (Qiagen). The RNA fraction was extracted using the Macherey-Nagel NucleoSpin kit. The DNA fraction (interphase after chloroform extraction) was isolated using a standard back extraction buffer protocol. DNA and RNA content were measured using standard protocol on a NanoDrop spectrophotometer (Thermo Scientific, Breda Netherlands). In addition, paraffin sections were stained with 4′,6-diamidino-2-phenylindool (DAPI) and DNA content was analyzed by fluorescence microcopy.
Scanning electron microscopy
Representative biopsies of two decellularized human livers were analyzed in a field emission electron microscope (JSM-7500F; Jeol, Peabody, MA). The liver pieces were frozen in liquid nitrogen and broken to mount the fold zone on the specimen holder. The fold zone was covered with 15 nm Au nanoparticles during 50 s in a dual target sputter coating machine (Quorum Q300TD; QuorumTech, East Sussex, United Kingdom) to make it electrically conductive.
Protein analysis
GAG assay
GAGs are measured as described previously [21]. In short, direct-frozen DHLS biopsies were mechanically fractionated using sonification, and subsequently digested using a papain solution (10 mg/mL in H2O, at 65°C, pH 6.5; Sigma). Subsequently, the sample was mixed with phosphate-buffered saline (PBS)-EDTA, substrate was added [1,9 dimethylmethylene-blue [DMB]; PolySciences, Eppelheim, Germany], and the extinction was measured in a spectrophotometer at 590/530 nm wavelength. The GAG content was calculated using a standard curve of chondroitin sulfate C (0.5 mg/mL; Sigma) in PBS.
High-resolution liquid chromatography–mass spectrometry
Protein samples were isolated from frozen biopsies taken from human decellularized livers (n = 3) and normal liver (n = 1) and processed for high-resolution liquid chromatography–mass spectrometry (Orbitrap LC-MS) analysis. In short, 6-μm-thick frozen sections were collected (in triplicate) using a cryostat microtome (Leica, Eindhoven, Netherlands). The use of optimum cutting temperature formulation was avoided to prevent contamination of the samples by this polymer. Frozen tissue sections were denaturized using 100 μL 0.1% RapiGest™ SF surfactant (Waters, Inc., United Kingdom) in 50 mM ammonium bicarbonate, followed by sonification for 2 min (amplitude 70%, 37°C). Protein sulfur bridges were reduced using dithiothreitol (5 mM, 30 min) in an Eppendorf thermomixer (56°C, 450 rpm). Alkylation was performed by addition of iodoacetamide (15 mM) and incubated in a thermomixer (30 min at 37°C, 450 rpm). Proteins were digested using trypsin gold mass spectrometry standard (Promega, Madison, WI) to a ratio of 1:100–1:20 w/w. Samples were incubated overnight (37°C, shaking, pH 8). Digestion was stopped using trifluoroacetic acid (0.5%, pH <2, 37°C, 45 min). The samples were separated by a nano-LC system followed by mass spectrometry [22]. Samples were analyzed by an Orbitrap Fusion™ Tribrid™ mass spectrometer (Thermo Fischer Scientific, San Jose, CA). Tandem mass spectrometry (MS/MS) spectra were extracted from raw data files and converted into mascot generic format files using extract msn (Xcalibur version 2.0.7.; Thermo Fischer Scientific). MS/MS spectra were sequenced through a Mascot MS/MS database search (version 2.3.2.; Matrix Science, Inc., London, United Kingdom) against the UniProt Swiss-Prot database (version 2014–09), taxonomy Homo Sapiens (20, 196 sequences). The search parameters used a maximum of four missed cleavages, tryptic cleavage, methionine oxidation (15.995 Da) as a variable, also proline and lysine hydroxylation (15.995 Da) were variable. Peptide mass tolerance was set at 10 ppm with a fragment mass tolerance of 0.5 Da. For peptide identification, a minimum ion score of 25 was required. These results were imported and further analyzed by Scaffold (version, 4.3.4; Proteome Software, Inc., Portland, OR), GenePattern (v3.9.1, January 2015) [23], and String (v9.1) [24].
Vascular integrity
To evaluate the integrity of the vascular network, Türk's solution (Merck) was injected into the portal vein and hepatic artery of a decellularized liver. In another scaffold, 50 mL iodine-containing positive X-ray contrast fluid (Iomeron® 300, 1:5 diluted; Bracco Imaging Deutschland GmbH) was injected into the portal vein and imaged immediately using a micro-computed tomography (CT) scan (SkyScan 1076 micro-CT; Bruker, Belgium). Scanning was done during 2.5 min. To demonstrate the integrity of the biliary tree, contrast fluid (diluted 1:20) was injected into the common bile duct and a CT image was made immediately using a clinical Somatom Force CT scanner (Siemens Healthcare).
Reendothelialization with HUVEC
HUVEC were kindly provided by Dr. Uitte de Willige (Erasmus MC) and expanded on gelatin-coated (1%) culture flasks (T25cm2; Greiner) in medium 199 (M190; Invitrogen), supplemented with endothelial cell growth factor (100 × ; Sigma), 1% pen/strep, and 20% fetal bovine serum. Cells are passaged 1:3 when 90% confluent. No passage numbers higher than passage 7 were used. HUVEC were incubated for 1–5 days with sections cut from DHLS (ø 8 mm and 250 μm thick) in a 48-well plate (Greiner Cellstar multiwell culture plate) in the HUVEC culture medium. Each section received 104 HUVEC. Every day (days 1, 2, 3, 4, and 5), sections were harvested, fixed in 4% paraformaldehyde, and embedded in paraffin. Immunohistochemistry was performed to detect endothelial cells using the endothelial markers: vimentin (Clone 9; Ventana) and Factor 8/von Willebrand Factor (dilution 1:8000, polyclonal; Dako). The sections were pretreated with cell conditioning solution-1 (CC1; Ventana Medical Systems, Inc., Tucson), 32 min at 97°C.
Results
Effective whole liver decellularization of human liver grafts
During the decellularization process, the human livers gradually lost their red-brown color to become pale beige (Fig. 1C, D). Interestingly, the characteristic transparency as seen in rodent livers was not obtained, although all human livers that were processed in this study were homogeneously perfused and fully decellularized. Overall, the 3D structure remained intact and did not collapse, deflate, or tear. Histological analysis of H&E-stained sections revealed the total absence of cells (Fig. 1E, F). A residual mesh of matrix structures remained present after the treatment. In accordance with this, the RNA content in the DHLS was decreased by up to 95% after decellularization (Fig. 2A). On average, 454 ± 401.6 ng RNA/mg wet weight [average ± standard deviation (SD)] was isolated in normal liver biopsies, whereas in the DHLS, 24.6 ± 7.0 ng/mg wet weight (average ± SD) remained. Also, DNA content of the liver was dramatically reduced to 2% [from 291.4 ± 231.8 ng/mg wet liver weight (average ± SD) before decellularization to 5.8 ± 3.8 ng/mg wet liver weight (average ± SD) in the DHLS] after the decellularization process followed by DNAse digestive treatment (Fig. 2A). DAPI-stained sections, made from biopsies taken after the decellularization process, were analyzed by fluorescence microscopy. No nuclei were found, confirming the removal of DNA and RNA in the DHLS (Fig. 1G, H). The pale beige color of the DHLS was most probably due to remaining lipofuscin, which was detected in predecellularized and postdecellularized human livers using Sudan Black B staining (Fig. 3).
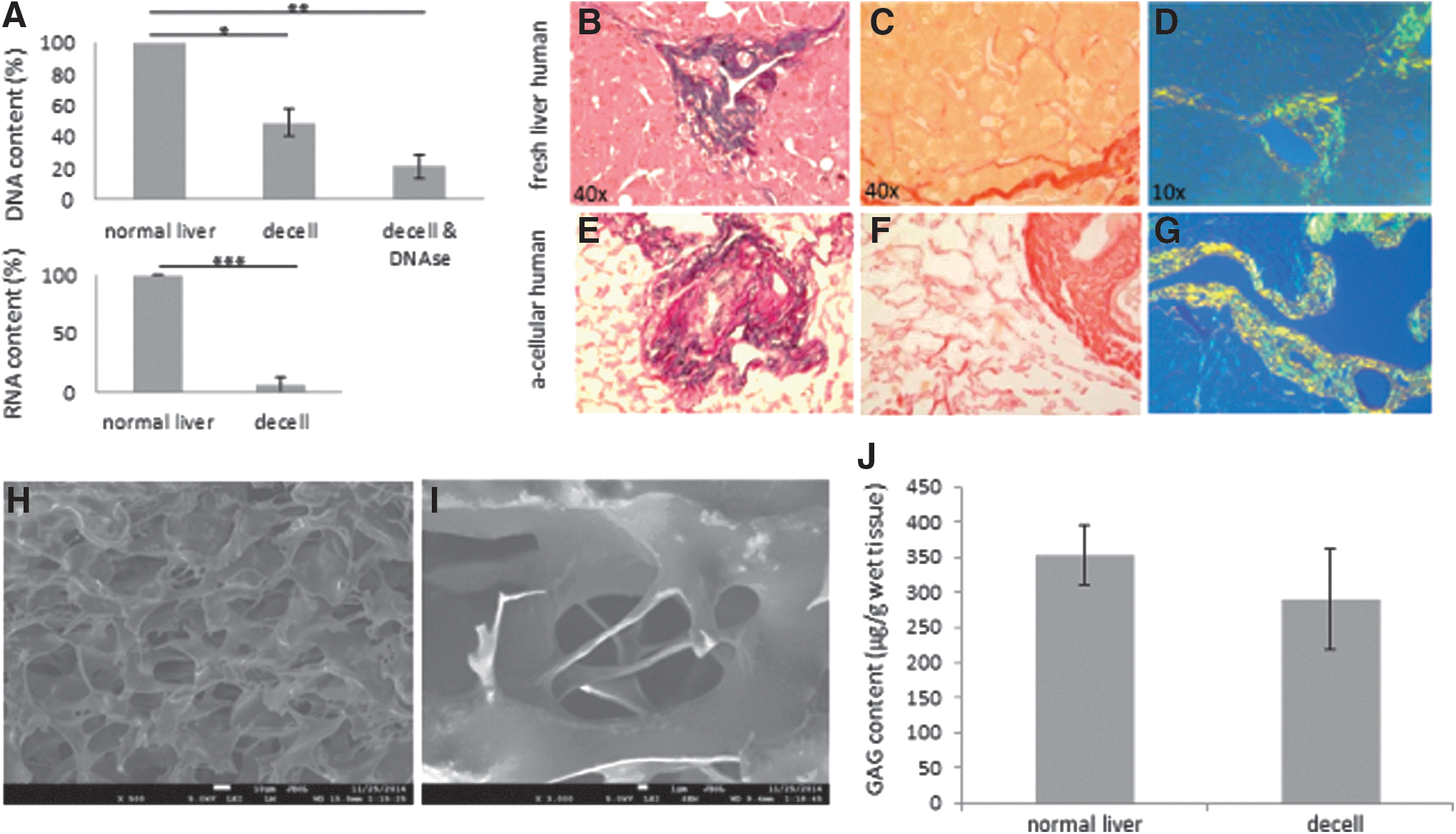
Decellularized livers preserve matrix proteins and their natural localization. DNA content decreased significantly (*P < 0.005) when treated with Triton X-100 and further decreased after washing with a DNase solution to 2% of DNA (**P < 0.0001, n = 4). RNA was washed as efficiently to 5% left after the decellularization process (***P < 0.002, n = 4). GAG content did not differ significantly (P = 0.18, n = 9) from normal liver contents after decellularization, and were not damaged or removed by the decellularization method used. DNA, RNA, and GAG content are expressed as microgram per gram wet liver weight
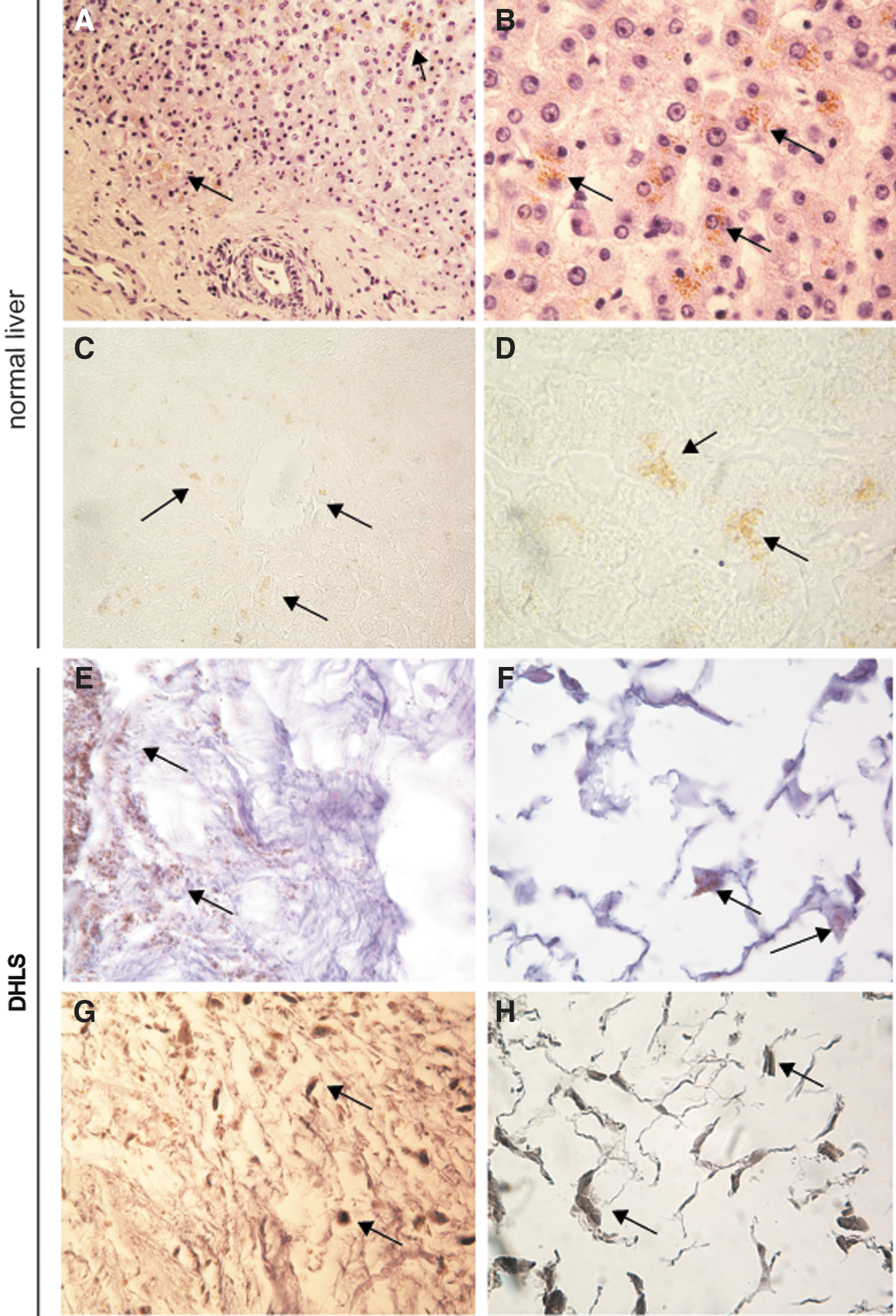
Lipofuscin-related pigment in the decellularized livers. Lipofuscin was detected in livers before decellularization as brown pigment in H&E-stained slides
Decellularized liver matrix integrity and GAG content
Left and right peripheral and core liver biopsies, taken before and after decellularization, were processed for histological analysis of matrix proteins collagen type I and type III and elastin by Weigert's resorcin fuchsin staining and Picro Sirius Red staining. Figure 2B–G show representative (DHLS) images. The most abundant types of collagen in the liver, collagen type I and III, including elastin, remained present at their natural location and were not affected by the decellularization process. To investigate the scaffold and its molecular surface in detail, field emission scanning EM was performed (Fig. 2H, I). The parenchymal space of the scaffold showed an open parenchymal structure with no cells left. Sulfated GAG content was measured using a direct spectrophotometric microassay. As shown in Fig. 2J, comparing the GAG content before to samples taken after the process, no significant differences were found (P = 0.18).
Protein content of decellularized human liver scaffolds
Proteomic analysis of the DHLS was compared to normal liver (t = 0), using LC-MS. As shown in Table 1, proteins most abundantly present were collagen type III and I, integrin, filamin, fibronectin, and other ECM-related proteins, as expected. Furthermore, some hepatocyte-abundant and/or liver-specific protein fragments (not matrix related) were found associated with the ECM. These included proteins related to fatty acid metabolism, complement and a few mitochondrial proteins (ATP synthase related). Although human leukocyte antigen (HLA)-related proteins were detected in the normal, nontreated liver samples, none of these immunogenic proteins were detected in the decellularized samples. Hierarchical clustering (GenePattern v3.9.1 [23] is graphically represented in a dendrogram; Fig. 4A) clearly illustrates the clustering of the samples taken before decellularization and the samples analyzed for three different decellularized livers. The analysis is done on the total set of proteins detected (1,761 proteins); however, only the top 25 most abundantly present proteins are shown in the heat map for clarity purposes. The interaction of the top 25 proteins expressed in decellularized liver was visualized using String analysis (v9.1) [24] (Fig. 4B), and clusters the matrix proteins, mostly collagens. A few nonmatrix related, but liver-specific protein fragments are present and clustered. Table 1 summarizes the top 25 most abundantly present proteins in the decellularized livers. Altogether these data show that matrix integrity and composition is well preserved during the decellularization process.
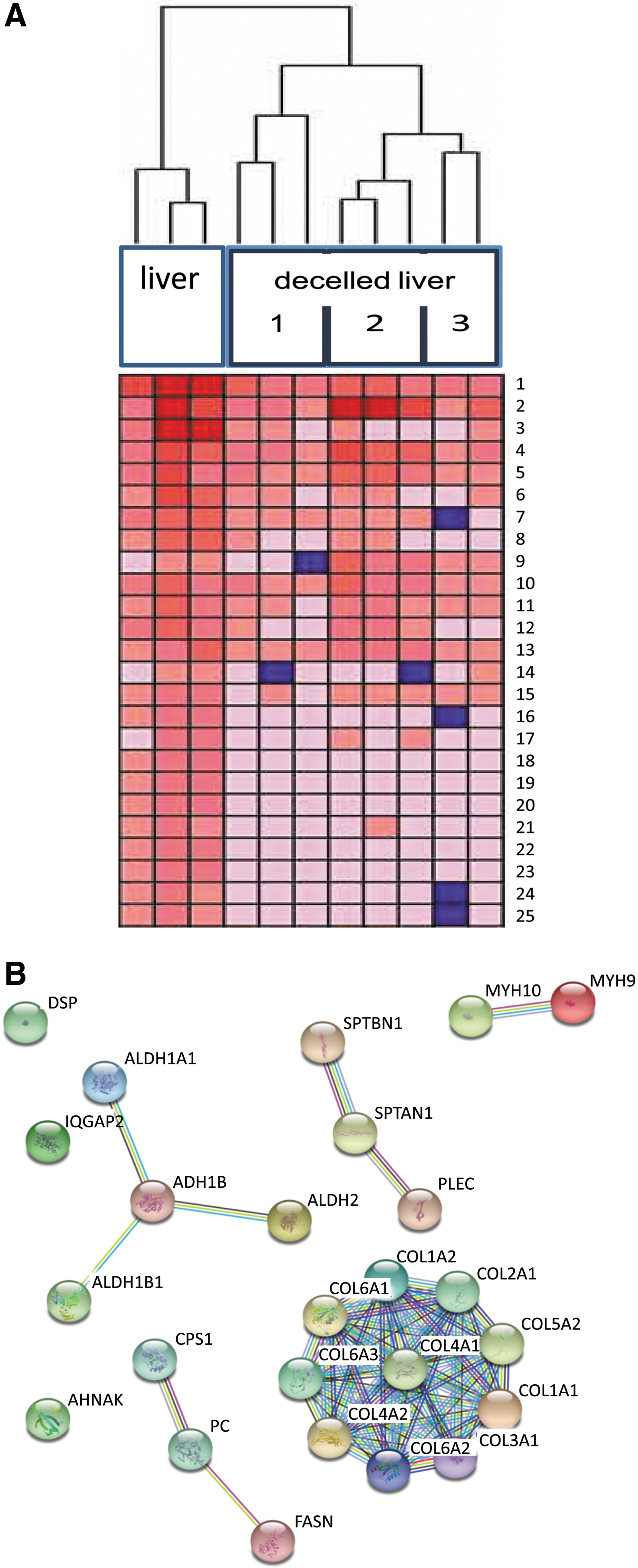
Differential expression of proteins present in the decellularized human whole liver scaffolds. Mass spectrometry analysis was performed on cryosections (6 μm) that were cut from a nontreated liver and three decellularized livers. Three sections (two in the case of liver #3) were collected from different areas in each liver and analyzed. Hierarchical cluster analysis clearly demonstrated the different protein expression between the samples taken before decellularization versus after (liver #1, #2, and #3). The three sections that were taken from each liver clustered together, but were different between the three decellularized livers demonstrating the differences between the livers after the treatment. The differential expression of the proteins (only the top 25 proteins are shown here) are visualized in the dendrogram
Top 25 most abundantly present proteins found in decellularized human livers as determined with liquid chromatography–mass spectrometry. Three decellularized livers were analyzed in triplo.
Decellularized human liver scaffolds are sterile and sustain cells in culture
To test the sterility of human decellularized liver, 200-μm-thick sections of DHLS (n = 15) were incubated in a culture medium at 37°C for over 2 months. The medium was refreshed twice weekly and did not contain any antibiotics. No bacterial, fungal, or yeast growth was found (data not shown), which shows that the decellularized human livers can be used in sterile cultures for recellularization. In addition, all proteins detected by mass spectrometry were matched to the whole bacterial database (Uniprot Swissprot) and no bacterial proteins were detected (data not shown).
Biliary and vascular network is preserved in human liver scaffolds
CT analysis of the biliary tree (Fig. 5) visualized the complete biliary tract providing evidence that this system is leak free, although denuded of biliary epithelial cells. Because of absence of tissue counter pressure, the bile ducts appear more bulgy than on normal cholangiography. LC-MS data also confirmed the presence of basement heparin glycoproteins in the decellularized samples, indicating that the basal membrane remains intact after decellularization. As also shown in other studies [12,13,17], the vascular network stays intact after the decellularization procedure. The blood vessels are transparent (Fig. 6A) and have sufficient strength to be sutured in a transplantation setting (Fig. 6B). Although the endothelial cells covering the vascular system in the liver are removed as a result of the treatment, the vascular network structure preserved its integrity, as demonstrated by injecting blue dye into the portal vein and hepatic artery (Fig. 6C). The vascular branches were visualized throughout the liver, and no leakage of the dye outside the blood vessels occurred. In addition, an intact vascular system was found when injecting iodine-containing contrast into the portal vein followed by micro-CT analysis (Fig. 6D). The presence of basement membrane proteins, mostly heparin sulfate proteoglycans, after decellularization was confirmed by proteomics (data not shown).
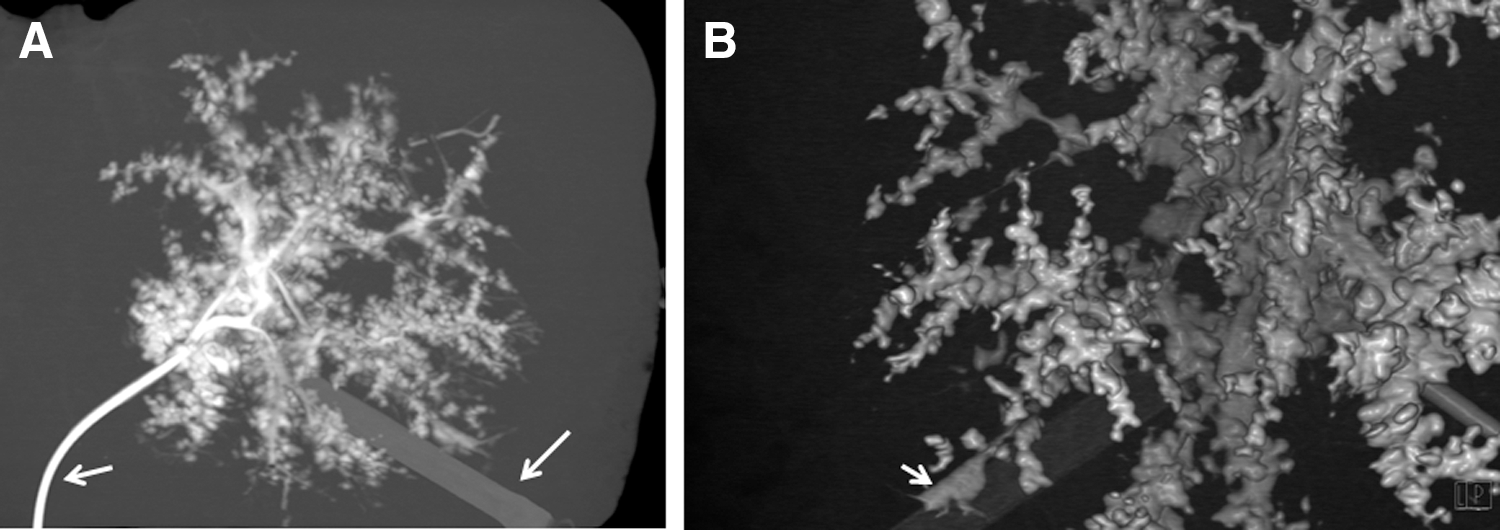
Biliary integrity. CT-scan in which the biliary tract is visualized using contrast fluid
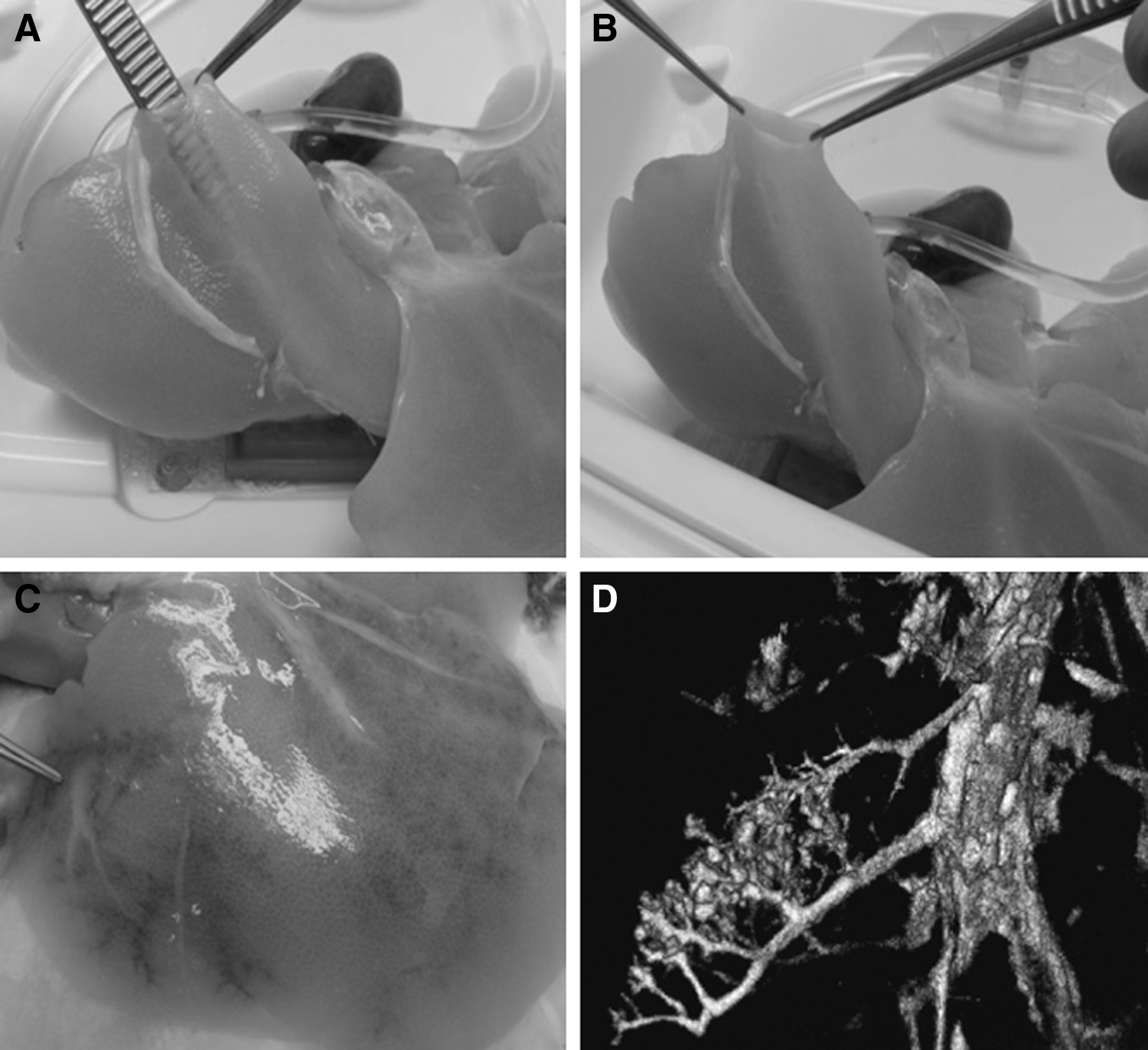
Vascular network integrity. The blood vessels of decellularized livers are transparent, but sturdy
Reendothelialization with HUVEC
Sections of human decellularized liver seeded with HUVEC were stained with antibodies against vimentin and von Willebrand Factor/F8, which are markers for endothelial cells. Vimentin and vWF/F8 staining were found in the lumina of blood vessels from day 1 onward to day 5 of incubation (Fig. 7, day 5 data are shown), indicating that the endothelial cells found their natural habitat and adhered there. As indicated, negative control sections (decellularized) did not stain (Fig. 7C, D) and positive control sections (normal liver) were all positive (Fig. 7A, B).
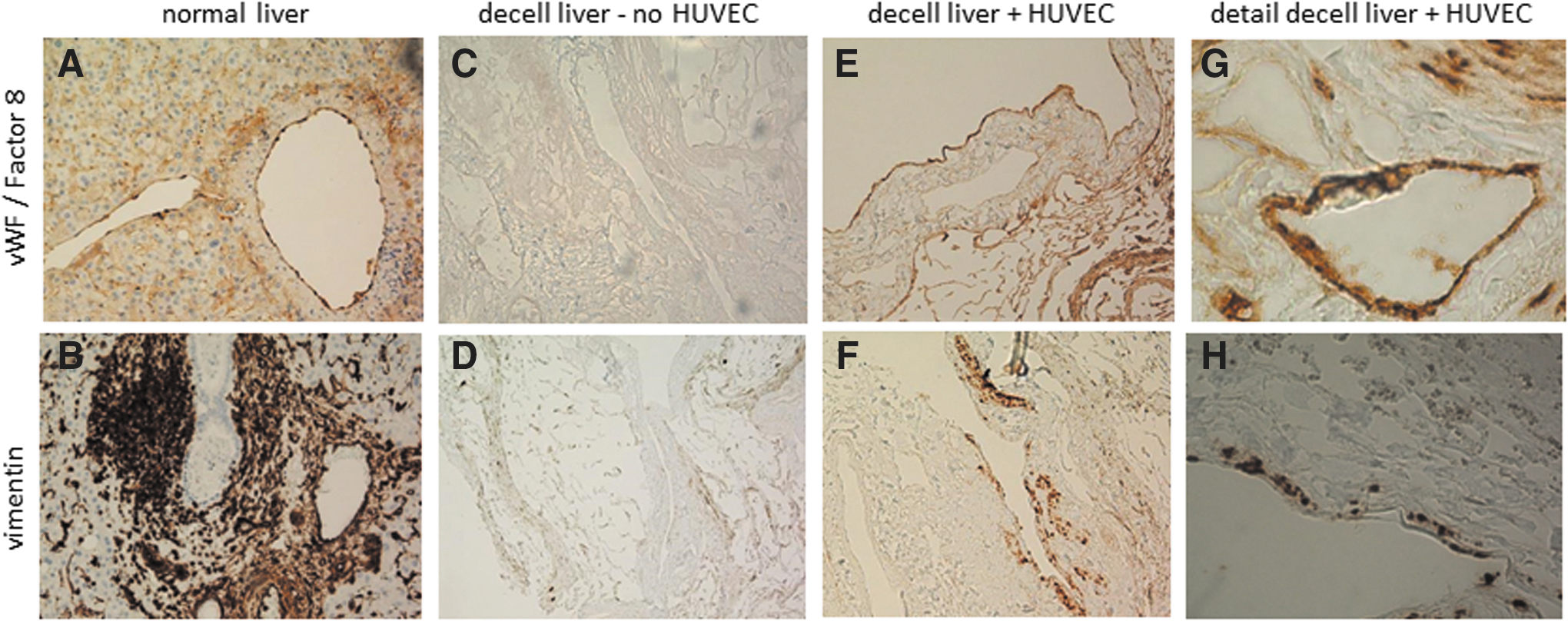
Reendothelialization of human decellularized matrix with HUVEC. Representative immunohistological staining (one representative of three experiments done) of decellularized liver sections incubated for 5 days with HUVEC. Control endothelial cells in normal liver stained positive for vWF/Factor8 and vimentin
Discussion
In this study, we report successful decellularization of human livers to obtain transplantable whole organ scaffolds. We show proof of concept that these scaffolds can serve as feasible resources for future tissue-engineering purposes. Using a controlled perfusion system, a complete 3D acellular human liver scaffold was generated on a clinically relevant scale and free of allo-antigens. The use of whole organ decellularization for the generation of vascularized liver scaffolds was pioneered by the Wake Forrest School of Medicine [12] and Harvard Medical School [7]. They, and others, demonstrated the feasibility to generate acellular liver matrices of different species origin in which components such as collagens, elastins, and GAGs are preserved [7,12,15,25 –32].
In this report, we present the feasibility of systematically upscaling the decellularization process to discarded human livers. Eleven human livers were efficiently decellularized by nonionic detergents by machine perfusion. A careful choice of the decellularization methodology is of great importance as methods described for decellularization may be well suitable for other organs than the liver, but may damage the composition of the matrix proteins [11,33 –35]. Interestingly, in our study, the decellularized human livers did not become as transparent as described in rodents. Even prolongation of the decellularization process using only the mild 4% Triton X-100 perfusion method did not result in a translucent liver. This is in line with previous studies, describing that SDS is typically more effective for removing cell residues from tissue compared to other detergents, but is also more disruptive to ECM [36 –38]. To prevent this, SDS was successfully left out of the perfusion solution with 100% removal of the cells without damage to the matrix proteins, and GAGs in particular.
Recently, Mazza et al. decellularized the left lobe of a human liver (segment 1, 2, 3, and 4) (n = 2) and one whole liver using a perfusion protocol with Triton X-100 in combination with 0.01%, 0.1%, and 1% SDS [5]. And, although they confirmed the presence of the liver architecture by the presence of collagen types I and IV, fibronectin, and other matrix proteins, there was no in-depth analysis of other important matrix components as GAGs that are usually severely damaged when using SDS in the decellularization procedure.
The remaining pale beige color of the livers described in our study was also noticed before in decellularization of kidneys from higher species [39]. It might possibly be due to accumulation of lipofuscin that accumulates with increasing age [40 –42], and which is obviously absent in the young laboratory animals used for decellularization. Lipofuscin appears as yellow-brown granules, alternatively called “dense bodies” that mainly contain oxidatively modified protein and lipid degradation residues, including triglycerides, free fatty acids, cholesterol, and phospholipids, but also carbohydrates and metals [43]. Lipofuscin granules can be detected in cell types that exhibit low rates of mitosis, or at least retain the potential for division, such as hepatocytes [40], and are not degraded or eliminated otherwise [44]. The accumulation of lipofuscin pigment in lysosomes is found in many tissues, including liver, and correlates with age [41]. The livers used in this study were all relatively old as opposed to the young laboratory animals used in other studies. Lipofuscin was detected in the livers before and after decellularization and is most probably responsible for the nontransparent appearance. Perfusion with SDS would most probably remove the lipofuscin, but as these “wear and tear” pigments are harmless and do not have any pathological implication, the milder, but effective decellularization method using Triton X-100 excels over the use of SDS that is known to damage the ECM [11,33 –35].
In this study, the human livers were cannulated through the portal vein and hepatic artery to enable perfusion of the whole organ. The choice for this route of entry and the use of Triton X-100 and ammonium hydroxide to wash out the cells were based on previous studies that compared several techniques for decellularization of rodent, sheep, and pig organs [15,16,45,46]. The intricate vascular network, including portal vein, hepatic artery inflow, and hepatic venous outflow, remains intact, even in the absence of vascular endothelium, which is in accordance with animal studies [12,13,17,47].
However, the integrity of the biliary tree was not thoroughly investigated before. Adequate biliary drainage is crucial and may form the “Achilles heel” of bioengineered livers. Only Yagi et al. showed that the larger bile ducts of the biliary tree seemed to have a natural structure [17]. However, the small peripheral bile ducts were affected, which might be explained by the use of more aggressive detergent (SDS) rather than using Triton X-100 [11,33]. In our study, the same integrity of the intrahepatic bile duct with adequate branching was shown. Histology and LC-MS proved conservation of the structure and composition of each specific ECM protein. In addition, LC-MS revealed the absence of major histocompatibility complex proteins in the scaffold, confirming the complete removal of cellular antigens and the suitability for use as allografts. Indeed, mixed lymphocyte reactions performed with rat matrix suggest that decellularized scaffolds have no allo-antigens that can induce lymphocyte proliferation or rejection [28].
Repopulation of a complex organ such as the liver poses numerous challenges. Using the extracellular matrix of the native liver obviously helps to create the most optimal niche for cells to repopulate, but the types of cells to be infused to create fully functional liver tissue remains to be elucidated. In addition to the liver-specific matrix proteins, the still present vascular and biliary system may also provide entry routes for the different cell types needed. Obviously, efficient recellularization is a complex process in which hepatocytes or other parenchymal cells need to pass the remnant basement membrane of the decellularized blood vessels or bile ducts to enter the parenchyma after vascular or biliary administration, respectively. In addition, cell numbers that are required for efficient recellularization are highly dependent on cell type and volume of the scaffold, as reviewed by Meng et al. [48].
Reendothelialization is a pivotal step to prevent thrombosis as a result of the massive collagen contact surface that blood will encounter upon reperfusion [47], and which cannot be prevented by coating with heparin [49]. We demonstrated, like others did in animal models [7,12], that matrix sections can be reseeded with endothelial cells and these cells end up at the location of the decellularized blood vessels and pave the basal membrane. In our studies, HUVEC were used as a source of endothelial cells, as in most studies in rodents and pigs [12,13,39,50], but other sources such as endothelial progenitor cells [51] are also used and show similar results.
The next hurdle to be taken toward clinical application is to choose a cell source for liver parenchyma repopulation. An adult liver contains ∼150–350 billion cells of which the largest part (70%–85%) is made up by hepatocytes. However, adult primary hepatocytes of high quality are scarce and therefore limit tissue-engineering applications. Ideally, autologous cells, isolated from the patients themselves, are used as these cells will have a low risk to trigger an immune response. However, cell numbers will be the limiting factor.
Alternatively, (autologous) pluripotent stem cells that self-renew and are able to differentiate into all cell types needed could be seeded. As embryonic stem cells are not available mainly due to ethical concerns, inducible pluripotent stem cells [52] are promising cell types to use in reseeding a liver scaffold [53]. Although these cells can be grown in large numbers and give rise to parenchymal and supportive cells, their use is not ideal due to their epigenetic modifications of DNA and the production of teratomas when transplanted in an undifferentiated state [54]. This may be overcome by using inducible human hepatocytes from pluripotent stem cells, generating large amounts of functional liver cells to reload the matrix [55]. However, the concern about the plasticity of these cells to form bile ducts is a main obstacle in clinically transplantable liver matrix engineering. In addition, to repair the endothelium of the vascular system, bile ducts also need to be repaved with biliary epithelium. A potential source of biliary epithelial cells may come from the recently discovered bipotential liver organoids, originating from Lgr5+ adult stem cells isolated from human liver, which are able to differentiate into both hepatocytes and cholangiocytes depending on culture medium composition [56]. Bipotential liver organoids could also be driven toward cholangiocyte differentiation and may be applied to repopulate the intrahepatic and extrahepatic bile ducts to gain functional bile ducts. These liver-derived organoids would be the most promising cell type to use, with or without infusion of additional accessory cells as mesenchymal stromal cells (MSC) [57]. The requirement of different cell types in various quantities might also be circumvented by using liver-derived organoids to repopulate the hepatocyte and cholangiocyte population. Instead of infusion of differentiated hepatocytes or cholangiocytes, organoids could be administered as undifferentiated stem cells that subsequently differentiate in situ according to the need and based on signals present in the decellularized scaffold [57]. In summary, human cadaveric livers can be successfully decellularized using machine perfusion and nonionic detergents, and can be repopulated with endothelial cells. The next steps toward clinical application involve finding a cell source or combinations of cell types to reseed the matrix, including the vascular and biliary system, to gain functional liver tissue.
Footnotes
Acknowledgments
The authors thank Christof Boers and Dorien Kimmenai, Department of Clinical Perfusion, Amphia Hospital Breda, for their assistance with the perfusion system. We are thankful for the help we received from Wendy Koevoet and Nicole Kops of the Department of Orthopedics and Otorhinolaryngology with GAG analysis and collagen histology. Joost Haeck and Yanto Ridwan of the Centre for Applied Molecular Imaging Erasmus MC (AMIE) are acknowledged for their help with visualizing the vascular system by CT analysis. We acknowledge Frans Oostrum, Faculty of Aerospace Engineering, TU Delft, for scanning electron microscopic analysis of the scaffold.
Author Disclosure Statement
No competing financial interests exist.