
Abstract
Chromodomain helicase DNA-binding protein 1-like gene (CHD1L) was initially isolated as a candidate oncogene in hepatocellular carcinoma, and it has been associated with many malignancies. Knockdown of Chd1l in zygote-stage mouse embryos resulted in developmental arrest, suggesting that Chd1l is required for mouse early development. However, the exact role of CHD1L in development, especially in humans, has not been reported. In this study, we found that overexpression of CHD1L in human embryonic cells (hESCs) upregulated the expression of ectoderm genes, especially PAX6. Furthermore, ectopic expression of CHD1L promoted hESCs to differentiate into neuroepithelium both in embryoid bodies and in directed neuronal differentiation. Knockdown of CHD1L significantly impaired neuroepithelial differentiation of hESCs. Interestingly, Chd1l colocalized with a PAX6-positive cell population and was highly expressed in the ventricular (germinal) zone of fetal mice. Taken together, these data suggest that CHD1L promotes neuronal differentiation of hESCs and may play an important role in nervous system development.
Introduction
I
Chromodomain helicase DNA-binding protein 1-like gene (CHD1L, also called ALC1) was first isolated as a candidate oncogene in hepatocellular carcinoma [8]. CHD1L is also an ATP-dependent chromatin-remodeling factor, and it contains a conserved SNF2-N domain, superfamily domain and macro domain. CHD1L has been shown to be associated with tumorigenesis, including hepatocellular carcinoma [8,9], ovarian carcinoma [10], bladder cancer [11], colorectal carcinoma [12], and gastric cancer [13]. Inhibition of CHD1L protein production with morpholino antisense oligomers in zygote-stage mice embryos blocks development before the blastocyst stage, which suggests that CHD1L is required for early development in mice [14]. However, the exact role of CHD1L in early development, especially in humans, has not been investigated.
In this study, we found that CHD1L promoted the differentiation of human embryonic cells (hESCs) into neuroepithelium. CHD1L overexpression in hESCs promoted neuroepithelial differentiation in both self-renewal and directional differentiation conditions. Knockdown of CHD1L impaired hESC differentiation into neuroepithelium. Overexpression of CHD1L dramatically upregulated PAX6 expression (a key regulatory gene in eye and brain development). Interestingly, Chd1l is highly expressed in cells of the ventricular (germinal) zone of E14 mouse embryos, and it colocalized with a PAX6-positive cell population. Our study suggests that CHD1L promotes the differentiation of hESCs into neuroepithelial cells.
Materials and Methods
Cultivation, spontaneous differentiation, and neural differentiation of hESCs
The hESC lines were established and cultured in our laboratory [15]. Briefly, hESCs were derived and cultured on a feeder layer of mitotically inactivated mouse embryonic stem fibroblasts at a density of ∼2,500 cells/cm2. The hESCs medium consists of DMEM/F12 supplemented with 15% knockout serum replacement, 4 ng/mL of basic fibroblast growth factor (bFGF), 2 mM nonessential amino acids, 2 mM
Establishment of CHD1L knockdown and overexpression of hESCs
For establishment of knockdown hESCs, two shRNAs (Supplementary Table S1; Supplementary Data are available online at
For establishment of overexpression of hESCs, full-length CHD1L cDNA was amplified and cloned into the expression vector pINDUCER20. The protocol was the same as the knockdown group with the only difference that the transfected cells were selected by G418 (11811023; Gibco) for 2 weeks.
Real-time PCR and quantitative PCR
Total RNA was isolated with TRIzol (15596018; Invitrogen) and cDNA was synthesized using 1 μg of total RNA, 20 μL reaction by the Transcriptor First Strand cDNA Synthesis Kit (04897030001; Roche) according to the manufacturer's instructions. Real-time PCR was performed in triplicate, using 6 μL cDNA (1:20 dilution) with LightCycler® 480 SYBR Green I Master (04887352001; Roche) and run on a 7500 Real-Time PCR Sequence Detection System (Applied Biosystems). The product sizes and primer sequences are provided in Supplementary Table S2. Relative RNA expression was given as 2−ΔCT, where ΔCt = Ct (target gene-28S). In some instances, the ΔCt values were further converted to relative expression levels normalized against the mean expression in the initial control. All data were presented as mean ± SD.
Western blotting analysis
Western blotting analysis was performed with the standard method with antibodies to rabbit anti-CHD1L (ab197019, 1:1,000; Abcam), mouse anti-β-Actin (MA5-15739-HRP, 1:1,000; Sigma), mouse anti-POU5F1 (sc-5279, 1:1,000; Santa Cruz), rabbit anti-SOX2 (ab92494, 1:1,000; Abcam), rabbit anti-NANOG (ab109250, 1:1,000; Abcam), and rabbit anti-KLF4 (11880-1-AP, 1:1,000; Proteintech). The densitometry data were analyzed with Scion Image Software (version beta 3b).
Immunocytochemistry staining
Cells were fixed in 4% paraformaldehyde at room temperature for 15 min, then blocked for 1 h with KB buffer (0.01 M Tris, pH 7.5; 0.15 M NaCl; 0.1% BSA) followed by incubation with primary antibodies diluted in KB buffer overnight at 4°C. For intracellular antigen retrieval, cells were permeabilized for 10 min in KB permeabilizing buffer (0.01 M Tris, pH 7.5; 0.15 M NaCl; 0.1% BSA; 0.2% Triton X-100) before blocking. The following antibodies were used, mouse anti-PAX6 (1:200; DSHB), rabbit anti-SOX1 (ab109290, 1:100; Abcam), rabbit anti-SOX2 (ab92494, 1:100; Abcam), mouse anti-NESTIN (sc-23927, 1:100; Santa Cruz), mouse anti-TUJ1 (480011, 1:400; Millipore), rabbit anti-MAP-2 (sc-20172, 1:200; Santa Cruz), and rabbit anti-CHD1L (ab197019, 1:500; Abcam). Unbound antibody was removed by extensive washing. Localization of antigens was visualized by using Alexa Flour 488 or 594 secondary antibodies (1:1,000; Invitrogen). Nuclei were counterstained with DAPI (D9564, 1:1,000; Sigma).
Karyotype analysis
Standard G-banding karyotypic analysis was performed after the hESC cells had been treated with Demecolcine solution (D7385; Sigma) for 4 h. At least 20 metaphase spreads were examined for each sample using an Olympus epifluorescence microscope BX51 (Olympus, Japan) with LUCIA KARYOTYPE software (Lucia, The Netherlands).
Cell proliferation assay
chESC-137 cells were detached by 0.02% EDTA and counted using a hemocytometer (Hausser Scientific). Then single cell was seeded at 8 × 104/well on Matrigel (354234; Corning)-coated 24-well plate. The cells were cultured with mTeSR 1 and changed every day, and the cells were counted at 1 day intervals using a hemocytometer.
Colony formation experiment
The 137-CHD1L shRNA were cultured in ES medium for 6 days then were digested into single cell. Following counting, the single cell (20,000 cells/dish) was seeded into 60-mm dish containing feeder layer. After 8 days, the colonies were fixed and stained with Alkaline Phosphatase (AKP). The positive AKP population was considered as a single colony.
Hematoxylin–eosin staining of EBs
EBs were collected and fixed in 4% paraformaldehyde. HistoGel (HG-4000-012; Thermo) was heated into a liquid state at 60°C upon use. After centrifugation and removing the supernatant fixatives, the fixed EBs were mixed with prewarmed liquid HistoGel and adjusted to relatively condensed state. The mixture was cooled down to get solid state at room temperature. Following the solidification, the gel within EBs was routinely processed to check the histology. Briefly, the solidified mixture was dehydrated, paraffin embedded, sectioned, and stained with Hematoxylin–Eosin. The sections were observed under microscope.
Animals
Female C57BL/6 mice (8–12 weeks old) were obtained from the Animal Resources Center of Guangzhou Medical University (SCXK-Yue-2012). Animal care and experimentation were performed under the guidelines of the Institutional Animal Use and Care Committee at the University. The female animals were mated with males of the same strain and the age of the embryos was determined depending on the appearance of vaginal plug and the noon of day plug appeared was considered as the 0.5-day postcoitus gestational stage (E0.5).
Collection of embryos or brains
The E14.5 embryos were isolated from the uteruses of pregnant mice in phosphate-buffered saline. The isolated embryos were decollated and the heads were transferred into 10% buffered formaldehyde for 24 h. Similarly, the newborn pups and adult mice were decapitated and their brains were removed from the skulls and used similarly. The fixed tissues were paraffin embedded. Sections of 5 μm thickness were cut with a microtome and subjected to further histological and immunohistochemical examination.
Immunohistochemistry
Sections were subjected to PAX6 (DHSB) and CHD1L immunohistochemical analysis. The adjacent sections were deparaffinized and rehydrated. The endogenous peroxidase activity was blocked with 3% H2O2 for 10 min. For the antigen retrieval, slides were immersed in 10 mM citrate buffer (pH 6.0) and boiled in a microwave oven for 15 min. Nonspecific binding was blocked by 5% normal goat serum for 10 min. The slides were incubated with a 1:100 dilution of monoclonal antibody against CHD1L (1:100 dilution; Abcam) and PAX6 (1:100 dilution; Abcam) at 4°C overnight. After washing in PBS, the slides were sequentially incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (1:100 dilution; Chemicon) for 1 h at room temperature. The 3, 5-diaminobenzidine (DAB) Substrate Kit (Dako) was used for color development followed by Mayer Hematoxylin counterstaining. Isotype-matched human IgG was used in each case as a negative control.
Statistical analyses
Unless otherwise cited, all data are expressed as the mean ± SD, and were analyzed using SPSS 19.0 software (SPSS, Chicago, IL). Intergroup differences were explored using one-way analysis of variance and the least significant difference post hoc tests. All experiments were representative of at least three independent experiments. The data were analyzed using GraphPad Prism, version 6.0 (GraphPad Software). P value <0.05 were considered statistically significant.
Results
CHD1L overexpression in human embryonic stem cells upregulates the expression of PAX6 under self-renewal conditions
To investigate the function of CHD1L in hESCs, we constructed a FLAG-tagged CHD1L conditional expression lentiviral vector with pINDUCER20 (Fig. 1A) and selected stably infected hESC colonies (137-CHD1L) by G418. The CHD1L mRNA level in 137-CHD1L was upregulated after overexpression for 1 day (Supplementary Fig. S1A). After treatment with doxycycline (DOX, NO. D9891; Sigma-Aldrich), expression levels of CHD1L mRNA and protein in 137-CHD1L were both upregulated (Fig. 1B, C). Immunocytochemistry staining with an anti-FLAG antibody revealed that ∼90% of G418-selected pool colonies were FLAG positive (Supplementary Fig. S1B). Karyotype analysis showed that 137-CHD1L cells have a normal karyotype (Supplementary Fig. S1C).
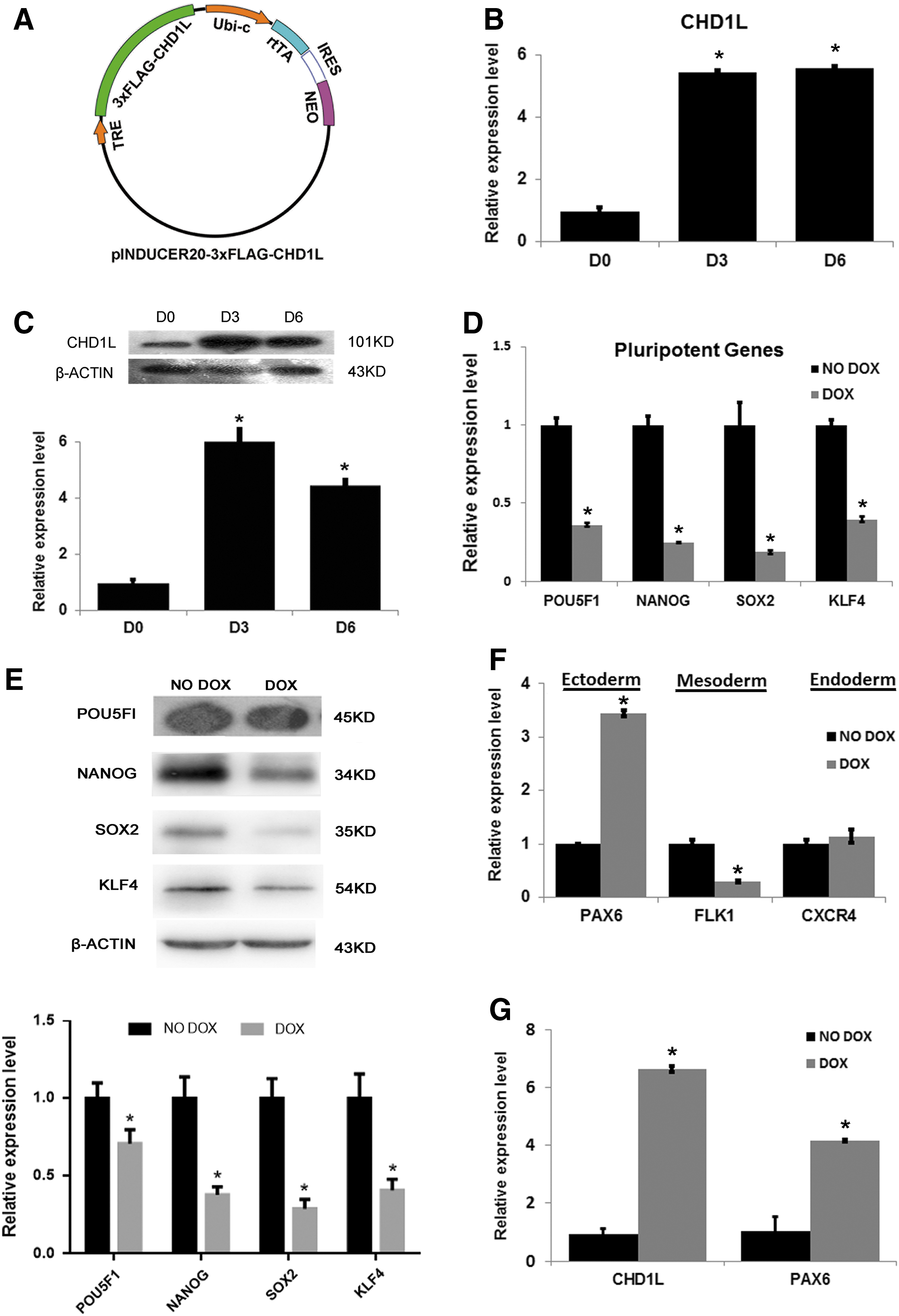
Ectoexpression of CHD1L in hESCs upregulated PAX6 expression under self-renewal conditions.
Although, the morphology of 137-CHD1L cells did not change after DOX treatment (Supplementary Fig. S1D), overexpression of CHD1L significantly inhibited hESCs growth (Supplementary Fig. S1E left). Consistent with the inhibition of growth, the colony formation ability of 137-CHD1L cells was significantly impaired after 5 days of DOX treatment (Supplementary Fig. S1E right). These results suggest that growth of hESCs was inhibited as a result of the ectoexpression of CHD1L, which may affect the self-renewal ability of hESCs.
To investigate whether CHD1L overexpression affects the pluripotency of hESCs, the expression level of pluripotency genes and three germ layer-related genes were checked by real-time PCR (RT-PCR). Upon overexpression of CHD1L, expression of pluripotency genes (POU5F1, NANOG, SOX2, and KLF4) decreased rapidly both in mRNA and protein levels (Fig. 1D, E). Among the three layer-related genes, the expression of endoderm marker (CXCR4) had no obvious change, mesoderm-related gene (FLK1) was markedly downregulated, and neuroectoderm marker (PAX6) was significantly upregulated (Fig. 1F and Supplementary Fig. S1F). Overexpression of CHD1L in another line of hESCs (8-CHD1L) showed the same results (Fig. 1G and Supplementary Fig. S1G). Taken together, these results indicate that overexpression of CHD1L induced ectodermal differentiation of hESCs and reduced their pluripotency, even in hESC culture conditions.
Overexpression of CHD1L promoted hESCs differentiation into neuroepithelium upon EB differentiation or neuroectodermal directional induction
Since PAX6 is necessary and sufficient for neuroectoderm specification, we hypothesized that CHD1L overexpression might promote neuroectodermal differentiation of hESCs. Thus, we investigated the influence of CHD1L overexpression in 137-CHD1L cells on the spontaneous differentiation of EBs. As shown in Supplementary Fig. S2A, EBs with CHD1L overexpression had very different morphology than EBs without CHD1L overexpression. Interestingly, although both ectoderm− and ectoderm+ EBs (Fig. 2A) could be found in EBs treated with or without DOX, there were more ectoderm+ structures in EBs with CHD1L overexpression than in the no DOX treatment control group (Fig. 2B). Moreover, although CHD1L mRNA levels were slightly downregulated in EBs without DOX treatment, PAX6 expression gradually increased during the formation of EBs. However, overexpression of CHD1L resulted in the upregulation of PAX6 during EB formation (Supplementary Fig. S2B, C). Next, we analyzed the expression of three germ layer-related genes in EBs with or without CHD1L overexpression. As shown in Fig. 2C, neuroectoderm markers (PAX6 and SOX1) were upregulated, whereas mesoderm (FLK1 and RUNX1) and endoderm markers (GATA4 and AFP) were downregulated. This suggests that CHD1L overexpression might promote neuroectodermal differentiation of hESCs and inhibit mesodermal and endodermal differentiation of hESCs.
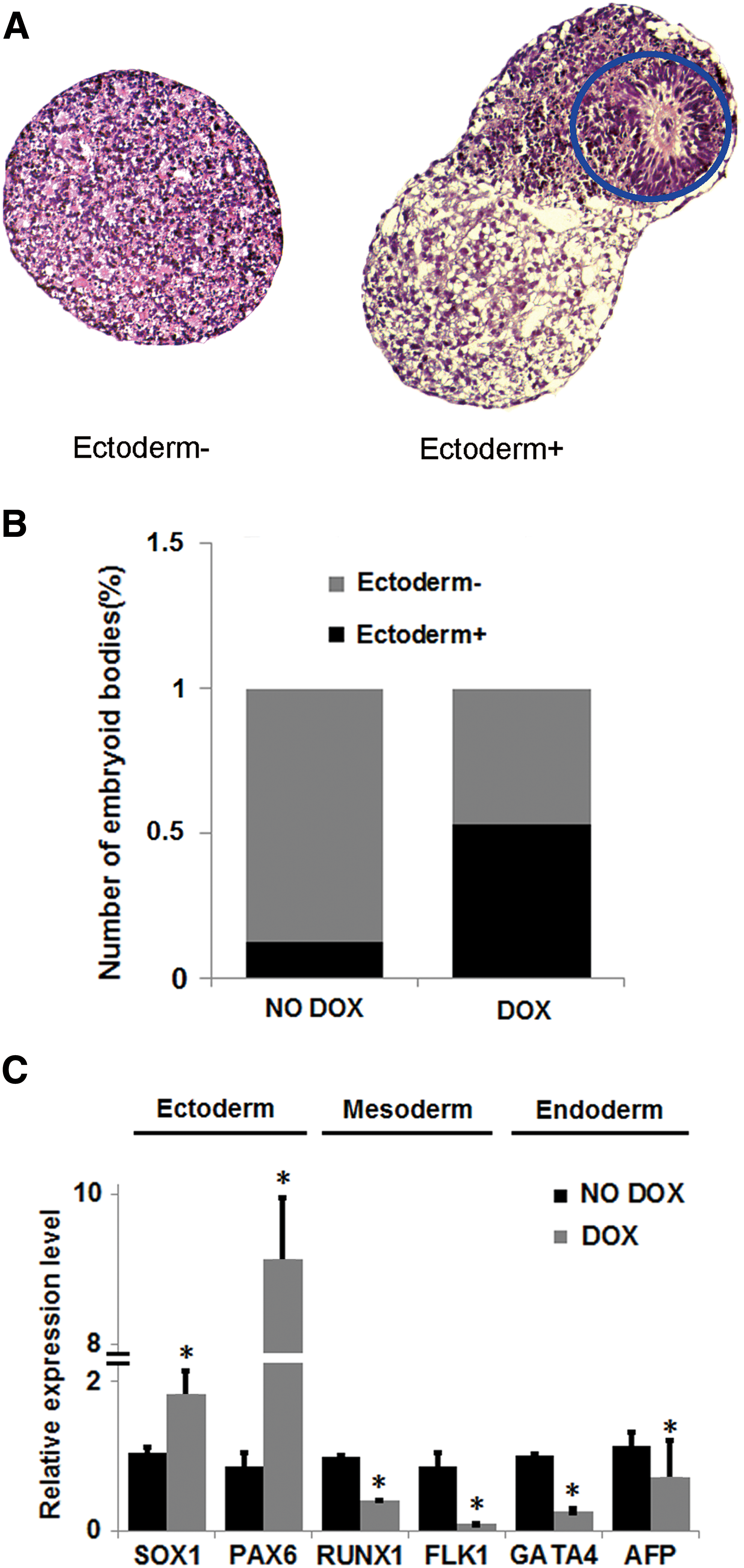
CHD1L overexpression promoted hESCs differentiation into neuroepithelial cells in spontaneous differentiation conditions.
We next investigated whether overexpression of CHD1L could promote the differentiation of hESCs into neuroectoderm in neuroectoderm induction conditions. The 137-CHD1L cells were cultured in suspension in EB medium for 4 days to form EBs, and then they were plated and fed with N2 medium supplemented with 20 ng/mL bFGF for several days. As shown in Fig. 3A, B, more neural rosettes formed more rapidly in hESC lines overexpressing CHD1L. To verify that the structures were really neural rosettes, we performed immunohistochemical analyses of early neural transcriptional factors commonly used as markers of neural rosettes, such as PAX6, SOX1, SOX2, and NESTIN. Staining with PAX6, SOX1, SOX2, NESTIN, and DAPI revealed the typical radial morphology of neural rosettes, and neural rosette markers only expressed on neural rosettes, except for SOX2 (it is also a pluripotency marker of hESCs) (Fig. 3C). A robust number of neurons were present which was positive for TUJ1 and MAP2 (marker of neuron) when these CHD1L-overexpressing rosettes were later cultured in neuronal medium for 6 days (Fig. 3D). Our results suggest that overexpression of CHD1L promotes neuroepithelial differentiation of hESCs.
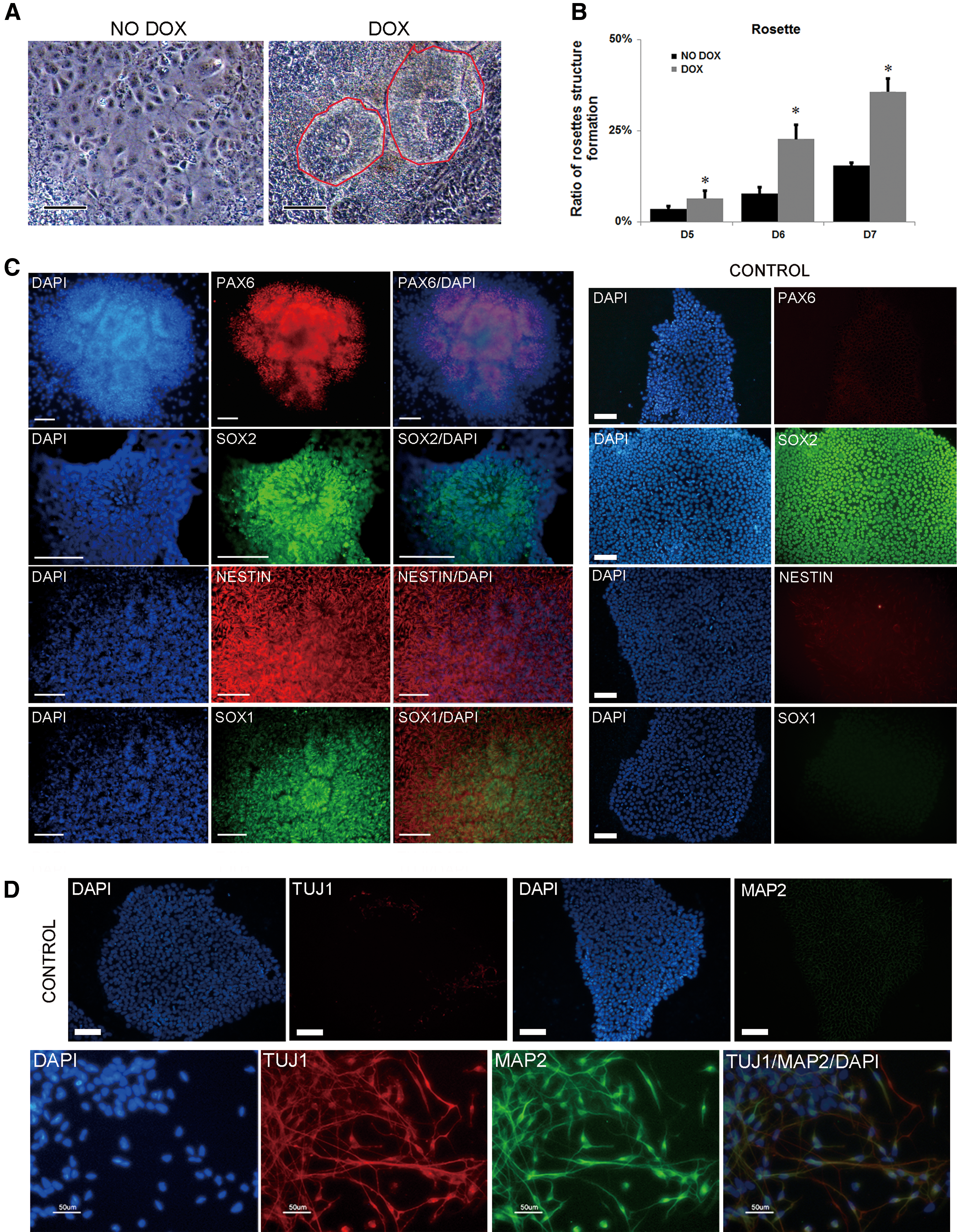
CHD1L overexpression promoted hESCs differentiated into neuroepithelial cells in induced differentiation condition.
Knockdown of CHD1L impaired the neuroectodermal differentiation of hESCs
To further investigate the role of CHD1L during hESC differentiation, we established hESC lines that stably integrated shRNA targeting CHD1L. To create stable CHD1L shRNA hESC lines, the DOX inducible (Tet-on) lentivector targeting an exon of CHD1L was used to produce lentivirus and transduce hESCs (Fig. 4A). Two colonies of CHD1L shRNA-1/2 hESC lines were used for subsequent experiments. After treatment with DOX, expression levels of CHD1L mRNA and protein in two 137-CHD1L cell lines were both downregulated (Fig. 4B, C).
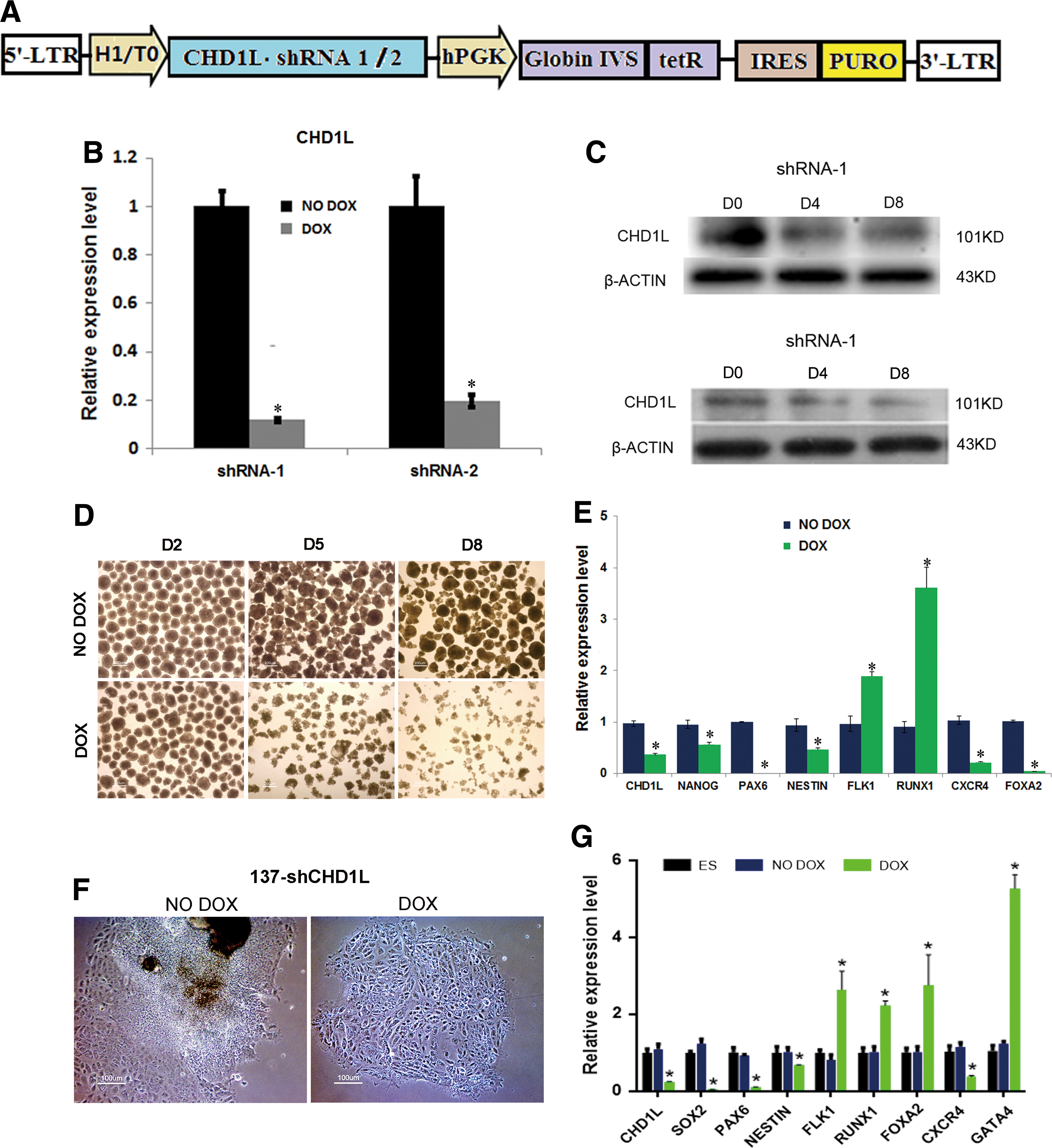
Knockdown of CHD1L impairs neuroepithelial differentiation in differentiating hESCs.
Our previous studies in hESCs have shown that knockdown of CHD1L compromised self-renewal, but did not affect pluripotency under self-renewal conditions. As shown in Supplementary Fig. S3A, overexpression of CHD1L significantly inhibited hESCs growth in two CHD1L knockdown lines. Immunocytochemistry staining of pluripotent genes (APK, POU5F1, NANOG, TRA-1-81, and SSEA4) showed no difference between DOX or NO DOX group in two CHD1L knockdown groups (Supplementary Fig. S3B). Results of western blotting and quantitative PCR also confirm this (Supplementary Fig. S3C). To determine whether CHD1L is indispensable for neuronal neuroepithelial differentiation of hESCs in differentiating conditions, 137-shCHD1L cells were induced to form EBs. Interestingly, EBs with CHD1L knockdown showed obvious changes in morphology (Fig. 4D and Supplementary Fig. S3D). EBs from the control and CHD1L knockdown groups could form sphere bodies at day 2. When cells were maintained in culture for an extended time, EBs from the control group remained compact, whereas EBs from the CHD1L knockdown group became smaller and loosely organized. To identify differences in the CHD1L knockdown group at the level of gene expression, we analyzed expression of CHD1L, a pluripotency marker (NANOG), and neuroectoderm markers (PAX6 and NESTIN). All of these markers were significantly downregulated, and PAX6 was downregulated to an undetectable level. Mesoderm markers (FLK1 and RUNX1) were upregulated and endoderm markers (CXCR4 and FOXA2) were downregulated (Fig. 4E). Similar results were observed in the 137-shCHD1L-2 cell line (Supplementary Fig. S3E). These results confirm that the expression of CHD1L was significantly downregulated during the differentiation of hESCs. Knockdown of CHD1L impaired expression of the neuroepithelial marker PAX6, which might impair the differentiation of hESCs into neuroectoderm.
To confirm that CHD1L knockdown specifically affects neuroectoderm differentiation, we differentiated 137-shCHD1L-1/2 cell lines using N2 supplement. The efficiency of neural induction in control (NO DOX) and CHD1L knockdown (DOX) lines was monitored by the appearance of cells with characteristic neuroepithelial cell morphology (neural rosette formation). Surprisingly, after 2 days of culture, control hESCs formed rosette structures, whereas hESCs with CHD1L knockdown did not form rosette structures, just a single layer of flat cells (Fig. 4F). RT-PCR analysis showed that the expression of neuroectoderm markers (PAX6, SOX2, and NESTIN) were significantly downregulated, whereas mesoderm (FLK1 and RUNX1) and endoderm markers (FOXA2 and GATA4) were all upregulated, with the exception of CXCR4 (Fig. 4G). We also confirmed the rosettes using immunocytochemistry, whereas control group could still form rosettes, as shown in Fig. 5A at day 4. The mesodermal and endodermal markers of rosettes are shown in Supplementary Fig. S3F. These rosette structures could still form neurons without DOX treatment, but the positive rate of rosettes was much lower than CHD1L overexpression group, as shown in Fig. 5B, whereas no rosette or neurons were found in DOX treatment group. Data from the knockdown and overexpression of CHD1L all suggest that CHD1L plays an important role in early neuroepithelial differentiation.

Characterization of neural rosettes and neuron-specific proteins.
CHD1L colocalized with PAX6 in the ventricular (germinal) zone in fetal mice
To determine whether CHD1L plays a role in mammalian neuroepithelial differentiation, we examined the expression of CHD1L in mice embryos by immunochemistry. As shown in Fig. 6, there was a high expression of CHD1L in the ventricular (germinal) zone (VZ), where PAX6 colocalized with CHD1L. PAX6 is a marker of neural stem or progenitor cells, and its expression persists in the VZ of the dorsal telencephalon. These results also suggest that CHD1L plays a role in neurogenesis.

CHD1L was colocalized with PAX6 in the VZ of mice embryos. Immunochemistry analysis of E14.5 mice embryos with antibodies for CHD1L
Discussion
Our previous studies and those of other groups have shown that the oncogenic function of CHD1L is associated with its antiapoptotic ability and the epithelial–mesenchymal transition. However, the exact role of CHD1L in the development of hESCs has not previously been reported. In this study, we demonstrate that CHD1L is indispensable for the neuroepithelial differentiation of hESCs, which is supported by the following evidence: overexpression of CHD1L promoted neuroepithelial differentiation, knockdown of CHD1L impaired neuroepithelial differentiation of hESCs, and CHD1L was highly expressed in the VZ where neuronal stem cells are enriched. We found that CHD1L overexpression impaired the ability of hESCs to proliferate and form colonies under self-renewal conditions. CHD1L overexpression led to the downregulation of pluripotency genes and the upregulation of the ectoderm marker, PAX6. These results differ from what has been shown in mouse ESCs (mESCs). In mESCs, overexpression of Chd1l increases its tumor susceptibility [14]. In our study, CHD1L overexpression did not affect the survival of hESCs, but growth more slowly (Supplementary Fig. S1E). Because hESCs and mESCs represent different stages of pluripotency [16], the results of our study may indicate that they also have distinct responses to the manipulation of CHD1L. A recent study showed that CHD1L and Parp1 interact and co-occupy the pluripotent loci of OCT4 and NANOG, and deletion of CHD1L would interfere with the acquisition of pluripotency during early reprogramming [17]. Consistent with our results, CHD1L overexpression upregulated PAX6 expression level, which was undetectable in undifferentiated hESCs. PAX6 overexpression would inactivate the promoter of pluripotency genes, such as OCT4 and NANOG, and convert hESCs into neuroectoderm [18]. We speculated that CHD1L may act directly or indirectly through the upregulation of PAX6 expression, then may inactivate some pluripotency genes and lead to the differentiation of hESCs into neuroectoderm.
Overexpression of CHD1L altered the morphology of EBs and induced EBs to form ectoderm+ structures during spontaneous differentiation. CHD1L overexpression resulted in the upregulation of neuroectoderm markers (PAX6 and SOX1) and the downregulation of mesoderm (RUNX1 and FLK1) and endoderm markers (GATA4 and AFP), which were consistent with the changes in histomorphology. Moreover, with induction by N2 supplement, overexpression accelerated the formation of neural rosettes. The EBs acquired this ability before culturing in N2 induction conditions, as previous studies have shown that the expression level of PAX6 had already been upregulated under self-renewal conditions. These two results show that overexpression of CHD1L induces hESCs to differentiate into neuroepithelium in differentiation conditions. In the future, studies on the overexpression of forebrain, midbrain, hindbrain, and spinal cord-associated genes during differentiation will facilitate our understanding of the mechanisms that regulate neurogenesis.
Our study has shown that knockdown of CHD1L in hESCs in the undifferentiated condition significantly impairs proliferation and colony formation, but does not affect pluripotency (Supplementary Fig. S3A–C). This result is partially inconsistent with research on mESCs. In mESCs, knockdown had no effect on proliferation and pluripotency [14], which may be due to differences in the manipulation of CHD1L between hESCs and mESCs. However, CHD1L knockdown during spontaneous differentiation caused downregulation of neuroepithelial genes (PAX6 and NESTIN), and PAX6 expression was undetectable. In the future, research on how CHD1L regulates the promoter activity of PAX6 will facilitate our understanding of the internal mechanism. Moreover, under induction by N2 supplement, CHD1L knockdown decreased the formation of rosette structures. This was confirmed by a decrease in neuroectoderm genes (PAX6 and NESTIN) and an increase in mesoderm (FLK1 and RUNX1) and endoderm-associated genes (GATA4). These results suggest that the lack of CHD1L specifically affected neuroectoderm differentiation and led hESCs differentiate into mesoderm and endoderm, although there was a slight downregulation in an endoderm gene (FOXA2). Further research will be necessary to establish CHD1L knockout hESCs lines and confirm the role of CHD1L during neuroectoderm differentiation.
Using in vivo experiments, we demonstrated that CHD1L was highly expressed in the VZ at E14.5, where it is the source of the central nervous system and enriched neural stem cells. Moreover, CHD1L partially colocalized with PAX6, a marker of neural stem cells. This suggests that CHD1L plays a role in embryonic neurogenesis. Establishing a line of CHD1L knockout mice will help us to determine whether the deletion of CHD1L would cause defects in nervous system development. In humans, CHD1L expression was highest in the fetal brain [19], which is consistent with our result on hESCs. These data suggest that CHD1L is indispensable for the development of human embryonic neuroepithelium.
Our results suggest that CHD1L promotes neuroepithelial differentiation of hESCs, and provides new insight into the roles of CHD1L in regulating self-renewal and differentiation of hESCs. Further investigations of the molecular mechanisms by which CHD1L regulates neuroepithelial differentiation will contribute to our understanding of hESCs differentiation and specification in early human development.
Footnotes
Acknowledgments
The authors thank Dr. Xinyuan Guan for providing the CHD1L plasmid. This study was supported by the National Natural Science Foundation of China (81372627), Natural Science Foundation of Hunan Province (13JJ3038), Graduate Student Research Innovation Project of Hunan Province (2016zzts113), and Innovation Funds for postgraduate of the Central South University (72150050335).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.