
Abstract
Induced pluripotent stem (iPS) cells possess pluripotency and self-renewal ability. Therefore, iPS cells are expected to be useful in regenerative medicine. However, iPS cells form malignant immature teratomas after transplantation into animals, even after differentiation induction. It has been suggested that undifferentiated cells expressing Nanog that remain after differentiation induction are responsible for teratoma formation. Various methods of removing these undifferentiated cells have therefore been investigated, but few methods involve morphological approaches, which may induce less cell damage. In addition, for cells derived from iPS cells to be applied in regenerative medicine, they must be alive. However, detailed morphological analysis of live undifferentiated cells has not been performed. For the above reasons, we assessed the morphological features of live undifferentiated cells remaining after differentiation induction as a basic investigation into the clinical application of iPS cells. As a result, live undifferentiated cells remaining after differentiation induction exhibited a round or oval cytoplasm about 12 μm in diameter and a nucleus. They exhibited nucleo-cytoplasmic (N/C) ratio of about 60% and eccentric nuclei, and they possessed partially granule-like structures in the cytoplasm and prominent nucleoli. Although they were similar to iPS cells, they were smaller than live iPS cells. Furthermore, very small cells were present among undifferentiated cells after differentiation induction. These results suggest that the removal of undifferentiated cells may be possible using the morphological features of live iPS cells and undifferentiated cells after differentiation induction. In addition, this study supports safe regenerative medicine using iPS cells.
Introduction
I
To resolve these problems of ES cells, Yamanaka and colleagues established induced pluripotent stem (iPS) cells with self-renewal ability and pluripotency equivalent to those of ES cells by introducing reprogramming factors (Oct3/4, Klf4, Sox2, c-Myc) into mouse and human somatic cells [3,4]. iPS cells can be produced using the somatic cells of a patient. Therefore, they overcome the ethical and tissue rejection problems of ES cells and can be used for transplantation into patients [4].
However, iPS cells form malignant immature teratomas after transplantation into animals [5]. When iPS cells are clinically applied, they are used after the induction of differentiation into target cell types. However, transplanted cells that have undergone differentiation induction also have the ability to form tumors [5 –7]. In our previous study, we confirmed that the tumors formed by these cells after differentiation induction were malignant immature teratomas [5]. It has been suggested that undifferentiated cells expressing Nanog that remain after differentiation induction are responsible for teratoma formation [5].
In mouse iPS cells, Nanog expression can be visually detected as fluorescence by introducing the green fluorescent protein (GFP) gene downstream of the Nanog promoter [8]. Therefore, in mouse iPS cells with the Nanog-GFP gene fusion, it is possible to remove undifferentiated cells expressing Nanog before transplantation.
However, when human iPS cells are clinically applied, methods that do not depend on genetic modification are necessary. For the above reasons, we report here the morphological features of live undifferentiated cells that remain after differentiation induction, termed residual Nanog-GFP-positive after differentiation (NPAD) cells, as part of a basic investigation into the clinical application of iPS cells.
Materials and Methods
Cell culture
The mouse iPS cell line iPS-MEF-Ng-20D-17 (APS0001; RIKEN Bioresource Center, Japan) used in this study expresses GFP under the control of the Nanog promoter [8]. Undifferentiated cells express Nanog, but its expression disappears with differentiation [8,9].
Gelatin from porcine skin (Sigma-Aldrich) was added as a 0.1% aqueous solution to a cell culture dish (Becton Dickinson Labware) and incubated at 37°C for 30 min, and a gelatin coat was applied to the culture dish. EmbryoMax® Primary Mouse Embryo Fibroblasts (MEFs; Millipore) treated with mitomycin C (Kyowa Hakko Kogyo Co., Ltd., Japan) were seeded in gelatin-coated culture dishes. The iPS cell line was seeded on mitomycin C-treated MEFs and cultured in ES medium [15% fetal bovine serum (FBS; MP Biomedicals), 1% non-essential amino acids (Millipore), 1% nucleosides (Millipore), and 1% Dulbecco's modified Eagle's medium (Wako Pure Chemical Industries, Ltd., Japan) containing 1%
iPS cells were passaged once every 2–3 days. For the dispersion of iPS cells, 2.5 g/L trypsin EDTA solutions were used.
Differentiation induction of iPS cells
iPS cells dispersed with enzyme (trypsin) were seeded at 5 × 104/mL on a low-adhesion culture dish (Corning) and suspension-cultured. For the culture, 10 mL of feeder cell culture medium (ES cell culture medium without LIF) was used. On day 7 after the seeding, the formed embryoid bodies (most of the embryoid bodies, including both GFP-positive and -negative cells) were transferred to gelatin-coated cell culture dishes, and adhered and cultured for 3 days (Fig. 1).
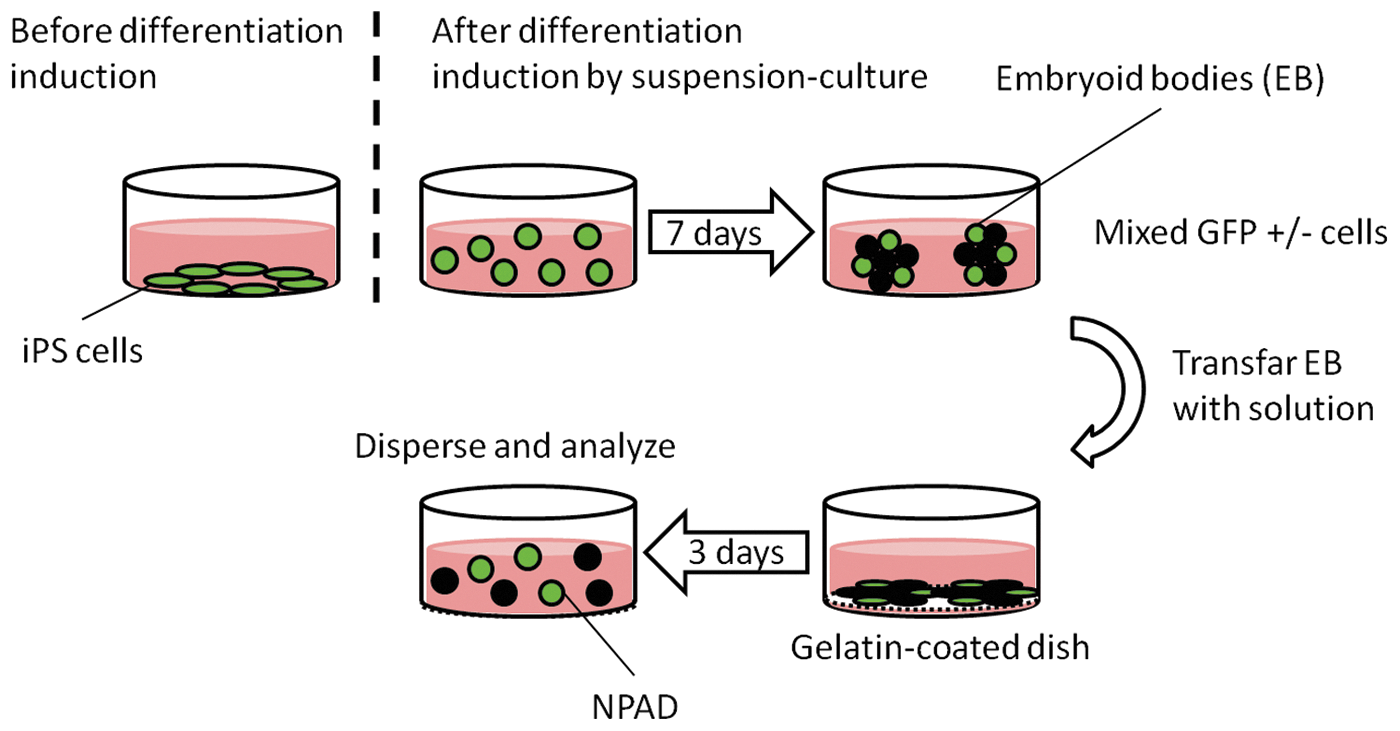
Induction of iPS cell differentiation. iPS cells dispersed with trypsin were seeded onto a low-adhesion culture dish and cultured, using the feeder cell culture medium. On day 7 after the seeding, the formed embryoid bodies were transferred to gelatin-coated cell culture dishes, where they were cultured for 3 days. Cells expressing Nanog-GFP for a total of 10 days were defined as NPAD cells, which were dispersed by enzyme and analyzed. GFP, green fluorescent protein; iPS, induced pluripotent stem; NPAD, Nanog-GFP-positive after differentiation.
Cells expressing Nanog-GFP even after the induction of differentiation for a total of 10 days were defined as NPAD cells (Fig. 2). NPAD cells were dispersed using an enzyme and morphologically analyzed.

NPAD cells among Nanog-GFP-negative cells derived from the embryoid bodies.
Cell sorting
iPS, NPAD, and GFP-negative cells were stained with 1.0 μg/mL propidium iodide solution (PI; Sigma-Aldrich) for 5 min. Stained cells were spun down at 250 g for 5 min and washed with sorting buffer [minimum essential medium (Gibco®)+2% FBS] and spun down again at 250 g for 5 min. Pellets were resuspended in 3 mL sorting buffer for sorting. iPS (PI−GFP+), NPAD (PI−GFP+), and GFP-negative (PI−GFP−) cells were sorted on a fluorescence-activated cell sorter (FACS; BD FACSAria™III, BD). Sorted cells were used for morphological analysis, growth rate, and colony formation assays.
Growth rate
NPAD and GFP-negative cells were seeded at 1.5 × 105 cells/well in six wells of a 48-well plate seeded with MEFs, and dispersed in three wells after 24 and 96 h. The Automated Cell Counter (TC 20; Bio-Rad) was used to determine cell numbers. For culturing, we used the feeder cell culture medium.
Colony formation assay
Equal amounts of 2 × ES cell culture medium and 2% agar (Difco Agar Noble, BD) were mixed (2 mL), poured into 6-cm dish, and allowed to solidify (bottom agar). The same culture medium and 1% agar seeded with iPS or NPAD cells at 105 cells/dish were mixed in equal amounts, and 2-mL volumes of this mixture were layered over the bottom agar. Furthermore, 2 mL of feeder cell medium was added and the cells were cultured for 20 days.
Morphological analysis
For living cells, iPS and NPAD cells were detected and observed using a confocal laser scanning microscope (FV 10i; Olympus, Japan) with or without GFP fluorescence as an indicator. iPS and NPAD cells were photographed under the confocal laser scanning microscope, and their morphological features were determined using the image analysis software ImageJ [10]. Features analyzed included the nuclear major axis, nuclear minor axis, nuclear area, cytoplasmic major axis, cytoplasmic minor axis, cytoplasmic area, and nucleo-cytoplasmic (N/C) ratio.
In addition, similar morphological analysis was performed on cells subjected to Papanicolaou (Pap.) staining and May-Grunwald's Giemsa (Giemsa) staining. In this case, the cells were separated by FACS depending on GFP fluorescence. Obtained cells were smeared, fixed, and then stained.
Statistical analysis
Statistically significant differences between groups (P < 0.05) were determined using the Mann–Whitney U-test.
Results
Cell sorting
We were able to sort iPS (PI−GFP+), NPAD (PI−GFP+), and GFP-negative (PI−GFP−) cells (Fig. 3).

Isolation of iPS, NPAD and GFP-negative cells using a fluorescence-activated cell sorter.
Growth rate
We considered the number of cells at 24 h as 100%, and NPAD cell number at 96 h was approximately eightfold higher, while that of the GFP-negative cells was shown to be increased approximately twofold (Fig. 4).
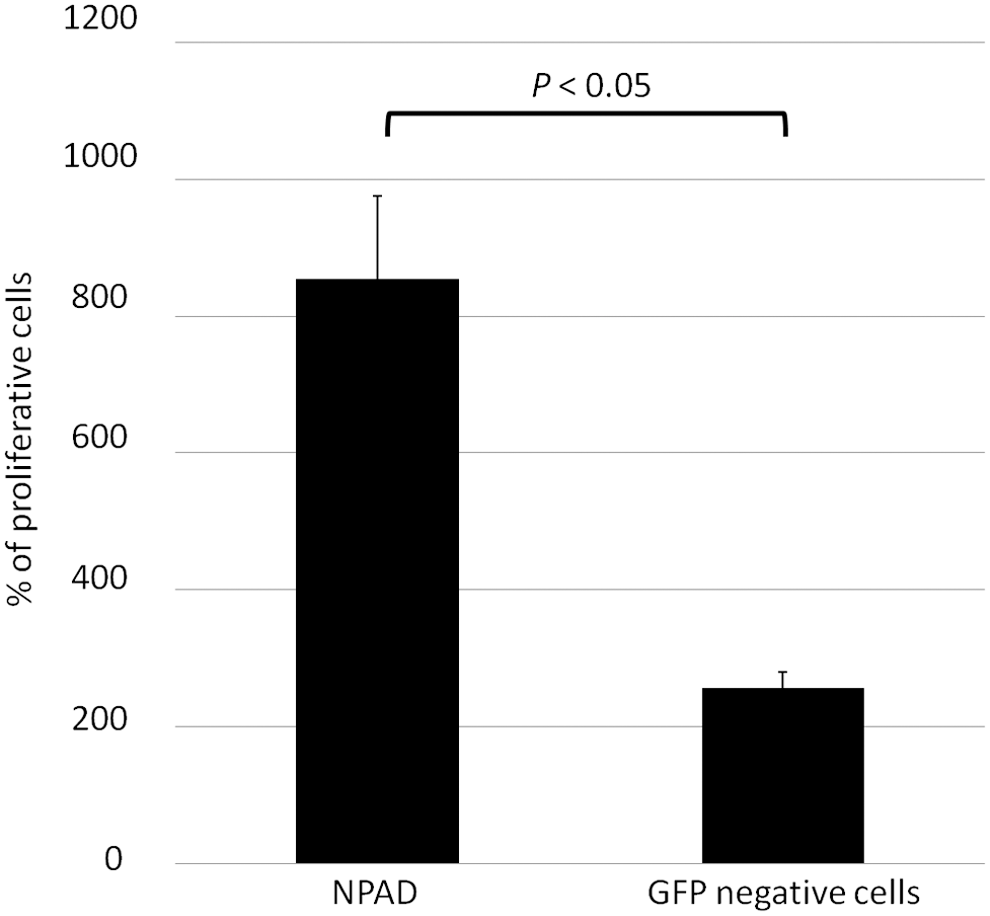
Growth rate of NPAD and GFP-negative cells. y-axis, the percentage of cells at 96 h, compared with that at 24 h (100%). NPAD cells grew faster than GFP-negative cells.
Colony formation assay
Colony formation assay results showed that four colonies were formed following the seeding of NPAD cells (Fig. 5A), but a large number of colonies were formed following the seeding of iPS cells (Fig. 5B).
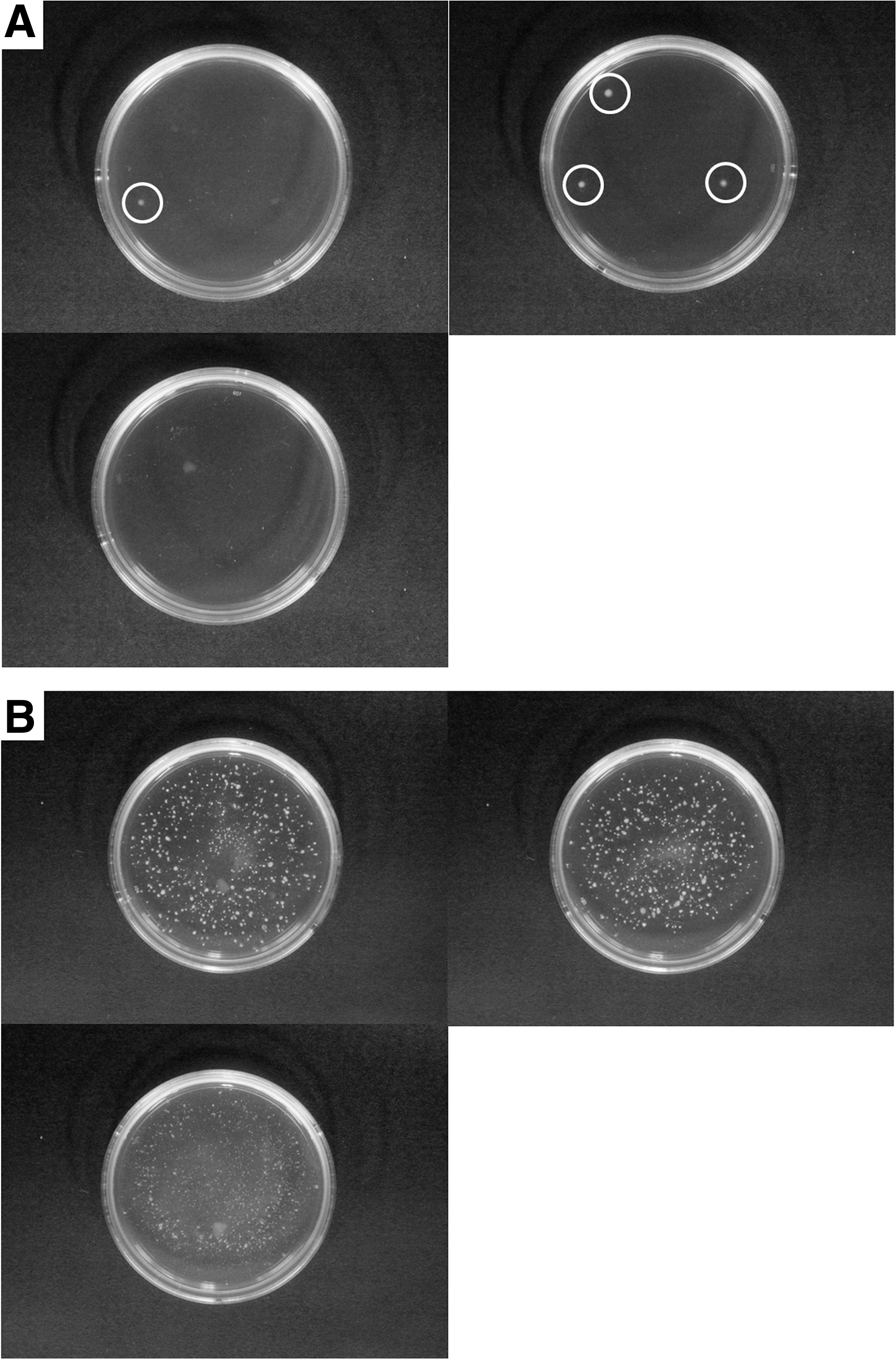
Colony formation of NPAD cells and iPS cells.
Analysis of live cells
According to the results of confocal laser scanning microscope analysis, iPS cells exhibited round or oval cytoplasms and nuclei (Fig. 6A). The nuclei of iPS cells tended to be eccentric, and one or more nucleoli were observed in the nucleus (Fig. 6B). There were regions with granule-like structures in the cytoplasms of iPS cells, while other regions in the cytoplasm were unstructured (Fig. 6C). Some iPS cells had round inclusion-like structures in the cytoplasm. According to the fluorescence images, iPS cells were Nanog-GFP-positive in both the cytoplasm and nucleus (Fig. 6A′–C′).
Viable iPS cells.
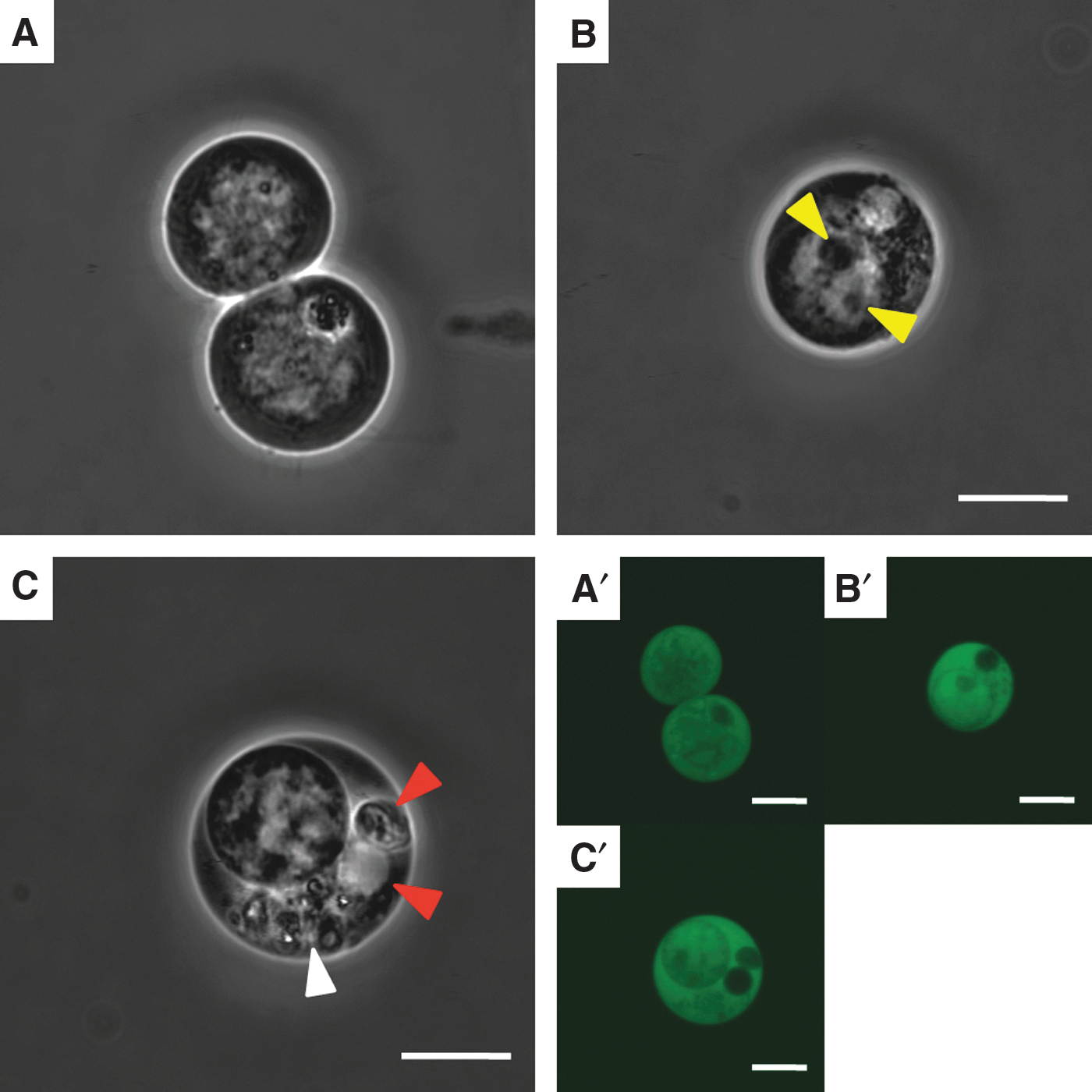
NPAD cells also exhibited round or oval cytoplasms and nuclei (Fig. 7A). The nuclei of NPAD cells tended to be eccentric, and one or more nucleoli were observed in the nucleus (Fig. 7A). NPAD cells showing round inclusion-like structures in the cytoplasm were observed (Fig. 7A). The nuclei of NPAD cells were brighter than the cytoplasm, and almost no structures other than the nucleolus was found in the nucleus (Fig. 7B). In addition, some NPAD cells had granule-like structures, but the rest of the cytoplasm was unstructured (Fig. 7B). According to fluorescence imaging, NPAD cells were Nanog-GFP-positive in both the cytoplasm and nucleus (Fig. 7A′–C′).
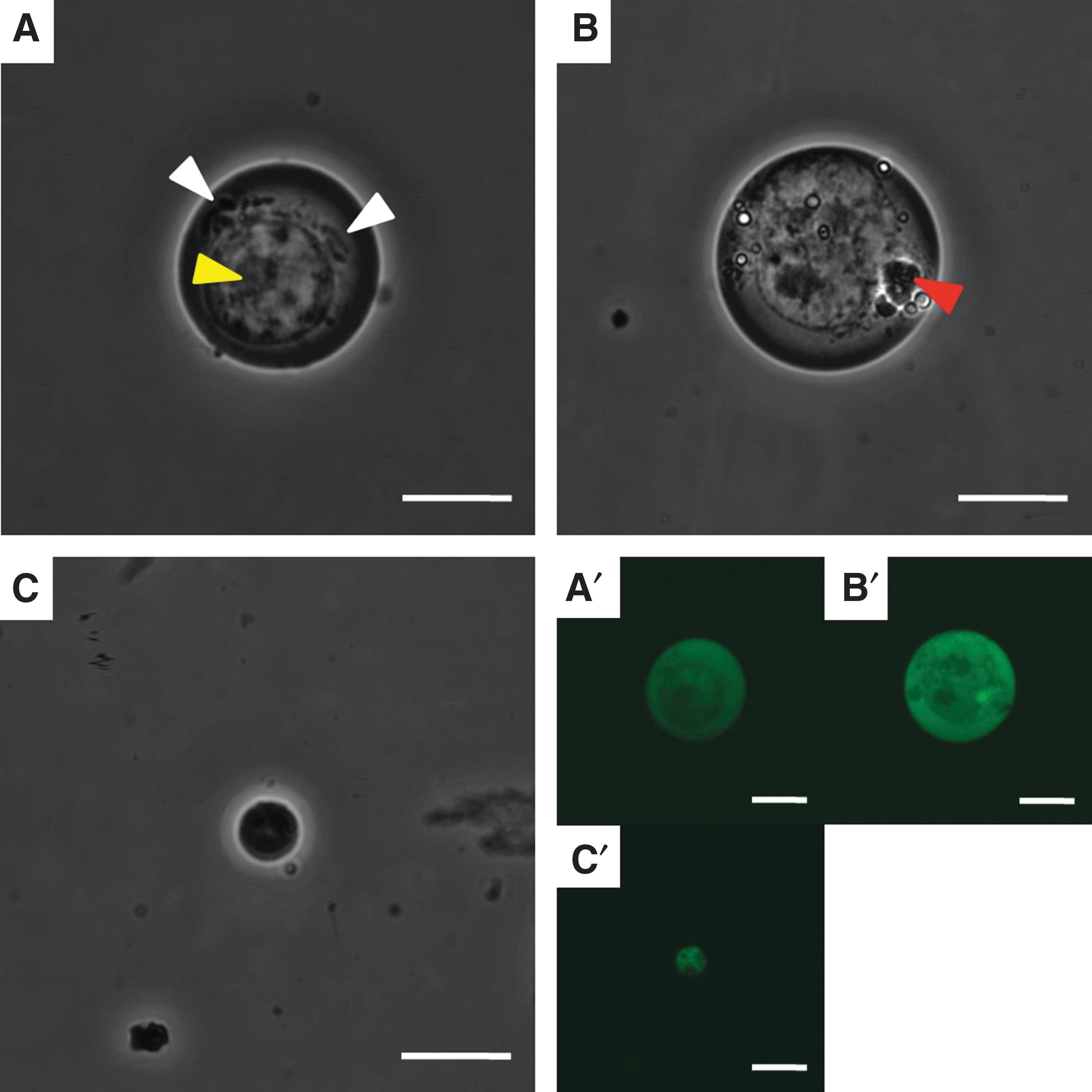
Viable NPAD cells.
In addition, extremely small cells were observed among NPAD cells (Fig. 7C). These small cells had round cytoplasms and nuclei (Fig. 7C). The interior of the nucleus was pyknotic, and the internal structures were unclear (Fig. 7C). In fluorescence images, small cells were Nanog-GFP-positive in both the cytoplasm and nucleus (Fig. 7C′).
According to analysis by ImageJ, iPS cells exhibited an average nuclear major axis of 10 ± 2 μm (mean ± standard deviation), nuclear minor axis of 9 ± 1 μm, nuclear area of 76 ± 26 μm2, cytoplasm major axis of 13 ± 2 μm, cytoplasm minor axis of 12 ± 1 μm, cytoplasm area of 120 ± 33 μm2, and N/C ratio of 63% ± 7% (Table 1). In contrast, NPAD cells exhibited an average nuclear major axis of 9 ± 2 μm, nuclear minor axis of 8 ± 2 μm, nuclear area of 62 ± 28 μm2, cytoplasm major axis of 12 ± 2 μm, cytoplasm minor axis of 11 ± 1 μm, cytoplasm area of 99 ± 28 μm2, and N/C ratio of 60% ± 14% (Table 1).

Following statistical analysis, statistically significant differences between iPS and NPAD cells were observed in terms of nuclear area, cytoplasm area, nuclear major axis, nuclear minor axis, cytoplasm minor axis, and N/C ratio (Table 1). Furthermore, NPAD cells were significantly smaller than iPS cells (Fig. 8).
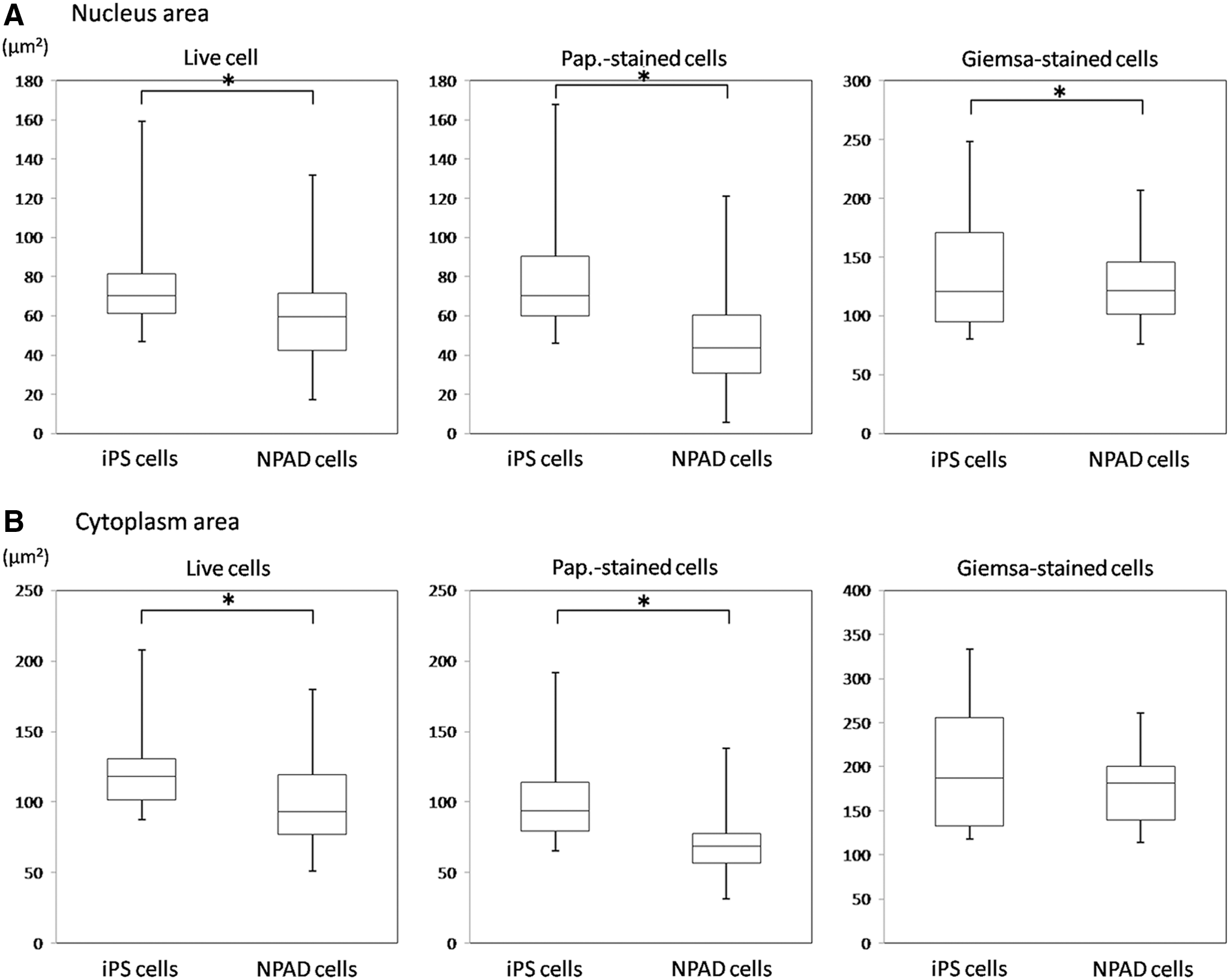
Distributions of the nuclear and cytoplasmic areas in iPS and NPAD cells.
Analysis of Pap.-stained cells
According to the microscopic analysis of Pap.-stained cells, iPS cells exhibited round or oval cytoplasms that were moderately to highly stained light green, with round or oval nuclei with euchromatic structures. There were like bare nuclei cells (Fig. 9A). The nuclei of iPS cells tended to be eccentric, the nuclear margins were clear, and one or more nucleoli were found in the nucleus (Fig. 9A). In the cytoplasms of some iPS cells, round inclusion-like structures that were moderately to highly stained light green were observed (Fig. 9A).

Papanicolaou-stained iPS and NPAD cells.
Similarly, NPAD cells exhibited round or oval cytoplasms that were moderately to highly stained light green, with round or oval nuclei (Fig. 9B). The nuclei of NPAD cells tended to be eccentric, the nuclear margins were clear, and one or more nucleoli were observed in the nucleus (Fig. 9B). Some NPAD cells had round inclusion-like structures that were moderately to highly stained light green in the cytoplasm (Fig. 9B). In addition, some small NPAD cells were observed (Fig. 9B). Small cells were characterized by round cytoplasms moderately to highly stained light green with pyknotic nuclei (Fig. 9B).
Following analysis by ImageJ, iPS cells had a nuclear major axis of 10 ± 2 μm (mean ± standard deviation), nuclear minor axis of 9 ± 1 μm, nuclear area of 75 ± 23 μm2, cytoplasm major axis of 12 ± 2 μm, cytoplasm minor axis of 10 ± 2 μm, cytoplasm area of 97 ± 31 μm2, and N/C ratio of 78% ± 10% (Table 2).

In contrast, NPAD cells had a nuclear major axis of 8 ± 2 μm, nuclear minor axis of 7 ± 1 μm, nuclear area of 46 ± 19 μm2, cytoplasm major axis of 10 ± 1 μm, cytoplasm minor axis of 9 ± 1 μm, cytoplasm area of 71 ± 21 μm2, and N/C ratio of 64% ± 17% (Table 2). There were statistically significant differences in the areas of the nucleus and cytoplasm between iPS and NPAD cells, in addition to the nucleus and cytoplasm major axis, minor axis, and N/C ratio (Table 2). Furthermore, NPAD cells were significantly smaller than iPS cells (Fig. 8).
Analysis of Giemsa-stained cells
According to microscopic analysis of Giemsa-stained cells, iPS cells exhibited round or oval cytoplasms that were basophilically stained and had round or oval nuclei with fine granular chromatin (Fig. 10A). The nuclei of iPS cells tended to be eccentric, the nuclear margins were clear, and one or more nucleoli were observed in the nuclei (Fig. 10A). In addition, perinuclear haloes were observed in iPS cells (Fig. 10A).
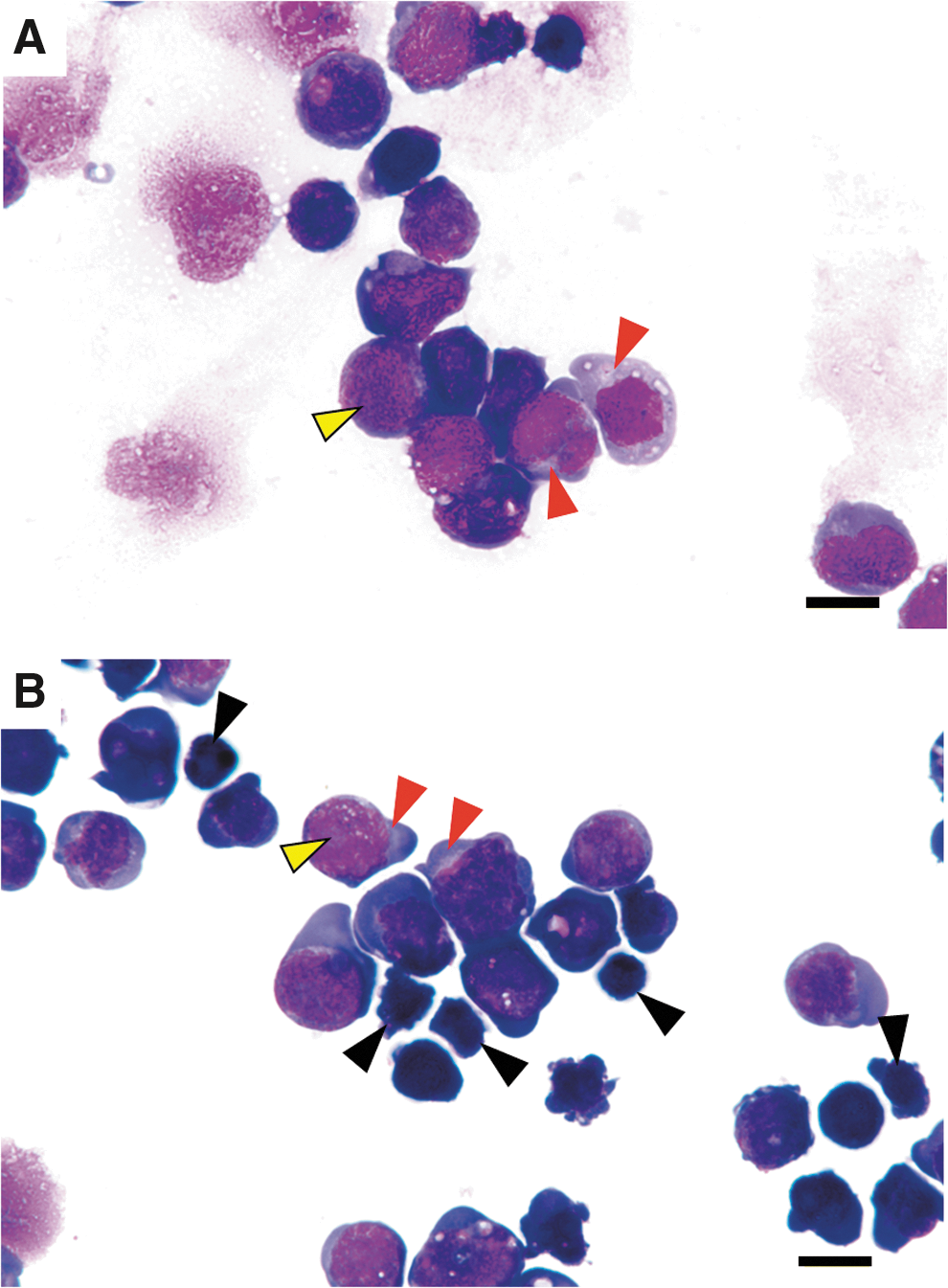
Giemsa-stained iPS and NPAD cells.
NPAD cells exhibited round or oval cytoplasms that were basophilically stained, and the nuclei had fine granular chromatin (Fig. 10B). The nuclei of NPAD cells tended to be eccentric, the nuclear margins were clear, and one or more nucleoli were observed in the nuclei (Fig. 10B). Perinuclear haloes were also observed in NPAD cells (Fig. 10B). In addition, small cells were observed among the population of NPAD cells (Fig. 10B). Small cells exhibited basophilic staining of the cytoplasm and pyknotic nuclei (Fig. 10B).
Analysis by ImageJ indicated that iPS cells had a nuclear major axis of 14 ± 2 μm (mean ± standard deviation), nuclear minor axis of 12 ± 2 μm, nuclear area of 127 ± 43 μm2, cytoplasm major axis of 17 ± 3 μm, cytoplasm minor axis of 14 ± 3 μm, cytoplasm area of 190 ± 66 μm2, and N/C ratio of 68% ± 8%. NPAD had a nuclear major axis of 14 ± 3 μm, nuclear minor axis of 11 ± 2 μm, nuclear area of 124 ± 43 μm2, cytoplasm major axis of 17 ± 2 μm, cytoplasm minor axis of 14 ± 3 μm, cytoplasm area of 184 ± 56 μm2, and N/C ratio of 67% ± 12%. Results for iPS and NPAD cells were significantly different in terms of nuclear area, nuclear major axis, nuclear minor axis, cytoplasm major axis, and cytoplasm minor axis (Table 3). Furthermore, the nuclei of NPAD cells were significantly smaller than those of iPS cells (Fig. 8).

Discussion
iPS cells are pluripotent stem cells that are able to differentiate into various cell types constituting the different tissues of the body, and they possess self-renewal ability. iPS cells that can regenerate and treat the damaged organs and cells of patients are expected to be useful to the field of regenerative medicine.
However, iPS cells form malignant immature teratomas after transplantation into animals [5]. When iPS cells are clinically applied, they are used after differentiation induction into target cell types, but if undifferentiated cells remain, they form teratomas after transplantation into animals as well [5 –7]. In particular, we reported in a previous study that tumors formed by iPS cells or cells derived from iPS cells were malignant immature teratomas [5]. It has been suggested that NPAD cells are responsible for this malignant immature teratoma formation [5].
In this study, NPAD cells were shown to have anchorage-independent growth, and their growth rate was confirmed to be significantly higher than that of the GFP-negative cells, indicating that NPAD cells cause tumorigenesis. Therefore, before cells derived from iPS cells can be applied in regenerative medicine, it is necessary to remove NPAD cells, including residual iPS cells [5], to remove the cause of malignant immature teratoma formation. In addition, when cells derived from iPS cells are applied in regenerative medicine, live cells must be used.
In this study, we revealed the morphological features of live iPS and NPAD cells. Based on our results, isolated, live iPS cells have round or oval cytoplasms and nuclei. In addition, there are regions of the cytoplasm with granule-like structures, but most of the cytoplasm is unstructured. Most of the insides of the nuclei were bright without a nucleolus. In addition, Pap.-stained iPS cells were bare nucleus-like cells with round or oval cytoplasms and euchromatic nuclei. Giemsa-stained iPS cells had basophilic round or oval cytoplasms and fine granular chromatin. The observations derived from stained cells were therefore similar to those of live iPS cells.
According to Zeuschner et al., iPS cells in colonies cultured with MEFs exhibited poor intracellular organelles as compared with MEFs according to analysis using an electron microscope [11]. Furthermore, they reported that iPS and ES cells had euchromatin and clear nucleoli [11,12].
We revealed that isolated, live NPAD cells exhibited round or oval cytoplasms and nuclei. In addition, granule-like structures appeared in some cytoplasmic regions, while most other regions were unstructured. Regarding the nucleus, it had not any more structure except for clear nucleoli.
In addition, small cells were observed among NPAD cells. These small cells exhibited round cytoplasms and nuclei. Virant-Klun et al. reported that pluripotent stem cells in the human ovarian surface epithelium were small and round (2–4 μm). They expressed SSEA-4, OCT-4, NANOG, SOX-2, and c-KIT as embryonic markers and formed embryoid-like structures in vitro [13]. Zuba-Surma et al. reported that SSEA-1, Oct-4, Nanog, and Rex-1-expressing stem cells derived from mouse bone marrow were extremely small (3.63 ± 0.09 μm) [14]. These reports support our evidence for the presence of small cells among NPAD cells.
As mentioned above, the morphological analysis in this study is applicable to human iPS cells, the genetic alteration of which presents ethical challenges. We observed that cells with different sizes can be observed in the population of NPAD cells, and these were shown to be considerably different morphologically from the iPS cells. The results of this study, showing that the removal of the pluripotent stem cells is possible with morphological techniques, may help improve the safety of iPS cell application in regenerative medicine.
Currently, methods for removing human pluripotent stem cells (hPSCs) that remain after differentiation induction include the introduction of suicide genes into hPSCs or the selective killing of hPSCs using cytotoxic antibodies, chemical inhibitors, or specific antibodies against hPSCs [15 –18]. However, these methods suffer from several issues with regards to specificity, efficacy, and safety for use in cell therapy for humans.
Our morphological study reveals the possibility of detecting and removing NPAD cells without the cell damage caused by genetic modification. Therefore, we provide a possible approach for using hPSCs while overcoming the ethical problems involving genetic modification and safety problems involving drugs used to kill hPSCs. Our findings therefore have important implications for the realization of regenerative medicine using iPS cells.
Footnotes
Author Disclosure Statement
No competing financial interests exist.