
Abstract
Neural crest (NC) cells are a migratory, multipotent population giving rise to numerous lineages in the embryo. Their plasticity renders attractive their use in tissue engineering-based therapies, but further knowledge on their in vivo behavior is required before clinical transfer may be envisioned. We here describe the isolation and characterization of a new mouse embryonic stem (ES) line derived from Wnt1-CRE-R26 Rosa TomatoTdv blastocyst and show that it displays the characteristics of typical ES cells. Further, these cells can be efficiently directed toward an NC stem cell-like phenotype as attested by concomitant expression of NC marker genes and Tomato fluorescence. As native NC progenitors, they are capable of differentiating toward typical derivative phenotypes and interacting with embryonic tissues to participate in the formation of neo-structures. Their specific fluorescence allows purification and tracking in vivo. This cellular tool should facilitate a better understanding of the mechanisms driving NC fate specification and help identify the key interactions developed within a tissue after in vivo implantation. Altogether, this novel model may provide important knowledge to optimize NC stem cell graft conditions, which are required for efficient tissue repair.
Introduction
N
Given their plasticity and possible cell derivatives, the NC-derived stem cells (NCSC) have raised a great interest for tissue regeneration and repair [8]. Several NCSC niches have already been described in the adult, in dorsal root ganglia [9], cornea [10], heart [11,12], bone marrow [9], dental pulp, and periodontal ligament [13 –15]. However, due to the limited availability of these cell sources, their use for therapies remains a challenge. Finding alternative NCSC unlimited sources has, therefore, been the focus of many laboratories and the differentiation of NCSC from pluripotent cells, embryonic stem (ES) cells, or induced pluripotent stem cells has appeared an attractive alternative [16 –20].
Although the gene regulatory network that governs the NC differentiation pathways is well described [21,22], there is still poor understanding of how NCSC behave in vivo and interact with their environment during development and in pathological situations. This is essential for developing successful therapeutic strategies. In this context, an important point is to obtain cellular tools allowing a precise tracking of the in vivo fate of NCSC, depending on where and when they are implanted. Labeled-lineage-specific ES cell reporter lines constitute such a tool and must facilitate the real-time monitoring of differentiation in live cells as well as the identification of factors governing these processes [23,24]. They easily enable purifying specific cell populations from heterogeneous differentiated progeny and to follow up their fate in normal physiological and pathological environments.
The Wnt1-CRE mouse is a well-established model [25], extensively used for lineage tracing of pre-migratory NC cells [26 –30], where the Wnt1 gene is activated in the NC at the time of its specification [31,32]. We have derived and characterized a novel fluorescent reporter ES cell line obtained from Wnt1-CRE-R26 Rosa TomatoTd mouse blastocysts. These ES cells should, therefore, “constitutively” express the Tomato protein, on reaching an NC phenotype after induction, thus giving access to an in vivo as well as in vitro follow-up of their fate.
Several protocols have been developed for deriving NC cells from ES cells; they usually involve forming three-dimensional (3D) embryoid bodies (EBs) or ES cell culturing in media containing complex or undefined components [16,18,19,33 –35]. We have recently set up a simple straightforward NC differentiation protocol, in monolayer, using a defined serum-free differentiation medium, and a treatment by fibroblast growth factor (FGF)2 followed by bone morphogenetic protein (BMP)4 [36] and used it to facilitate the follow-up of the Wnt1-CRE Tomato cell differentiation process.
Here, we show that this novel Wnt1-CRE-R26 Rosa TomatoTd ES cell line presents all the characteristics expected for ES cells and can be efficiently committed toward an NC cell fate following our induction protocol. Once specified, the ES-NC-derived (ES-NCD) cells can be sorted out, based on their Tomato fluorescence, and give rise in vitro, on specific signaling, to typical NC cell type derivatives such as osteoblasts, neurons, glial cells, or pericytes, therefore faithfully mimicking the differentiation process of the post-migratory cranial NC cells in vivo. Further, the specified ES-NCD cells are able to respond in vivo and ex vivo to the signaling action of embryonic tissues, thus making this novel cell model a powerful tool to get insight into the interactions they develop in vivo within a host tissue.
Materials and Methods
Mice and tissues
Wnt1-CRE transgenic mice [25] were crossed with fluorescent reporter B6.Cg-Gt(ROSA)26Sortm9(CAGtdTomato)/J mice (The Jackson Laboratory, Janvier Labs, Saint Berthin, France; 007909) to obtain blastocysts from which the ES cell line was isolated.
For teratoma formation, 1 × 106 Wnt1-CRE-Rosa
Tomato
ES cells in 50 μL of CM+/− (detailed composition in Supplementary Fig. S1; Supplementary Data are available online at
For embryonic epithelium tissue and differentiated Wnt1-CRE-Tomato Rosa ES cell re-association, heterogeneous populations of NC differentiated Wnt1-CRE-Tomato Rosa cells at day 15 of differentiation and embryonic day (ED) 12 oral/mandibular epithelium were formed on semi-solid medium and cultured during 12 or 14 days (38/57 surviving re-associations) as described [37]. Re-associations between ED12 oral epithelium and corresponding dental mesenchyme (25/44 surviving re-associations) or between ED14 dental epithelium and unsorted populations of Wnt1-CRE Tomato (11/25 surviving re-associations) were observed at day 15 of differentiation. Twelve re-associations were also performed as controls. The number of cells per pellet was approximately 1.86 ± 0.18 × 105 D15 differentiated Wnt1-CRE-Rosa Tomato cells for one ED12 oral epithelium or one ED14 dental epithelium as previously determined [38].
The outlines of the re-association protocol are presented in Fig. 4f. Re-associations were fixed for 24 h in Bouin-Hollande and embedded in paraffin. Serial sections (10 μm) were stained with either Mallory's staining [39] or hematoxylin/eosin. For detection of cartilage, re-associations were stained with alcian blue and safranin O. Sections were observed under a Leica DM4000B microscope.
All mouse procedures were approved by the Ethical Committee of Paris Descartes University (Agreement CEE nr: CEEA34.AP.031.02).
Derivation and culture of mouse Wnt1-CRE-Tomato Rosa ES cell lines
Super-ovulated B6. Cg-Gt(ROSA)26Sortm9(CAGtdTomato)/J females mated with Wnt1-CRE transgenic males were sacrificed, and late blastocyst-stage embryos were collected by flushing out isolated uteri by using M2 medium (Sigma Aldrich; M7167). Blastocysts were successively washed three times in drops of sterile M2 medium, TYRODE'S medium (Sigma Aldrich; T1788), M2 medium, and finally knockout serum replacement (KOSR) ES medium (Supplementary Table S1). The ES cell line was then generated as described [40]. Briefly, blastocysts were plated on mouse embryonic fibroblasts (MEFs) in KOSR ES medium to allow their attachment and inner cell mass outgrowth. On day 3 after plating when the blastocysts had attached to the feeders, half of the medium was replaced by fresh medium and cells were fed in this manner every other day until enzymatic disaggregation and cell plating to a new feeder covered plate.
At this step named Passage 1, outgrowths were moved from KOSR ES medium to ES fetal bovine serum (FBS) medium. Isolated cell clones (16) were tested for the presence of the Wnt1-CRE gene, and three positive cell clones were chosen and expanded as described [41]. The composition of the culture media used is described in Supplementary Table S1. One clone that displayed all the typical characteristics of an ES cell line was selected for further analysis (NC kinetics of differentiation, cell sorting and differentiation, ex vivo and in vivo interactions). This is the ES 1.1 cell line.
ES cell differentiation toward the three germ layers
Spontaneous differentiation was induced with the formation of EBs by using the hanging drop method [42]. Briefly, drops of 22 μL containing 1.2 × 103 Wnt1-CRE-Rosa Tomato cells were deposited on lids of bacterial Petri dishes for 5 days. For mesoderm differentiation, EBs were formed and cultured in CM+/− medium supplemented with 50 ng/mL vascular endothelial growth factor (VEGF; Peprotech, Neuilly sur Seine, France; 450-32) and 100 ng/mL of basic FGF2 (Peprotech; 100-18B). At day 5, EBs were placed onto a 3D Type I collagen gel (Corning 354249) in CM+/− supplemented with 50 ng/mL of VEGF, 100 ng/mL FGF2 (Peprotech; 100-18B), erythropoietin at 2 U/mL (Roche; 11 120 166 001), and interleukin-6 at 10 ng/mL (Peprotech; 216-16). The next day, 1 mL of the same medium was added and then renewed at 50% daily until day 14.
For endoderm differentiation, ES cells were cultured in hanging drops in CM+/− medium. At day 5, EBs were collected from the lid of the bacterial dishes and plated on 0.1% gelatin in P24 wells. EBs were then cultured in CM+/− medium until day 14.
For ectoderm differentiation, EBs were cultured in hanging drop in CM+/− medium supplemented with 5 μM retinoic acid [43]. At day 5, EBs were transferred in neurobasal medium supplemented with B27 (without vitamin A; Invitrogen-Gibco; Fisher Scientific France, Illkirch-Graffenstaden, France; 12587-010) and 5 mM retinoic acid (N2/B27/RA medium) on poly-
ES cell induction toward an NC phenotype
Wnt1-CRE-Tomato Rosa ES cells were inoculated on 0.1% gelatin-coated plates (2 × 104 cells/cm2) in CM+/+ medium (detailed composition in Supplementary Table S1) and induced to differentiate as previously described [36].
In brief, the next day [considered as differentiation day 0 (D0)], the medium was switched to a completely defined differentiation medium (DDM medium, see Supplementary Table S1) containing 1× ITS 100× (Gibco; Fisher Scientific France; 51500-056), FGF2 (10 ng/mL) (Peprotech; 450-33), and 100 ng/mL heparan sulfate (Sigma Aldrich; 51541). From D4, BMP4 (10 ng/mL) (Peprotech; 315-27) was added and the medium was subsequently renewed every other day until D10. To reach differentiation D15, D10 cells were trypsinized (TVP1X, Supplementary Table S1), plated on laminin-coated plates at a density of 4 × 105cell/cm2, and cultured in the DDM medium as earlier. The time course of NC induction is summarized in Supplementary Fig. S1.
NC cell sorting and in vitro differentiation
Flow cytometric analysis was performed at the Centre d'Imagerie Cellulaire et de Cytométrie, Centre de Recherche des Cordeliers, UMR S 1138, Paris, France, with an FACS ARIA III
For pericytic differentiation, cells were plated on gelatin-coated slides at the density of 5 × 103 cells/cm2 and cultured in Endothelial Cell Growth Medium EGM™ BulletKit™ (Lonza, Levallois-Perret, France; CC-5036) supplemented as recommended by the manufacturer with the corresponding EGM Plus SingleQuots solutions (Lonza; CC-4542), that is, FBS, epidermal growth factor, hydrocortisone, insulin-like growth factor, ascorbic acid (AA), and 50 U/mL penicillin/streptomycin (Invitrogen; Fisher Scientific France; 151070-063) as described [45]. The differentiation was carried out for 8 days.
For osteoblastic differentiation, cells were plated on gelatin-coated slides at the density of 2 × 104 cells/cm2 and cultured in alpha modified Eagle medium (αMEM) (Gibco; Fisher Scientific France) with 10% FBS, 7 mM β-glycerophosphate (CalBiochem; Fisher Scientific France), 0.2 mM AA, 100 mM dexamethasone, and 50 U/mL penicillin/streptomycin [46]. The differentiation was carried out for 21 days.
For neuronal differentiation, cells were plated on poly-
Immunofluorescence and immunohistochemistry
Cells were fixed in paraformaldehyde (PFA) 4% for 15 min, washed with PBS, permeabilized in 0.2% Triton X-100 in PBS (Sigma Aldrich; T-8787) for 15 min, and finally blocked with a solution of 1% BSA in PBS for 2 h. Coverslips were incubated overnight at 4°C with primary antibodies in PBS plus 0.2% BSA. After washing, coverslips were incubated for 1 h at 20°C with secondary antibodies diluted in PBS with 0.2% BSA and were mounted in DAKO mounting medium (S3023; Dako, Trappes, France). Negative controls were performed with secondary antibodies only (not shown) and isotype mouse or rabbit serum (shown in Supplementary Fig. S2). All antibodies are listed in Supplementary Table S2. Images were captured with a confocal laser-scanning microscope SP5 (Leica Microsystemes SAS, Nanterre, France).
Genotyping
Genotyping of the Wnt1-CRE mice was done by PCR using tail gDNA. DNA extraction was made with the EZ-10 Spin Genomics DNA Miniprep kit (Euromedex, Souffelweyersheim, France), according to the manufacturer's recommendation. The following primers were used for Wnt1-CRE: FOR 5′-GTCCGGGCTGCCACGACCAA-3′ and REV 5′-ACGGAAATCCATCGCTCGACCAGTT-3′ generating a 350 bp amplicon. RosaTomato (mT/mT) mice B6 Cg-GT(ROSA)26Sor tm9(CAG-tdTomato)Hze/J (Jackson laboratory) were genotyped as recommended by the provider. One standard PCR reaction was performed with the Kit TaKaRa ExTaq Polymerase (Chemicon; Fisher Scientific France; RR001A).
Quantitative PCR analysis
Samples were collected at various time points of the differentiation process. Total RNA extraction was performed with the RNeasy Mini Kit (Qiagen S.A.S., Courtaboeuf, France; 74134). Synthesis of cDNAs was made with the Verso Thermo Scientific kit (AB14531B) according to the manufacturer's recommendations.
Quantitative PCR reactions were performed with SYBR Green Master Mix 2X kit in a thermocycler LightCycler480® (both Roche Diagnostics, Meylan, France). The specificity of amplification was confirmed by visualizing the PCR products on agarose gels and by analyzing the melting curve after the completion of 45 cycles. The values for each gene were normalized to expression levels of a reference pool of genes, including β-actin and succinate dehydrogenase complex subunit A flavoprotein (SDHA). Results were analyzed by the ΔΔCt method. Each PCR was performed in triplicate. A list of the primers used in this study is presented in Supplementary Table S3.
Cell transplantation in avian embryos
NC-derived Wnt1-CRE-Tomato Rosa ES cells at day 15 of differentiation were injected into the presumptive territory forming the first branchial arch in E2 chick embryos (HH12 stage, ie, 48 h of incubation). The dispersal and migration of Tomato+ cells were observed 24 h (n = 3) and 7 days (n = 3) after grafting, and their phenotypic differentiation was analyzed on 10 or 30 μm cryostat sections of embryos fixed in PFA 4% and embedded in Optimum Cutting Temperature compound. Visual observation of Tomato+ cells was performed with a Leica microscope (DM6000B).
Statistical analyses
The statistical analyses were conducted by using Prism 4 (GraphPad Software). Experiments were repeated at least three times. Depending on the number of samples and their distributions, one-way analysis of variance test was used to compare mean with standard error of the mean in the case of normal distributions. *P < 0.05, **P < 0.005, ***P < 0.001. n indicates the number of samples analyzed. For the cell sorting experiments, the statistical analyses were done with mean ± standard deviation.
Results
The Wnt1-CRE-RosaTomato ES 1.1 cells display the characteristics of an ES cell line
To identify and characterize mouse cranial NC progenitor cells and to follow their in vitro as well as in vivo differentiation potentials, we have generated a Wnt1-CRE-Rosa Tomato mouse ES cell line, the ES 1.1 line, from blastocysts resulting from a cross between Wnt1-CRE male [25] and R26-Rosa Tomato females. The Wnt1-CRE-Rosa Tomato line displayed a typical ES colony morphology (Fig. 1a) with no Tomato protein expression in the naive state (Fig. 1b). ES colonies displayed alkaline phosphatase activity (Fig. 1c) as well as an immunoreactivity toward the typical pluripotency markers NANOG, OCT4, SOX2, or SSEA-1 (Fig. 1d–g). They also presented a capacity to differentiate in vivo (Fig. 1h–m) as well as in vitro (Fig. 1n–t) toward derivatives of the three embryonic germ layers. Indeed, on implantation of 1 × 106 Wnt1-CRE-Rosa Tomato ES cells in a BALB/c nude mice mouse, teratoma tumors developed within 4 weeks. Sections at different levels of the tumor revealed, among others, the presence of neural rosette and glial tissue (Fig. 1h, i), cartilage, and mineralized structures (Fig. 1j, k), together with glandular tissue and ciliated cells (Fig. 1l, m), typical from the ectoderm, mesoderm, and endoderm layers, respectively. As expected, zones of red fluorescent cells could be detected in the tumors, demonstrating the functionality of the CRE system in our ES cell derivatives (not shown). In vitro, specific induction of the ES 1.1 cells toward a neuronal fate allowed revealing, after 12 days, the presence of numerous cells expressing neuron-specific class III β-tubulin (TUJ1) or glial fibrillary acidic protein (GFAP) (Fig. 1n, o respectively). Induction toward a vascular fate led to the formation of sprouting structures after 14 days (Fig. 1p), positive for alpha smooth muscle actin (αSMA) or CD31 (Fig. 1q, r). In addition, 14 days of spontaneous differentiation allowed observing extensive zones of cells expressing the early endodermal markers Sox17 (Fig. 1s), HNF1 (not shown) and the pancreatic marker FoxA2 (Fig. 1t). Altogether, these data, thus, demonstrated that the ES 1.1 cell line presents all the characteristics of a stable pluripotent ES cell line, able to be differentiated toward ectodermal, mesodermal, and endodermal cell fates.
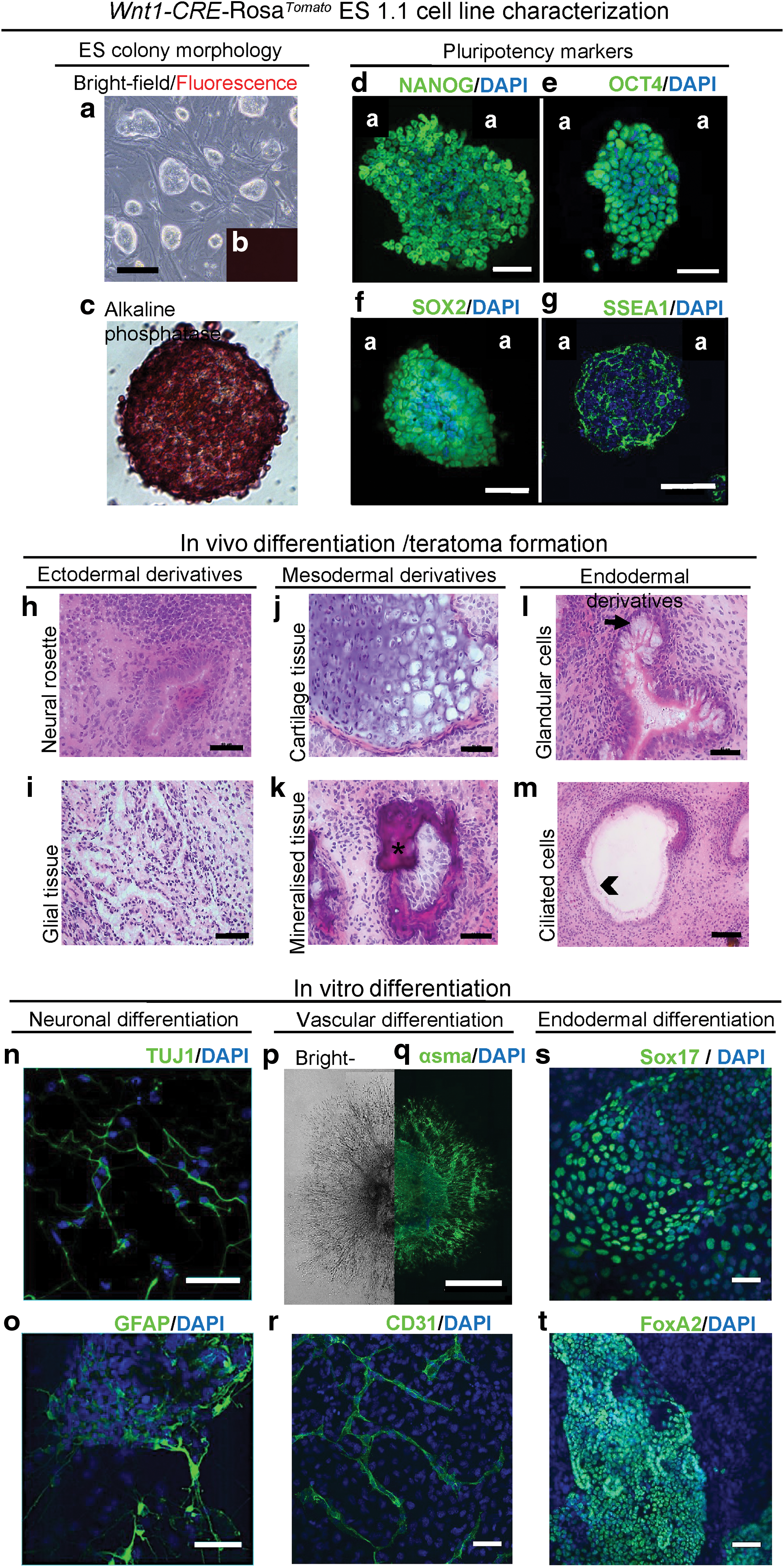
Wnt1-CRE-Rosa
Tomato
ES 1.1 cell line characterization
The Wnt1-CRE-RosaTomato ES 1.1 cells can be induced toward an NC phenotype
The Wnt1-CRE-Rosa tomato ES 1.1 cells were induced to differentiate toward an NC fate in the successive presence of FGF2 and BMP4 (Supplementary Fig. S1). Quantitative PCR analysis performed at various times of the differentiation kinetics showed a drastic decline in the pluripotency gene expression, Sox2, Nanog, and Oct4, shortly after differentiation induction [36]. This classically signals the ES cell loss of pluripotency and commitment.
The induction of typical NC specifier gene expression such as Snail, AP-2α, Twist, Pax3, FoxD3, Sox9, and Sox10 as well as Nestin was observed between day 6 and 10 of the differentiation (Fig. 2a) in parallel with the Tomato protein detection in a subset of cells (Fig. 2b). This reflected the acquisition of an NC identity by the ES 1.1 cells, thereafter referred to as ES-NCD cells, and demonstrated the functionality of the CRE recombinase system in our cells. An FACS analysis revealed the existence of three cell sub-populations in regard to the Tomato protein expression levels: Tomatohigh, Tomatolow, and Tomato- (Fig. 2c, d). These cell populations represented, on average (nine independent analyses), respectively, 39.2%, 10.2%, and 50.6% of the cell culture. Our protocol for NC differentiation was, therefore, efficient with half of the cell population expressing the Tomato protein (Fig. 2c) and very reproducible, as shown by the statistical analysis (Fig. 2d).

NC phenotype induction in Wnt1-CRE-RosaTomato ES 1.1 cells. In vitro, the Wnt1-CRE-Rosa
Tomato
mouse ES cells are induced toward an NC phenotype in a defined culture medium with FGF2 and BMP4. Real-time PCR analysis shows the activation of early NC specifier genes such as Snail and AP2-α, Twist and Pax3 starting at day 6 of differentiation. Later, NC typical genes, such as FoxD-3, Sox family members, and Nestin, become detected around day 10 of differentiation. Expression levels are normalized to β-actin and SDHA transcripts. The mRNA levels in differentiated cells are expressed relative to those of naive ES cells, which are taken as 1. The vertical scale bar represents standard deviation of three independent experiments after normalization. Statistical significant difference between samples, *P: 0.05; **P: 0.005; ***P: 0.0005. D, differentiation day
At the single-cell level, immunofluorescence performed on the total population of ES-NCD confirmed the expression of AP-2α and Snail proteins in the Tomato-positive ES-NCD cells (Fig. 2e, f). However, although the majority of the Tomato+ cells expressed Snail, AP2α expression was restricted to a subpopulation of the ES-NCD Tomato+ cells. This heterogeneity, also reflected in the FACS analysis, could reveal the presence of cells with different degrees of differentiation/maturation: immature/Tomatolow and more differentiated/Tomatohigh ES-NCD cells.
Day 10 ES-CND Tomato cells are able to differentiate into typical NC-derived cell types
NC cells are known to be multipotent and able to differentiate toward a variety of cell types such as osteoblasts, chondrocytes, adipocytes, neurons, melanocytes, pericytes, or odontoblasts. To evaluate whether the day 10 ES-NCD Tomato+ cell population presented the typical multipotent properties of the NC, the sorted Tomato+ (Tomato high subpopulation) population was induced in vitro toward different classical NC-derived cell types.
Sorted Tomato+ cells were first induced toward an osteoblast-like phenotype in a mineralization medium for 21 days. At this time point, mineralization nodules could be observed in the culture (Fig. 3a, b). They could be stained by Alizarin Red (Fig. 3c, d), in contrast to control cells cultured in normal medium (not shown). Further, immunofluorescence analyses revealed that the sorted Tomato+ cells expressed bone sialoprotein and osteopontin (Fig. 3e, f, respectively), two proteins classically associated with the osteoblast phenotype.
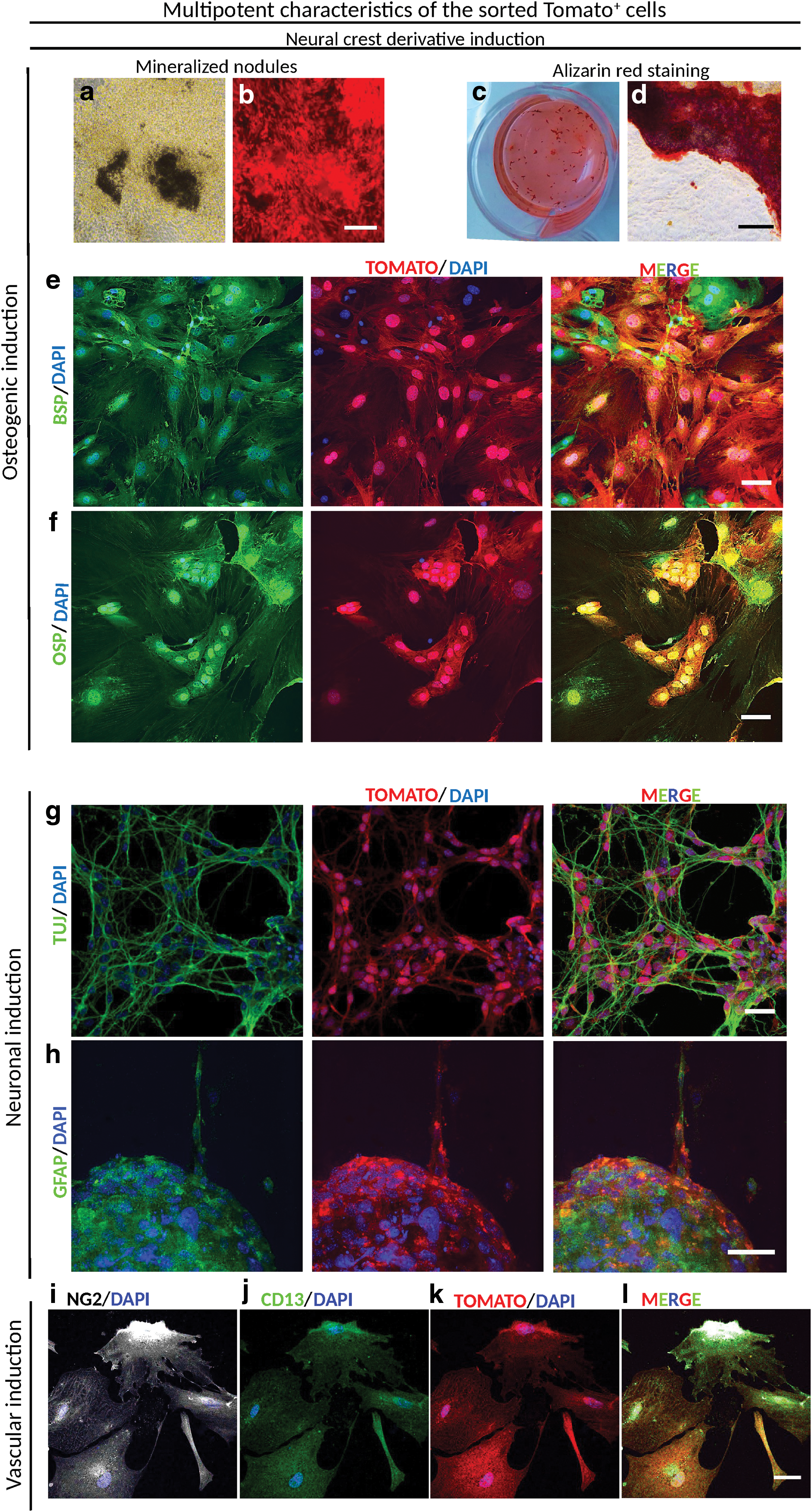
Multipotent characteristics of the sorted Tomato+ cells. Day 10 ES-NCD cells sorted on Tomato expression were induced in vitro toward different NC derivative programs. After 21 days of differentiation in osteogenic conditions
In parallel, the sorted Tomato+ cells were induced for 14 days toward a neuron- or a glial-like cell phenotype. In these conditions, cells displayed a typical morphology of neural-like cells and became stained for markers of glial and neuronal cells, GFAP and TUJ1 respectively (Fig. 3g, h).
Finally, culture of the sorted Tomato+ cells for 8 days in the presence of a pericytic differentiation medium allowed them to be directed toward a pericytic-like phenotype as shown by their immunoreactivity for NG2 and CD13 (Fig. 3i, j and l) co-localized with the Tomato protein (Fig. 3k, l).
Taken together, these results indicated that the induced ES-NCD Tomato+ cells could be committed in vitro toward classical NC-derived cell types. They, thus, behaved as a multipotent population as expected for NCSC cells.
Day 15 ES-NCD Tomato cells are able to interplay with embryonic tissues ex vivo and in vivo
In view of getting more homogenous and potentially more mature ES-NC cells, the D10 ES-NCD cells were re-plated on laminin-coated plates and their specification was pursued for five more days. At D15, the ES-NCD cell population still presented a typical ectomesenchymal morphology (Fig. 4a). FACS analysis showed two clear peaks of Tomato− and Tomato+ cells, and the percentage of Tomato+ cells in the total population reached 58% (Fig. 4b). Accordingly, the NC markers Snail, Twist, Ap-2α, Sox9, Sox10, and Nestin were significantly upregulated compared with the D10 cell population (Fig. 4c); whereas Pax3 and FoxD3 were downregulated (not shown).
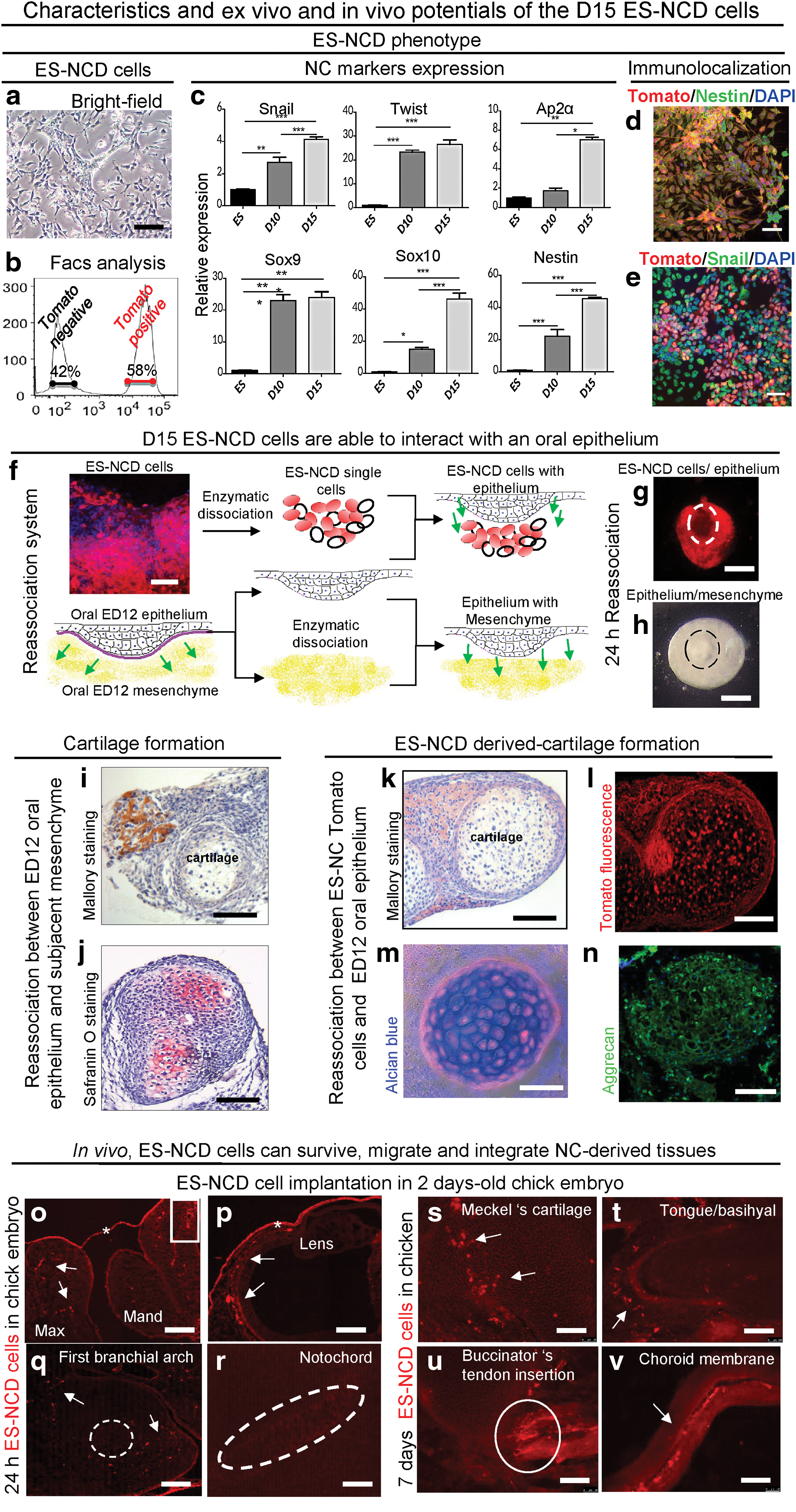
Phenotype characteristics and ex vivo and in vivo potentials of the D15 ES-NCD cells. ES-NCD phenotype: Ectomesenchymal morphology of D15 ES-NCD cells
At the single-cell level, immunofluorescence analysis performed on unsorted cells revealed that a majority of Tomato+ cells expressed Nestin, a classical hallmark of NC cells, whereas the early NC marker Snail was expressed by a subset of cells (Fig. 4d, e, respectively and Supplementary Fig. S3). This suggested that the cells had, indeed, progressed in the differentiation process between D10 and 15.
To further determine whether the ES-NCD cells could be used for cell/cell or cell/tissue interaction analysis and cell tracking in embryonic tissues, we first used a classical model of re-association largely exploited for the study of epithelium–mesenchymal interactions during development [6,38,47]. Mouse oral epithelium, between ED10 and 12, is known to be instructive toward the underlining ectomesenchyme and to be involved in the specification of odontogenesis and formation of Meckel's cartilage [22,48,49].
In contrast to D10 cells (Supplementary Fig. S4), D15 ES-NCD cells were able to form stable re-associations with a D12 oral epithelium (Schematic representation Fig. 4f, g), similar to its D12 ectomesenchymal cell counterparts (Fig. 4h). After a 12-day in vitro culture, cartilaginous-like structures formed in 82% of the ES-NCD/epithelium re-associations as revealed by Mallory, alcian blue staining, and aggrecan immunofluorescence (Fig. 4k, m, and n, respectively). In these structures, Tomato+ cells (Fig. 4l) systematically co-localized with the alcian blue staining (Fig. 4m). This demonstrated a direct participation of D15 ES-NCD cells in cartilage formation. Similarly, control re-associations between ED12 oral epithelium and its subjacent mesenchyme presented cartilaginous structures in 76% of the cases as revealed after staining with Mallory (Fig. 4i), safranin O (Fig. 4j), or alcian blue (not shown).
In contrast, ED12 ectomesenchyme alone or D15 ES-NCD cells cultured alone or after re-association with a non-signaling dental epithelium did not generate any recognizable differentiated structure (not shown). Taken together, these results showed that D15 ES-NCD cells were capable of responding to the signaling action of an embryonic epithelium in a similar manner to ED12 ectomesenchymal cells.
To also determine whether the ES-NCD cells were able to survive, migrate, and integrate within a physiological embryonic environment, advantage was taken from the chicken embryo, a well-characterized model in which mammalian cells have been shown to survive and differentiate in response to avian environmental signals [50].
Cells were implanted near the maxilla-mandibular commissure in the first branchial arch of ED2 chick embryos. Forty-eight hours after grafting, Tomato+ cells were revealed within the first branchial arch, the mandibular and maxillar processes, and the choroid membrane of the eye but were notably absent from the mesoderm-derived notochord and the mesenchymal nucleus of the first branchial arch (Fig. 4o–r, respectively), substantiating the ectomesenchymal tropism of the ES-NCD. Seven days after implantation, they were observed within cranio-facial structures such as Meckel's cartilage, basihyal bone, and insertion site of the buccinators tendon along with perivascular derivatives as evidenced in the choroid membrane (Fig. 4s–v, respectively).
These observations showed that the Tomato+ cells were able to survive and migrate in the chick embryonic environment. Besides, their patterns of integration were consistent with the potentials displayed by the ES-NCD cells in our in vitro differentiation assays. Altogether the data of re-associations and in vivo implantation demonstrated that the ES-NCD cells were responsive to ex vivo and in vivo embryonic tissue signaling.
Discussion
We have described the isolation of a new ES line, the Wnt1-CRE-Rosa Tomato ES 1.1 line, derived from blastocysts of a cross between Wnt1-CRE [25] and R26 Rosa Tomato mice. We revealed that this cell line: (1) displayed all the characteristics expected for an ES cell line, (2) could be efficiently committed in vitro toward an NC-like phenotype, through a straightforward protocol and in 10 days, (3) behaved at this stage as multipotent NC cells that can be further specified toward classical NC-derived cell types as well as ectomesenchymal cells, and (4) could respond to signals from an embryonic epithelium tissue as well as survive and get integrated within craniofacial tissues of the chick embryo.
Over the past decades, the development of the stem cell research field has been exponential and the hope of using stem cells for developing innovative cell or tissue-engineering strategies has been growing in all fields, with some already in the first phase of clinical trials [51 –53]. However, if the potentials of various stem cell types, such as MSCs or ES cells, in tissue engineering/therapeutic approaches have been demonstrated in animal models, their transfer to clinical applications remains limited. This is explained, in part, by the lack of knowledge on the in vivo behavior of these cells. For instance, the survival as the direct participation of implanted cells in the tissue reconstruction process is still a matter of debate and appears to directly depend on the type of implanted cells and the microenvironment of host tissues [54].
To get insight into this matter, development of new tools is required. Lineage-specific ES cell reporter lines are important in this context [23,24] since they can facilitate the real-time monitoring of differentiation in vitro, the purification of specific progenitor subpopulations, and the longitudinal analysis of their survival and behavior in implanted tissues.
Our ES 1.1 cell line specifically expressing a fluorescent protein in NC cells and their derivatives was generated from blastocysts derived from a cross between the Wnt1-CRE mice and Rosa Tomato mice. TheWnt1-CRE mice have been extensively used for NC lineage tracing and NC-specific gene inactivation (see, for instance, [26]). In mice carrying both the Wnt1-CRE transgene [25] and R26RlacZ reporter allele [55], the lacZ gene was shown to be expressed in cranial NC derivatives in addition to the region where Wnt1 is usually expressed [56]. The transgene expression is exclusive to the cranial NC mesenchymal lineages [26], hence providing a distinct feature and sharp contrast to the mesoderm-mesenchymal derivatives that never activate Wnt1-CRE transgene activity.
In their naive state, the newly isolated Wnt1-CRE-Rosa Tomato ES line displayed the typical characteristics of pluripotent ES cells. As expected, they were able, on in vivo implantation in the mouse, to form typical teratoma, therefore containing various tissue types derived from the three germs layers among which NC derivatives as suggested by the presence of red fluorescent cells were observed in the tumors. Likewise, under appropriate in vitro induction, they could be committed toward a mesodermal, endodermal, or neuroectodermal fate. Altogether, they behaved as a true ES cell line.
In view of their use for following the fate of NCSC in tissue-engineering repair models, Wnt1-CRE-Rosa Tomato ES cells were first induced toward an NC phenotype through a two-step protocol [36]. NC development is perceived as a step-wise progression from inductive events to transcription factor expression and modulation of migration and differentiation. FGF and BMP together with Wnt and Shh signaling have been shown to be involved in NC specification in vitro as well as in vivo [16,18,20,57,58]. Studies in different model systems have led to observing that NC cells are specified in the region where neural-promoting FGF signals and epidermal-promoting BMP activity overlap to establish a balance of these two signals [58]. Both these signaling factors were used in our protocol [36].
On such a treatment, ES Wnt1-CRE-Rosa Tomato cells changed their morphology and started expressing the classical early neural NC specifier genes between day 6 and 10, in parallel with the Tomato fluorescent protein. This observation demonstrated the functionality of the CRE recombinase system in our conditions. Differentiation was efficient: At D10, the expression of all the classical NC specifier genes was activated and more than 40% of the cells were Tomato+. The presence of both a “low” and a “high” Tomato-positive cell subpopulation suggested that a maturation process is involved and that some cells had not yet reached the full NC fate. This hypothesis was supported by the observation that, on further specification (D15), only one peak of Tomatohigh cells was detected.
This population expressed the set of markers, characteristic of NC phenotype. At the single-cell level, most of the D10 ES-CN Tomato+ cells displayed expression of Snail, whereas Ap-2α expression was more restricted. Both genes are typical early NC specifiers, involved in epithelial-mesenchymal transition [21], with AP-2α often associated with the migratory NC phenotype. It seemed reasonable to hypothesize at this point that Ap2α+ and − cell populations corresponded to mixed migratory and non-migratory NC cells. Alternatively, this heterogeneity could underline different stages of specification [59].
Cell-sorted D10 ES-NCD Tomato+ cells behaved as a multipotent population as expected for NC stem cells [7]. They did not spontaneously differentiate and required specific induction for engaging into an osteogenic, neuronal, glial, or vascular program with kinetics similar to what had been described for NCSC [28,46]. Future studies will have to address the stem cell nature of the sorted Tomato+ population and its self-renewal ability as its capacity to maintain a multipotentiality in defined culture conditions [28,60].
Altogether, these data show that the protocol used to generate NC cells from the ES 1.1 cells is efficient and reproductive. Wnt1 expression, in addition to NC expression, has been described to also mark dorsal neural stem cells [56]. Although we cannot rule out a small contamination of non-NC Tomato+ cells here, we are focusing on a Tomato+ cell population, which, after differentiation and sorting, displays the characteristic multipotency of NCSC, which is capable, in particular, of efficient osteogenic differentiation.
The interaction of ES-NCD cells with other tissues was first explored by establishing classical in vitro tissue recombination with an embryonic oral ectoderm as described [61]. D10 ES-NCD unsorted cells were not able to establish a dialogue with the oral epithelium, leading to tissue formation and, consequently, rapidly degenerated, suggesting that, at this stage, the cells were still immature. A further 5-day differentiation allowed acquisition of this property. Indeed, on re-association with an ED12 oral epithelium, not only did the D15 ES-NCD and their Tomato+ subpopulation survive but they also efficiently participated in the formation of cartilage nodules in which numerous Tomato+ cells were present.
Similarly, cartilage structures were observed on re-association of an ED12 oral epithelium with its subjacent ectomesenchyme. This cartilage formation appeared therefore as a consequence of the dialogue between the two tissues since no cartilage ever formed on culturing D15 ES-NCD alone or in re-association with an ED14 oral epithelium. This acquisition of a capacity of interaction with an oral epithelium was paralleled with a change in NC gene expression pattern with, for instance, expression stimulation of Snail, Twist, Ap-2α, Sox10, and Nestin and downregulation of FoxD3 and Pax3, two early NC characteristic genes whose expression is known to be downregulated on differentiation [7,62]. A further characterization of the D10 and D15 ES-NCD cells by RNA-Seq analysis should allow in the future a better understanding of the mechanisms underlying their maturation.
As native NC cells, D15 ES-NCD cells could integrate an in vivo environment, such as the chick embryo [28]. Indeed, they survived for several days after their graft in the developing chick, and integrated various tissues such as cartilage, bone, or blood vessels. Their pattern of integration was consistent with the fates they were able to display in our in vitro/ex vivo analysis and with published data from other laboratories obtained after in ovo implantation of NC-like stem cells [17,28,63,64]. Future studies should address the nature of the integrated Tomato+ cells as well as their fate and behavior at later time points during development.
In summary, we have isolated a new fluorescent ES-NC lineage-specific cell line and showed that it was capable of (1) efficiently acquiring, after induction, a typical NC phenotype together with expression of Tomato fluorescence, allowing specific cell tracking and sorting; (2) in vitro differentiating toward various NC-derived cell type phenotypes, thus behaving, as a whole, as an NCSC population; and (3) interacting with embryonic tissues and participating in the formation of new structures.
This cell line is, therefore, a new cell model that will enable studying the signals governing the NC fate acquisition and underlying differentiation to their specific subtype derivatives. Further, thanks to its fluorescent labeling, it should allow analyzing their interplays with the host tissue microenvironment after implantation and, consequently, pave the way for optimizing the conditions of efficient repair processes promoted by the use of NCSC or progenitors.
Footnotes
Acknowledgments
This work has been supported by grants from Institut Français d'Odontologie (IFRO), Université Paris Descartes, DIM (Ile de France) Biothérapies and the National French Agency for Research (grant ANR PulpCell 2014–2017), and FRM VisualSystem (SC laboratory). S.A.-M. has been supported by a doctoral fellowship from DIM Biothérapies. The authors thank Dr P. Opolon for help with analysis of the mouse teratoma and E. Devevre (CICC, CRC, Paris) for her help with the cell sorting analyses.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.