
Abstract
The enzyme aldehyde dehydrogenase (ALDH) is found in developing and multipotent cell populations, and is important for the production and regulation of retinoic acid, which controls β-cell differentiation in the pancreas. The role of ALDH-expressing cells in the formation of endocrine-like cells and co-localization with the putative stem cell marker CD133 has not been examined during human pancreatic development. This study focuses on the co-expression of CD133 on ALDH+ cells from the human fetal pancreas (18–22 weeks of fetal age) with transcription factors (TFs) central to endocrine cell development. Fluorescence-activated cell sorting demonstrated that cells with high ALDH activity (ALDHhi) had increased co-expression of CD133 and endocrine-lineage TFs when compared with cells with low ALDH (ALDHlo) expression. Hormone-expressing (insulin, somatostatin) and ductal cells (CK19) were noted in the ALDHhi population, while mesenchymal (vimentin) and endothelial (CD31) markers were predominantly found in ALDHlo cells. Culture of sorted ALDHhi or ALDHhi/CD133+ cells resulted in loss of endocrine TF, insulin, and CK19 expression. The formation of cell clusters from cultured ALDHhi or ALDHhi/CD133+ cells led to restored CK19 expression and showed endocrine TFs and insulin expression. In summary, pancreatic ALDHhi cells contain a heterogeneous CD133-enriched population with a subset of β-cell associated markers in the developing human pancreas.
Introduction
T
ALDH activity stimulates cellular differentiation by facilitating the conversion of retinaldehydes to retinoic acid (RA), a critical regulator of organogenesis during embryonic development [9 –11]. RA treatment also reestablishes differentiation in precursor populations after reversible ALDH inhibition through diethylaminobenzaldehyde (DEAB) [9,10,12]. During human pancreatic development, ALDH1A1+ cells correlate with neurogenin 3 (NGN3)- and insulin-expressing cells [10,13]. Thus, ALDH is a potential marker for examining and isolating cells that contribute to pancreatic endocrine cell development.
The presence of other reported adult stem cell markers have been identified within the developing and mature pancreas, and have been isolated using cell surface expression of CD34, CD117, and CD133 [14 –17]. Co-localization of these surface markers with ALDH activity has been used to analyze pancreatic cells with multilineage potential, as seen in ALDH+ adult murine centroacinar cells co-expressing CD133 [18]. Cells isolated from human bone marrow and umbilical cord blood demonstrated high ALDH activity and co-localization with other putative stem cell markers, and these cell populations promote effective regenerative properties in STZ-damaged islets [19]. Therefore, examining ALDH activity in conjunction with other stem cell markers in the developing human pancreas may lead to the identification of a population with the capacity to produce islet-associated cell types. We have previously characterized ALDH1 activity during different stages of human pancreatic development [10], yet no study to date has examined the co-expression of sorted ALDH+ cells with putative stem cell markers from the dissociated human fetal pancreas and their contribution to pancreatic endocrine cell differentiation. In this study, we hypothesized that cells with high ALDH activity (ALDHhi) and CD133 co-expression in the developing human pancreas represent a pancreatic progenitor cell population with the potential to express transcription factors (TFs) and hormones present in pancreatic endocrine cells.
Materials and Methods
Preparation and dissociation of human fetal pancreata
Human fetal pancreata between 18 and 22 weeks of fetal age were obtained and approved by the Health Human Sciences Research Ethics Board at Western University, in accordance with the Canadian Council on Health Sciences Research Involving Human Subjects guidelines [10]. Pancreata were immediately digested with dissociation buffer containing DNase I (Roche, Mississauga, ON, Canada) and collagenase V (1 mg/mL; Sigma, Oakville, ON, Canada) [10], and further treated with trypsin-ethylenediaminetetraacetic acid (Invitrogen, Burlington, ON, Canada) to prepare single cell suspension. Cell suspensions recovered in CMRL-1066 media supplemented with 10% fetal bovine serum (FBS; Invitrogen) until the time of cell sorting at the London Regional Flow Cytometry Facility, Robarts Research Institute (London, ON, Canada) [10].
Fluorescence-activated cell sorting
Cell preparations were suspended in Aldefluor buffer (StemCell Technologies, Inc., Vancouver, BC, Canada) at a density between 6 × 106 and 1 × 107 viable cells. Aldefluor reagent (StemCell Technologies, Inc.) and anti-human fluorophore-conjugated CD34, CD117, and CD133 antibodies (Supplementary Table S1; Supplementary Data are available online at
Anti-human fluorophore-conjugated CD235a antibody (Supplementary Table S1) and viability marker 7-actinomycin (7-AAD; BD Biosciences, Mississauga, ON, Canada) were used to remove erythrocytes and nonviable cells from the final sorted populations, respectively. Cells were separated using multiparametric cell sorting on the FACSAria III Cell Sorter (BD Biosciences, San Jose, CA), and paired into two sorting groups for our primary focus: (1) ALDHhi and ALDHlo populations; and (2) ALDHhi/CD133hi and ALDHlo/CD133lo populations. All analyses were conducted using FACSDiva software (BD Biosciences).
Expansion and differentiation culture of sorted cells
To determine the extracellular matrix (ECM) composition that best promoted the viability of sorted ALDHhi cells, matrix proteins of fibronectin, laminin, and collagen I and IV (BD Biosciences) were used as coating in 96-well plates at 5 μg/mL, with control wells coated using 1% BSA solution. ALDHhi cells were plated at a density of 1 × 104 cells per well in Dulbecco's modified Eagle's medium (DMEM)/F12 media supplemented with 10% FBS (Invitrogen). Cellular viability and metabolic activity was measured 24 h and 5 days after plating using a MTT assay (3-[4,5-dimethyl-2-thiazolyl]-2,5-diphenyl-2H-tetrazolium bromide; Sigma). MTT+ cells were imaged by selecting five consistent fields in each well and quantified. Data were normalized to the total number of adhered cells per experimental group (three replications per experimental group).
Based on our initial tests and our previously published reports [20,21], collagen I coating (5 μg/mL) was chosen for expansion of the sorted ALDHhi, ALDHlo, ALDHhi/CD133hi, and ALDHlo/CD133lo cell groups in culture. Cells were cultured with DMEM/F12 + 10% FBS media. Once wells were 90% confluent, expanded cells were passaged and harvested for analysis. For differentiation cultures, expanded cells were seeded in 12-well plates and cultured with serum-free differentiation media [22 –24]. After 7 days in differentiation media, clusters were collected for RNA extraction and histological analysis.
Immunofluorescence analyses of cell populations
Sorted, cultured, and cluster-forming cell pellets were fixed in 4% paraformaldehyde and suspended in 2% agarose gel before embedding in paraffin [10]. Sections were cut at 4 μm and stained with the appropriate dilutions of primary and secondary antibodies (Supplementary Table S2). Analysis and quantification was conducted using the Image ProPlus software (Media Cybernetics, Inc., Rockville, MD) by counting positive-staining cells over total 4′,6-diamidino-2-phenylindole (DAPI)-labeled cells.
Quantitative reverse transcription polymerase chain reaction (qRT-PCR) analyses of cell populations
RNA from sorted, cultured, and cluster-forming cells was extracted using the RNAqueous-4PCR kit (Invitrogen) [10]. SuperScript II Reverse Transcriptase (Invitrogen) was used alongside isolated RNA for the generation of complementary DNA (cDNA). Real-time polymerase chain reactions (PCRs) were performed using the iQ SYBR Green Supermix kit (Bio-Rad Laboratories Canada Ltd., Mississauga, ON, Canada) and designed primer sequences (Supplementary Table S3). Data were analyzed using the Bio-Rad CFX Connect real-time PCR detection system (Bio-Rad Laboratories Canada Ltd.). Relative gene expression was quantified using the comparative CT method (2−ΔΔCT) and normalized to the internal control 18S rRNA [10,25].
Statistical analyses
Data are expressed as mean ± standard error of the mean, and statistical analyses using paired or unpaired Student's t-tests were conducted using GraphPad Prism program (Version 5.0c; GraphPad Software, Inc., La Jolla, CA). Data were considered statistically significant at P < 0.05.
Results
Sorted ALDHhi cells highly co-expressed CD133 in the developing human pancreas
ALDHhi cells from dissociated pancreata at 18–22 weeks of fetal age were isolated after gate establishment using a DEAB control (Fig. 1A) and comprised the majority (>60%) of viable cells (7-AAD−) retrieved (P < 0.001; Fig. 1B). Examination of stem cell–associated surface markers showed CD133 was prominently co-expressed in the ALDHhi population (66% ± 5%) compared with the ALDHlo sorted cells (8% ± 3%) (P < 0.001; Fig. 1C, F). In contrast, the majority of CD34+ cells were observed in ALDHlo sorted cells (42% ± 3%) compared to the ALDHhi cell population (24% ± 3%; P < 0.001; Fig. 1D, F). A low frequency of CD117+ cells was noted in both the ALDHhi (13% ± 8%) and ALDHlo (11% ± 4%) sorted groups (Fig. 1E, F).
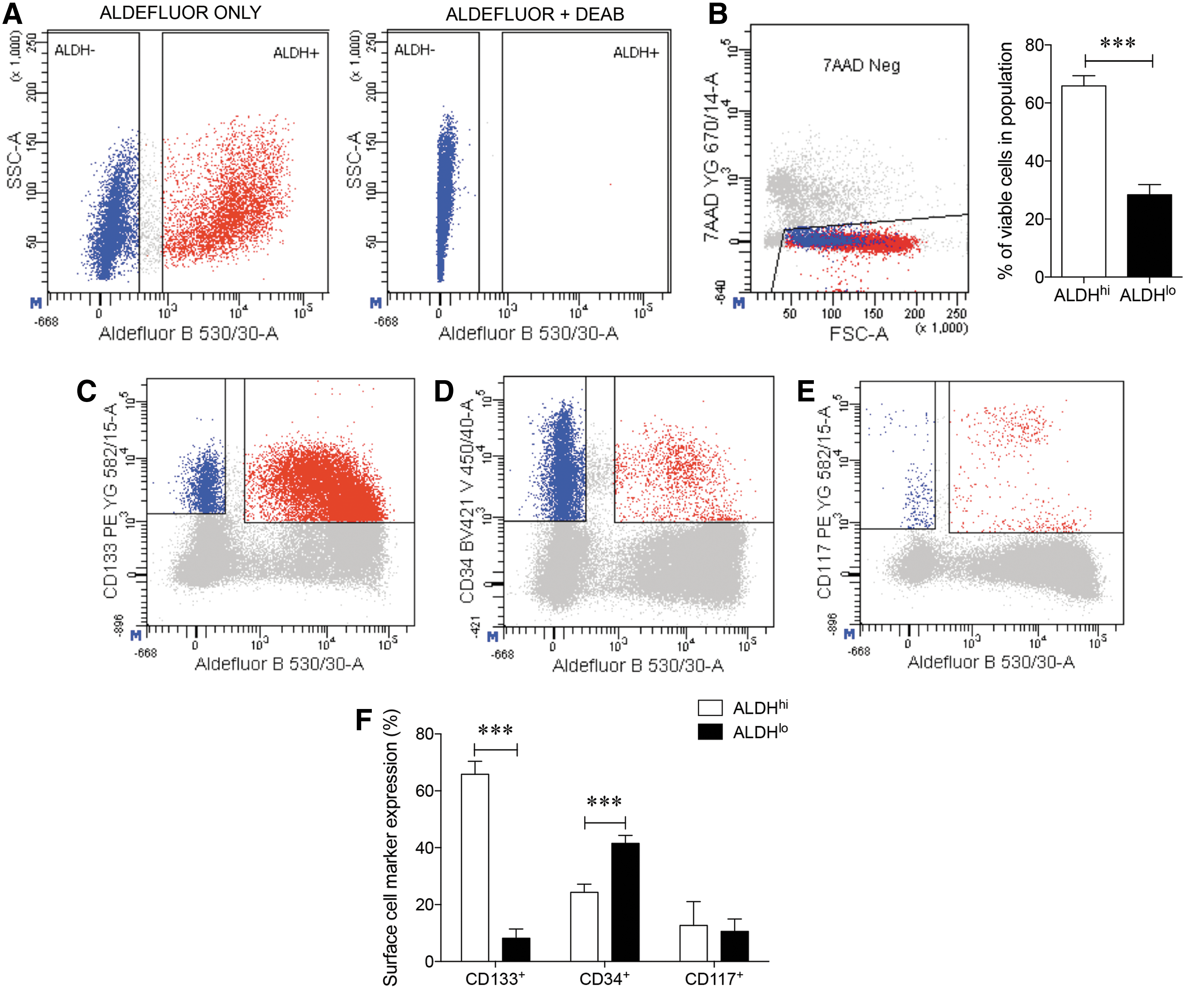
Analysis of ALDH activity, cell viability, and surface stem cell markers in the human fetal pancreas.
Using multiparametric analyses, overlapping co-localization for these surface markers in the ALDHhi and ALDHlo sorted cells was further examined. Co-expression of CD133+ cells with CD34 (Fig. 2A), CD117 (Fig. 2B), or CD34+ cells with CD117 (Fig. 2C) in both ALDHhi and ALDHlo sorted groups were observed at low levels (4%–10%). Thus, CD133 expression highly correlated with high ALDH activity, but not with CD34 or CD117 co-expression.
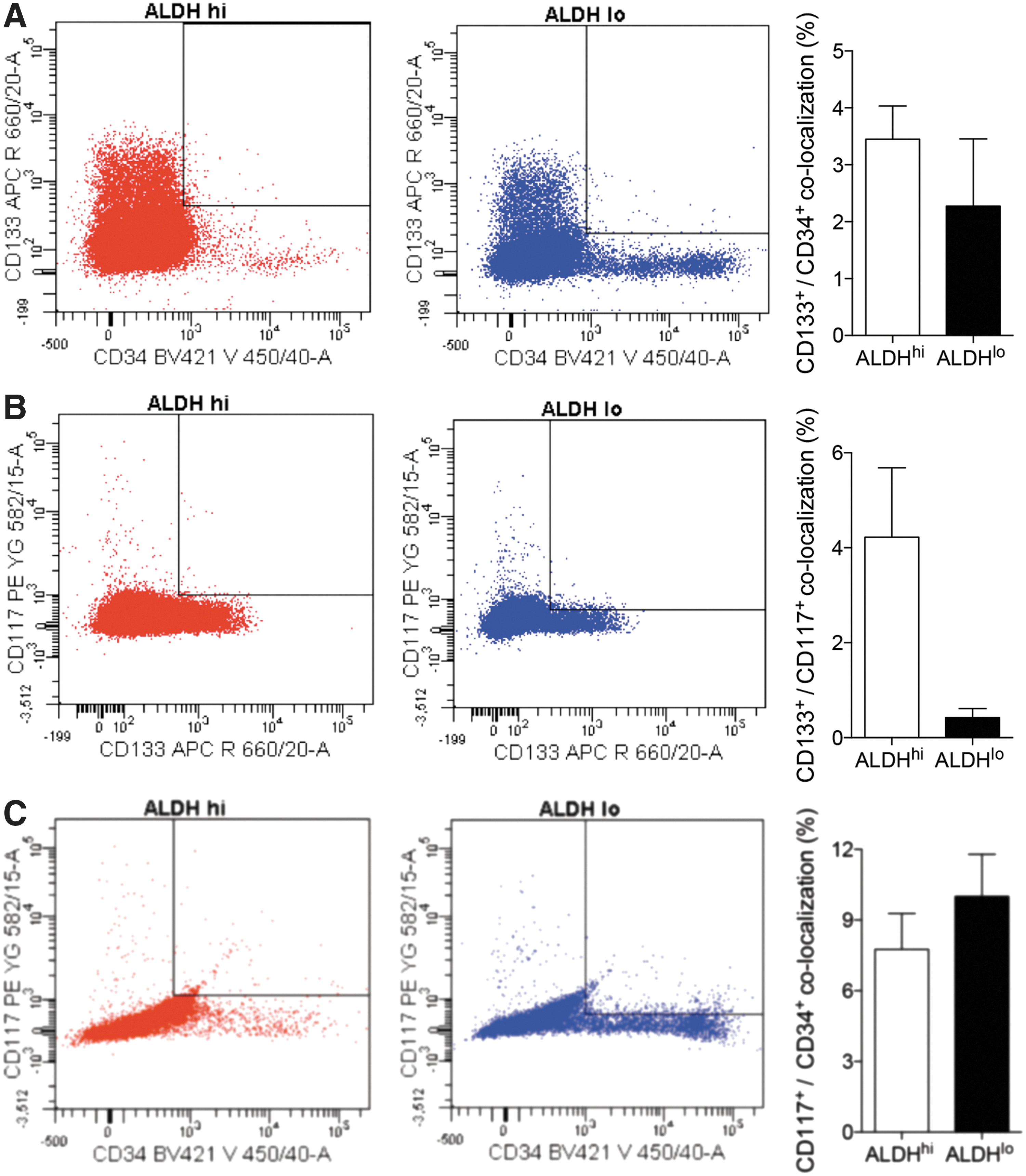
Co-localization between cell surface markers CD133, CD34, and CD117 in ALDHhi and ALDHlo subpopulations. Sorting gates (left) and quantification (right), in separated ALDHhi (red) and ALDHlo (blue) populations, of cells that co-localize markers CD133+/CD34+
Sorted ALDHhi cells showed higher expression of endocrine TFs
In addition to high CD133 co-localization, sorted ALDHhi pancreatic cells also demonstrated higher expression of endocrine cell-associated TFs (Fig. 3). Increased expression of SOX9, PDX-1, NGN3, and NKX6.1 mRNA was observed in ALDHhi cells, but statistical significance was not reached due to high variability between sample pancreata collected (Fig. 3A). To verify the nuclear localization of these TFs, immunohistochemical analyses were performed on sorted cells (Fig. 3B).
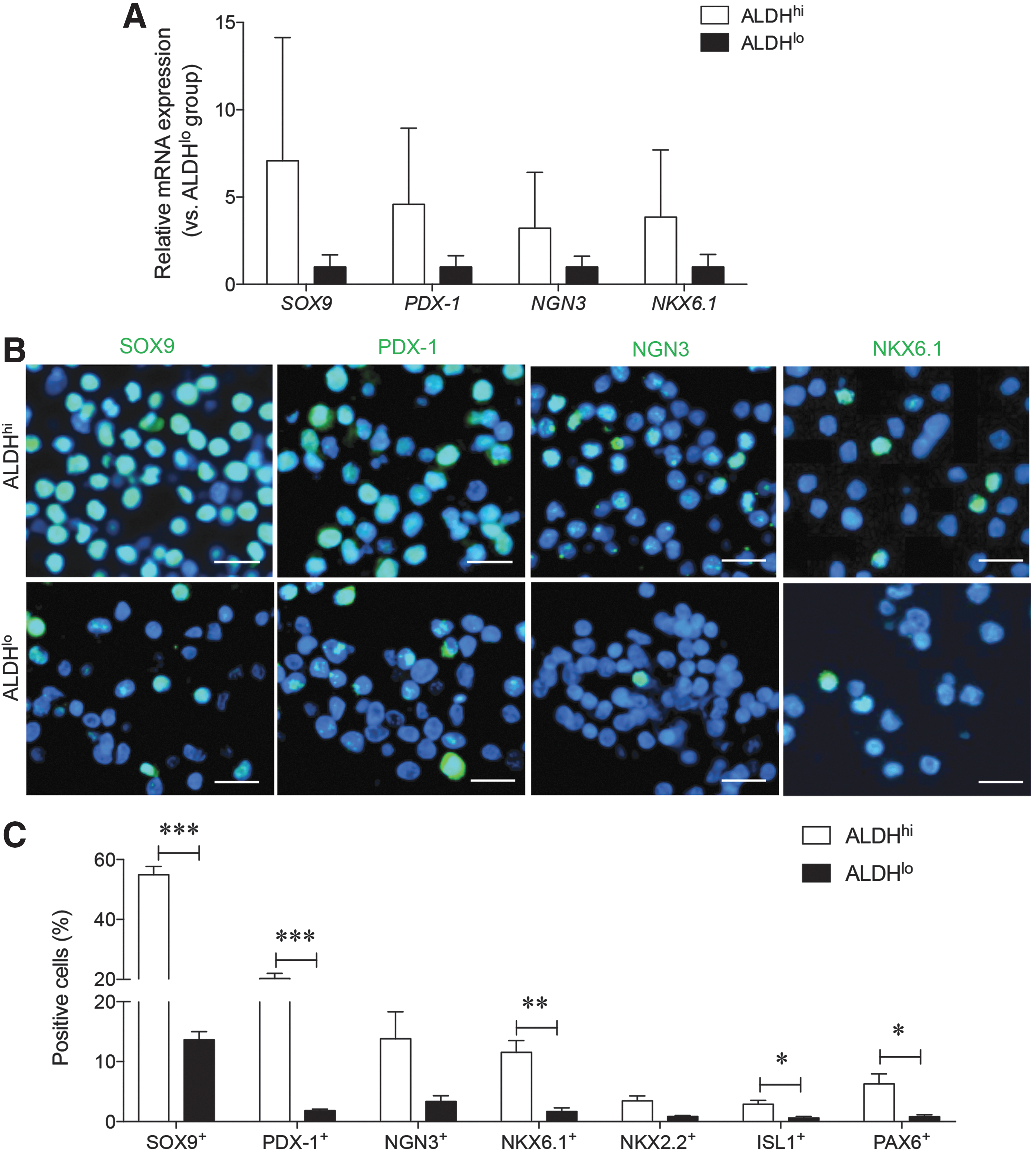
Pancreatic endocrine-committing TF expression in ALDHhi and ALDHlo sorted populations of the human fetal pancreas.
Sex determining region Y-box 9 (SOX9) expression was found in the nucleus of approximately half (55%) of sorted ALDHhi cells, while only 10% of ALDHlo cells contained nuclear SOX9 staining (P < 0.001 vs. ALDHlo; Fig. 3C). In addition nuclear pancreatic and duodenal homeobox 1 (PDX-1) and NK6 homeobox 1 (NKX6.1) expression were both significantly higher in the ALDHhi sorted cells (Fig. 3C). A high percentage of neurogenin 3+ (NGN3) cells were also noted in the ALDHhi group, but statistical significance was not detected (Fig. 3C). Finally, islet 1 (ISL1) and paired box gene 6 (PAX6) expression were also significantly increased in sorted ALDHhi cells compared with ALDHlo cells (P < 0.05; Fig. 3C). Similar to NGN3 expression, there was no significant difference in NK2 homeobox 2 (NKX2.2) expression between ALDHlo and ALDHhi cells. These data indicate that ALDHhi pancreata examined immediately after sorting contained a high percentage of cells expressing endocrine TFs necessary for islet cell differentiation.
ALDHhi cells represent a heterogeneous pancreatic population showing low proliferation
The presence of hormone+ cells and markers of nonendocrine pancreatic cell lineages were also examined in the sorted cells using immunofluorescence staining. A significantly higher frequency of insulin+ cells was observed in the ALDHhi population (5% ± 1%), while insulin+ cells were nearly undetectable in ALDHlo cells (0.8% ± 0.2%; P < 0.05 vs. ALDHhi; Fig. 4A). A high percentage of somatostatin+ cells were also present in the ALDHhi group (P < 0.05 vs. ALDHlo; Fig. 4A), yet both sorted populations contained a similar percentage of glucagon+ cells (Fig. 4A).
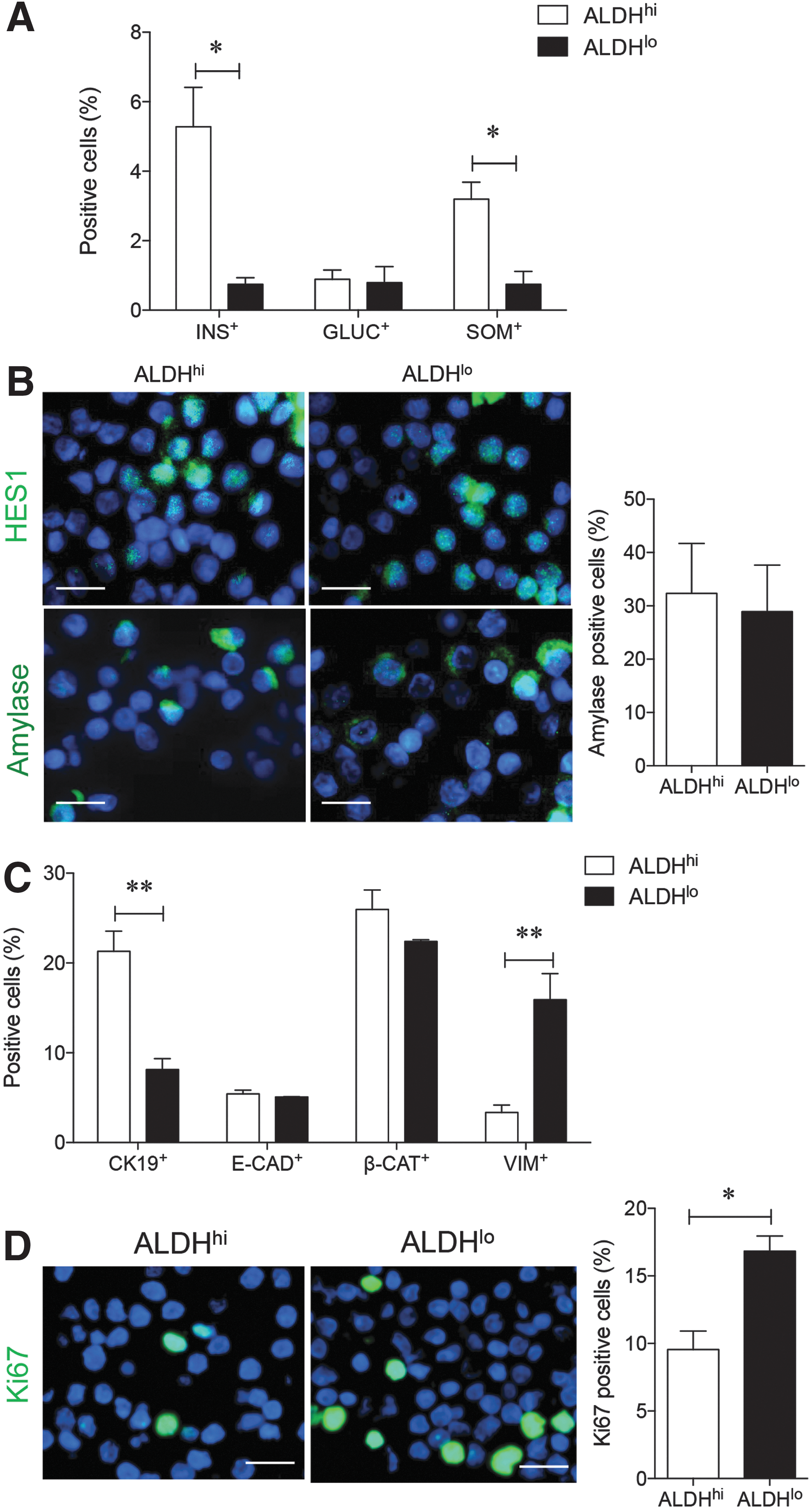
Characterization of cell phenotypes and cell proliferation in ALDHhi and ALDHlo sorted populations. Quantitative analysis for endocrine
Exocrine cell markers HES1 and amylase showed a similar distribution within both ALDHhi and ALDHlo groups (Fig. 4B). Expression of the pancreatic ductal epithelial cell marker CK19 correlated with the ALDHhi population, while no correlation was observed with E-cadherin and β-catenin expression (Fig. 4C). Sorted ALDHlo cells contained a high percentage of cells positive for the mesenchymal cell marker vimentin (P < 0.01 vs. ALDHhi; Fig. 4C). Furthermore, Ki67 detection in actively proliferating cells was higher in ALDHlo cells compared with the ALDHhi population (P < 0.05; Fig. 4D), indicating decreased cell cycle activity in the ALDHhi population at the time of selection.
Sorted ALDHhi cells lose expression of endocrine transcript factors during in vitro expansion
Due to the lower proliferative capacity identified in sorted ALDHhi cells, selected ECM proteins were used to promote ALDHhi cell viability and growth at 24 h and 5 days in expansion culture. At 24 h, viable adherent ALDHhi cells were scattered throughout the well in both control and ECM-coated wells (Fig. 5A). However, viable monolayers of expanded cells were significantly higher on laminin and collagen I-coated wells at day 5 (Fig. 5A). Both ALDHhi and ALDHlo cells displayed a spindle-shaped morphology during the expansion culture. Gene expression analysis, measured in the expanded ALDHhi and ALDHlo cells, showed decreased SOX9, PDX-1, NKX6.1, and INS mRNA in ALDHhi cells, yet levels of NGN3 appeared to be retained when compared with ALDHlo cells (Fig. 5B).
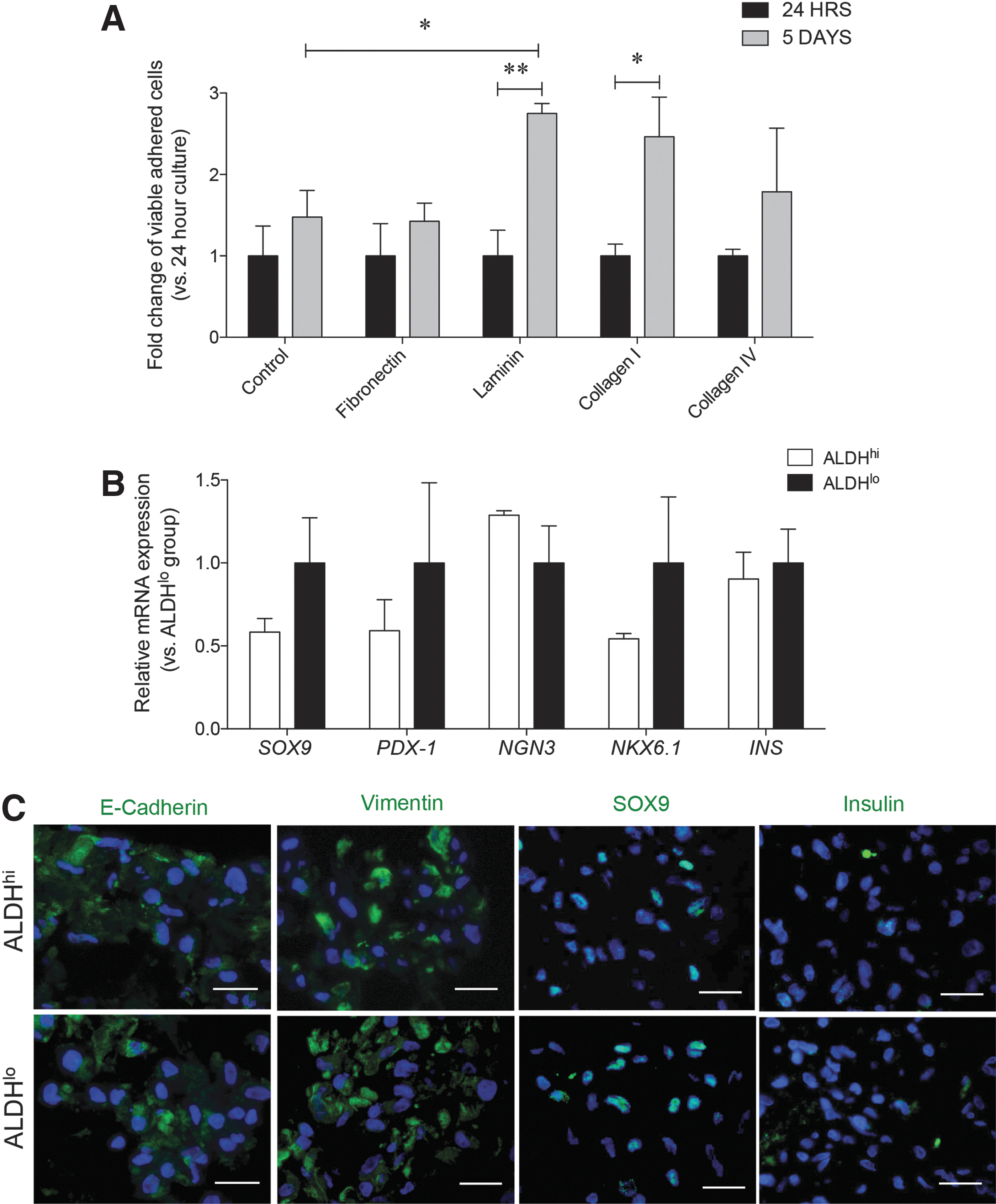
Analysis of cell phenotypes in ALDHhi and ALDHlo cells during expansion culture.
Immunostaining determined that expanded ALDHhi and ALDHlo cells positively labeled for both E-cadherin and vimentin (Fig. 5C), with loss of CK19 staining in both populations (images not shown). Further examination of nuclei staining for endocrine TFs revealed that SOX9 labeling was retained in ALDHhi and ALDHlo cells, but PDX-1+ and NGN3+ nuclear labeling was lost (images not shown) along with insulin expression after culture expansion of both cell population (Fig. 5C).
Cultured ALDHhi cells restore ductal phenotype during the formation of cell clusters
Treatment of cultured ALDHhi and ALDHlo populations with a modified differentiation media led to the formation of cell clusters within 36 h of administration (Fig. 6A). Quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis of TF expression showed equivalent expression of PDX-1, NGN3, and INS mRNA between differentiated ALDHhi and ALDHlo clusters (Fig. 6B). Immunofluorescence staining of fixed clusters showed the continued expression of vimentin and restoration of CK19 detection (Fig. 6C). Similar to cells expanded under nondifferentiated conditions, nuclear SOX9 staining was maintained in ALDHhi clusters. However, clusters derived from ALDHhi and ALDHlo cell populations showed weak staining for insulin (Fig. 6C).
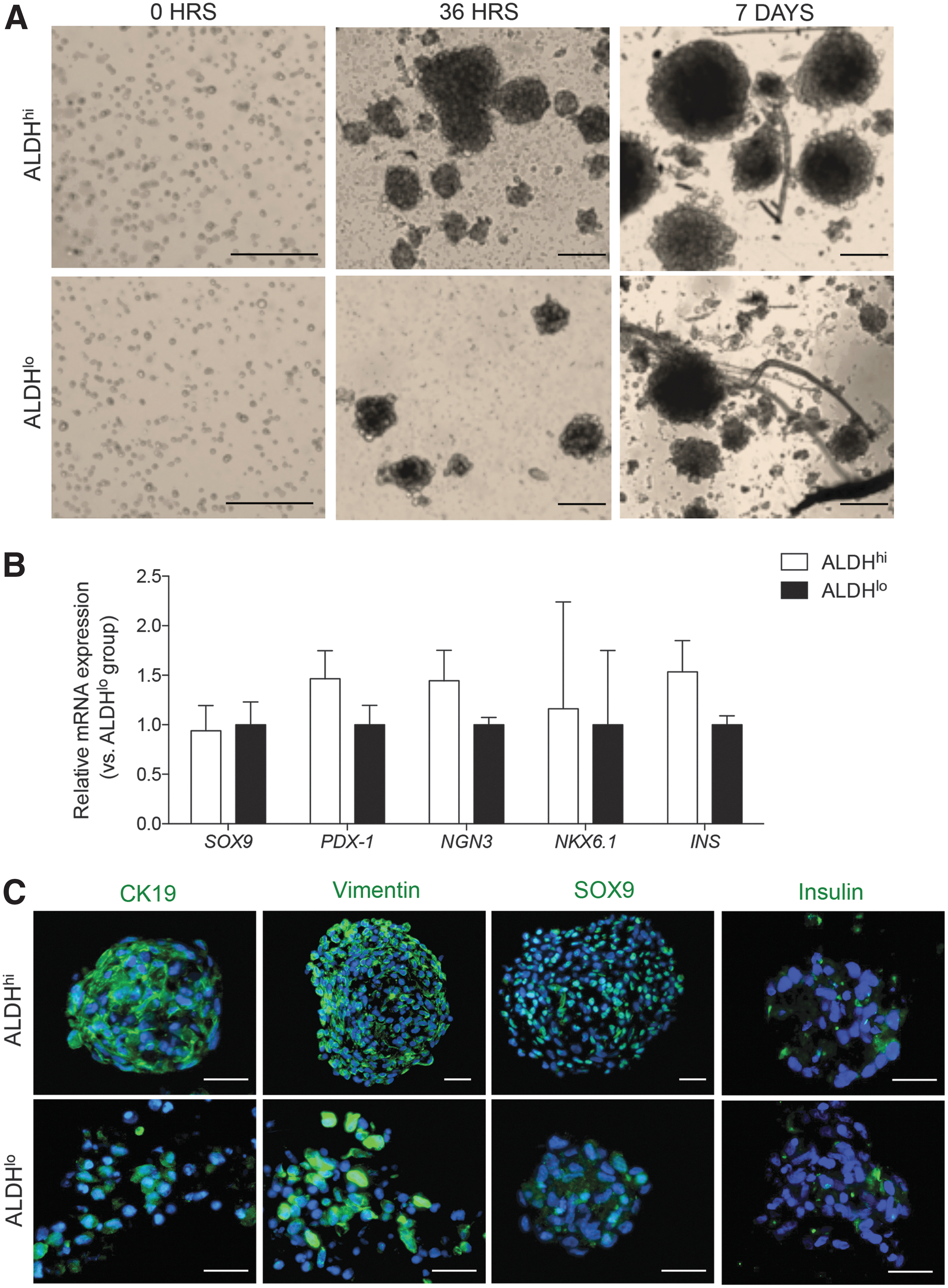
The presence of pancreatic TFs and hormones in clusters differentiated from expanded sorted populations.
Examination of ALDHhi/CD133 + sorted cells in vitro
Since sorted ALDHhi cells contained CD133+ cells with high SOX9, PDX-1, and NKX6.1 co-expression (Fig. 7A), we focused on expansion of the rare ALDHhi/CD133+ subpopulation to determine whether this cell type could produce an endocrine cell population in vitro. Although qRT-PCR analysis of the gene expression for TFs in expanded ALDHhi/CD133+ and ALDHlo/CD133− cells showed no significant differences in expression, higher levels of HES1 and AMYLASE (AMLY) genes were detected in the expanded ALDHhi/CD133+ cells (Fig. 7B).
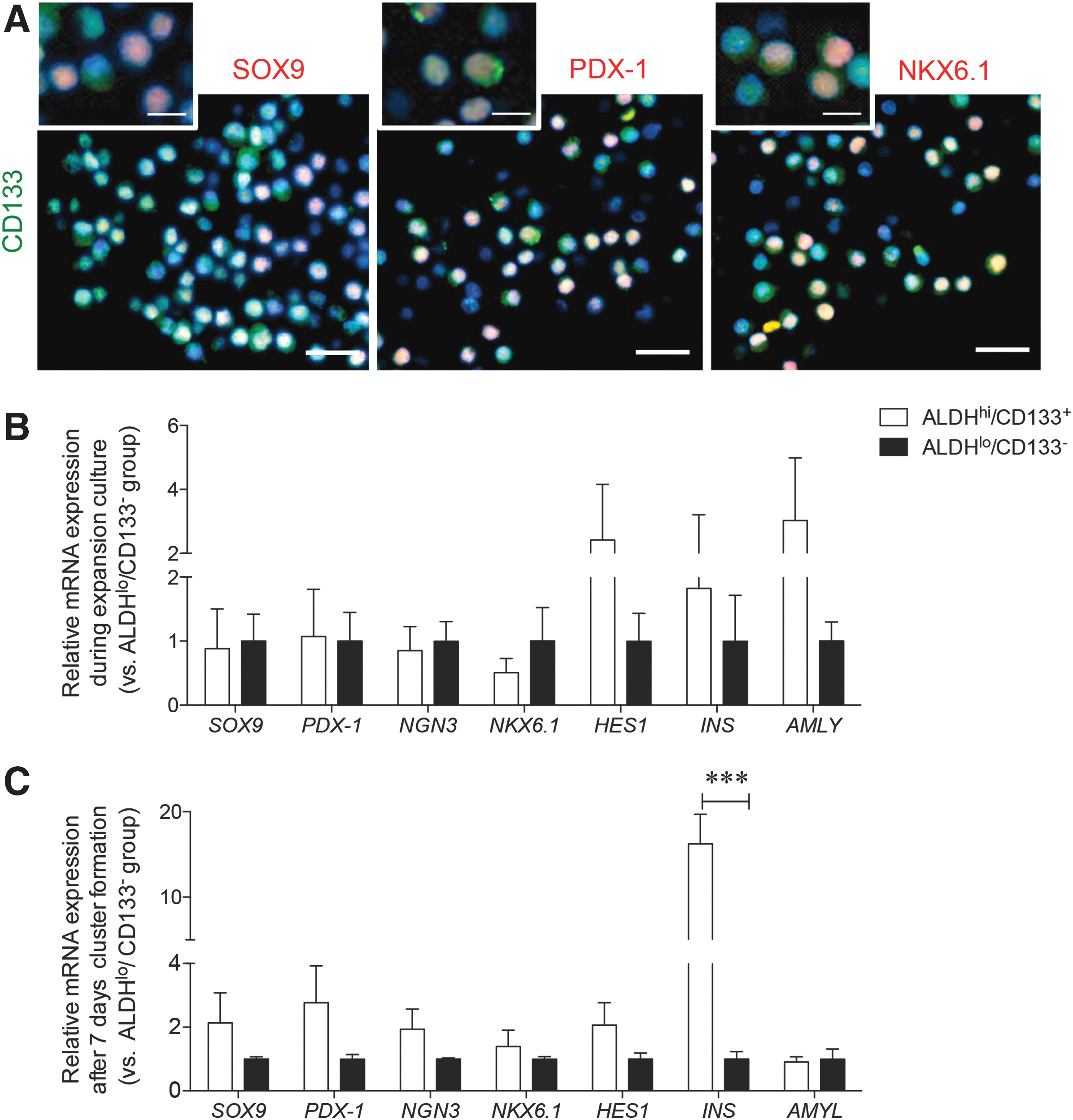
Analysis of ALDHhi/CD133+ and ALDHlo/CD133− expanded populations.
After the 7-day differentiation culture of ALDHhi/CD133+ and ALDHlo/CD133− cells, SOX9, PDX-1, NGN3, and NKX6.1 mRNA expression was increased in clusters derived from ALDHhi/CD133+ cells (Fig. 7C). Elevated INS gene expression was also detected in ALDHhi/CD133+ clusters (P < 0.001; Fig. 7C), but there was no change in AMLY gene expression.
Confirmation of qRT-PCR analyses was conducted using immunofluorescence staining of fixed cell clusters, and increased nuclear labeling for SOX9, PDX-1, and NKX6.1 in the ALDHhi/CD133+ clusters was detected with a few NGN3 nuclei positive cells present (Fig. 8A). Although these TFs were also present in the ALDHlo/CD133− clusters, staining was localized in their cytoplasm (Fig. 8A). Insulin staining was detected in both groups after treating cells with differentiation media, and quantification of the cell clusters found a higher percentage of insulin+ cells within the ALDHhi/CD133+ population (P < 0.01 vs. ALDHlo/CD133−; Fig. 8B). C-peptide staining was comparatively stronger in the ALDHhi/CD133+ clusters, while no difference was observed for glucagon and somatostatin staining (Fig. 8C). Both CK19+ and desmin+ cells were observed in the ALDHhi/CD133+ and ALDHlo/CD133− clusters (Fig. 8).

Immunofluorescence staining evaluation of ALDHhi/CD133+ islet-like clusters. Immunofluorescence labeling (green) of endocrine TFs and ductal marker CK19
Discussion
In this study, we examined sorted ALDHhi cells from the developing human pancreas at 18–22 weeks of fetal age, found that ALDHhi cells highly co-expressed CD133 and early endocrine-associated TFs, and insulin and somatostatin. During expansion under nondifferentiative culture conditions, sorted ALDHhi and ALDHhi/CD133+ cells lost ALDH, endocrine TFs, insulin, and CK19 expression, with an increase in vimentin+ cells. In contrast, culture of these cells under differentiation-promoting conditions induced cluster formation and led to increased expression of endocrine TFs and INS mRNA in ALDHhi and ALDHhi/CD133+ clusters. These data suggested that ALDHhi and ALDHhi/CD133+ populations represent a heterogeneous pancreatic pool with an increased subset of cells expressing TFs required for the generation of endocrine cells in the developing human pancreas.
Sorted ALDHhi cells highly co-expressed CD133, which is found on ductal cells that acquire NGN3 expression during human pancreatic development [26]. CD133 also co-localized with ALDH+ centroacinar cells in adult murine pancreata that can form insulin-producing cells [18], suggesting that this marker can be used to isolate cells with the potential to form endocrine hormone-producing cells. Sorted ALDHhi/CD133+ cells show increased SOX9, PDX-1, and NKX6.1 expression, which reflects the association of CD133 as a pancreatic ductal cell marker and with the developing endocrine lineage [26 –29].
Sorted cells with high ALDH activity and CD133 expression did not correlate with CD34+ cells. CD34+ cells, which were found prominently in the ALDHlo subset, have been shown to co-localize with the vasculature in developing pancreata [30,31] and indicates a potentially separate pancreatic cell fate than ALDHhi and CD133+ cells.
Although both sorted ALDHhi and ALDHlo populations were heterogeneous and formed multiple pancreatic cell lineages, endocrine-associated TFs were increased in the ALDHhi cell subset. The pancreatic progenitor TF SOX9 [32] was highly expressed in ALDHhi cells and has been previously associated with cells demonstrating high ALDH activity in isolated human fetal islet-epithelial clusters [10]. NGN3+ cells were increased in purified ALDHhi cells, which require early SOX9 expression and eventual downregulation to commit toward an endocrine cell fate [29], but also indicates that the expression of both TFs in the ALDHhi group represent a pool of cells more closely related to early pancreatic progenitor fate and not of mature endocrine cells. NKX6.1 and ISL1, two TFs that are essential for β-cell maturation and regulation [33 –36], were also expressed in ALDHhi cells.
PDX-1 is expressed throughout the pancreatic epithelium during early development (8–11 weeks) and co-localize with insulin+ and CK19+ cells at 19–21 weeks of development, indicating islet neogenesis and subsequent endocrine cell commitment at this stage [33,37]. The presence of increased PDX-1+, CK19+, insulin+, and somatostatin+ cells in the sorted ALDHhi cells indicates a population that were composed predominantly of cells with ductal markers and early endocrine-associated TFs, with a small population of cells that are positive for endocrine hormones. Exocrine-linked markers HES1 and amylase expression were equivalent in both sorted cell groups. Because exogenous RA treatment has been shown to increase [38] and decrease [39] the number of amylase+ cells in the developing murine pancreas, enhanced RA production within ALDHhi cells may also regulate exocrine commitment during the second trimester of pancreatic development.
Under expansion conditions, both vimentin and E-cadherin were expressed in ALDHhi and ALDHhi/CD133+ populations. Epithelial-to-mesenchymal transition (EMT), previously observed in insulin-expressing cells migrating from human fetal islets in vitro [22], is one hypothesized mechanism for β-cell formation in vivo [40]. Similar to our cultured ALDHhi and ALDHhi/CD133+ cell subsets, human pancreatic ductal cells and nonendocrine PDX-1+ human pancreatic cells demonstrate increased vimentin expression with repeated passages [41,42]. Interestingly, both of these studies achieved endocrine-like fate and upregulation of insulin+ cells when exposed to differentiation conditions. However, conflicting results refute EMT from β-cells in murine islets in vitro [43], and it should be taken into consideration that adherent vimentin+ cells were initially present in the heterogeneous sorted populations.
Serum-free differentiation media have been used in early β cell differentiation protocols [22 –24,44 –46]. The treatment of expanded mesenchymal-like precursor cells from adult islets with serum-free media induced cluster formation and increased expression of markers associated with endocrine cells [22,45,46]. When mesenchymal-like expanded ALDHhi and ALDHhi/CD133+ cells were treated with a similar media, both populations demonstrated cell cluster aggregation, increased expression of endocrine TFs, and the presence of insulin+ and c-peptide+ cells. Although this was also achieved in ALDHlo and ALDHlo/CD133− populations, the gene expression of endocrine TFs and INSULIN was lower than in ALDHhi and ALDHhi/CD133+ clusters.
The findings in this study are supported by similar experiments using NGN3+ cells from the adult pancreatic exocrine compartment with high CD133 co-localization, where pancreatosphere formation and upregulation of associated endocrine TFs were seen upon serum-free expansion in vitro [47]. However, it is important to note that increased expression of insulin+ cells in the ALDHhi and ALDHhi/CD133+ clusters may not indicate fully differentiated β cell–like cells. The upregulation of early endocrine TFs were observed in these clusters, and it has been previously established that insulin+ cells are detected as early as 9–10 weeks of fetal age in the developing pancreas [37].
Another limitation in this study is that current culture conditions in this experiment could only produce a small percentage of insulin+ cells in the ALDHhi/CD133+ group, which limits the potential to test clusters for glucose responsive and insulin secreting β cell–like cells. Both ALDHlo and ALDHlo/CD133− clusters were also able to yield hormone-expressing cells with this protocol, and indicates that ALDH activity is not mandatory for the in vitro differentiation of pancreatic precursor cells. Additional factors used previously to stimulate insulin production [48,49] may be necessary to overcome this limitation and bolster glucose responsiveness from formed ALDHhi and ALDHhi/CD133+ β cell–like clusters.
Adapting multiple step protocols that are able to induce insulin-producing cells from cells with a pancreatic progenitor cell type similar to ALDHhi and ALDHhi/CD133+ populations can also be ideal for further efficient maturation of the generated clusters. This has recently been shown in progenitor stem cell-derived NGN3+/NKX6.1+ cells, where the use of a guided differentiation protocol resulted in the production of glucose-sensitive insulin-secreting cells [50].
In conclusion, we have shown that sorted pancreatic ALDHhi cells that expressed the CD133 surface marker receptor represented a pool of TFs associated with the endocrine precursor population, and demonstrated the capacity to increase endocrine-associated TFs in vitro. Characterizing factors in human pancreata that form islets and drive β cell commitment is necessary for understanding the generation of β cells in vivo, and will aid in furthering therapies that target β cell restoration in diabetic patients. Additional studies are also required to examine optimized culture environments for efficient generation of functional insulin-producing islet cells in vitro.
Footnotes
Acknowledgments
The authors would like to thank Dr. Kristin Chadwick from the London Regional Flow Cytometry Facility for her assistance with cell sorting of pancreatic samples. This work was supported by grants from the Canadian Diabetes Association (CDA, grant no. OG-3-11-3318-RW).
Portions of the data in this article were presented at the 51st European Association for the Study of Diabetes (EASD) Annual Meeting, held September 14–18, 2015.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.