
Abstract
Ovarian cancer is most lethal among gynecological cancers with often fatal consequences due to lack of effective biomarkers and relapse, which propels ovarian cancer research into unique directions to establish solid targeted therapeutics. “Ovarian stem cells” expressing germline pluripotent markers serve as novel paradigm with potential to address infertility, menopause, and probably influence tumor initiation. Cancer stem cells (CSCs) pose vital role in tumor recurrence and hence it is extremely important to study them with respect to ovarian stem cells across various cancer stages and normal ovaries. Pluripotent (OCT4, NANOG, SOX2, SSEA1, and SSEA4), germline (IFITM3, VASA/DDX4), and cancer stem (CD44, LGR5) cell specific markers were characterized for protein and mRNA expression in tumor tissues to understand their distribution in the surface epithelium and ovarian cortex in benign, borderline, and high-grade malignant stages. To elucidate whether pluripotent ovarian germline stem cells and CSCs are common subset of stem cells in tumor tissues, VASA was colocalized with known pluripotent stem (OCT4, SSEA1, SSEA4) and CSC (CD44, LGR5) specific markers by confocal microscopy. Single, smaller spherical (≤5 μm), and larger elliptical fibroblast like (≥10 μm) cells (also in clusters or multiples) were detected implying probable functional behavioral significance of cells in tumor initiation and metastasis across various cancer stages. Cells revealed characteristic staining pattern in ovarian surface epithelium (OSE) and cortex regions exclusive for each marker. Co-expression studies revealed specific subpopulations existing simultaneously in OSE and cortex and that a dynamic hierarchy of (cancer) stem cells with germline properties prevails in normal ovaries and cancer stages. Novel insights into CSC biology with respect to ovarian and germline stem cell perspective were obtained. Understanding molecular signatures and distribution within ovarian tissue may enable identification of precise tumor-initiating CSC populations and signaling pathways thus improving their efficient targeting and strategies to prevent their dissemination causing fatal relapse.
Introduction
S
Cancer initiation and resultant propagation/invasion entail a complex series of cross talk of events at cellular and molecular levels [15], while the role played by CSCs requires to be rigorously investigated. There is no consensus regarding true identity of ovarian CSCs and their signaling mechanisms, their location, precise cell(s) of origin, and their individual response to novel drugs. Moreover, the putative CSC populations with ovarian stem cell perspective have never been studied across various stages of ovarian cancer. Tissue specific CSCs have already been identified in ovarian cancer, but it remains to be determined if germline stem cells reported in normal ovaries express these pluripotent stem and CSC specific markers. To reiterate, the goal of present study was to correlate the expression of pluripotent stem cells with germ lineage markers.
Presence of germline stem cells termed “oogonial stem cells” (OGSCs)/“female germline stem cells” (FGSCs) was first put forth by Prof. Jonathan Tilly and his group in adult mice [16] and later in adult mice and human ovaries [17]. Similarly, pluripotent stem cells have been demonstrated in adult human ovaries [2,3,18 –22] and mice [22,23] by certain groups independently. Parallel to these reports, a pluripotent stem cell population with embryonic characteristics named very small embryonic-like stem cells (VSELs) has been identified in various adult tissues and organs. These peculiar, tiny spherical stem cells are remnants of migratory PGCs gone astray during embryogenesis as they share common markers [24,25]. Similarly mouse hematopoietic stem progenitor cells (HSPCs) show germline connection, and thus, a hierarchical link among murine VSELs, HSPCs, and primordial germ cells (PGCs) was recently speculated [26]. Likewise a germline connection with the tumors was proposed [27] with pluripotent adult stem cells showing embryonic characteristics (VSELs) as the putative connecting link. These VSEL cells have been speculated to differentiate into CSC populations [28,29].
VASA/DDX4 is a member of DEAD box protein family involved in processes such as embryogenesis, gametogenesis, cellular growth, division and PGC development, and so on [30]. Besides its multifaceted cellular functions, recently its expression was linked to tumor formation and maintenance in drosophila brain [31], as well as in tumor progression and poor patient prognosis in case of epithelial ovarian cancer cells [32,33]. DEAD-box helicase 4 (DDX4) is also implicated in cell cycle regulation in multipotent and proliferating cells by influencing 14-3-3 sigma at G2/M checkpoint. Hence it becomes imperative to pinpoint the expression of pluripotent stem cell population(s) with germline characteristics existing within the ovarian tumors in comparison to normal ovaries and investigate their relation with other pluripotent and CSC markers.
Identification of unique gene signatures for detection of early onset, predicting patient response to treatment and their survival, has been the Achilles' heel so far in cancer research. The known pluripotent stem cell population namely ovarian (germline) stem cells and (cancer) stem cells both were studied systematically in various stages of ovarian cancer in comparison to normal ovaries. Single and double immunolabeling experiments interestingly revealed dynamic populations of pluripotent stem cells with germline and CSC specific properties within the OSE and cortex regions of the ovary. Current study provides novel insights into the stem cell profiles in normal versus ovarian tumor tissues and consolidates the germline connection with normal stem cells, as well as CSCs.
Materials and Methods
Ovarian tissue acquisition and ethical approval
Ovarian tissues were obtained from patients undergoing surgery as part of their clinical management for gynecological problems and as a therapeutic procedure against cancer from James Graham Brown Cancer Center, University of Louisville in compliance with the standards and protocols approved by the Institutional Review Board (IRB). Consent from the patient was obtained before surgery, and tissues were acquired. A total of 50 ovarian samples were used in the study of which 16 fresh and frozen tissues (n = 4 for each category-NO, BN, BL, and HG) were obtained from institutional biorepository and were used for RNA extraction and reverse transcription polymerase chain reaction (RT-PCR) purpose. Thirty four ovarian tissue sections [Normal ovaries (4), Benign tumor (10), Borderline tumor (10), and High Grade tumor (10)] were obtained from Department of Pathology, University of Louisville under supervision of an expert pathologist and were used in the study for immunolocalization purpose. Experiments were repeated at least thrice on independent occasions. All chemicals, reagents, kits, and plastic ware utilized in present study were purchased from Sigma-Aldrich or otherwise specified. All data generated or analyzed during this study are included in this published article.
RNA extraction and cDNA preparation
RNA was purified from n = 16 normal ovarian and tumor tissues using TRIzol reagent (Invitrogen) as described previously [34] following the instructions from the supplier. DNA contamination was eliminated by incubating RNA with DNaseI following standard protocol as suggested by the manufacturer. DNaseI was inactivated by heating the sample at 70°C for 5 min followed by snap chill on ice, and RNA was stored at −80°C. RNA for each sample was quantified with a NanoDrop spectrophotometer (Fisher Scientific). One microgram of RNA from each sample was converted to cDNA using iScript cDNA Synthesis Kit from Bio-Rad Laboratories according to instructions from the supplier.
PCR amplification of genes
Two microliters of first strand cDNA from each sample was amplified in PCR using 10 pmol of each primer (Table 1) in a 20 μL of reaction mix containing 12.5 μL of REDTaq ReadyMix comprising MgCl2, dNTPs, Taq polymerase (Sigma), and 10.5 μL of H2O. All the contents were mixed and amplified in a thermal cycler (Eppendorf). Amplification was carried out by initial denaturation at 95°C for 5 min, followed by 94°C for 1 min, annealing temperature as optimized for each primer (Table 1) for 1 min followed by elongation at 72°C for 1 min for 30–35 cycles with a final extension at 72°C for 7 min. The PCR product was electrophoresed on 2% agarose gel containing 5 μg/mL ethidium bromide in 1 × TBE buffer at 100 V. Amplicon size was determined using 100-bp DNA ladder to confirm specific band size.
House-keeping gene.
Pluripotent stem cell.
Cancer stem cell.
Germline stem cell genes.
DDX4, DEAD-box helicase 4; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Immunohistochemical analysis and colocalization of various stem cell/CSC markers
To document the localization of stem cells/CSCs, we performed immunohistochemistry (IHC) analysis for the normal and cancer tissues as described previously [35] with modifications. Briefly, paraffin embedded sections were deparaffinized in xylene, treated with 0.3% hydrogen peroxide in 100% methanol for 60 min at room temperature to quench endogenous peroxidase, and rehydrated in decreasing graded series of ethanol. Sections were heated at 95°C for 10 min in 10 mM sodium citrate buffer pH 6.0, for antigen retrieval followed by two rinses with phosphate buffered saline (PBS). Tissues were permeabilized with 0.3% Triton X-100 (3–5 min for surface; 5–7 min for cytoplasmic; and 10–12 min for nuclear markers, respectively). The sections were blocked with serum from ABC kit (Vector Laboratory, Inc.) for 60 min followed by incubation with marker specific antibody (Table 2) at 4°C overnight. After rinsing the sections thrice with PBS (5 min each), sections were incubated with biotinylated anti-mouse or anti-rabbit secondary antibody for 45 min at room temperature followed by three rinses with PBS and incubation with streptavidin as described previously [35]. The sections were washed with PBS and incubated with 3,3′-diaminobenzidine (DAB; Sigma) for color development and were examined using Aperio scanner and Olympus IX71 inverted microscope (Olympus, Tokyo, Japan) and photographed. Average staining intensity of DAB was optimized systematically for each primary antibody against a negative control used by omitting primary antibody (Supplementary Fig. S1; Supplementary Data are available online at
C, cytoplasmic; CSC, cancer stem cell; GSC, germline stem cell; HSC, hematopoietic stem cell; N, nuclear; PSC, pluripotent stem cell; S, surface.
We investigated if germline stem cell marker (specific for ovarian stem cells) is coexpressed with CSC marker by performing dual labeling for VASA and pluripotent stem cell or CSC marker by fluorescence confocal microscopy. After incubation with the primary antibody (as described above), sections were rinsed with wash solution (0.5% bovine serum albumin in PBS) and incubated with secondary antibody [Alexa Fluor 488/568 labelled goat anti-mouse IgG, or goat anti rabbit-IgG (1: 1,000)] (Molecular Probes, Life Technologies) and counterstained with nucleus specific dye 4′, 6-diamidino-2-phenylindole (DAPI; Sigma). After rinsing with PBS the slides were dehydrated in alcohol grades and mounted (Eukitt® Quick-hardening; Sigma-Aldrich) in mounting medium, and representative images were captured using Nikon (Eclipse TI) laser scanning confocal microscope and documented with the help of NIS Elements AR software (version 4.51). Similar procedure was followed for single marker immunofluorescence staining of samples. Special attention was paid to optimize image acquisition parameters for each antibody especially for colocalization studies of VASA with other markers, and representative images were captured at constant settings to maintain uniformity in the documentation of results. For both immunolocalization techniques on an average 7–15 images were captured per sample/antibody used and representative images from one or several fields were compiled to prepare the composite result panels.
Results
Identification of the putative populations with pluripotent stem and germline stem cell characteristics in normal ovary is very important to implicate these cells in the etiology of ovarian cancer or search for the cell/(s) of origin for ovarian cancer. Besides, it is essential to understand the prevalence and distribution of various subpopulations of these tumor initiating cells across cancer stages (described in Supplementary Tables S1 and S2). We thus studied various putative markers for expression of mRNA transcripts and proteins in situ in normal ovarian and tumor tissue sections (Table 3). Negative controls by omission of primary antibody were used which revealed nil staining (Supplementary Fig. S1).
Pluripotent stem cell gene.
Adult stem cell gene.
Cancer stem cell gene.
Germline stem cell gene.
House-keeping gene.
IF, immunofluorescence; PCR, polymerase chain reaction; ✓, gene employed in the technique; —, gene not employed in the technique.
Expression of pluripotent stem and germline stem cell specific mRNA transcripts in normal ovary and different stages of cancer
RT-PCR was performed for detecting mRNA transcripts specific for various pluripotent stem cell markers such as Oct4, Nanog, and Sox2, germline stem cell specific Fragilis and Vasa; and housekeeping gene Gapdh (Table 1). Amplicons of expected size were amplified across four sets of samples comprising normal ovary (NO), benign (BN) tumor, borderline/low malignant potential (BL), and high grade/high malignant potential (HG) ovarian tumor (Fig. 1). Variations in band intensity of the amplicons of mRNA transcripts for genes especially Oct4 and Vasa were prominently observed from patient to patient. These results were congruent with those observed in terms of protein expression in vivo by immunohistochemical analysis (Figs. 2 –14) within the ovarian tissue and tumor tissue sections. Reverse transcriptase and no template cDNA (negative) control samples were amplified in separate experiments using the same primers, and no amplification was confirmed.
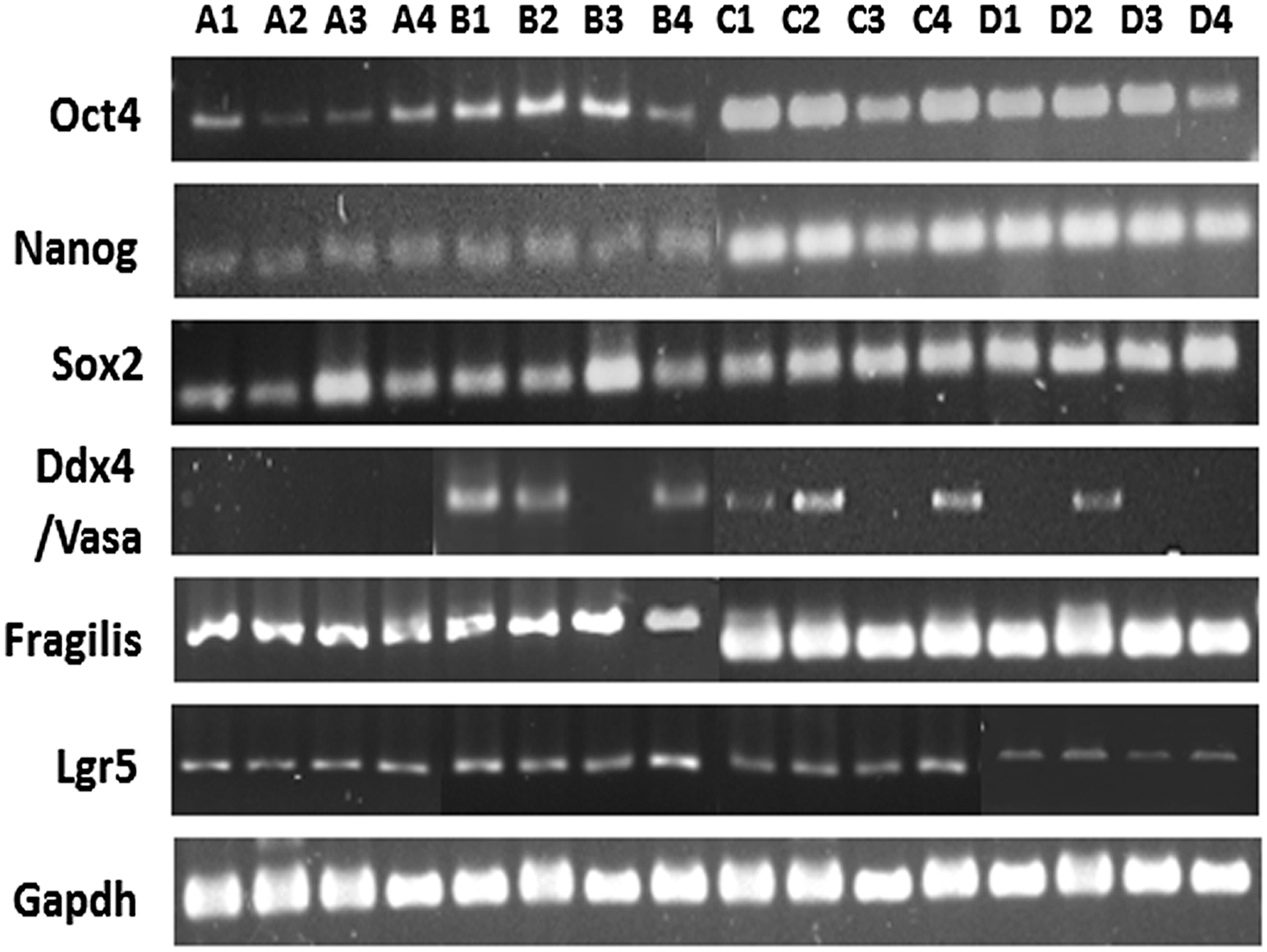
Gene expression analysis by RT-PCR for pluripotent, germline, and cancer stem cells from ovarian and tumor tissues: Presence of various mRNA transcripts was investigated by RT-PCR analysis followed by gel electrophoresis, and amplicons of desired base pair lengths were observed for various genes such as pluripotent stem (Oct4, Nanog, Sox2), germline stem (Vasa, Fragilis), cancer (stem) (Lgr5) cell, and housekeeping gene (Gapdh), which were detected in various normal human ovarian and tumor tissue samples. 1 = NO (normal) ovarian tissue; 2 = BN (benign); 3 = BL (borderline); 4 = HG (high grade) ovarian tumor, respectively; A = set 1; B = set 2; C = set 3; D = set 4 of independent patient samples. RT-PCR, reverse transcription polymerase chain reaction.
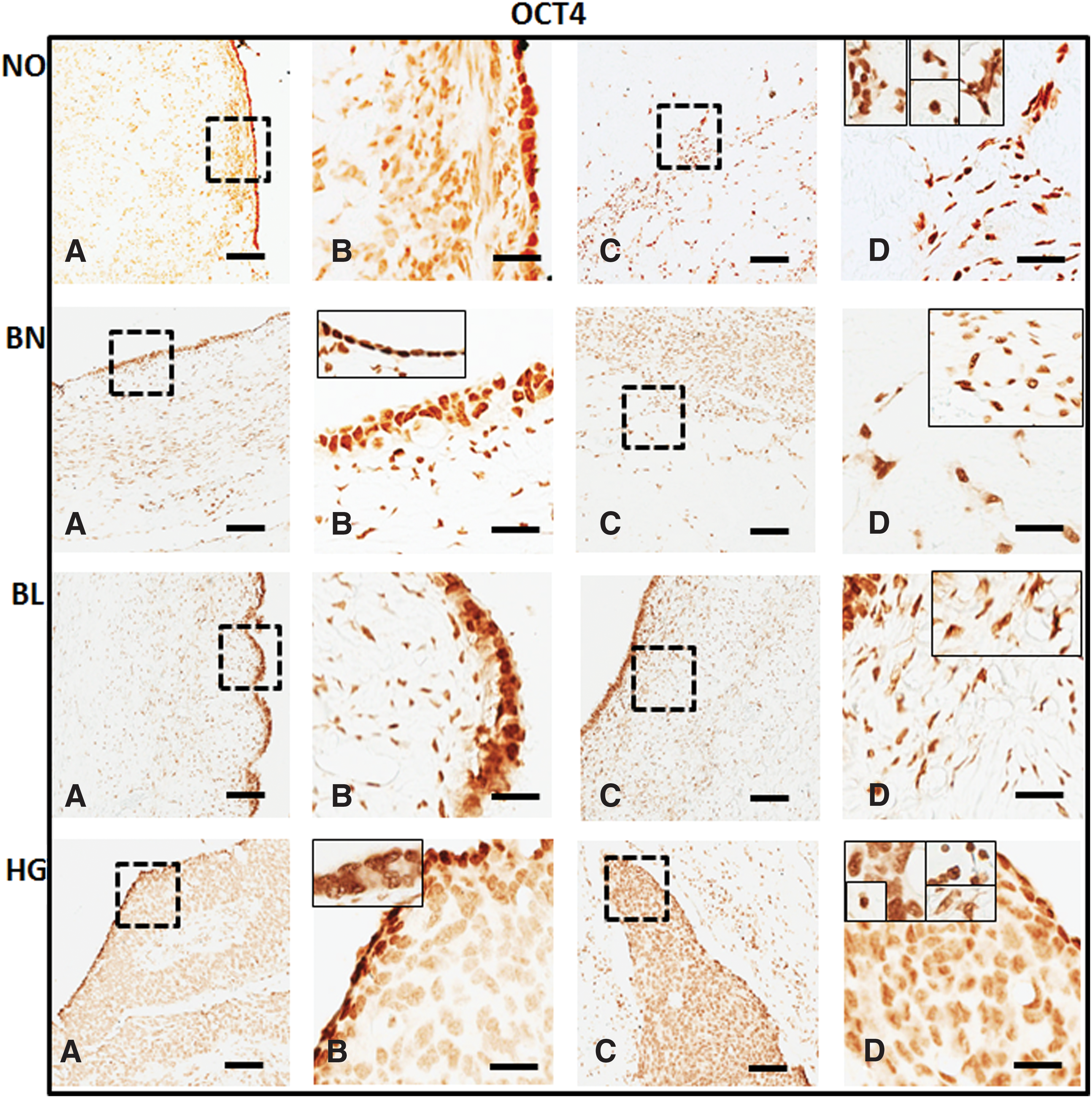
Detection of pluripotent stem cell marker OCT4 in normal (NO), benign (BN), borderline (BL), and high grade (HG) ovarian cancer tissues: mouse monoclonal anti-OCT4 antibody was localized within normal ovarian and tumor tissue sections. OCT4 is localized in both OSE
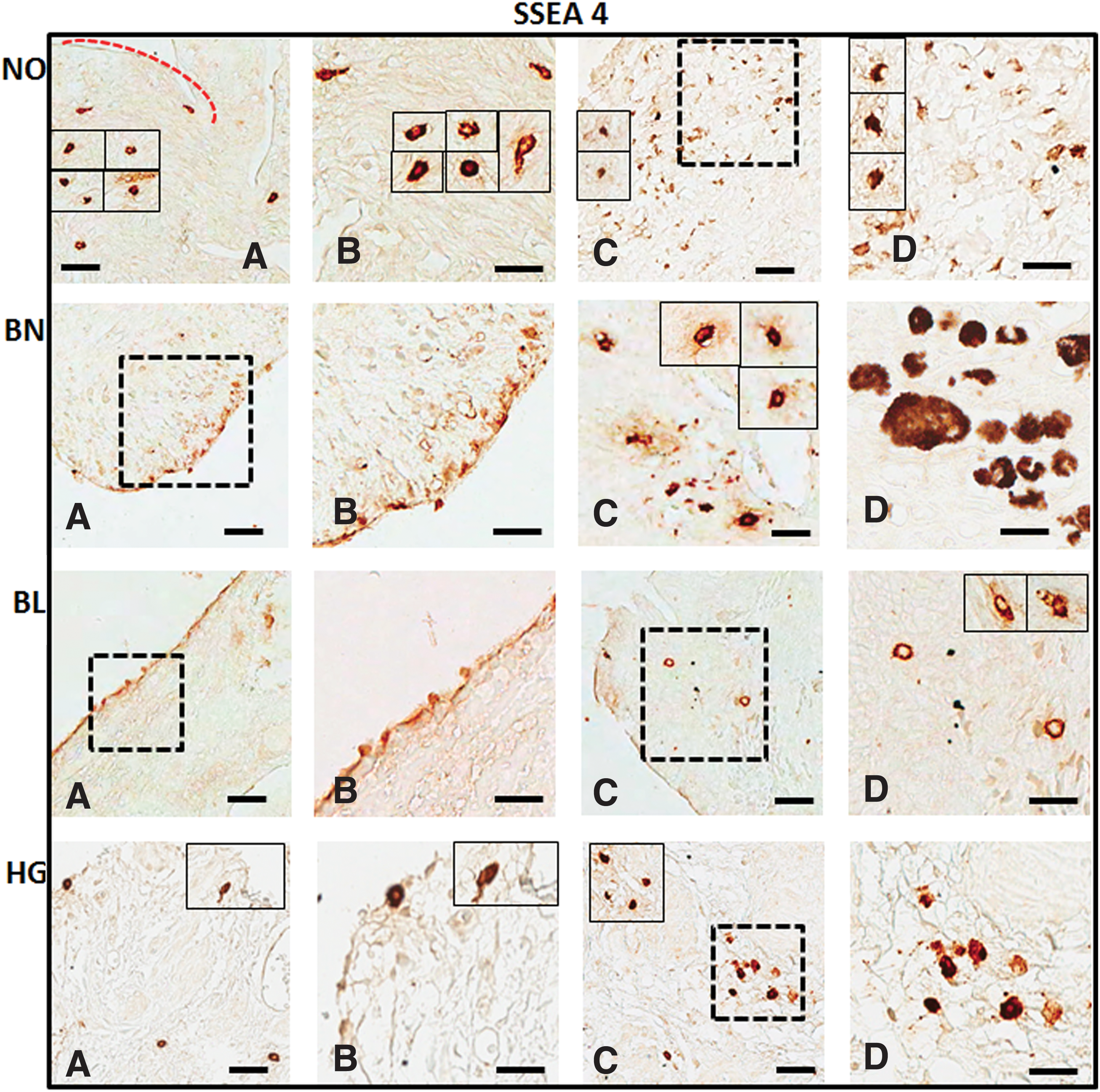
Expression of cell surface pluripotent stem cell marker SSEA4 in normal ovarian (NO), benign (BN), borderline (BL), and high grade (HG) ovarian cancer tissues: mouse monoclonal anti-SSEA4 antibody was localized in both OSE
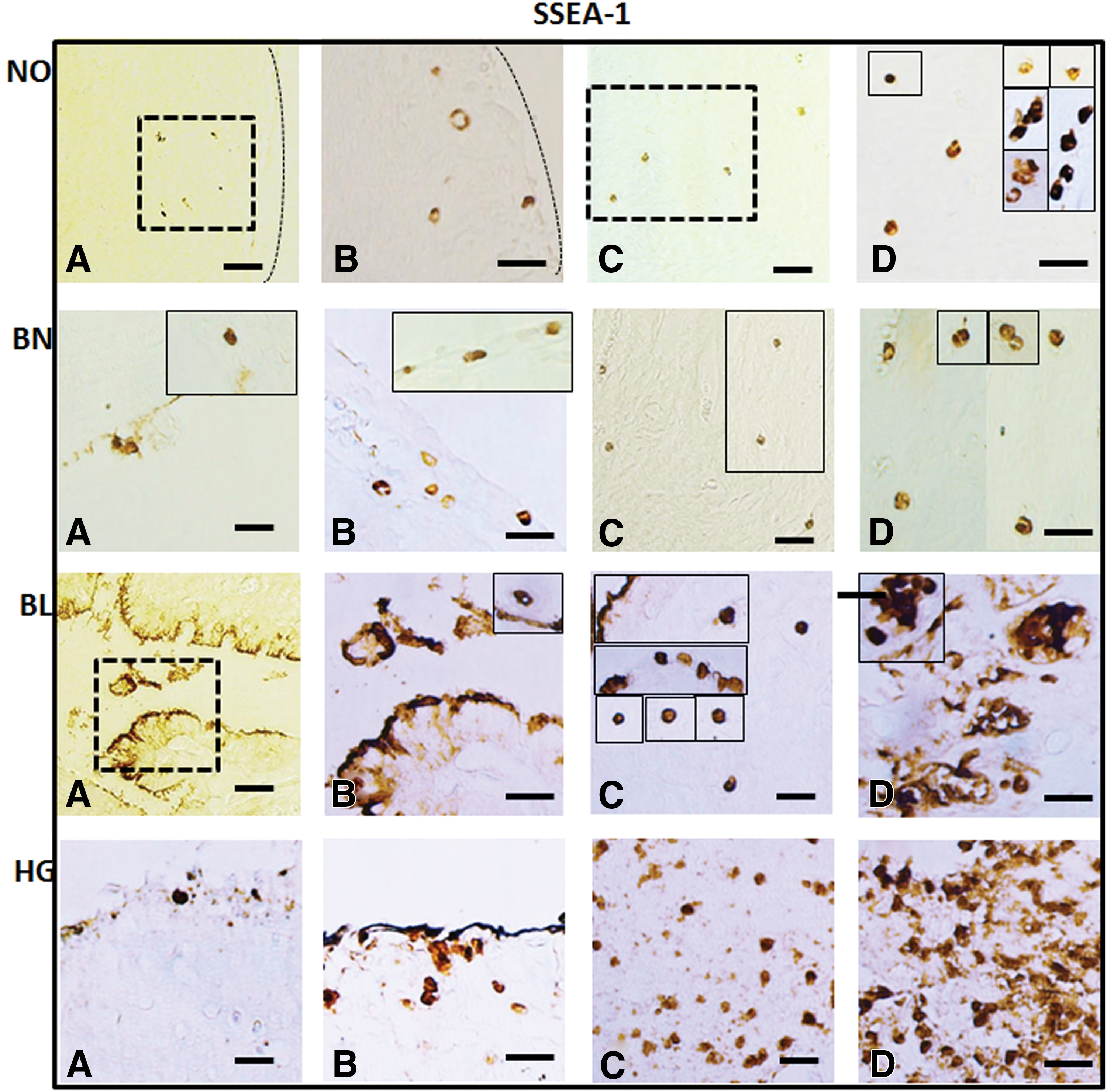
Detection of cell surface marker SSEA1 in normal (NO), benign (BN), borderline (BL), and high grade (HG) ovarian cancer tissues: mouse monoclonal anti-SSEA1 antibody was localized in both OSE
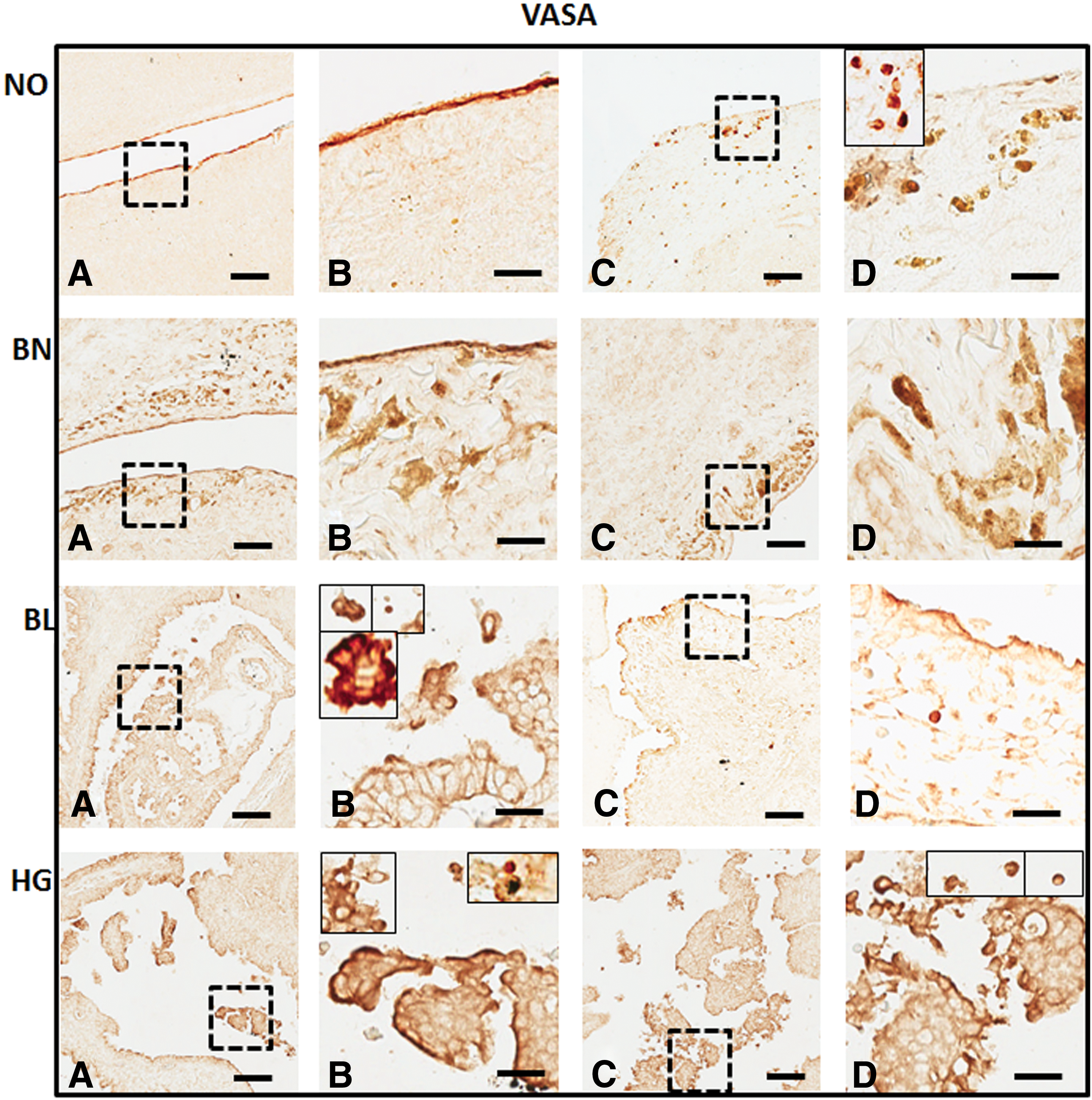
Immunohistochemical detection of VASA from tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) ovarian cancer: polyclonal anti-VASA antibody was localized in the OSE
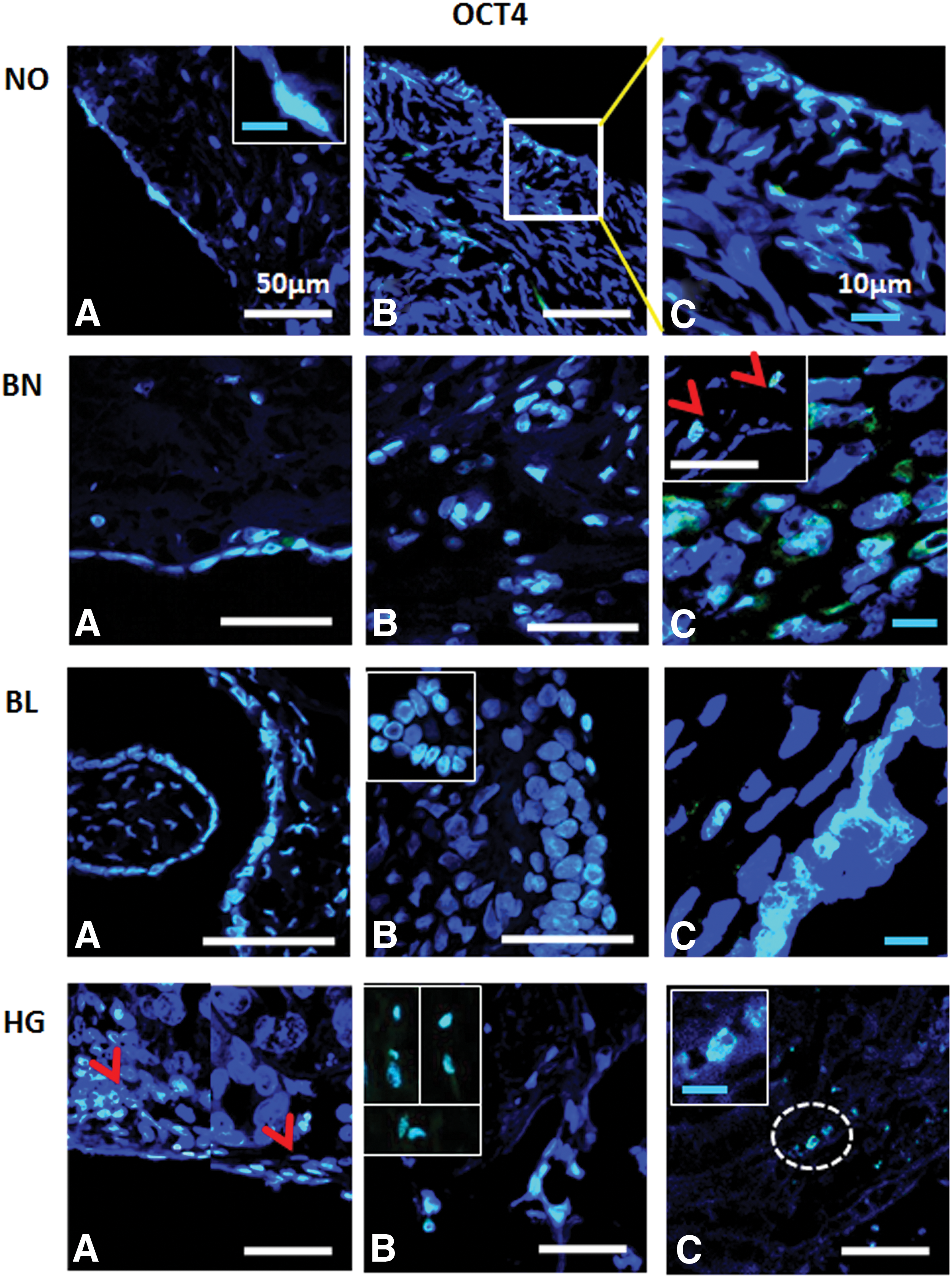
Immunofluorescence detection of OCT4 from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: OCT4+ signals were observed in OSE
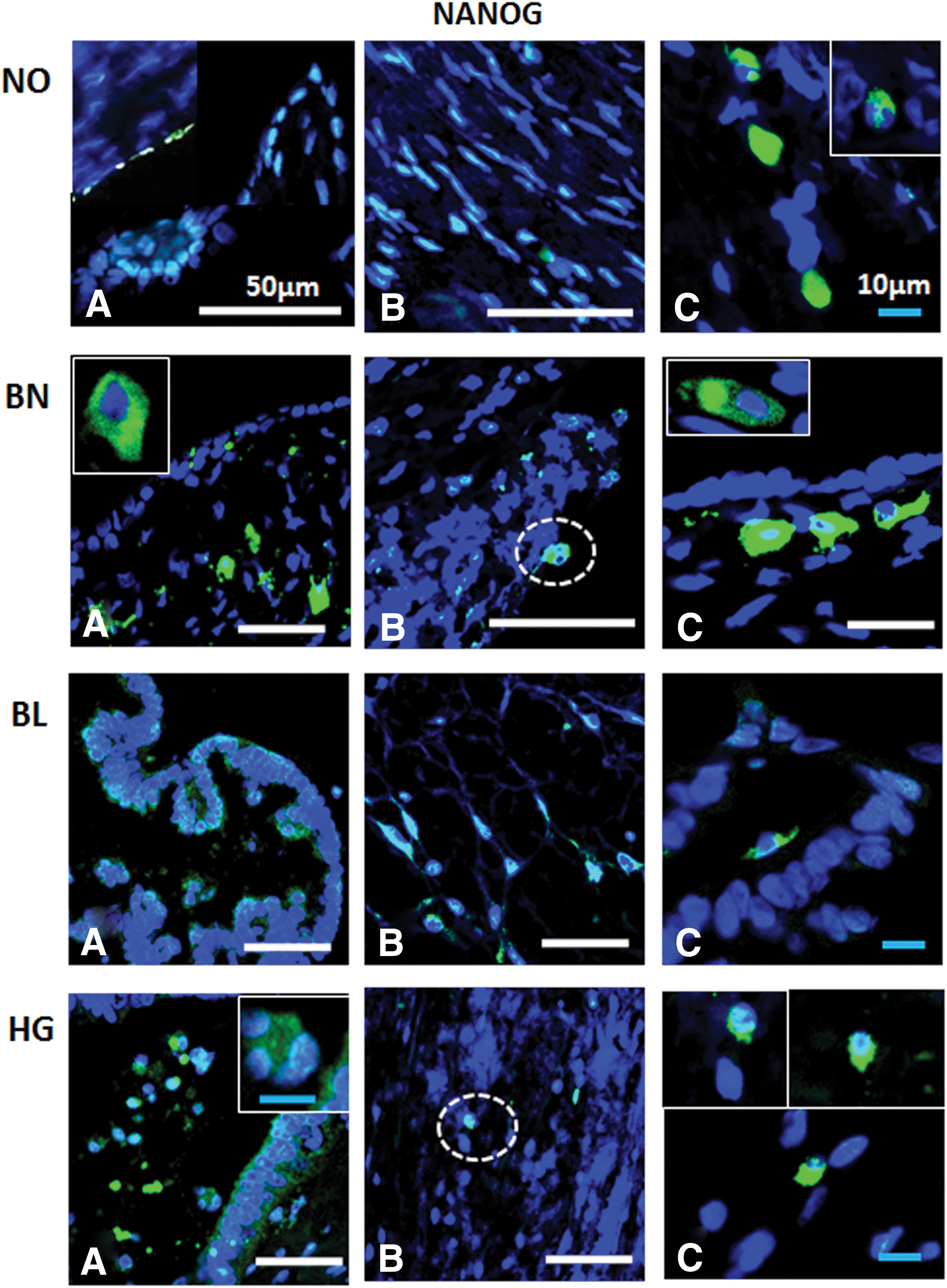
Immunofluorescence detection of NANOG from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: NANOG+ cells were observed in OSE
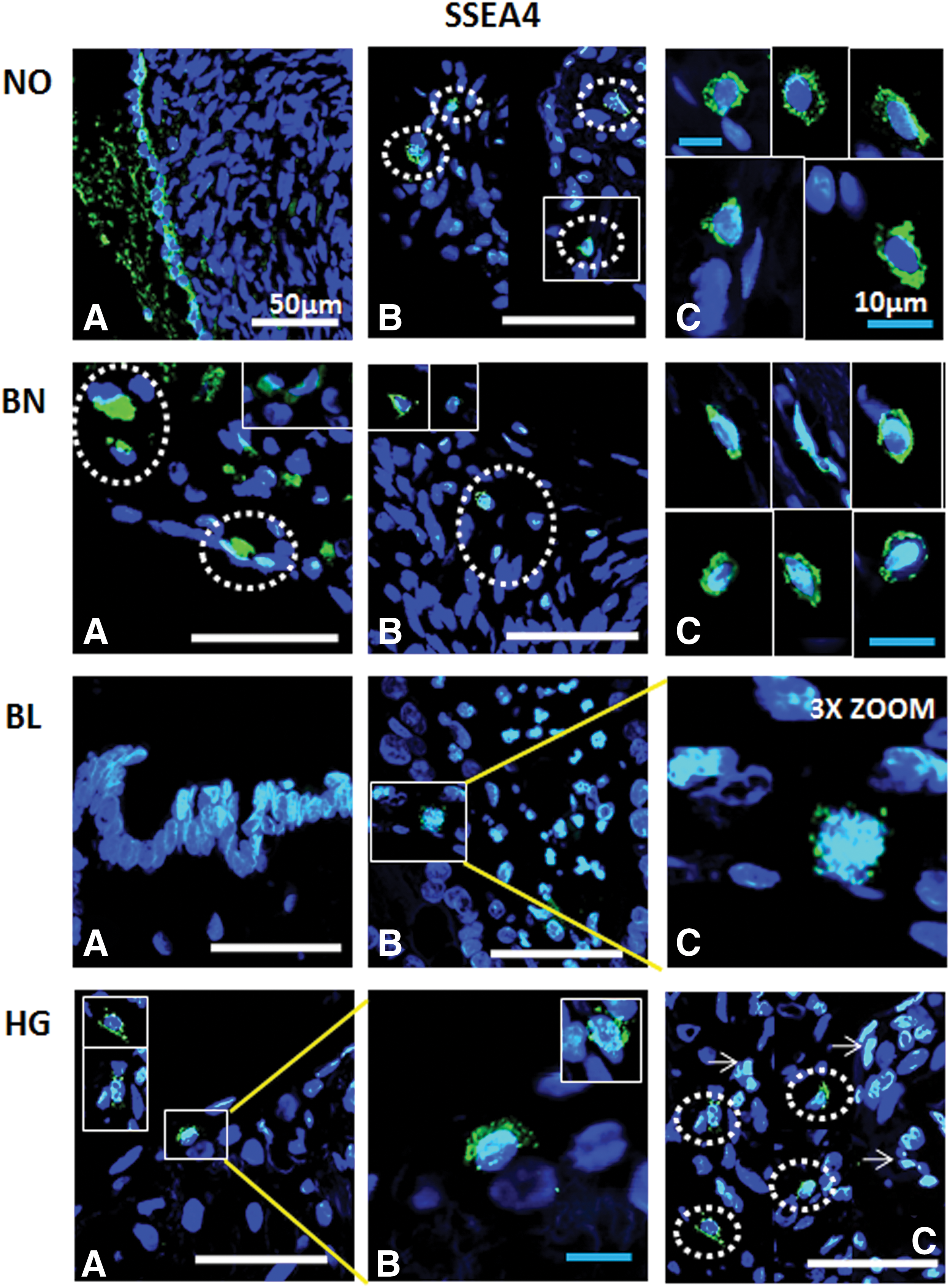
Immunofluorescence detection of SSEA4 from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: OSE layer
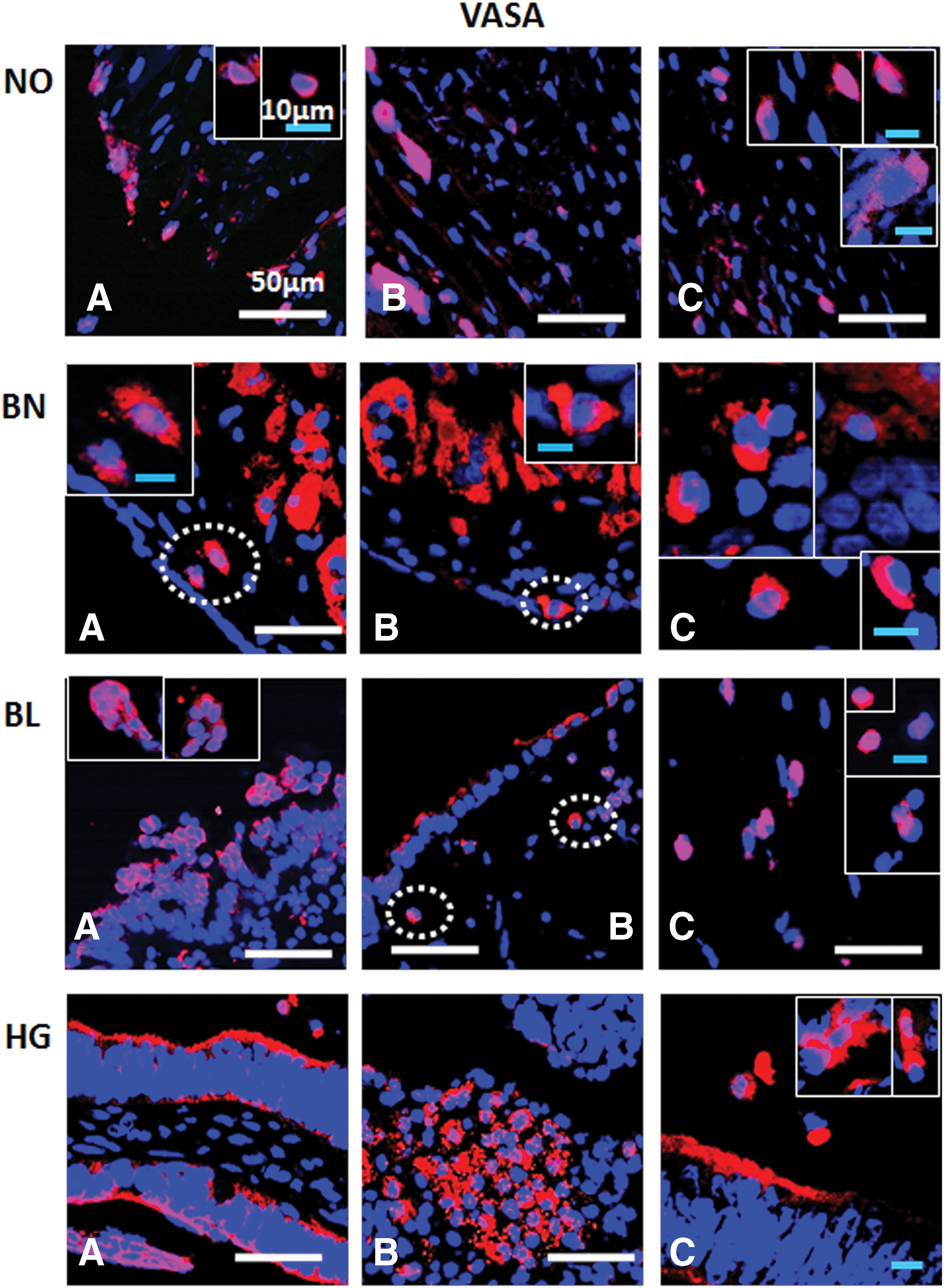
Immunofluorescence staining of VASA/DDX4 from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: VASA+ cells are consistently observed in OSE layer
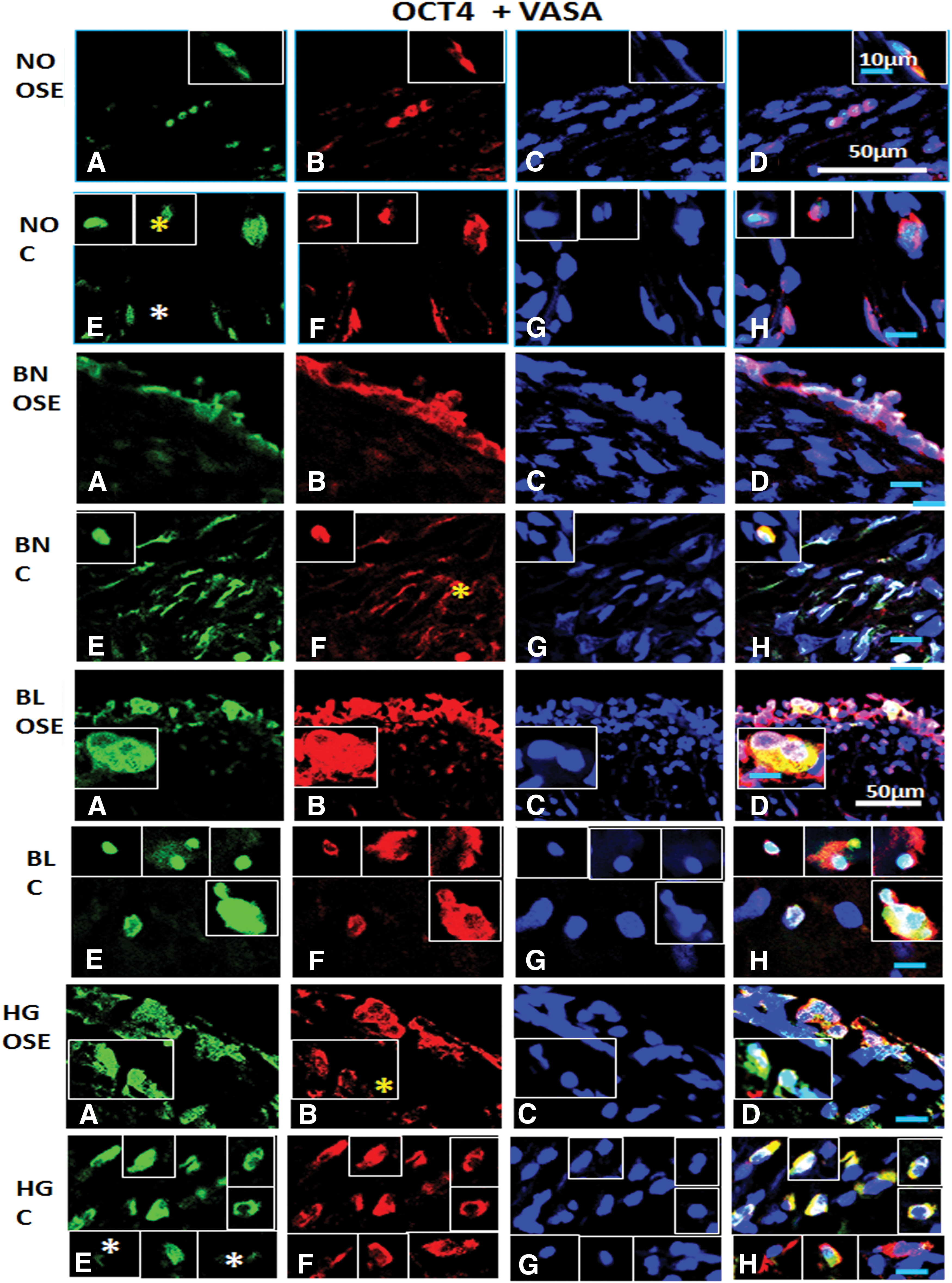
Immunofluorescence staining for OCT4 and VASA/DDX4 from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: OCT4+/VASA+ cells are consistently observed in
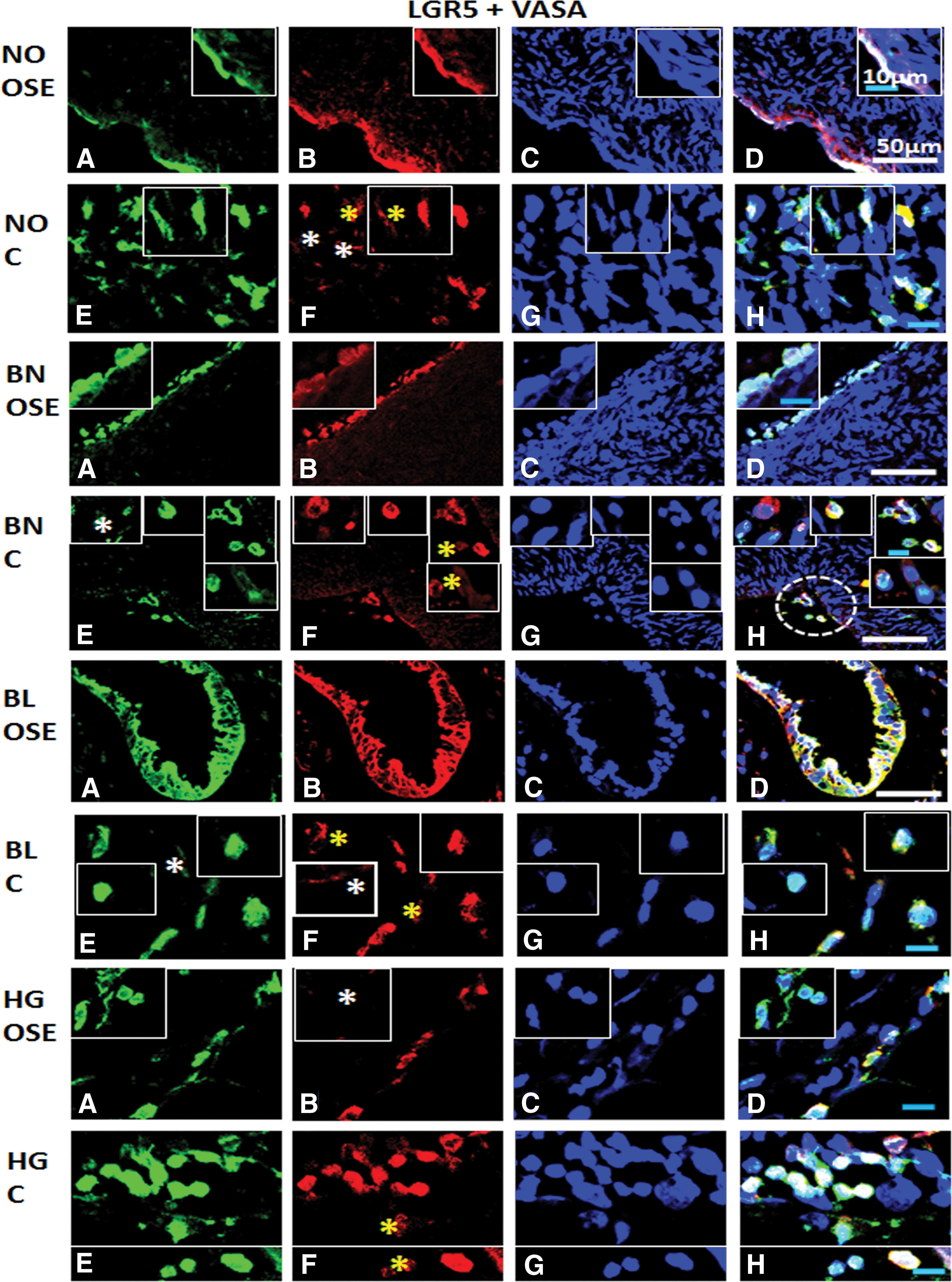
Immunofluorescence staining for LGR5 and VASA/DDX4 from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: Panel comprises of
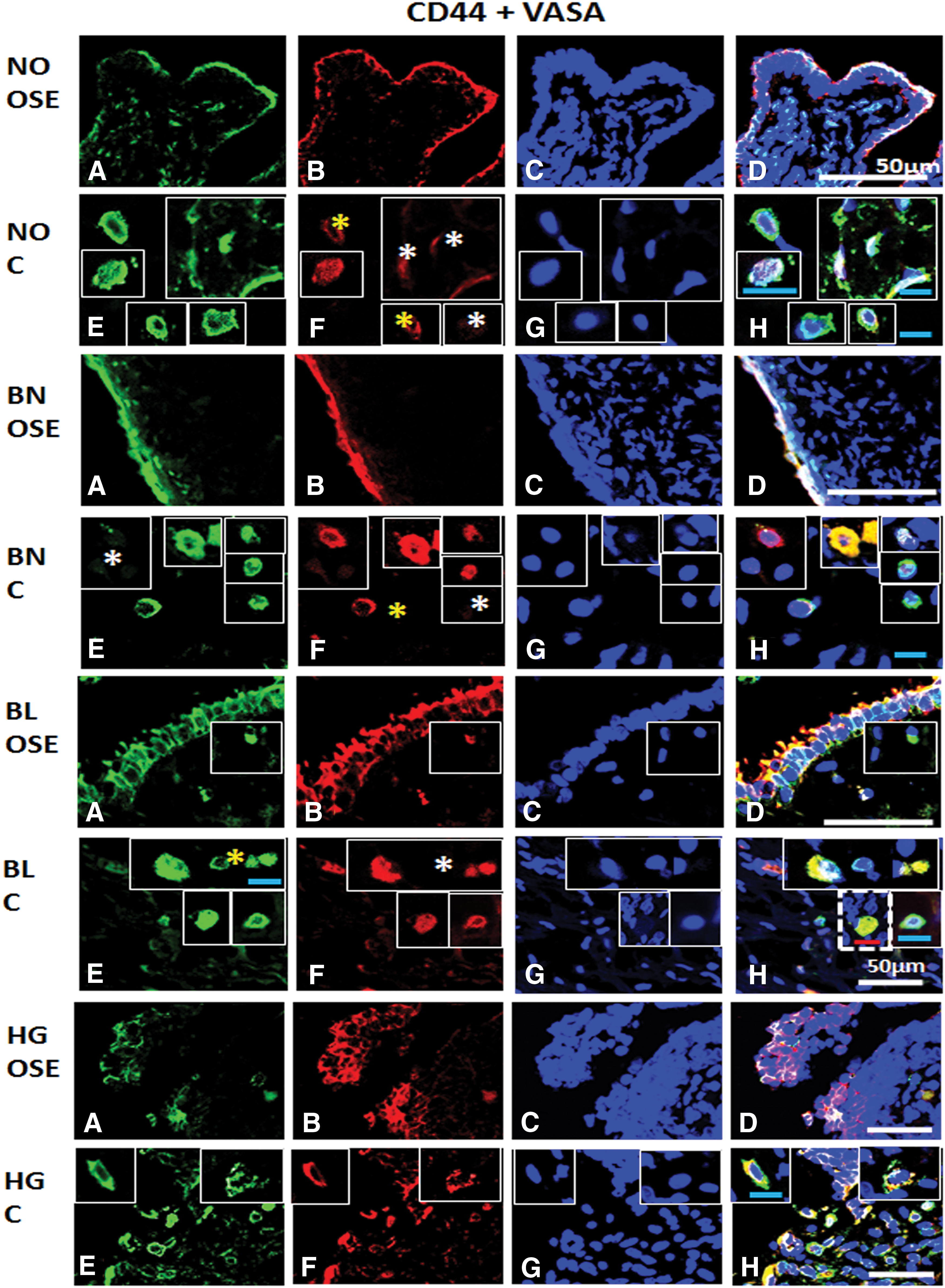
Immunofluorescence staining for CD44 and VASA/DDX4 from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: Panel comprises
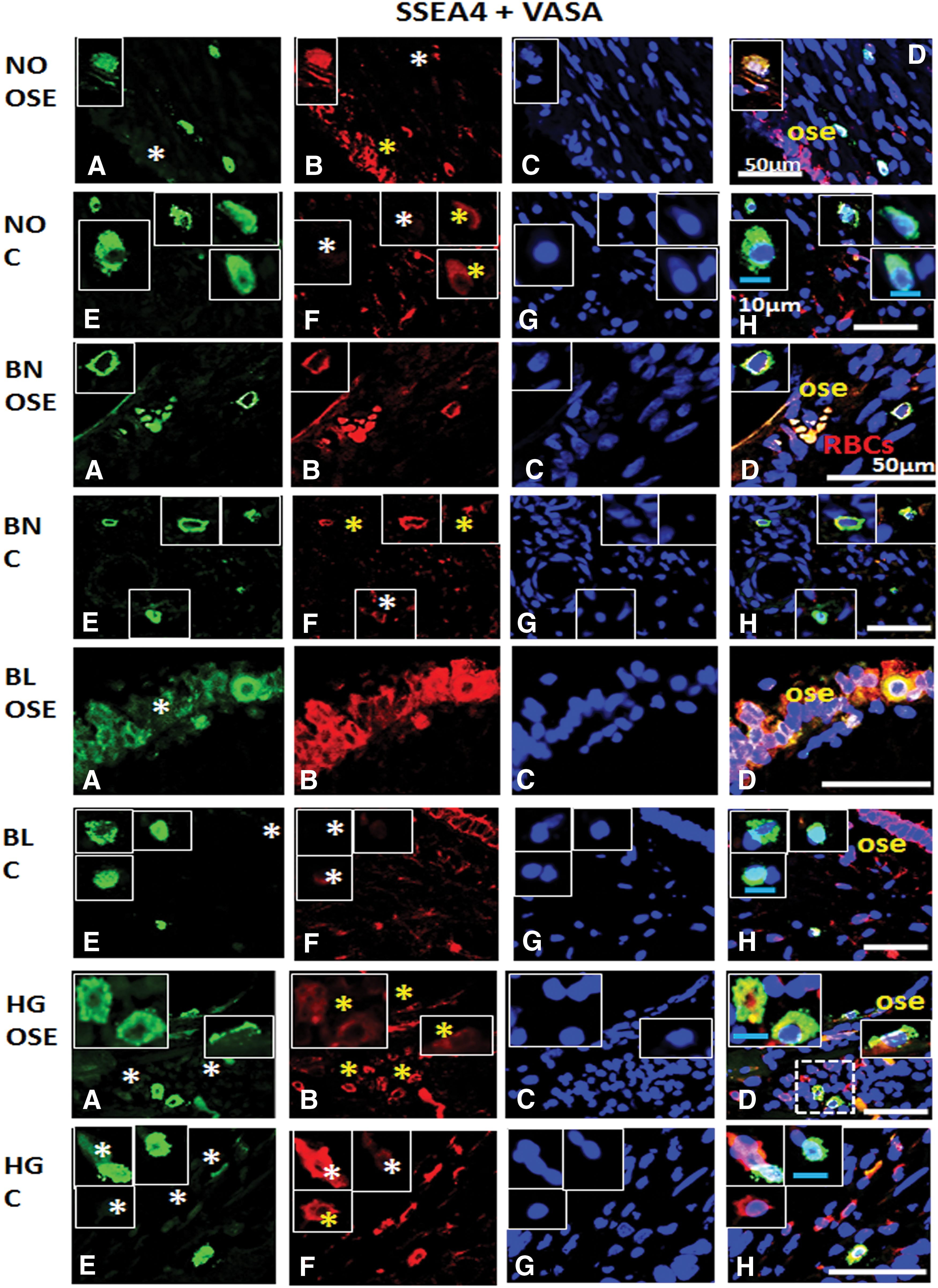
Immunofluorescence staining for SSEA4 and VASA/DDX4 from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: Panel comprises
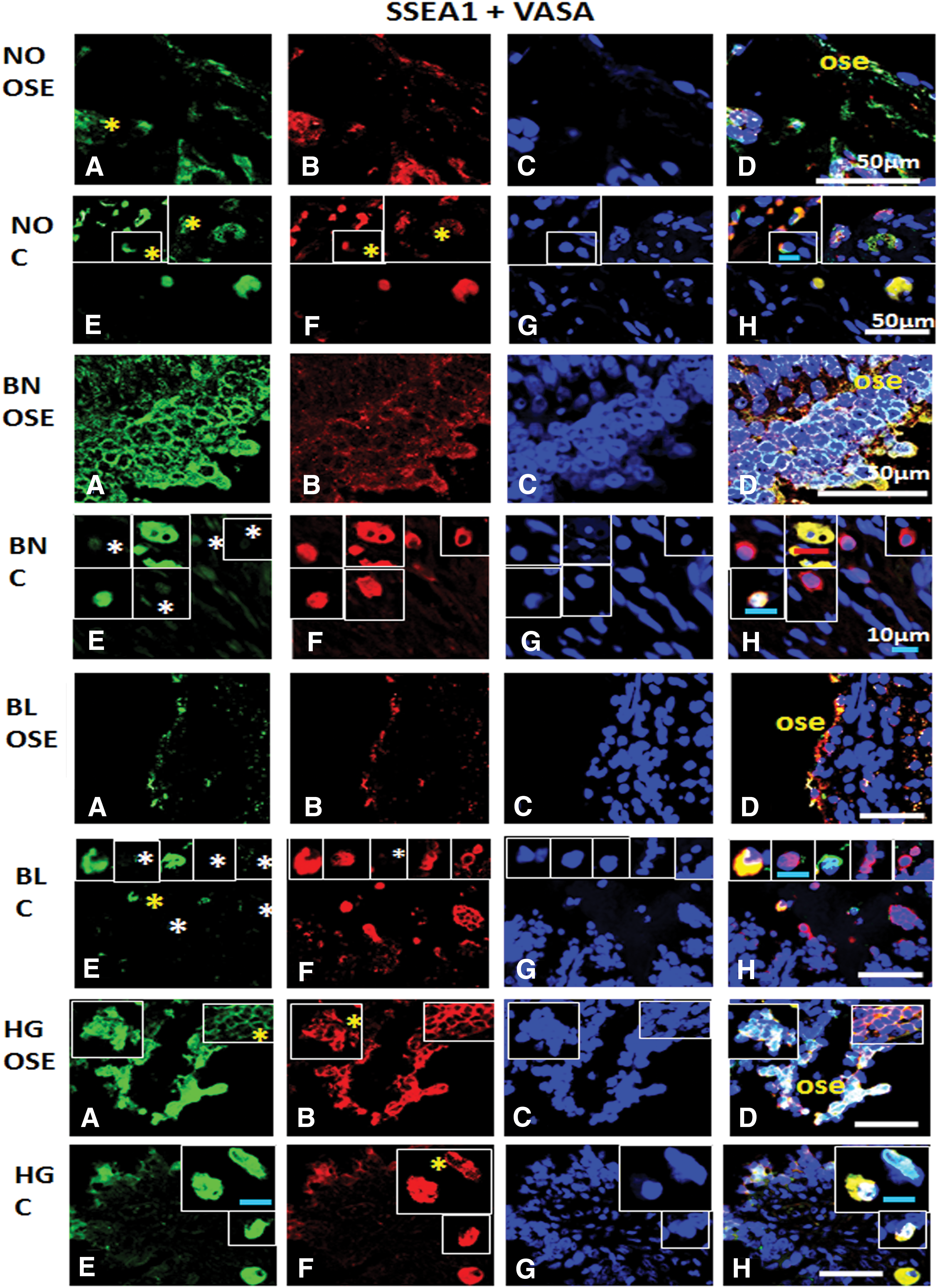
Immunofluorescence staining for SSEA1 and VASA/DDX4 from ovarian tissue sections in normal (NO), benign (BN), borderline (BL), and high grade (HG) cancer: Panel comprises
Characterization of stem cells in ovaries
Immunohistochemical localization of pluripotent stem and germline stem cell markers in normal and ovarian tumor tissues
With the help of specific antibodies (details in Table 2) various pluripotent stem cell (OCT4, SSEA1, and SSEA4) and germline (VASA) specific proteins were localized within the normal ovaries and their expression levels were compared in BN, BL, and HG tumors (Figs. 2 –5). Against a blank background, antibody-specific signals were acquired in all the tissue sections examined in target (OSE and cortex) regions within the normal ovary and tumor tissues. Individual cells in OSE layer, multilayered OSE cells, single cells, and/or clusters of cells in tumors were observed. Typically small spherical cells (∼5 μm diameter) identical to the ones reported as ovarian stem cells in earlier reports [2,3,19,21,22,36] were identified on the basis of their cell size and marker expression in or near the OSE layer in all samples and more prominently in the cortex region in tumor tissue sections. Another cell population with elongated/elliptical shape (∼7–10 μm diameter) was abundantly observed across normal tissue, as well as tumor tissue sections. In addition, cells in proximity to the adipose cells specifically in HG were found to express SSEA4 and VASA markers (observations not shown).
Immunofluorescence staining and confocal microscopy of ovarian tissues
Various antibodies specific for pluripotent stem, cancer stem, and germline stem cells (Table 2) were tested for their expression in NO, BN, BL, and HG ovarian tumor tissues (Figs. 6 –9). OCT4 was localized in majority of the cells in OSE and specific regions within the cortex, while BL and HG OSE revealed multilayered NANOG+ cells. Cytoplasmic NANOG+ cells within cortex were observed more often. Versatile populations of OCT4+ and NANOG+ cells with nuclear and cytoplasmic signals denote primitive stem cells and their differentiated progenitors as explained earlier [3]. Similarly, SSEA4+ cells were found to be equally distributed within the OSE layer, as well as the cortex in NO, BN, and BL, while in HG they were moreover prevalent in the cortex region. Elongated/elliptical shaped cells (∼7–10 μm diameter) were abundantly observed across the tissues. VASA+ cells slightly spherical and elongated in appearance were present in both OSE layer and cortex regions across NO, BN, BL, and HG. Of all samples BN and HG cortex composed of more VASA+ cells distributed throughout the tissue sections, where BN cortex reflected larger round and elliptical fluffy-appearing (>10 μm diameter) cells. NO and BL contained sparsely distributed VASA+ cells visible in each field of focus. Multilayered OSE cells in BL and HG cortex samples exhibited clusters of VASA+ cells, while elliptical/spindle shaped VASA+ cells in the NO cortex and slightly spherical VASA+ cells in BN, BL, and HG cortex were visible (Figs. 6 –9).
Colocalization of markers
Colocalization of germline stem cell marker (VASA) with pluripotent stem cell (OCT4, SSEA4, and SSEA1) and CSC (LGR5 and CD44) markers was analyzed by confocal microscopy (Figs. 10 –14) and summarized in Table 4.
+, positive expression; −, negative/nil expression; BN, benign tumor; BL, borderline tumor; C44, CD44; HG, high grade tumor; low, low levels of marker expression below normal expression; NO, normal ovary; OSE, ovarian surface epithelium; L, LGR5; O, OCT4; S1, SSEA1; S4, SSEA4; V, VASA.
Normal ovary
Pluripotent stem cell markers Oct4 and SSEA4 consistently reveal dual positive cells in OSE and cortex. Cortex revealed some germline cells with low or undetectable Oct4 expression. Some SSEA4+ cells with loss of germline marker and only VASA+ cells were observed in OSE layer. SSEA1+ cells revealed faint expression in cortex and gradual loss of germline features.
BN ovary
Either OCT4+ or VASA+ cells, as well as dual positive cells, were observed in both OSE and cortex. SSEA4+ cells with gradual loss of VASA expression were evident. VASA+ cells lacked SSEA1 expression, while dual positive cells were observed in both OSE and cortex regions.
BL ovary
Dual positive cells (OCT4 and VASA) in both OSE and cortex were visualized. Cells with more germline characteristics were present in the OSE layer rather than the cortex region. Individual marker positive and dual marker positive cells were located in the cortex region. SSEA1/VASA expressing cells revealed prominent dynamics in the cortex region.
HG ovary
Germline and pluripotent stem cell markers were present in both OSE and cortex regions. SSEA4+ cells revealed lesser germline potential, and dual positive cells were present in the cortex. Some SSEA1+ cells revealed low VASA expression in cortex, and most of the cells were dual positive.
OCT4 and VASA expression
Dual positive cells in NO signify both pluripotent stem and germline stem cell characteristics in both OSE and cortex regions, while three subpopulations showing each individual marker and combination of both in BN were observed. BL and HG probably reflected subpopulations with combination of both markers in OSE and cortex regions (Fig. 10). Most of the cells revealed both pluripotent stem cell (OCT4) and germline stem cell (VASA) markers across all samples, that is, NO, BN, BL, and HG, and highest number was observed in HG compared to NO. Some subset of cells expressed OCT4 and VASA independently. However, some subsets of cells showed high expression of both germline and stem cell specific markers.
LGR5 and VASA expression
Majority of cells showed pluripotent and germline stem cell characteristics both in NO OSE and cortex regions, while few cells with low expression of both markers reflected heterogeneity in normal ovaries. In BN, cells with pluripotent stem cell characteristics revealed low expression of germline features and vice versa, while third population of cells exhibited both cell type features. Similar populations of cells were observed in both OSE and cortex regions reflecting their dynamism. A clear hierarchy of subpopulations was visible in BL where pluripotent stem cells showed low to undetectable levels of expression of germline marker with subsequent acquisition of both types of markers. HG OSE reflected more dynamic subpopulations of cells compared to the cortex of BN and BL. Similar combination of cell populations persisted in both OSE and cortex, while individual marker positive cells were present in HG OSE. LGR5+ cells showed acquisition of germline characteristics (from low to normal/detectable levels of expression). In cortex, LGR5+ cells seemed to acquire GSC characteristics (Fig. 11). LGR5+ stem cells almost coincided with germline stem cell populations. Versatility in terms of expression of both markers was evident in the cortex region of BN, BL, and HG and OSE layer of HG stage. Similarly high proliferation index was observed in HG ovarian cancer compared to other stages of ovarian tumor and normal ovaries, suggesting a low number of proliferating (stem) cells.
CD44 and VASA expression
OSE of NO, BN, BL, and HG showed both cell type characteristics. Cortex of NO and BN shows similar characteristics, that is, CD44+ cells showed gradual acquisition of GSC characteristics and finally cell populations exhibiting expression of both markers. BL cortex shows relatively more dynamic subpopulations (CD44 low→+; VASA− →+). Besides, large, round, fluffy-appearing dual positive cells were also observed. Clearly dual positive cells in HG were visible within the cortex (Fig. 12). CD44 expression marks highly proliferating cells abundant in both OSE layer and cortex regions, while CD44+/VASA+ cells with variable expression of VASA but consistent CD44 expression were noticed within the cortex region of NO, BN, and BL. As expected, HG OSE and cortex both revealed highly proliferating dual positive cells. NO OSE and cortex also revealed CD44+/VASA+ cells but their numbers being low compared to tumor tissues.
BL with low malignant potential tumors reveals typical populations of CD44+ and LGR5+ cells, which failed to coexpress VASA independently signifying highly proliferating pluripotent cancer (stem) cells. However it remains to be determined whether the CD44+ and LGR5+ cells belong to similar subset of cells and whether they express germline markers at any point of development and/or differentiation. Further analysis is warranted to understand the complex translineage differentiation of CSCs, which impart them dynamic capabilities of sustaining tumorigenesis while resisting oncotherapy by hijacking signaling pathways of normal stem cells and posing a serious hurdle to eradicate them.
SSEA4 and VASA expression
Dual positive cells were present in NO OSE and cortex, while few VASA+ cells suggested the presence of germline stem cells in OSE. BN OSE revealed presence of SSEA4+/VASA+ (dual positive) cells with low expression of markers. BN cortex revealed presence of more pluripotent stem cells rather than germ lineage specific cells. Dual positive and VASA+ germline cells were observed in BL OSE, whereas in BL cortex, SSEA4+ cells seem to slowly acquire germline characteristics or vice versa. HG OSE revealed presence of more SSEA4+ pluripotent stem cells with simultaneous expression of VASA− and VASAlow cells. Two distinct populations exhibiting pluripotent stem and germline characteristics individually were observed in HG cortex region, while gradual acquisition of the other marker type was typically visible in these cell subpopulations (Fig. 13).
SSEA1 and VASA expression
NO OSE harbored SSEA1+ cells showing VASAlow and VASA+ cells—that is, slow acquisition of germline stem cell characteristics. Similar to OSE, cell hierarchy was observed in NO cortex as well. Dual positive cells were observed. In BN cortex SSEA1− cells showed gradual acquisition of germline characteristics and more cells were abundant in the cortex region of BL. Also individual marker positive cells with gradual acquisition of the other marker (SSEA1/VASA) were clearly detected in BL cortex. In addition, BN, BL (low expression), and HG OSE revealed dual positive cells (SSEA1+VASA). HG cortex revealed VASA+ cells with low expression of SSEA1, and another subpopulation with vice versa expression was prominently observed (Fig. 14).
Based on our results, we propose that some intermediate stages of cells revealing other combination of markers in terms of low, high, or undetectable expression might be present. But these may remain undetected due to low cell numbers, sensitivity to antibody used, and its cross-reactivity. Whether heterogeneity/hierarchy of cells persists, whether independent subpopulations exist, or whether process of differentiation and/or dedifferentiation occurs is not well understood. Further precise experiments targeted for lineage tracing of individual subpopulation will lead to more relevant/sound information and improve our understanding of how these subpopulations exist independently, coexist, interact with each other, and bring about the dynamic cell–cell interactions eventually leading to a complicated network/scenario, which we witness only superficially within the limitations of our scientific knowledge and approach.
Discussion
OGSCs (5–8 μm in size by labelling with anti-VASA/DDX4 primary antibody) have been demonstrated successfully and differentiated to produce female gametes in vivo in mice and in vitro in human [1]. Similarly, rare but distinct pluripotent stem cell populations within the OSE layer (comparable to VSELs) in the OSE scrapings and ovarian cortical cultures of menopausal human, as well as other adult mammalian ovarian tissues, were isolated with the aid of different strategies. These cells undergo in vitro differentiation into oocytes (germ cells) [2,3,19,21,22]. A tiny rare, but novel, population of pluripotent stem cells with embryonic characteristics (expressing CXCR4, OCT4, SSEA4, Sca-1, NANOG, and LIN−/CD45−) named VSELs was first reported within the adult bone marrow, cord blood, epidermis, heart, pancreas, testis, bronchial epithelium, etc. [37]. These cells are hypothesized to be deposited within adult tissues and organs during gastrulation whereby they undergo deviation from their default migratory route to the presumptive gonads and hence express epiblast and germ cell specific markers [23,38].
Similarly, a potential germline origin of the tiny pluripotent stem cells in human OSE layer (with embryonic characteristics) is also speculated [26]. The investigators arrived at the conclusion due to their previous observations of SSEA4+ cells (4 μm in size, isolated by both magnetic-activated cell sorting and fluorescence-activated cell sorting methods) from OSE layer of normal ovaries from females of reproductive, as well as menopausal, age [36]. These cells exclusively expressed PGC [PRDM1, PRDM14, DPPA3 (STELLA) DAZL] and embryonic stem (ES) (OCT4A, SOX2, NANOG, TERT, SALL4, CDH1, and LEFTY1) cell specific markers. Similarly, NANOG+ cells/structures with unknown biological and clinical significance were also reported within BL ovarian cortex region by same group [39]. In another similar study same authors investigated pluripotency (OCT4, SOX2, NANOG, and SSEA4) and germinal lineage (VASA/DDX4 and DPPA3/STELLA) related markers in tiny spherical cells (5 μm) found in close vicinity of borderline ovarian tumor cell cultures. Genes concerning regulation of transcription, cell signaling, cell cycle, apoptosis, secretion, phosphorylation, and cell junction were found to be differentially regulated thus suggesting these CSCs with tumor-initiation potential [40]. These authors also detected small putative stem cells (5 μm in size) in human ovarian cancer tissues expressing NANOG, SOX2, and SSEA4 besides normal healthy ovaries, which assumed cluster-like formation in cancerous OSE layers. In addition cells also expressed epithelial mesenchymal transition (EMT) marker Vimentin. Besides NANOG+, VIMENTIN+, SSEA4+, and SOX2+ cells (10–15 μm in size) with large nuclei appeared to be sloughed off from the OSE layer. Recently, these cells were typically observed in association with EMT and morphologically transformed OSE cells, implicating as CSCs [41].
Pluripotent stem cells, including VSELs, residing in various adult tissues and organs and in human menopausal ovaries have been proposed. Recently, these have been proposed as cancer initiating cells [29]. Typically high telomerase activity, high cell proliferation and migratory properties, immortalization, invasion, lack of cell adhesion properties, demethylation, immune evasion, and marker expression are common cellular characteristics found in both PGCs and cancer cells [42]. This unique link would possibly explain the century old hypothesis of Embryonic Rest/Germ line origin of cancer development put forth by Rudolf Virchow, Julius Conheim, and John Beard [27].
The possible reasons for origin of cancer cells are linked to perturb genetic, epigenetic, and proteomic profiles of these putative CSCs, their faulty intercellular cross talk, acquisition of unique methylation patterns, and fusion with other somatic cells [15]. It was recently speculated that presence of cancer cell subpopulations (including CSCs) located in specified niches within a single tumor could be attributed to variation in expression of genes/markers due to molecular heterogeneity, cellular adaptations, and surrounding microenvironmental influences as per Darwinian dynamics of evolution [43].
The suggested origin of CSCs arising from reprogrammed progenitors according to the CSC hypothesis together with the observation of pluripotent stem cells in normal, as well as tumor, tissue lays down the foundation for our quest for identifying predominant subpopulations of cells in OSE and ovarian cortex responsible for tumor initiation. To accomplish this, we investigated a panel of markers specific for pluripotent stem and germline specific markers by RT-PCR (Fig. 1) and immunolocalization by immunohistochemistry (Figs. 2 –5), as well as confocal fluorescence microscopy (Figs. 6 –14) procedures to confirm mRNA and protein expression while comparing tumor tissue with normal ovaries.
Various patient samples (normal ovaries, as well as BN, BL, and HG tumor tissues) revealed transcripts for pluripotent stem cells (Oct4, Nanog, and Sox2) and germline stem (Vasa and Fragilis) and CSC (Lgr5). Considering the rarity of the cells contributing to all the transcripts detected (Fig. 1), a quantitative estimate of expression or comparison across various samples and across different cancer stages was not possible, thus demanding enriched populations for further studies. Pluripotent ES cell markers such as OCT4, SOX2, and NANOG are crucial factors for malignancy and provide evidence for CSCs operating as dynamic entities while maintaining equilibrium between CSCs and cancer progenitor cells [44].
Using IHC and immunofluorescence techniques, characteristic expression of pluripotent markers (OCT4 and NANOG) (Figs. 2, 6, and 7) in nucleus, as well cytoplasm, was observed in most samples. Typical cytoplasmic localization of pluripotent marker OCT4 has been reported earlier in lineage committed progenitor stem cells in both male [45] and female [3] gonads. Similarly, Gkountela et al. [46] reported that localization of OCT4A protein within the cytoplasm instead of the classic “nucleus” of cKIT+ human PGCs during fetal development suggests cellular heterogeneity. Present study results demonstrate a hierarchy of PGC subpopulation and complex biology with regards to the pluripotency and acquisition of germline characteristics. In context of cancer, Baillie et al. [47] reported cytoplasmic localization of OCT4, SALL4, SOX2, and NANOG in oral tongue squamous cell carcinoma consistent with other investigators [48] in glioma cells where nuclear and cytoplasmic expression of OCT4 and NANOG was reported whose significance remains unknown at present. Santaliz-Ruiz et al. [49] reported cytoplasmic expression of NANOG in ovarian cancer similar to our observations as reported in Fig. 7. A possible role of this unconventional expression and significance of differentiated progenitors detected with respect to normal and tumor physiology needs to be investigated independently.
Coexpression of LIN28+/OCT4+ proteins in ovarian cancer [50], OCT4+/NANOG+ in lung adenocarcinoma [51], and OCT4+, c-Myc+/NANOG+ cells in ovarian tumors [52] signifies presence of various CSC populations. Cell hierarchy of breast cancer cells based on phenotype and functions was recently indicated on the basis of heterogeneous cell populations both in vitro and in vivo [53]. A thorough characterization of pluripotent stem, cancer stem, and germline stem cell markers in ovarian cancer across various stages of tumorigenesis has never been reported, and the current study is by far the first to describe this systematically.
Other markers such as surface glycoproteins SSEA1 and SSEA4 are implicated not only in embryonic cells, ES cells, germ cells, etc. but also their expression is reported in other types of cancers, and possible prognostic role is suggested [3,35,54 –56] apart from few studies in ovarian cancer [39 –41]. With limited studies reported so far, the expression pattern, their importance, and implications in ovarian cancer are understudied. Besides VASA, another germ cell marker IFITM3/FRAGILIS was utilized to isolate FGSCs [57], while few studies suggesting its versatile role in breast, colorectal, and gastric cancer in addition to endometriosis [58 –61] warrant further investigation. Similarly VASA expression was demonstrated not only in mice and human OGSCs [1,21], migratory PGCs [30], and multipotent and proliferating cells [62] but also rare studies suggesting its role as CSCs as well [32,63] are reported. In addition coexpression of CD133 and VASA and increased expression of both in ovarian cancer attribute a CSC identity to VASA [63]. Next, the adult stem cell marker LGR5 reported in intestinal crypt, stomach, hair follicle bulge, eye, mammary gland, in mice embryonic gut, skin, kidney, etc. [64 –66] is strongly implicated in ovarian epithelial biology [67] and it is speculated that neoplastic transformation of these LGR5+ stem cells would lead to ovarian cancer [7,68]. CD44 is known to regulate key processes like cell adhesion, migration, tight junction assembly, recirculation and homing of lymphocytes, hematopoiesis, and angiogenesis [69] and was expressed with pluripotent marker SSEA4 [70] and associated with highly metastatic tumors and poor prognosis in patients [71].
In our studies, we report for the first time that pluripotent stem cell markers OCT4 and SSEA4, as well as embryonic and germ cell marker SSEA1, implicated in ES cells and PGCs also coexisted with the VASA+ germline stem cell subsets of the ovary. Interestingly, LGR5, a CSC marker and characteristic of highly proliferating adult stem cells, coexpressed with germline stem cell marker VASA originally utilized for characterizing ovarian stem cell populations. Of key note was the prominent coexpression of CSC marker CD44 with VASA within the OSE, as well as cortex, regions and possesses no known germline connection. Careful and unbiased analysis of immunostaining pattern suggests that CD44+ cells slowly acquired germline characteristics, and VASA expression could be interpreted as differentiation of progenitor cells in tumor tissues. Upon examining normal ovaries, presence of VASA+ cells reveals presence of germline stem cells and hence alternately loss of VASA rather than gain of its expression would better explain the CD44+ cell profile in OSE and cortex regions. This information collectively came across as excellent novel data that has never been explored to date. Primarily establishing and defining a true identity for the various stem cell subsets/populations not only in normal ovarian tissue but also in various stages of ovarian tumor will be extremely helpful to define the role of CSCs in tumorigenesis.
Coexpression of pluripotent stem, CSCs, and adult stem cell markers individually with VASA in the same cell at a given point of time when the tumor tissue and NO were dissected from the human subjects denotes probable existence of interconvertible cell populations either by differentiation or dedifferentiation of specific clones. Similar cellular dynamics and reciprocal conversion of PGCs and pluripotent stem cells have been defined in normal physiological conditions [72] while that in tumor state require further evaluation. In fetal germ cells an intricate balance between the repressions of pluripotency related mechanism concerning the germ cell differentiation is essentially maintained in normal scenario; however, a lapse in this mechanism explains the possible propensity of germ cells toward tumor formation instead of germ lineage [73]. Differential expression of markers within the tumor cells may be explained by the differentiation state of cancer cells constituting the tumor [74]. Besides in vivo tumor, ovarian cell line based study also indicated a versatile fluctuating population of cancer cells with acquisition of stem cell phenotype and development of metastatic potential [75]. Existence of these hierarchical (sub) populations probably is meant to serve a specific function as per the microenvironmental cues from surrounding tumor cells and niche components and a bidirectional complex cross talk supporting cellular heterogeneity. However migration of stem cell populations with similar characteristics from extra-ovarian tissue like oviduct, fimbriae, and fallopian tube remains undetermined in current study and can be researched on similar lines further to address this possibility.
Conclusion
In view of the current understanding of CSC biology to date, our study provides novel and important information regarding pluripotent stem cell populations existing in normal ovarian tissue and persisting in various stages of ovarian tumor, which could be implicated in the possible origin of cancer. Novel insights into CSC biology with respect to ovarian stem cells with germline perspective were obtained. Unique stage and location specific information about the molecular signatures and distribution pattern within the ovarian tissue may enable identification of specific predominant tumor initiating CSC population. This understanding would thus improve their efficient targeting and aid in devising strategies to prevent their dissemination to distant organs causing fatal relapse. Information regarding pluripotent stem cells in cancer stages will provide important leads to understand their downstream signaling as candidates for predicting patient survival, as well as for testing and developing therapeutics to treat ovarian cancer more effectively. Stage specific characterization of CSCs may be helpful in developing precision medicine and/or personalized medicine to treat ovarian cancer at specific stage of development and differentiation and probably eliminate the possibility of dissemination of CSCs to distant organs causing relapse and mortality.
Footnotes
Acknowledgments
The authors acknowledge the technical assistance and insights provided by Dr. Colin Rogers, bio-imaging expert, NIKON for confocal microscopy imaging and the help rendered by trainee student Mr. Ankit Kanotra in compiling some part of literature and for processing tissue slides during immunolocalization experiments under the supervision of S.P. Authors also thank Dr. V.R. Jala for technical help with the confocal microscope. This work was supported by grants from NIH NCI CA124630 (S.S.K.) and U01CA185148 (S.K.B.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.