
Abstract
Lysophosphatidic acid (LPA) is a membrane-derived lysophospholipid that exists in the plasma and platelets. It exerts its functions through activation of various LPA receptors (LPARs), which belong to the family of G protein-coupled receptors. Activation of LPARs has important roles in stem cell differentiation. However, how LPA affects human hematopoietic stem cell (HSC) differentiation remains elusive. In our previous studies, we have suggested that LPA receptor 2 (LPA2) and LPA receptor 3 (LPA3) play opposing roles and may act as a molecular switch during megakaryocytic differentiation in K562 cells. In this study, human CD34+ HSCs and zebrafish are adopted to investigate the roles of LPA3 during megakaryopoiesis/thrombopoiesis in vitro and in vivo. Our results show that LPAR3 mRNA expression level is decreased upon induction by thrombopoietin and stem cell factor in human HSCs. Using pharmacological activators and shRNA knockdown experiments, we demonstrate that activation of LPA3 inhibits megakaryopoiesis in human HSCs. In addition, pharmacological activation of LPA3 suppressed thrombopoiesis in zebrafish. Furthermore, blockage of LPA3 translation by morpholino increased the number of CD41-GFP+ cells in Tg(CD41:eGFP) zebrafish. Moreover, the mRNA expression level of zCD41 increased significantly in LPA3-knockout zebrafish. These results clarify the negative role of LPA3 during megakaryopoiesis and provide important information for potential treatments of related diseases, such as megakaryopenia.
Introduction
M
Lysophosphatidic acid (LPA) is a small glycerophospholipid derived from cell membrane phospholipids by metabolism of autotaxin [6]. To date, at least six G protein-coupled LPA receptors (LPARs) have been identified. LPA1, LPA2, and LPA3 are classified in the endothelial differentiation gene (EDG) family [7,8], whereas LPA4, LPA5, and LPA6 are categorized as non-EDG family [9]. Recent studies have indicated that autotaxin and lipid phosphate phosphatase 2A are differentially expressed in the bone marrow [10]. During early vertebrate development, the autotaxin-LPA axis has been reported to participate in hemangioblast differentiation through LPA1 activation [11]. Furthermore, gene profiling confirms the expression of LPARs in myeloid progenitors [10]. In contrast, the LPA/LPA4 axis regulates hematopoiesis indirectly through affecting stromal cell activity in bone marrow [12]. These results suggest the novel and critical roles of LPA signaling in the microenvironment during early hematopoiesis. Nevertheless, the roles of LPAR in terminal differentiation, such as erythropoiesis and megakaryopoiesis, have not yet been fully confirmed.
In our previous study, we demonstrated that LPA is involved in EPO-dependent erythrocyte differentiation by activating LPA3 and β-catenin pathways [13]. We also reported the specific roles of LPAR subtypes in three different species [14]. In the present study, we sought to investigate the functional roles of LPA3 during megakaryopoiesis/thrombopoiesis in human HSCs and zebrafish embryos. Our results demonstrate that LPA3 negatively regulates early platelet differentiation in vitro and in vivo. Furthermore, the thrombopoietic phenotype of LPA3-knockout zebrafish is also reported in this study.
Materials and Methods
Cell culture
The isolation and culture methods for human HSC were described previously [13]. HSCs were isolated from umbilical cord blood after obtaining the donor's consent according to governmental regulations (“Guidelines for collection and use of human specimens for research,” Ministry of Health and Welfare, Taiwan) approved by the Institutional Review Board of the Taoyuan General Hospital, Taiwan. In stage I, 2.5 × 105 human HSCs were cultured in 5 mL of erythrocyte development medium (EDM) (5 × 104 cells/mL) in the presence of stem cell factor (SCF) (50 ng/mL) and EPO (6 IU/mL) for 6 days. In stage II, cells were cultured in 10 mL of EDM (3 × 104 cells/mL) in the presence of SCF (50 ng/mL) and TPO (50 ng/mL) for 7 days. All cell cultures were incubated at 37°C in 5% CO2 under humidified conditions.
Treatment of agonist and antagonist
Stock solution of LPA (18:1; Sigma-Aldrich), (2S)-3-[(hydroxymercaptophosphinyl) oxy]-2-methoxypropyl ester (2S-OMPT) (Cayman Chemical), and Ki16425 (Cayman Chemical) were prepared in different solvents, respectively, as described in our previous report [15]. CD34+ HSC was cultured with the test compounds diluted in the medium with 0.005% fatty acid-free bovine serum albumin (Gibco).
Lentiviral infection
Lentiviral-shRNA expression vectors were purchased from National RNAi Core Facility Platform (Academia Sinica, Taiwan). Lentiviral stocks were produced by Lipofectamine 2000 (Invitrogen) transfection. A range of 30%–40% confluent of HEK293T cells was prepared in T75 flasks and transfected with a DNA mixture containing 7.5 μg packaged plasmids and 7.5 μg vector of shRNA for the target gene for 16 h. The transfected condition medium was replaced with 12 mL fresh DMEM containing 10 mM sodium butyrate (Sigma-Aldrich). The conditioned media were replaced and harvested 3 days after transfection. The packaged lentiviral particles in clarified conditioned media were harvested by ultracentrifugation at 25,000 rpm for 3 h at 4°C. The viral pellets were resuspended in 1 mL fresh DMEM and stored at −80°C. Human CD34+ HSCs were infected by packaged lentivirus. After 2 days infection of lentivirus, cells were treated with cytokines, LPA, and LPAR agonist/antagonist.
RNA extraction and quantitative real-time polymerase chain reaction
Total cellular RNA was extracted using the TRIzol reagent (Invitrogen). Complementary DNA was synthesized with 1 μg total RNA using a Toyobo reverse transcription (RT)-polymerase chain reaction (PCR) Kit (Toyobo). The real-time PCR with the mixture reagent KAPA SYBR Green as the fluorescent dye (Bio-Rad) was carried out on a MiniOpticon real-time detection system (Bio-Rad). Gene-specific primers were used, and the specificity was confirmed by single melting curve following the real-time PCR. Cycling conditions are 95°C for 3 min, followed by 30 cycles of 95°C for 30 s, 62°C for 30 s, and 72°C for 30 s. For quantification, the target gene was normalized against the amount of GAPDH. Primers for the real-time PCR were as follows: lpar1: forward (5′→3′): TTCAACTCTGCCATGAACCCC, reverse (3′→5′): CTAAACCAC AGAGTGGTCATT; lpar2: forward (5′→3′): ACACTTCTGGCACTGCCTCT, reverse (3′→5′): AGGCTGAGTGTGGTCTCTCG; lpar3: forward (5′→3′): TCAGCAGGAGTGACACAGGCAG, reverse (3′→5′): GGAAGTGCTTTTATTGCA GACT; CD41: forward: (5′→3′): GGGCCTTCCTCGGGAT, reverse (3′→5′): AGCACAATTGGGCTCAGCTT; CD61: forward (5′→3′): GTGACCTGAAGGA GAATCTGC, reverse (3′→5′): TTCTT CGAATCATCTGGCC-5′); zCD41: forward (5′→3′): GCAAATCTTGAGCTTTGT GATG, reverse (3′→5′): 5-GCCACTCCATAT AAATCACACTG; and zEF1α: forward (5′→3′): CGGTGACAACATGCTGGAGG, reverse (3′→5′): ACCAGTCTCCACACGA CCCA.
Flow cytometry assay
5 × 105 of human CD34+ HSCs were collected in phosphate-buffered saline and stained with 10 μL of anti-CD61-FITC antibodies and 10 μL anti-CD41-PE antibodies (BD Bioscience Pharmingen) for 30 min simultaneously. All experiments were analyzed using cyto-flow instrument (BD FACSCanto II) and FCS Express software (De Novo).
Maintenance of zebrafish
The zebrafish were maintained according to an approval from National Taiwan University Zebrafish Core Laboratory standard operating protocol for animal use. The zebrafish handling, breeding, and staging methods were performed as described previously [13,16]. Briefly, wild type (AB strain), Lpa3 +/− , and Lpa3 −/− strains were maintained at 28°C in egg water (6 g sea salt in 20 L sterilized H2O).
Knockout zebrafish line generation
Transcription activator-like effector nuclease (TALEN) gene knock system was designed by ZGene Biotech, Inc. L- and R-arm TALEN mRNA was injected with concentration 100 ng/μL, respectively, in total volume of 4 μL. The genomic DNA was purified with digesting buffer (1 mg/mL proteinase K in TE buffer) and analyzed with high-resolution melting PCR.
Morpholino oligonucleotides
The stocks of morpholino oligonucleotides (MOs) were prepared in sterile double-distilled water at a final concentration of 24 μg/mL. Four nanograms MO was injected with 5% phenol red. MO sequences were as follows: MO-zLpar3: 5′-CAGCCCTAAAACA CAGGAAGACAT-3′.
Quantification of thrombocytes in CD41:GFP transgenic zebrafish
Tg(CD41:GFP) zebrafish embryos of 24 hours postfertilization (hpf) were injected with either zLpa3 morpholino or control morpholino. After 72 h, CD41-GFP+ cells were examined by a DMIL epifluorescent microscope (Leica). The images were acquired using the EOS Utility program (Canon), and the average intensity of fluorescent was quantified by ImageJ.
Statistical analysis
All the experiments were repeated at least thrice. Significant differences were calculated using Student's t-test or one-way analysis of variance. The statistical analyses were performed using StatView software (Abacus Concept). Results are expressed as the mean ± standard deviation based on at least three independent experiments. P < 0.05 was considered statistically significant in all the tests.
Results
LPAR3 expression is negatively regulated during TPO/SCF induced megakaryopoiesis in human CD34+ HSCs
Our previous study demonstrated that pharmacological activation of LPA3 enhances erythropoiesis [13,14]. However, the role of LPA signaling in megakaryopoiesis is still unclear in human HSCs. To examine the expression patterns of LPAR during MK differentiation, a TPO/SCF-induced megakaryopoiesis model system was applied to human CD34+ HSCs. HSCs were cultured in serum-free EDM with 50 ng/mL of TPO and SCF for 7 days. At day 0, the mRNA expression level of LPAR2 mRNA is significantly fewer than LPAR1 and LPAR3 in HSCs (Supplementary Fig. S1; Supplementary Data are available online at
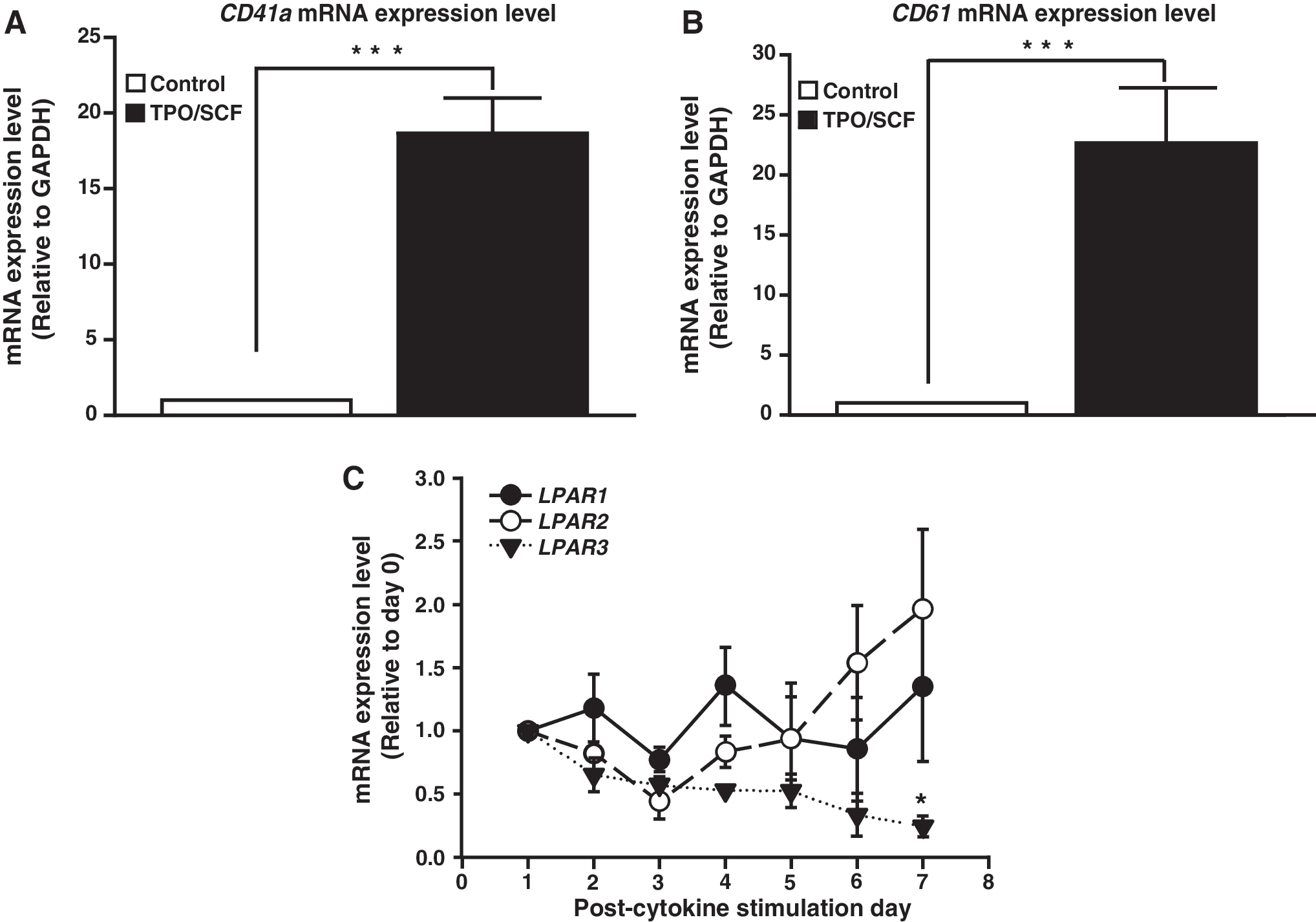
mRNA expression of LPAR under TPO/SCF induction. Human HSCs were treated with 50 ng of TPO and SCF for 7 days to induce megakaryopoiesis. Induction efficiency was quantified by mRNA expression of
LPA3 negatively regulates megakaryopoiesis
The decrease of LPAR3 mRNA expression upon HSC induction with TPO/SCF indicated a potential role in the regulation of megakaryopoiesis. We hypothesized that downregulation of LPA3 might promote MEPs to differentiate into MKs and not erythrocytes. To confirm our hypothesis, we transfected CD34+ HSCs with LPAR3 shRNA (shLPAR3) with 60% knockdown efficiency compared to control (shLacZ, Supplementary Fig. S2). Upon cytokine induction, mRNA expression level of CD41 and CD61 increased significantly. Knockdown of LPAR3 remarkably increased the mRNA expression of CD41 and CD61, indicating that knockdown of LPA3 promotes MK differentiation (Fig. 2). Moreover, LPA treatment suppressed the LPAR3-knockdown induced increase in mRNA expression of CD61 mRNA. These results suggest that LPA might repress megakaryocytic differentiation through activation of LPA3.
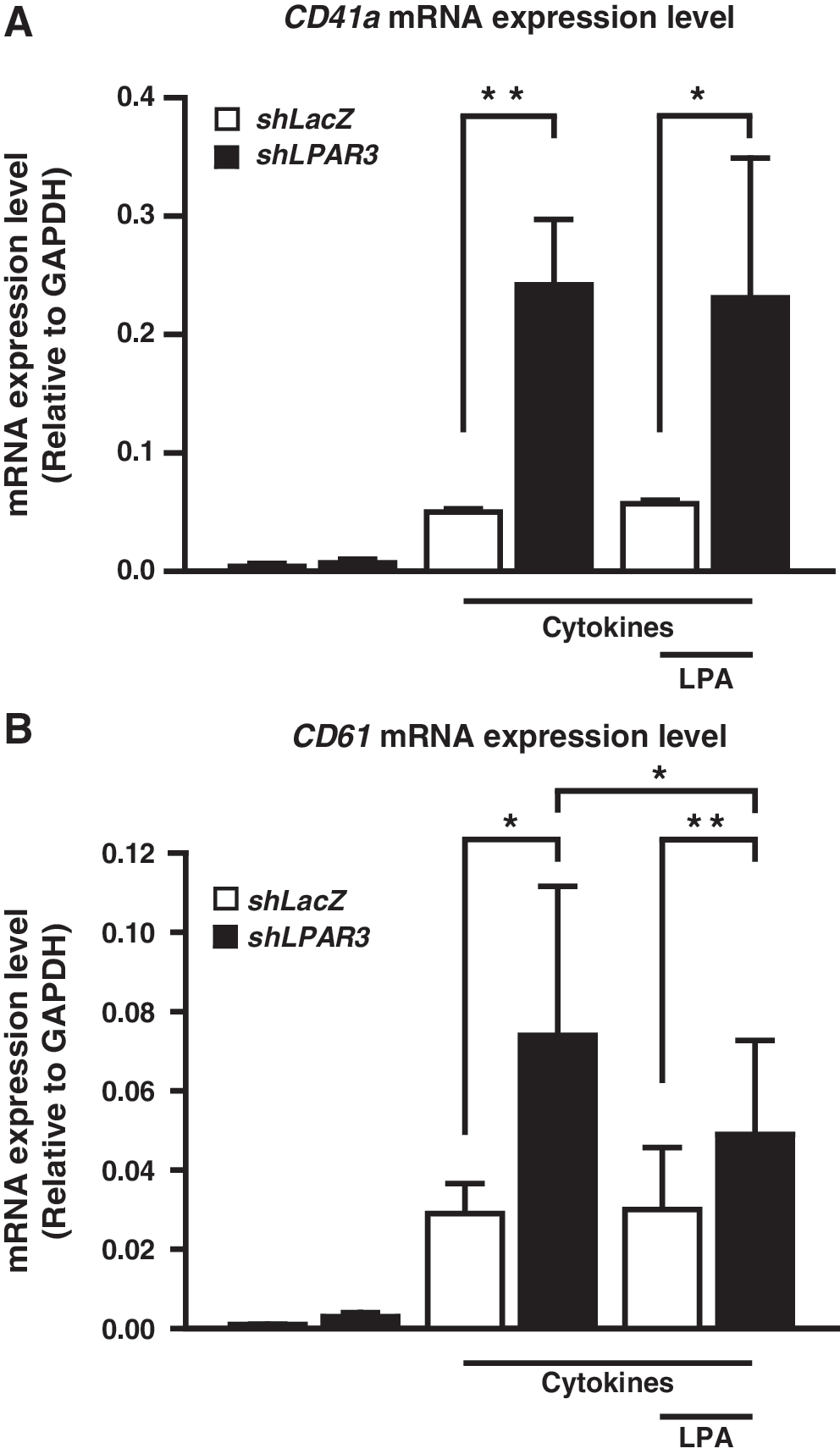
Knockdown of LPAR3 promotes platelet differentiation in HSCs. CD34+ HSCs were cultured with 50 ng/mL of TPO/SCF and treated with 5 μM of LPA. HSCs were simultaneously infected with LacZ shRNA (shLacZ, indicated as white bars) and LPAR3 shRNA (shLPAR3, indicated as black bars). The mRNA expression level of
Pharmacological activation and blockage of LPAR signaling affects TPO/SCF induced megakaryopoiesis in CD34+ HSCs
We sought to confirm the findings of our knockdown experiments using pharmacological activators and inhibitors targeting LPARs. Human CD34+ HSCs were stimulated with Ki16425, an LPA1/3 selective antagonist. Flow cytometry analysis showed that single treatment of HSCs with LPA inhibited CD41 and CD61 protein expression (double positive area), whereas treatment with Ki16425 rescued the inhibitory effects caused by LPA (Fig. 3). Furthermore, megakaryopoiesis was significantly inhibited by treatment with 2S-OMPT, an LPA3 selective agonist (Fig. 4). Since LPA1 expression is unchanged during early platelet differentiation, these findings strongly suggest that activation of LPA3, not LPA1, inhibits megakaryopoiesis.

Pharmacological manipulation of LPA3 in CD34+ HSCs. Isolated HSCs were cultured with TPO/SCF induction medium for 7 days. Cells were simultaneously treated with methanol/chloroform (M/C, solvent of LPA) and DMSO (solvent of Ki16425), 5 μM of LPA, and 20 μM of Ki16425. Fluorescence intensity greater than 103 was defined as positive area.

Pharmacological activation with 2S-OMPT in CD34+ HSCs. Isolated HSCs were cultured with TPO/SCF induction medium for 7 days. Cells were treated with DMSO and 500 nM, 1 μM, or 5 μM of 2S-OMPT. Fluorescence intensity greater than 103 was defined as positive area.
Pharmacological manipulation of LPA3 signaling affects thrombopoiesis in vivo
Previous studies have shown that LPA treatment enhances erythrocyte differentiation by activating LPA3 [13]. Since LPA3 is present at all developmental stages [18,19], it might play important roles during early embryogenesis in zebrafish. To investigate whether LPA3 regulates thrombocyte differentiation, we applied Ki16425 to repress the signals of LPA1 and LPA3. Since zCD41 mRNA expression was first detected at 42 hpf in zebrafish [20], we exposed embryos to Ki16425 for 48 h. The expression level of zCD41 was significantly increased following treatment with Ki16425 (Fig. 5A). Activation of LPA3 by 2S-OMPT inhibited thrombopoiesis (Fig. 5B). Consistent to what we found in human CD34+ HSCs, these results suggest that LPA3 may participate in the LPA-dependent thrombopoiesis in zebrafish.
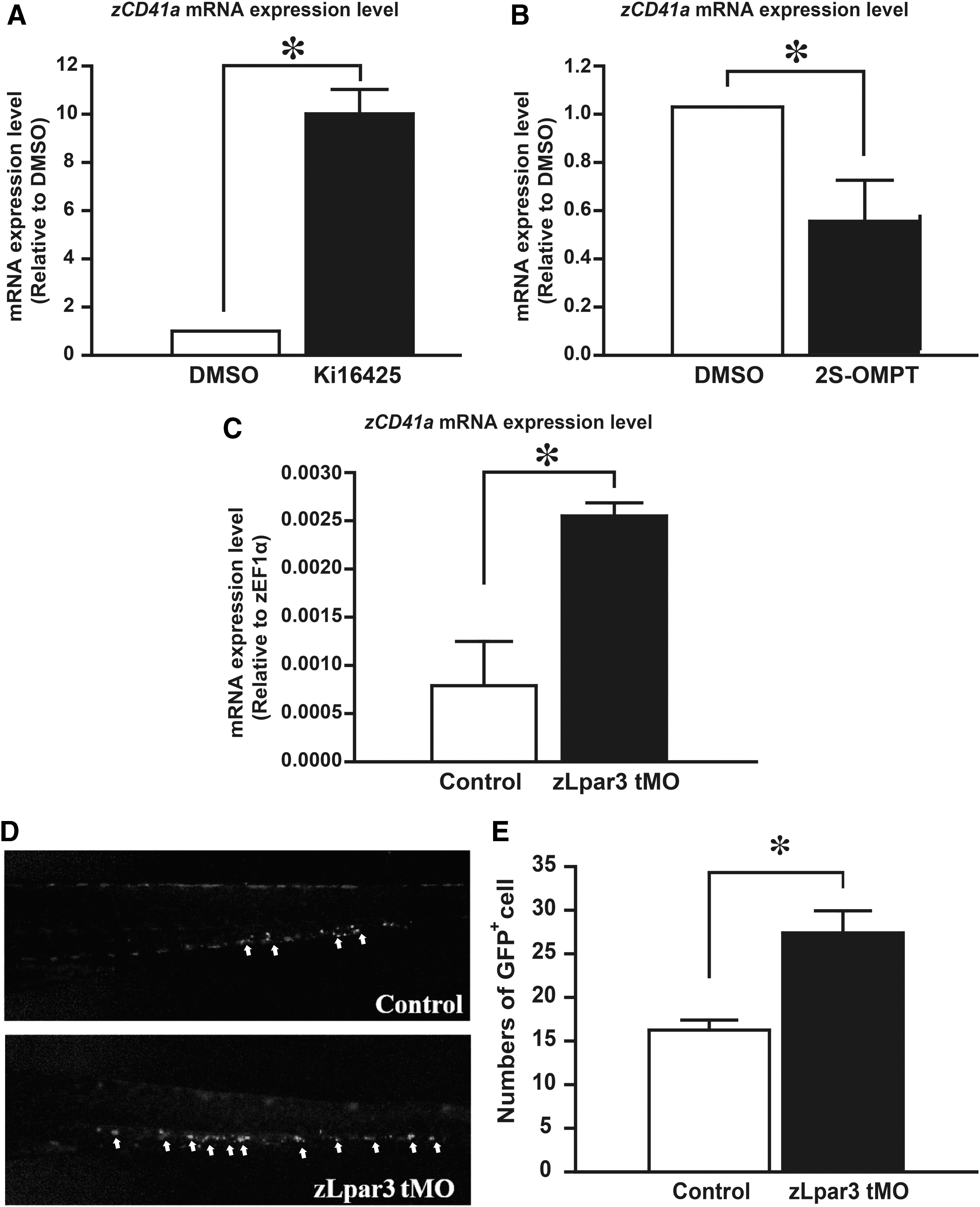
LPA3 inhibits thrombopoiesis in zebrafish. A total of 30 zebrafish embryos from 48 hpf were treated with
Blockade of LPA3 promotes thrombopoiesis during the definitive wave of hematopoiesis in zebrafish
To further confirm the results from pharmacological experiments in zebrafish, we applied zLpar3 Morpholino in zebrafish embryos. Knockdown of zLpar3 enhanced mRNA expression of endogenous zCD41 (Fig. 5C). Furthermore, we use Tg(CD41:eGFP) zebrafish to observe the level of thrombocytes. Previous studies have established Tg(CD41:eGFP) zebrafish to study the development of thrombocyte in zebrafish [20,21]. CD41-GFP+ cells are represented as the numbers of thrombocyte in this transgenic zebrafish. The number of CD41-GFP+ cells increased during embryonic development, and circulating thrombocytes were readily observed after 3 days postfertilization (dpf) in the cardiac sinus region [20]. After injection of zLpar3 Morpholino, the number of circulating CD41-GFP+ cells was increased at 4 dpf (Fig. 5D). In addition, CD41-GFP+ cells, which were isolated from the whole kidney marrow, were analyzed by flow cytometry. Previous studies suggest that GFPhi cells represent the mature thrombocytes, whereas GFPlo cells represent an immature phenotype [20]. We found that isolated CD41-GFP+ cells from zLpar3 knockdown embryos showed higher intensity of fluorescence compared to controls (Fig. 5E). These results indicate that LPA3 exerts an inhibitory role during thrombopoiesis in zebrafish. We confirmed these findings in an LPA3 knockout transgenic zebrafish model. Consistent with the results observed previously, mRNA expressional level of zCD41 was increased significantly in LPA3-KO zebrafish embryos at 7 dpf (Fig. 6). Overall, our observations demonstrate that activation of LPA3 is an important regulatory signal during the definitive wave of thrombopoiesis in zebrafish.

Knockout of LPA3 promotes thrombocyte differentiation in zebrafish. LPA3 was depleted from the genome of zebrafish using TALEN method. Total RNA was extracted from 30 zebrafish embryos, and mRNA expression of zCD41 was analyzed by qPCR. Values were normalized to zEF1A. The results are shown as the mean ± SD from six independent experiments and analyzed by Student's t-test (*P < 0.05). TALEN, transcription activator-like effector nuclease.
Discussion
HSC differentiation is a complicated process and it is controlled by numerous factors. Within hematopoietic organs, appropriate niches provide essential conditions for HSC lineage commitment [22]. Recent studies have reported the importance of LPA signaling in the microenvironment during mesenchymal differentiation [23,24] and in the regulation of multipotency of neuronal stem cells [25]. Furthermore, LPA plays a crucial role during embryonic hematopoiesis [11] and in the early stages of myeloid differentiation [10]. However, little is known about the effects of LPA on terminal commitment in myeloid lineages. Our previous studies demonstrated that LPA promotes erythropoiesis through LPA3 signaling and inhibits erythropoiesis through LPA2 signaling in vitro and in vivo [13,14]. In the present study, using in vitro and in vivo models, we investigate the role of LPA3 during megakaryopoiesis in human HSCs and zebrafish for the first time. We also highlight that megakaryopoiesis can be modulated by treatment with specific agonists of LPARs.
2S-OMPT, a selective LPA3 agonist, has physiological functions on renal injury [26], neurite growth [27], and smooth muscle contraction [28]. Our previous study demonstrated that 2S-OMPT significantly enhances hemoglobin mRNA transcripts and the expression level of erythroid markers in a leukemia cell line and in human HSCs. In addition, activation of LPA3 in zebrafish embryo and adult mice increases the number of circulating RBCs [14]. Our present study has clarified that activation of LPA3 represses platelet differentiation in human HSCs and zebrafish. CD41 and CD61 protein levels were decreased by LPA treatment, but co-treatment of LPA and Ki16425 was able to recover the expression level of the two surface markers. Most importantly, deletion of LPAR3 in zebrafish resulted in enhanced expression of zCD41 mRNA, suggesting that LPA3-mediated signaling represses thrombopoiesis in zebrafish. Although the expressional level of LPAR2 was increased for 60%–70% during the 7-day induction, we think that it is not important during megakaryopoiesis because of its low expressional level (Supplementary Fig. S1). These results suggest that LPA2 does not participate in LPA-dependent megakaryopoiesis inhibition in normal physiological conditions. In contrast, since the mRNA expression level of LPAR1 did not change under induction of megakaryopoiesis by TPO/SCF in HSCs, we conclude that LPA1 has no critical functions during megakaryopoiesis. Consistent with previous observations that LPA1 has been reported as a developmental cue to regulate early hematopoiesis [11], our current results show that LPA3 plays a major role during platelet differentiation.
This is the first study to confirm the role of LPA in the early stages of megakaryopoiesis, but other LPARs have been reported to participate in the maturation of platelets. Activation of LPA5, one of the non-EDG families of LPAR, has been suggested to regulate platelet activation and shape change of MK cells [29]. In addition, our previous study indicated that LPA2 repressed, whereas LPA3 activated, megakaryopoiesis in the K562 cell model [15]. In our previous study, we found that LPA3 promoted megakaryocytic differentiation through reactive oxygen species activity and enhancement of FLI-1 mRNA expression, which is an important transcription factor involved in directing megakaryopoiesis by antagonizing EKLF [30,31]. This study is in contradiction to our latest finding that LPA3 promotes megakaryopoiesis. Nevertheless, evidence suggests that there might be differences between K562 and human HSCs. One interpretation might be that the two cell models represent different stages of myeloid precursors. K562 cells were derived from a 53-year-old female chronic myelogenous leukemia patient, whereas umbilical cord blood-derived HSCs are embryonic stem cells. Recent studies also suggest that MK ontogeny might have a differential route compared to conventional hematopoietic hierarchy [32], supporting that LPA3 might play opposed roles in K562 and HSCs. Another possibility is that K562 cells lack TPO receptors [33]. In our previous study, we used phorbol 12-myristate 13-acetate (PMA) to induce megakaryopoiesis in K562 cells, which suggests that terminal differentiation driven by PMA may activate different molecular pathways from conventional TPO signaling. The detailed mechanism by which LPA3 signaling regulates the megakaryopoiesis in human HSC requires further examination.
Taken together, our study suggests that the expressional pattern of LPA3 might be a key factor for MK lineage commitment. In addition, our results also suggest that chemical agents, including 2S-OMPT, might be future drug candidates to regulate megakaryopoiesis. These observations may facilitate the development of novel treatment strategies for thrombocythemia.
Footnotes
Acknowledgments
This research was supported by grants (MOST 103-2311-B-002-015 and MOST 104-2311-B-002-013) to H.L. and grants (MOST 104-2628-E-155-002-MY3) to C.L.Y. from the Ministry of Science and Technology of Taiwan.
Author Disclosure Statement
I declare that all of the authors have no competing interests or other interests that might be perceived to influence the results and/or discussion reported in this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.