
Abstract
Tip110 plays important roles for stem cell pluripotency and hematopoiesis. However, little is known about the regulatory mechanisms of Tip110 expression in this process. In this study, we first showed that constitutive Tip110 expression was cell proliferation and differentiation dependent and self-regulated in both human cord blood CD34+ cells. Using a series of molecular techniques, we found that ectopic Tip110 expression led to increased constitutive Tip110 expression through its 3′-untranslated region (3′UTR), specifically through preferential usage of proximal polyadenylation sites within its 3′UTR in cells, including human cord blood CD34+ cells, which indeed led to an increased number of CD34+ cells during differentiation of those cells. Lastly, we showed that Tip110 protein interacted with cleavage stimulation factor 64 (CstF64) protein and that more CstF64 was recruited to the promixal polyadenylation site than the distal polyadenylation site within its 3′UTR. These finding together demonstrates that constitutive Tip110 expression is regulated, at least in part, through its interaction with CstF64, recruitment of CstF64 to, and selective usage of those two polyadenylation sites within its 3′UTR.
Introduction
H
The 3′ end processing of pre-mRNA is an important part of RNA metabolism and plays important roles in eukaryotic mRNA maturation [15,16]. Almost all eukaryotic pre-mRNA and several noncoding RNA transcripts are polyadenylated [17]. The polyadenylation process consists of cleavage of the pre-mRNA at a polyadenylation site, followed by addition of a poly(A) tail to the 5′ cleavage product. More than 60% of human genes have multiple polyadenylation sites; their expression involves alternate cleavage and polyadenylation [18], giving rise to two or more distinct mRNA isoforms with different 3′ untranslated regions (3′UTR) and as a result regulating the repertoire and expression level of proteins [19]. Importantly, 3′UTR have increasingly been recognized as sites for microRNA–mRNA interactions [20]. Thus, a longer 3′UTR is often associated with increased usage of the distal polyadenylation site (dPAS) and decreased protein expression. Changes in the polyadenylation site usage and 3′UTR length, resulting in changes of protein expression, have now increasingly been linked to oncogene activation and various disorders, including cancers [19 –23]. However, it is not known whether Tip110 plays a role in other RNA metabolic processes besides pre-mRNA splicing.
In the current study, using a series of molecular biology strategies, we demonstrate that Tip110 expression was cell proliferation and differentiation dependent and was self-regulated through selective usage of the polyadenylation sites within its 3′UTR and interaction with its 3′UTR and cleavage stimulation factor 64 (CstF64). These findings will help to clarify the role of Tip110 expression and function of human cord blood CD34+ HSC and progenitor cells.
Materials and Methods
Plasmids and siRNA
pCSC.GFP and pCSC.Tip110.GFP were previously described [4]. Full-length CstF64 cDNA was purchased from OriGene (Rockville, MD). pc3.CstF64 was constructed using a standard polymerase chain reaction (PCR) cloning technique using CstF64-specific primers (Table 1). pGL3.Tip110-5′UTR-Luc and pGL3.c-Myc-5′UTR-Luc were previously described [5]. pLighSwitch.Luc-Tip110-3′UTR was purchased from SwitchGear Genomics (Carlsbad, CA). It contained a constitutive prompter PRL10 and the full-length Tip110 3′-UTR (1,346 nt). Tip110 siRNA, CstF64 siRNA, and control siRNA were purchased from Thermo Scientific (Lafayette, CO).
3′UTR, 3′-untranslated region; dPAS, distal polyadenylation site; ORF, open reading frame; pPAS, proximal polyadenylation site.
Cells, cell transfection, and luciferase reporter gene assay
The 293T and K562 were purchased from American Tissue Culture Collection (ATCC, Manassas, VA) and maintained in Dulbecco's modified Eagle's medium or RPMI 1640 medium, respectively, supplemented with 10% fetal bovine serum. Human cord blood was provided to us by the Cord:Use CB Bank, Orlando, FL and used as reported [5,6]. CD34+ cells were purified within 24 h of collection using immunomagnetic selection (Miltenyi Biotec). CD34+ cells (>93% pure) were cultured in Iscove's modified Dulbecco's medium (IMDM) containing 10% fetal bovine serum (FBS) with the cytokine combination of: 100 ng/mL of human stem cell factor (SCF), 100 ng/mL human FLT3 ligand (FL), and 100 ng/mL of human thrombopoietin (Tpo) from R&D Systems (Minneapolis, MN). Cell transfections were performed on the Nucleofector device (Amaxa Biosystem) according to the manufacturer's instructions [24]. pcDNA3 was used to equalize amounts of DNA transfected for all transfections. Luciferase reporter gene activities were determined using the Firefly/Renilla luciferase system (Promega, Madison, WI). pTKβGal was included to normalize variations in transfection efficiency for all transfections. siRNA transfections were performed using Lipofectamine 2000 LTX (Invitrogen, Carlsbad, CA).
Preparation of whole cell lysates, immunoprecipitation, and western blotting
Cells were washed twice with ice-cold phosphate-buffered saline (PBS) and harvested for whole cell lysates according to the previously described methods [25]. Briefly, cell pellets were suspended in two volumes of whole cell lysis buffer (10 mM NaHPO4, 150 mM NaCl, 1% Triton X-100, 0.1% sodium dodecyl sulfate (SDS), 0.2% sodium azide, 0.5% sodium deoxycholate, 0.004% sodium fluoride, and 1 mM sodium orthovanadate) and incubated on ice for 10 min. Whole cell lysates were obtained by centrifugation and removal of cell debris. For western blotting, cell lysates or immunoprecipitates were electrophoretically separated on 10% SDS–polyacrylamide gel electrophoresis and analyzed by immunoblotting using an anti-CstF64 antibody (Santa Cruz Biotechnologies, Santa Cruz, CA), and anti-Tip110 antibody [5] and appropriate horseradish peroxidase-conjugated secondary antibodies, followed by visualization with the enhanced chemiluminescence (ECL) system (GE Healthcare Biosciences, Pittsburgh, PA).
RNA isolation and quantitative reverse transcription polymerase chain reaction
Total RNA was isolated using TRIzol (Life Technology, Grand Island, NY) according to the manufacturer's instructions. cDNA was synthesized using the ScriptII RT Reagent Kit (Promega). cDNAs were used for quantitative polymerase chain reaction (qPCR) using the Power SYBR® Green PCR Kit (Life Technologies, Grand Island, NY) according to the manufacturer's instructions. The qPCR program consisted of 1 cycle at 95°C for 10 min; 40 cycles at 95°C for 15 s and at 60°C for 1 min. The Bio-Rad CFX manager software was used to calculate gross-threshold (CT) values. The 2(−ΔΔCT) was calculated to represent the fold change of gene expression and normalized using β-actin as a reference.
Northern blotting
Total RNA was isolated using TRIzol for further extraction with acid phenol and treated with RNase-free DNase I (Roche, Indianapolis, IN). Twenty-microgram RNA was separated on 1% agarose formaldehyde denaturing gel, followed by transfer to nylon membrane overnight using 10 × SSC (1.5 M NaCl and 0.15 M Na Citrate, pH 7.0). A probe (Table 1) was synthesized and biotin labeled using the Biotin RNA Labeling Mix (Roche). Hybridization was performed at 55°C overnight; Tip110 mRNA was detected using the LightShift Chemiluminescent RNA EMSA Kit (Thermo Fisher Scientific, Waltham, MA). Membrane was stripped for reprobe with a human β-actin cDNA as an internal control as described (24).
3′ rapid amplification of cDNA ends and semiquantitative RT- PCR
Total RNA (1 μg) isolated above was reverse transcribed to cDNA using d(T)nP2 anchored primer (Table 1). Aliquots of cDNA were used as a template for PCR with specified primers and a program consisting of 1 cycle at 95°C for 5 min; 40 cycles at 95°C for 1 min, 50°C–54°C (depending on primer Tm) for 10 s, 72°C for 30 s, and 1 cycle of 72°C for 10 min. PCR products were analyzed on a 1.5% agarose gel.
Southern blotting
Following 3′ rapid amplification of cDNA ends (3′ RACE), semiquantitative PCR, and gel electrophoresis, the gel was alkaline denatured and transferred onto a positively charged nylon membrane (GE Healthcare, Pittsburgh, PA). A DNA fragment covering from 1 to 249 bp of Tip110 3′UTR was isolated through full digestion with EcoRI and PstI followed by biotin labeling using Biotin-High Prime (Roche, Indianapolis, IN) as a probe. Southern blot hybridization was performed at 65°C for 16 h. Stringent washing and immunological detection were performed following kit instructions (Thermo Fisher Scientific, Waltham, MA).
RNA immunoprecipitation and semiquantitative reverse transcription polymerase chain reaction
The 293T (10 × 106) cells were transfected with Tip110 or cDNA3, cultured for 48 h, and fixed by including 540 μL formaldehyde to 20 mL DMEM for 10 min at room temperature, followed by glycine buffer to stop the reaction. The cells were then lysed in the lysis buffer [50 mM HEPES, pH 7.0, 60 mM KCl, 5 mM MgCl2 and 0.5% NP-40, 1 mM DTT, 0.1 U/mL RNasin, and protease inhibitor cocktail (Roche)] and sonicated to shear RNA to 200–500 bp fragments. The lysates were precleared by incubation with protein A sepharose beads (GE Healthcare Biosciences, Pittsburgh, PA) at 4°C for 1 h. The cleared lysates were incubated with an anti-CstF64 antibody (Santa Cruz), or isotype-matched IgG control at 4°C overnight with gentle mixing. Fresh protein A/G sepharose beads were added, the mixtures were incubated at 4°C for one more hour. The beads were recovered by centrifugation. After washes with lysis buffer twice and PBS twice, the beads were suspended in TRIzol for RNA isolation. Acid phenol extraction was performed to prevent any amplification of contaminated genomic DNA in the RNA preparation. The RNA was analyzed by semiquantitative Titan One Tube reverse transcription polymerase chain reaction (RT-PCR) (Roche) using indicated primers (Table 1) followed by running on 1% agarose gel for detection. Cell lysates and immunoprecipitates were analyzed by western blotting.
Flow cytometric analysis of human cord blood CD34+ cells
Freshly purified and cultured human cord blood CD34+ cells were washed twice with PBS and stained with mouse anti-human CD34-PE/CD34-APC/CD34-FITC antibody, CD38-PE antibody, CD45RA-PECF594, mouse anti-human CD90-PECy7, CD49f-PerCPCy5.5, and CD41a-PE from BD Biosciences (San Jose, CA); CD235a-APC/Cy7, CD14-Pacific Blue, and CD66b-PE/Cy7 from BioLegend (San Diego, CA); and Tip110 in the dark at room temperature for 30 min. Staining with mouse IgG-FITC, mouse IgG-APC, or mouse IgG-PE alone were included as negative controls. Cells were subjected to FACS flow cytometric analysis (Becton Dickinson, Franklin Lakes, NJ).
Statistical analysis
Data were expressed as mean ± standard deviation unless otherwise stated and representative of at least three independent experiments. All experimental data were analyzed by two-tailed student's t test. A P value <0.05 was considered to be statistically significant and marked as *; a P value <0.01 was considered to be statistically highly significant and marked as **.
Results
Constitutive Tip110 expression in response to cell proliferation and differentiation and ectopic Tip110 expression
Tip110 expression in normal and nonproliferating cells is low, but is highly elevated in proliferating stem cells and cancer cells/tissues [3 –6]. To determine Tip110 expression in relationship to cell proliferation and differentiation, we took advantage of human cord blood CD34+ cells and used them as the model. Human cord blood CD34+ cells were freshly purified and cultured in the presence of 10% FBS and a defined set of cytokines, SCF, FL, and TPO stimulation (19,22), which allow those cells to transition from the quiescent stage (0 h) to the proliferation stage, and then to the differentiation stage. Constitutive Tip110 mRNA expression, determined by semiquantitative RT-PCR and quantitative reverse transcription polymerase chain reaction (qRT-PCR) increased rapidly from the quiescent state to the proliferation stage as early as within 2 h, up to 1 day, during which there were no changes of the CD34+ cell percentage (Fig. 1A). After extended culturing of those cells for up to 10 days, CD34+ cells differentiated, as CD34+ cell percentage decreased from 97% to 20%, constitutive Tip110 mRNA became less detectable. The results were consistent with our previous publication [5]. In the meantime, we also monitored the differentiation of those cells using several defined cell lineage markers and flow cytometry and determined constitutive Tip110 expression in those cells. The markers were CD235a+ for mature erythrocytes and erythroid precursor cells, CD41a+ for platelets, CD14+ for macrophages, and CD66b+ for neutrophils and eosinophils. Compared with day 1, day 3 to day 7 all showed gradual and considerable increases for CD235a+, CD14+, and CD66b+ cell percentages, whereas considerable decreases in CD34+ cell percentage (Fig. 1B). Meanwhile, CD41a+ platelets remained relatively stable, between 13% and 20%. Except CD41+ cells, constitutive Tip110 protein was highest in CD34+ cells, and significantly lower in all other cells, including CD34− cells at day 7 (Fig. 1C). Similar results were obtained for constitutive Tip110 protein expression in those cells on day 5 (data not shown). Those results confirm the cell proliferation- and differentiation-dependent nature of constitutive Tip110 expression.
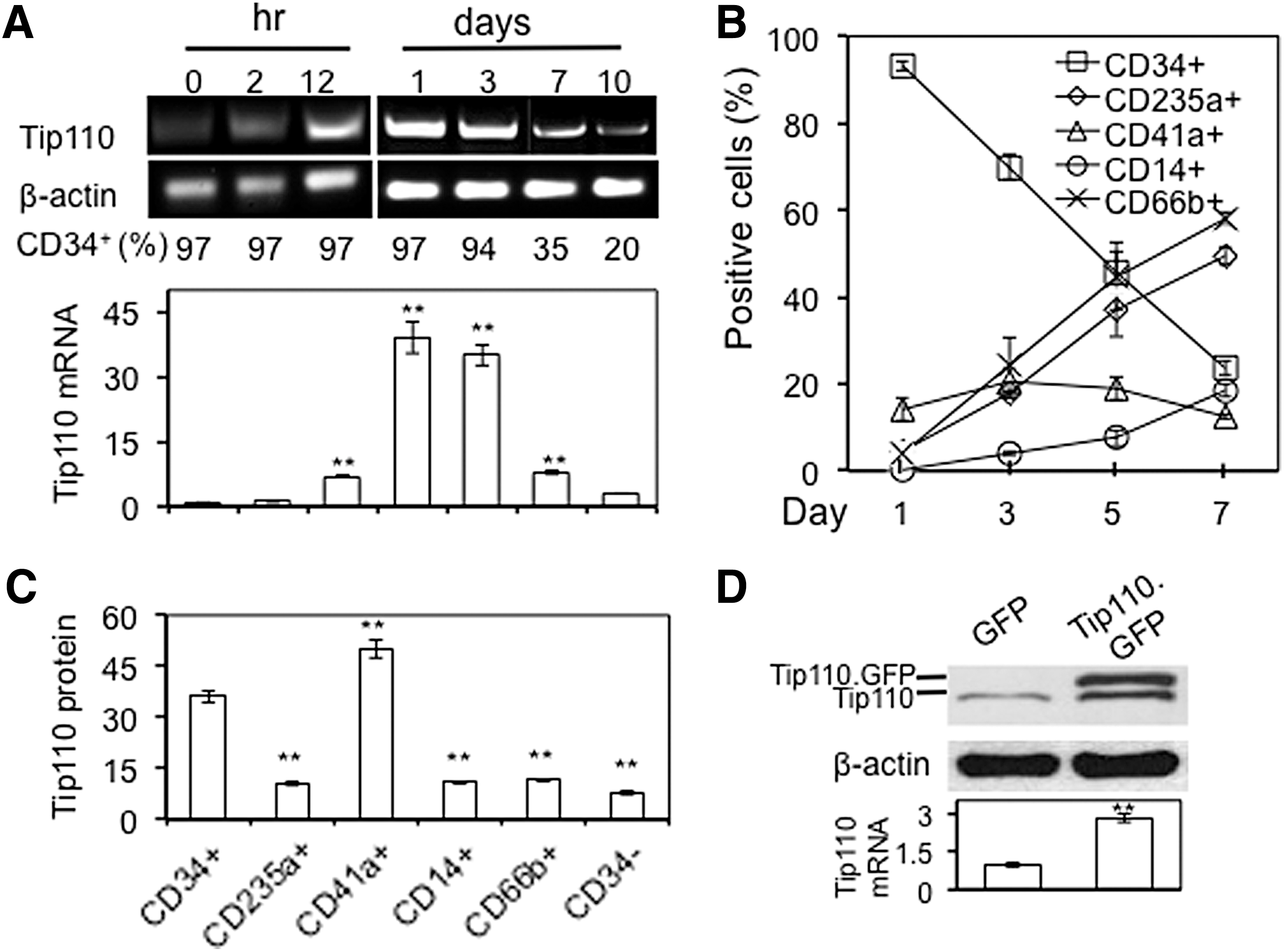
Changes of Tip110 expression in proliferating and differentiating human cord blood CD34+ cells and K562.
To further validate the findings and to overcome the difficulties in detecting Tip110 protein expression in the limited number of primary cells above, we used the in vitro induced K562 differentiation model to determine the relationship between constitutive Tip110 protein expression and cell proliferation and differentiation. K562 are well characterized for its property of proliferation and differentiation into megakaryocytes with PMA and ionomycin treatment [26 –29]. Similarly, to the findings in human cord blood CD34 cells above, Tip110 mRNA expression increased during proliferation and decreased during differentiation (data not shown). In addition, we found that ectopic Tip110 expression in the form of Tip110.GFP, which allowed us to distinguish constitutive Tip110 from exogenous Tip110 by the difference of molecular weights, resulted in significant increases in constitutive Tip110 protein expression (Fig. 1D).
Tip110 3′UTR but not Tip110 5′UTR was involved in self-regulation of Tip110 expression
Tip110 regulates expression of several genes through their promoters, including c-Myc [6]. To evaluate the possibility of self-regulatory mechanisms for Tip110 expression, we first determined the effects of Tip110 on its own promoter (5′UTR). The 293T cells were transfected with a Tip110 promoter-driven luciferase reporter gene pGL3.Tip110-5′UTR-Luc (Fig. 2A) and Tip110 expression plasmid, or Tip110 siRNA. Tip110 overexpression and knockdown showed no effects on the Tip110 promoter-driven reporter gene expression (Fig. 2B). To validate this strategy, a similarly designed c-Myc promoter-driven reporter gene was included as a positive control (Fig. 2C). As expected, Tip110 overexpression increased the c-Myc promoter-driven reporter gene expression, whereas Tip110 knockdown decreased the c-Myc promoter-driven reporter gene expression (Fig. 2D). Then, we evaluated whether Tip110 3′UTR could be involved in Tip110 self-regulation. The 293T cells were transfected with Tip110 3′UTR-containing luciferase reporter gene pLightSwitch.Luc-Tip110-3UTR (Fig. 2E) and increasing amounts of Tip110 or Tip110 siRNA. Tip110 overexpression led to dose-dependent increases in the Tip110 3′UTR reporter gene activities (Fig. 2F), whereas Tip110 knockdown led to dose-dependent decreases in the reporter gene expression (Fig. 2G). The results together suggest that the Tip110 3′UTR, but not 5′UTR, is likely involved in Tip110 self-regulation.
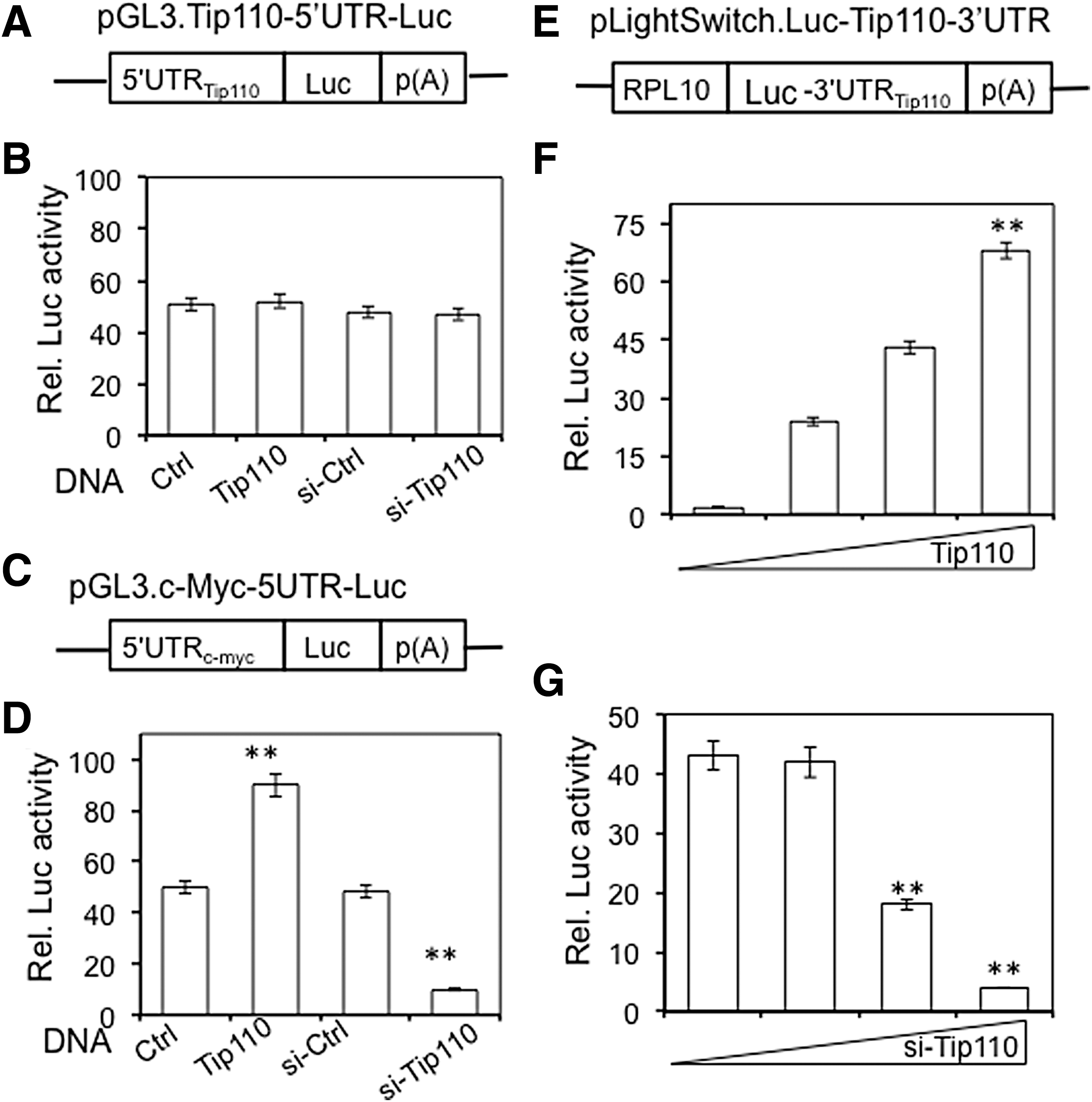
Tip110 self-regulation mediated through its 3′UTR, but not 5′UTR.
Ectopic Tip110 expression led to less dPAS-containing Tip110 mRNA and more CD34+ cells
Bioinformatic analysis revealed two alternate polyadenylation (APA) sites within the Tip110 3′UTR: one proximal polyadenylation site (pPAS, nt. 3876–3881) and one dPAS (nt. 4347–4353) (Fig. 3A). APA has been shown to be a major regulatory mechanism of gene expression through miRNA [30 –32]. Thus, we next determine whether APA was involved in regulation of Tip110 expression and CD34+ proliferation and differentiation. Human cord blood CD34+ cells were transduced with lentivirus-expressing Tip110 or its control lentivirus, cultured, harvested, and determined expression of different Tip110 mRNA species at day 3 and 7, whereas monitored CD34+ cell differentiation. Tip110 mRNA species were identified by qRT-PCR using primer pairs for different regions of the full-length Tip110 mRNA transcripts (Fig. 3A, Table 1). Compared with the control, ectopic Tip110 expression led to significantly higher Tip110 mRNA expression at both day 3 and 7, determined by primer pair Q0, which amplified both constitutive and exogenous Tip110 mRNA (Fig. 3B). In addition, as shown above, more constitutive Tip110 mRNA was noted in CD34+ at day 3 than that at day 7. Primer pairs Q1 and Q3 were targeted at the junction of Tip110 open reading frame (ORF) and 3′UTR, therefore allowing detection of constitutive Tip110 species only, as they were not present in the Tip110 mRNA derived from ectopic Tip110 expression. At day 3, primer pair Q1 detected significantly more constitutive Tip110 mRNA in Tip110-transduced CD34+ cells than that of the control (Fig. 3C, left panel), whereas primer pair Q3 showed comparable constitutive Tip110 mRNA between Tip110-transduced and control CD34+ cells. Nevertheless, constitutive Tip110 mRNA detected by primer pair Q3 was significantly lower than that by primer pair Q1. At day 7, primer pair Q1 detected more constitutive Tip110 mRNA in Tip110-transduced CD34+ cells than that of the control (Fig. 3C, right panel). However, primer pair Q1 detected significantly less constitutive Tip110 mRNA in Tip110-transduced CD34+ cells than that of the control. Similar to day 3, at day 7, constitutive Tip110 mRNA detected by primer pair Q3 was significantly lower than that by primer pair Q1. Meanwhile, a comparable CD34+ percentage was noted between Tip110-transduced and control-transduced cells (Fig. 3D) at day 3. But, at day 7, there were significantly more CD34+ cells derived from Tip110-transduced CD34+ cells than that from the control. These results provide further evidence that ectopic Tip110 expression increases constitutive Tip110 expression and the number of CD34+ cells and suggest that regulation of Tip110 expression during CD34+ cell differentiation involves APA and selective use of pPAS.
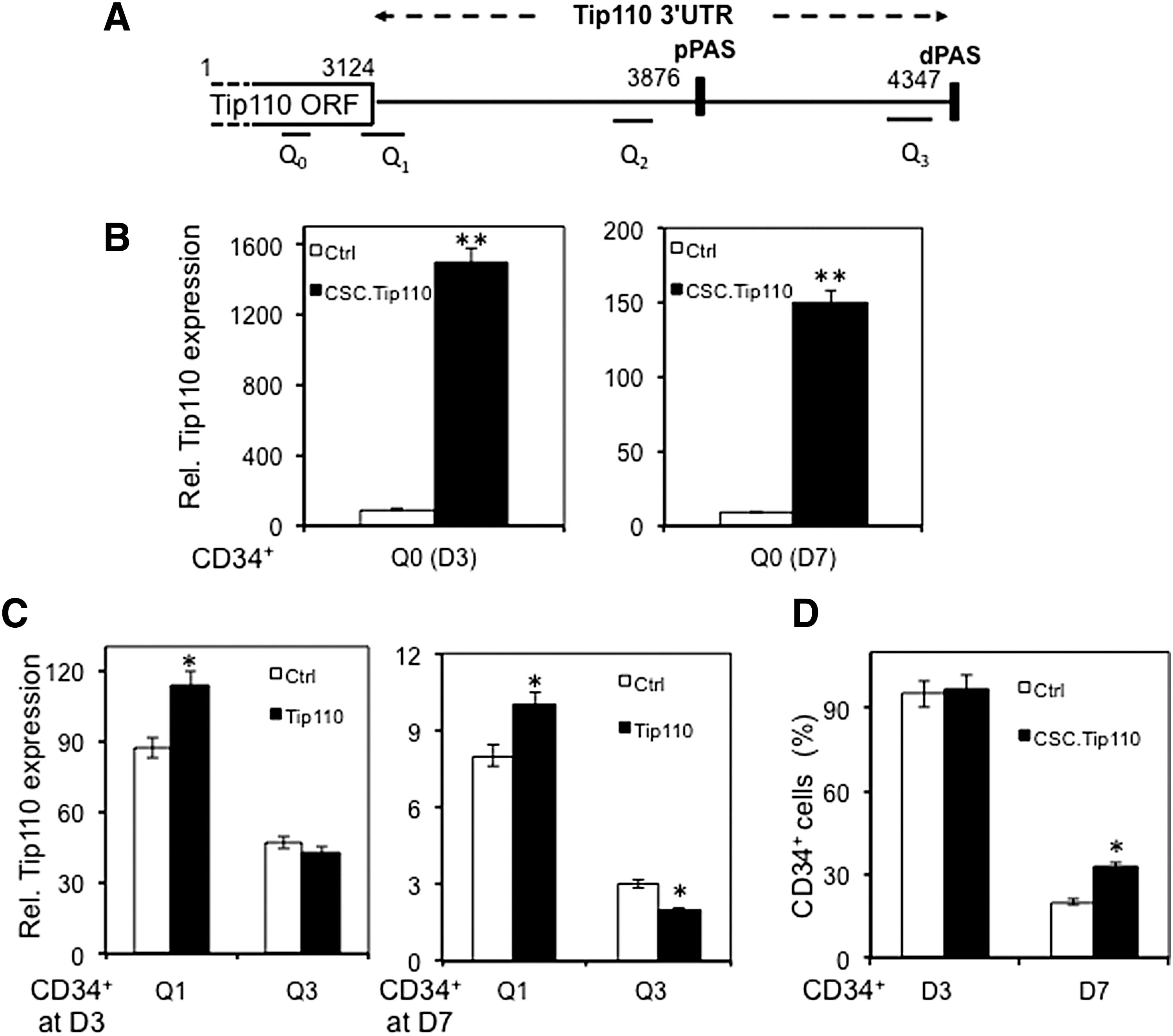
Effects of ectopic Tip110 expression on human cord blood CD34+ differentiation and expression of constitutive Tip110 mRNA transcripts.
Ectopic Tip110 expression led to preferential usage of pPAS
APA of 3′UTR and increased usage of pPAS, aka shortened 3′UTR mRNA have been noted in proliferating stem cells and cancer cells [20,23,33,34]. Thus, to ascertain the role of APA and relative usage of pPAS and dPAS in relation to Tip110 expression, we transfected 293T with Tip110 expression plasmid or its control and determined the relative level of constitutive Tip110 mRNA species using northern blotting, 3′ RACE with specific primers (Fig. 4A, Table 1), and Southern blotting. Tip110 protein expression was confirmed using western blotting (Fig. 4B). Northern blotting detected the full-length Tip110 mRNA transcript (4,353 nt.) in the transfection control cells, and a shorter Tip110 mRNA transcript (3,881 nt.) in Tip110-transfected cells (Fig. 4C). The shorter Tip110 transcript was estimated to be the size of the Tip110 mRNA that only contained the pPAS site. To ensure these findings, 3′RACE was performed. Specifically, the reverse transcription was performed using the anchored primers dTnP2, followed by PCR using primer pair with P1 and P2. While two PCR products (440 and 917 bp) were detected in the control cells, only the short PCR product (440 bp) was detected in Tip110-transfected cells (Fig. 4D). Those results were confirmed using another nest primer P3 and primer P2, which gave rise to two PCR products (385 and 862 bp) in control cells and only the 385 bp PCR product in Tip110-transfected cells (Fig. 4E). To further ensure the specificity of amplifications by primer pairs P1 and P2, Southern blotting was performed using a DNA probe located within the amplified products. A PCR product within the Tip110 ORF was included as a specificity control (Fig. 4D) for Southern blotting. Only those two 3′RACE products were detected by Southern blotting (Fig. 4F). These results together demonstrated that ectopic Tip110 promoted APA and preferential usage of pPAS, increasing constitutive Tip110 expression.
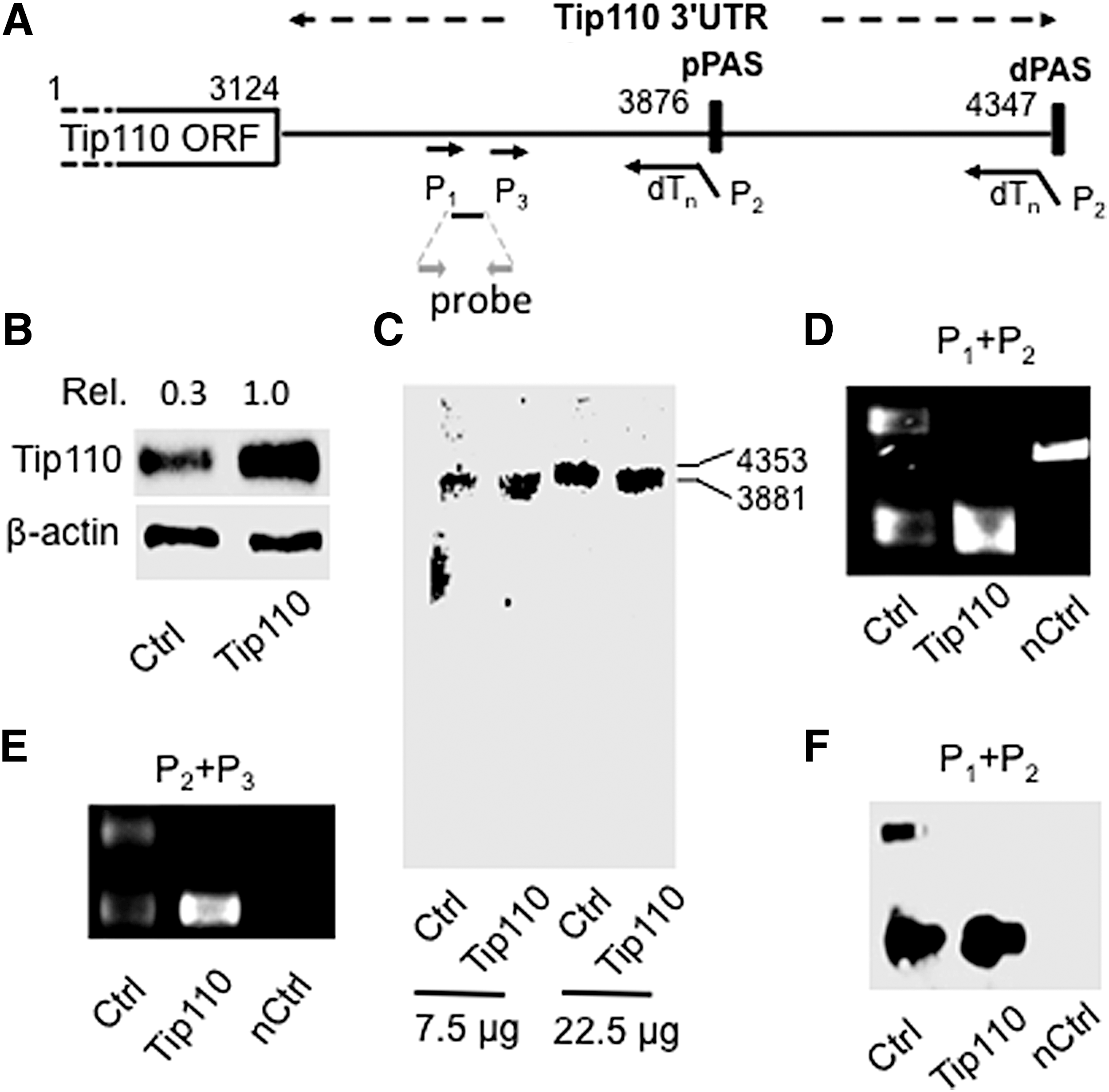
Effects of ectopic Tip110 expression on expression of constitutive Tip110 mRNA transcripts in 293T.
Tip110 bound to cleavage stimulation factor CstF64 and regulated each other's expression
Tip110 protein binds to RNA polymerase II (RNAP II) [10], which is directly involved in and important for every step of mRNA transcription of most genes, including cleavage and polyadenylation [35,36]. During cleavage and polyadenylation, polyadenylation site (PAS) recognition is achieved by RNA–protein interaction. The major factors involved in PAS are cleavage and polyadenylation specificity factor (CPSF) and cleavage stimulation factor (CstF). CstF is composed of three subunits, CstF77, CstF50, and CstF64 [19]. CstF64 directly binds to RNA through its RNA recognition motif and is an important regulator of APA. Higher levels of CstF64 expression are associated with increased use of pPAS in certain mRNA transcripts [33,34]. Thus, we assessed possible roles of CstF64 on Tip110 expression-associated preferential usage of the pPAS with Tip110 3′UTR. We first determined whether Tip110 bound to CstF64. Immunoprecipitation followed by western blotting detected Tip110 protein in CstF64 immunoprecipitates, not other control immunoprecipitates, suggesting that Tip110 and CstF64 formed a complex (Fig. 5A). We next evaluated whether CstF64 overexpression had any effects on the Tip110 3′UTR-mediated reporter gene activity. The 293T cells were transfected with pLightSwitch.Luc-Tip110-3′UTR and CstF64 expression plasmid. CstF64 overexpression led to significantly increased Tip110-3′UTR reporter expression (Fig. 5B, top panel). CstF64 overexpression was detected by western blotting (Fig. 5B, bottom panel). CstF64 knockdown by CstF64 siRNA led to significantly decreased reporter gene expression (Fig. 5C, top panel). CstF64 knockdown was confirmed by western blotting (Fig. 5C, bottom panel). We further showed that Tip110 knockdown resulted in decreased CstF64 protein expression (Fig. 5D). No change in CstF64 expression was observed with ectopic Tip110 overexpression (data not shown). These results raised the possibility that CstF64 might be involved in self-regulation of Tip110 expression.
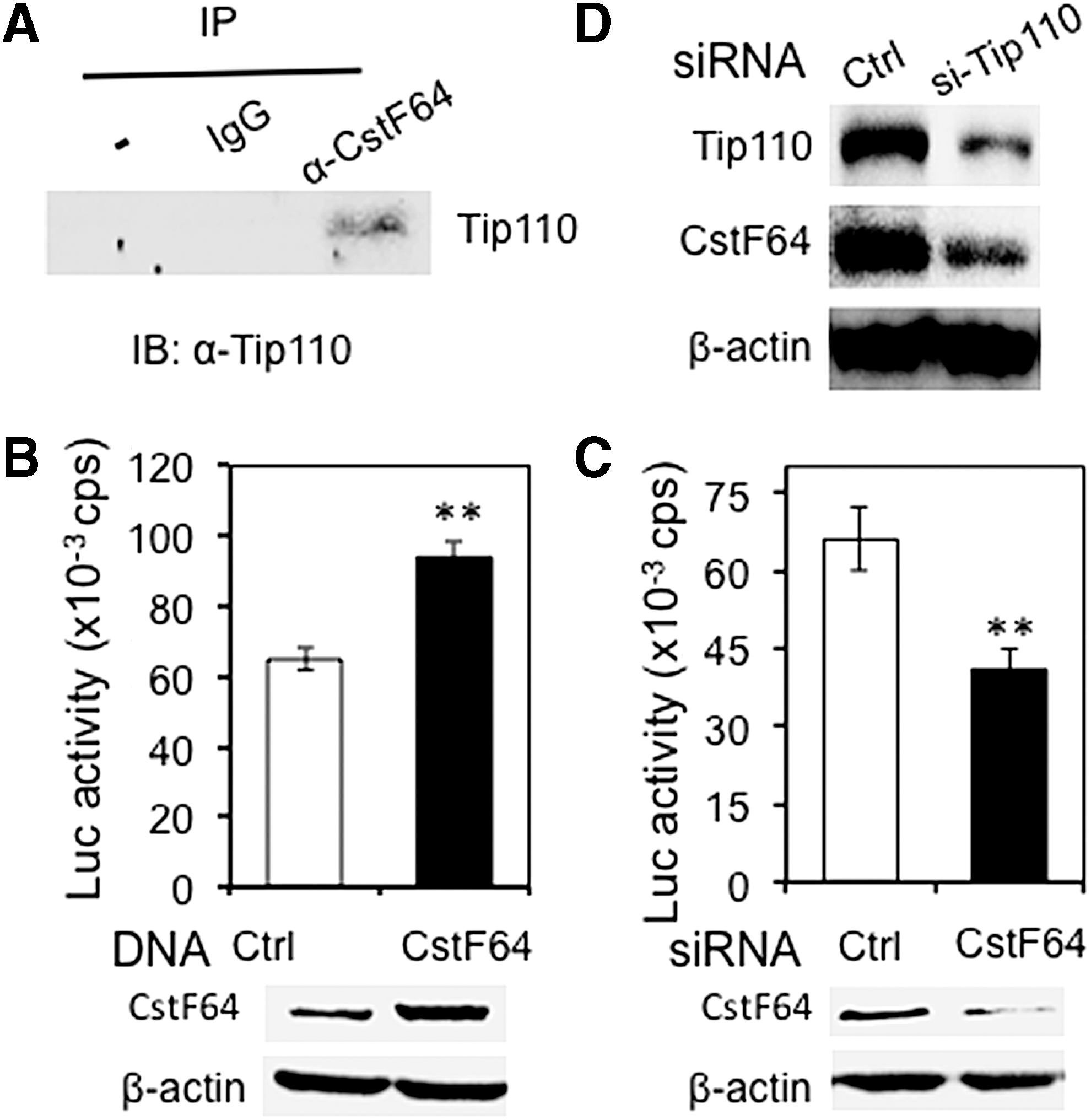
Tip110 interaction with CstF64 and their reciprocal regulation.
Ectopic Tip110 led to decreased CstF64 recruitment to the dPAS
Higher levels of CstF64 expression are associated with increased usage of the pPAS [37 –39]. We then determined whether Tip110 overexpression would alter CstF64 recruitment to pPAS and dPAS. The RNA immunoprecipitation assay was performed; CstF64 recruitment to pPAS and dPAS was compared between control and Tip110-transfected cells. The 293T cells were transfected with Tip110 or its control. Increased Tip110 protein expressed was confirmed by western blotting (Fig. 6A). CstF64 protein in CstF64 immunoprecipitates was also confirmed to be comparable between control and Tip110-transfected cells by western blotting (Fig. 6B). Little CstF64 protein was detected in isotype-matched IgG-immunoprecipitates. CstF64-bound RNA transcripts from immunoprecipitates were determined for qRT-PCR using primer pairs (Fig. 3A, Table 1). There was no detection of RNA with primer pair Q0 located within ORF, a very low detection of RNA with primer pair Q1 located at the junction of ORF and 3′UTR, a higher detection of RNA with primer pair Q2 located close to pPAS, and a lower detection of RNA with primer pair Q3 located close to dPAS (Fig. 6C). No difference of RNA level was detected between the control and Tip110-transfected cells. However, a significant lower of RNA was detected in Tip110-transfected cells than the control. The results provide strong evidence that ectopic Tip110 result in less CstF64 recruitment to dPAS and suggest that Tip110 self-regulation is a likely result of preferential usage of pPAS with the Tip110 3′UTR.

Effects of ectopic Tip110 expression on CstF64 binding to pPAS and dPAS. The 293T cells were transfected with Tip110 expression plasmid, or cDNA3 control, cultured for 48 h, crosslinked with formaldehyde, and harvested for cell lysates, followed by western blotting using an anti-Tip110 or anti-CstF64 antibody
Discussion
We demonstrated that constitutive Tip110 expression was cell proliferation and differentiation dependent that ectopic Tip110 expression resulted in increased constitutive Tip110 expression in immature human cord blood CD34+ cells, and that the Tip110 3′was involved in this process. Tip110 expression resulted in selective usage of the PAS within its 3′UTR using northern blotting, 3′RACE, and Southern blotting analysis. Moreover, CstF64 was identified to interact with Tip110 and regulate Tip110 3′UTR activity. CstF64 was also recruited to Tip110 poly(A) sites within the Tip110 3UTR, with stronger more to pPAS than dPAS in the presence of Tip110 overexpression. These findings support the notion that Tip110 expression is self-regulated through selective usage of the PAS within its 3′UTR.
Activation of HSC is often accompanied by cell proliferation. We have shown that Tip110 was expressed in human CD34+ cells and decreased with differentiation of CD34+ cells. CD41- HSC are more proliferative [40] and thus showed a high level of Tip110 expression (Fig. 1C). There are several other genes with regulatory patterns of expression similar to that of Tip110 during cell proliferation and activation. Oncoprotein 18 (Op18) was identified as a proliferation-related phosphoprotein and expressed at high levels in leukemia cells [41]. When normal peripheral blood lymphocytes were activated with PHA, Op18 expression increased, peaked, and then declined [41]. Proliferating cell nuclear antigen is another example. It is expressed in different types of proliferating cells, including mitogen-stimulated human peripheral blood lymphocytes [42]. Based on the structure and function of these genes, it would be interesting to determine whether self regulation through selective usage of the PAS in their 3′UTR is involved in the expression of those genes.
Several RNA-binding proteins are capable of binding their own pre-mRNA and regulating their own expression. HuR protein regulates its own expression through alternative usage of the PAS and interaction with its PAS-associated GU-rich element (GRE) sequence [43]. Another example is TDP-43 protein, which regulates its own expression by targeting the poly(A) site within its 3′UTR [44]. Consistent with those findings, we showed that Tip110 regulated its own expression through interaction with its own pre-mRNA and its interaction with CstF64 and RNAP II [10]. Tip110 binds to RNAP II C-terminal domain (CTD) [10]. RNAP II is enriched downstream of the poly(A) site; high usage proximal sites have higher upstream nucleosome occupancy and lower downstream nucleosome occupancy [45]. RNA- immunoprecipitation by using CstF64 antibody showed that CstF64 protein bound to Tip110 3′UTR proximal GU-rich domain in the presence of Tip110. This finding suggests that Tip110 may be presented in the CPSF (cleavage/polyadenylation specificity factor) complex, as CstF64 is present in the CstF complexes. Both CstF and CPSF are multisubunit factors for RNA recognition and interact with each other [46]. RNAP II CTD as a scaffolding factor binds to both CPSF and CstF [47]. Thus, Tip110 interaction with RNAP II may mediate interaction with the corresponding processing factors for poly(A) site cleavage. Changes in the levels of CstF64 expression have been shown as a general processing factor to control poly(A) selection [48,49] and in most cases promotes usage of pPAS [38]. Interestingly, we showed that endogenous CstF64 expression level was not altered by ectopic Tip110 expression, but was downregulated by Tip110 knockdown (Fig. 5). Tip110 is a nuclear protein and regulates pre-mRNA splicing and gene transcription that might be important in Csft64 expression. Nevertheless, this unexpected finding does not argue against the notion that Tip110 interaction with CstF64 and RNAP II may contribute to the preferential selection and usage of the pPAS within its 3′UTR.
Tip110 protein has been proposed to regulate alternative splicing of pre-mRNA through cooperation with RNPS1 protein, a pre-mRNA splicing factor [50] and through interaction with U4/U6 [51,52]. In the current study, we demonstrated that Tip110 regulated its own expression through selective usage of the PAS in its 3′UTR. It is very likely that Tip110 regulates expression of other genes through similar mechanisms. Recent studies have shown global changes in 3′UTR lengths in rapidly proliferating cells, including stem cells and cancer cells [53,54]. The 3′UTR shortening results in a loss of negative regulatory elements, mostly the microRNA-binding sites. Therefore, 3′UTR shortening directly gives rise to increased protein expression. Thus, the results from our current study not only provide mechanisms for elevated levels of Tip110 protein in human cord blood CD34+ cells, and possibly cancer cells, but also raise the possibility that Tip110 may also regulate pre-mRNA splicing and expression of other genes through alternate cleavage and polyadenylation.
Thus, when cells are in the proliferation state, Tip110 and polyadenylation factors are activated resulting in enhanced proximal 3′ cleavage and polyadenylation activity. Shortened 3′UTR of Tip110 escapes targeting by miRNAs due to elimination of their binding sites [55]. The upregulated Tip110 expression forms a positive feedback for its mRNA shortening. When cells are in a nonproliferating state, there is less Tip110 and polyadenylation factors recruited to the proximal 3′ cleavage and polyadenylation site, so there exist more noncleaved 3′UTR. The longer 3′UTR provides more miRNA-binding sites that eventually reduce the efficiency of protein production (Fig. 7).

Scheme of the regulatory mechanisms of constitutive Tip110 expression in human cord blood CD34+ cells. In freshly isolated human cord blood CD34+ cells, more CstF64, part of the proccessive polyadenylation complex binds to the dPAS, more full-length Tip110 RNA transcripts (containing both dPAS and pPAS) are utilized. Those full-length transcripts allow more binding sites for Tip110 miRNA and result in less Tip110 protein expression. When those cells begin to proliferate, more CstF64 binds to pPAS, and more shorter Tip110 transcripts (only containing pPAS) are utilized. Those transcripts allow fewer binding sites for Tip110 miRNA and result in more Tip110 expression. When those cells begin to differentiate, Tip110 expression follows the similar regulatory mechanism to that in freshly isolated quiescent CD34+. In addition, there is a self- regulatory feedback loop for Tip110 expression.
Footnotes
Author Disclosure Statement
No competing financial interests exist.