
Abstract
Various microenvironments influence the multiple differentiation potential of mesenchymal stromal cells. For example, inflammatory microenvironment can suppress the myogenic differentiation capability of laryngeal mucosa mesenchymal stromal cells (LM-MSCs). The present study therefore sought to identify the underlying molecular mechanisms regulating these processes. We isolated a novel population of MSCs, LM-MSCs, from the laryngeal mucosa tissues. The cells were cultured in osteogenic, adipogenic, and myogenic differentiation media in the presence or absence of interleukin-1β and tumor necrosis factor α (to simulate inflammatory microenvironment). The expression of active β-catenin, p-GSK3β, and GSK3β were detected by western blot and real-time polymerase chain reaction. The myogenic differentiation of LM-MSCs in inflammatory microenvironment and the regulation by Dickkopf-1 (DKK1) were tested both in vivo and in vitro. Inflammatory microenvironment could suppress the osteogenesis, adipogenesis, and myogenesis of LM-MSCs. The Wnt/β-catenin signaling pathway was activated during myogenesis in inflammatory microenvironment. The suppressed myogenic differentiation capability of LM-MSCs in inflammatory microenvironment was reversed by DKK1. By regulating the Wnt/β-catenin signaling pathway, DKK1 can improve the myogenic differentiation of LM-MSCs in inflammatory microenvironment. Thus, the results of this study may help improve the efficacy of LM-MSCs injection therapy for vocal fold regeneration.
Introduction
M
It is generally accepted that tissue regeneration is necessary in areas of the body where there is significant damage and hence, an inflammatory reaction [9]. The inflammatory cytokines, interleukin-1β (IL-1β) and tumor necrosis factor α (TNF-α), were upregulated in the acute phase of the rat vocal fold wound healing [10]. Additionally, IL-1β and TNF-α are likely to be present in joints where cartilage is undergoing attempted repair or regeneration [11]. MSCs are thought to have a strong potential for tissue regeneration due to their unique properties of self-renewal and their multiple differentiation capability [12]. Inflammatory microenvironment has been shown to affect the functions of MSCs, including cell proliferation and multilineage differentiation [13 –15]. The osteogenic differentiation of the MSC population was suppressed by IL-1β and TNF-α [9]. It is demonstrated that local dentin/pulp inflammation interfered with odontoblast differentiation and dentin repair [16]. It has also been shown that fetal liver hematopoietic progenitor cells express the IL-1 receptor, type 1, and that IL-1 increases fetal liver hematopoiesis and progenitor cell activity, thereby promotes hematopoietic cell survival [17]. In the brain, TNF-α was shown to influence the proliferation, survival, and neuronal differentiation of neural stromal/progenitor cells [18 –20]. Thus, to achieve successful stromal cell-mediated repair, it is necessary to overcome inflammation-mediated inhibition of tissue regeneration.
The Wnt signaling pathway regulates the development, cell proliferation, multilineage differentiation, and cell fate during embryonic development and tissue homeostasis [21,22]. Human birth defects, cancer, and other diseases are associated with genetic mutations of Wnt pathway [23,24]. Among the Wnt pathways, the canonical Wnt/β-catenin signaling pathway has been extensively studied, and it controls gene expression by stabilizing β-catenin protein. The Wnt/β-catenin signaling pathway was also shown to be involved in the regulation of inflammatory processes and associated with tissue/organ injury [25,26]. The pathway can be activated by Wnt3a or inhibited by Dickkopf-1 (DKK1). Blocking the Wnt/β-catenin signaling pathway can restore the osteogenic differentiation of periodontal ligament stromal cells under inflammatory conditions [27].
From the evidence above, we became intrigued by the influence of inflammatory microenvironment on the multilineage differentiation of LM-MSCs, especially the myogenic differentiation capability, and its potential mechanisms. The purpose of this study was to improve the myogenic differentiation capability of LM-MSCs by regulating the Wnt/β-catenin signaling pathway. This investigation may help improve the efficacy of LM-MSCs injection therapy for vocal fold scarring.
Materials and Methods
Materials
Minimum essential medium α (α-MEM), fetal bovine serum (FBS), and glutamine were purchased from Invitrogen. Type I collagenase was purchased from Gibco. The Cell Proliferation Kit (MTT) was purchased from Sigma (St. Louis, MO). PE or FITC-conjugated antibodies for human CD105 (PE), CD14 (PE), CD90 (FITC), CD73 (FITC), CD45 (FITC), and CD31 (PE) were purchased from eBioscience. Antibodies for human CD44 (PE) and CD34 (PE) were purchased from BioLegend. IL-1β, TNF-α, and DKK1 were purchased from PeproTech. 5-Azacytidine (5-Aza) was purchased from Melonepharma. Primary antibodies, such as p-GSK3β was purchased from Cell signaling Technology; active β-catenin was purchased from Millipore; Myod1, Myogenin, and MyHC were purchased from R&D system; and β-actin was purchased from Santa Cruz Biotechnology. Secondary conjugated horseradish peroxidase (HRP) antibodies were purchased from Beyotime.
Study samples and ethics statement
The human laryngeal mucosa samples (n = 6) were isolated from patients undergoing surgery for laryngopharyngeal carcinoma (all males, 49–60 years of age) and were free from any recent clinical acute or chronic infections. Before the investigation, the participants were informed about the objectives of this study and signed the informed consent forms. Ethical approval was obtained from the Institutional Review Board at Xijing Hospital, Fourth Military Medical University.
Cell isolation and culture
LM-MSCs were isolated as previously described [8,28]. Briefly, the harvested normal laryngeal mucosa were excised and rinsed with phosphate-buffered saline containing 1% penicillin, streptomycin, and 1% amphotericin B. After the blood and muscle tissues were removed, the mucosa was cut into 1-cm-wide stripe and placed into centrifuge tubes. Dispase (Gibco BRL) was added to the tube for 30 min at 37°C, then the stripped mucosa was tore to remove epidermal cells. The rest of lamina propia were then digested with type I collagenase at 37°C for 50 min. Cell suspension was obtained by removing undigested tissue pieces through a 100 μm cell strainer. The cells were then cultured in α-MEM supplemented with 10% FBS, 0.292 mg/mL glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C. After 3 days, cells were adherent and passaged at 70%–80% confluency. All cells used for our experiments were in their third passage.
Colony-forming and cell proliferation assay
Cells were cultured in 10-cm diameter culture dishes at 2,500 cells per well for 2 weeks. Cell colonies were fixed with 4% paraformaldehyde and stained with 0.1% Toluidine Blue. For proliferation analysis, the third passage LM-MSCs were cultured into 96-well plates at the density of 1,000 cells per well for 8 days, MTT assay was carried out for 8 days according to the Cell Proliferation Kit protocol. The plate was read at 570 nm by a spectrophotometer.
Flow cytometry analysis
LM-MSCs were stained with antibodies for stromal cell surface markers and analyzed by flow cytometry. Of these, 5 × 105 cells were incubated with PE or FITC-conjugated antibodies for human CD105, CD14, CD90, CD31, CD44, CD73, and CD45 at 4°C. The samples were measured by flow cytometric analysis using a Beckman Coulter Epics XL cytometer (Beckman Coulter, Fullerton, CA). The experiment was repeated at least three times.
Osteogenesis
Cells were cultured in 6-well plates at a density of 2 × 105 cells per well. To induce osteogenic differentiation, 5 mM β-glycerophosphate, 100 nM dexamethasone, and 50 mg/mL ascorbic acid were added to the medium. To evaluate inflammation-related changes in the osteogenic potential of human LM-MSCs, IL-1β (5 ng/mL) and TNF-α (10 ng/mL) were added to the osteogenic differentiation medium to generate an inflammatory microenvironment. After induction for 7 days, alkaline phosphatase (ALP) staining was performed with the BCIP/NBT Alkaline Phosphatase Color Development Kit (Beyotime Co., Shanghai, China) following the standard protocol, and the quantification assay was performed using Plus 5.0 software. After 21 days of osteogenic differentiation, 1% Alizarin Red S (Sigma) was used to detect calcium accumulation according to the manufacturer's suggested protocol. Then, Alizarin Red was incubated with 100 mM cetylpyridinium chloride (Sigma) for 30 min at room temperature and quantified by spectrophotometer at 560 nm for statistical analysis.
Adipogenesis
Cells were seeded into 6-well plates at a density of 2 × 105 cells per well and maintained in α-MEM. Upon reaching confluency, cells were incubated in adipogenic medium [10% FBS, 2 mM insulin, 0.5 mM isobutylmethylxanthine; Sigma) and 10 nM dexamethasone (Sigma)] in the presence or absence of IL-1β (5 ng/mL) and TNF-α (10 ng/mL). After 14 days of adipogenic differentiation, the cells were fixed with 4% paraformaldehyde for 20 min and stained with 0.3% Oil Red O. Intracellular droplets were dissolved in isopropanol and absorbance was quantitatively measured at 560 nm for statistical analysis.
Myogenesis
To induce myogenesis, cells were cultured in α-MEM with 10 μM 5-Aza for 24 h, and then changed to the medium supplemented with 5% horse serum (Gibco, BRL) without 5-Aza for the follow-up experiment. The medium was changed every 3 days for the entire experimental time course. For different groups, corresponding cytokines were added: (1) control group: α-MEM without any cytokine; (2) DDK1 group: α-MEM with DKK1 (100 ng/mL), IL-1β (5 ng/mL), and TNF-α (10 ng/mL); (3) infla group: α-MEM with IL-1β (5 ng/mL) and TNF-α (10 ng/mL); (4) wnt3a group: α-MEM with Wnt3a (25 ng/mL).
Real-time polymerase chain reaction
Total RNA was isolated using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. Reverse transcription polymerase chain reaction (RT-PCR) was performed using the QuantiTect SYBR Green PCR Kit (Toyobo, Osaka, Japan) and the Applied Biosystems 7500 Real-Time PCR Detection System following the manufacturer's protocol (Applied Biosystems, Darmstadt, Germany). All samples were run in triplicates and cDNA levels were normalized to those of GAPDH. The sequences of primers used are shown in Table 1.
Western blot analyses
Total protein was extracted from the cells by cell lysis buffer (10 mM Tris–HCl,1 mM EDTA, 1% sodium dodecyl sulfate, 1% Nonidet P-40, 1:100 proteinase inhibitor cocktail, 50 mM β-glycerophosphate, and 50 mM sodium fluoride). For each sample, aliquots of 20–50 μg of the cell lysates were separated by 10% SDS-PAGE and transferred to a PVDF membrane. The membranes were blocked with 5% nonfat milk powder for 2 h and incubated with the following primary antibodies: p-GSK3β (1:1,000), active β-catenin (1:1,000), Myod1 (1:1,000), Myogenin (1:1,000), MyHC (1:1,000), and β-actin (1:4,000). After incubation with HRP-conjugated secondary antibodies (1:4,000) for 2 h at room temperature, the blots were visualized using an Enhanced Chemiluminescence Kit (Amersham Biosciences, Piscataway, NJ).
In vivo experiments
The effects of the Wnt/β-catenin signaling pathway and inflammatory microenvironment on the myogenesis of human LM-MSCs were also investigated in vivo. Animal protocols were approved by the Animal Care Committee of Fourth Military Medical University. LM-MSCs were cultured in medium for 3 weeks to induce myogenesis, as described above: (1) control group: α-MEM without any cytokine; (2) DDK1 group: α-MEM with DKK1 (100 ng/mL), IL-1β (5 ng/mL), and TNF-α (10 ng/mL); (3) infla group: α-MEM with IL-1β (5 ng/mL) and TNF-α (10 ng/mL); (4) wnt3a group: α-MEM with Wnt3a (25 ng/mL). Cells of each group were resuspended in 0.5 mL type I collagen. The mixture of cells and collagen were incubated at 37°C for 30 min, and were injected subcutaneously under the dorsal skin of immunocompromised nude mice (4-week-old males; n = 3, each group; Fourth Military Medical University, Animal Center, Xi'an, China). Two weeks postimplantation, mice were sacrificed and the implant was taken out, fixed in 4% paraformaldehyde and embedded in paraffin. Then, the samples were sectioned and subjected to Hematoxylin–Eosin, Masson's Trichrome, and immunohistochemical analysis.
Immunofluorescence
After rehydration, the sections were boiled in sodium citrate buffer (pH 6.0) for antigen retrieval, permeabilized with 0.2% Triton X-100, and blocked in 1.5% goat serum at 37°C for 20 min, then, incubated in primary antibody overnight at 4°C. Primary antibodies used in this study included mouse Myod1 (1:150; R&D), mouse myogenin (1:150; R&D), and mouse MyHC (1:150; R&D). The samples were treated with fluorescence-labeled secondary antibodies and the nuclei were stained with Hoechst (Biyuntian, China). All samples were examined under a confocal microscope (FV1000; Olympus, Japan) and were analyzed using the National Institutes of Health ImageJ software.
Statistical analysis
All results are presented as mean with standard deviation. For the analysis of two unpaired groups, a Student's t-test was used. For the analysis of multiple groups, a one-way ANOVA was used. We utilized statistical program for social sciences (SPSS) software for statistical tests. All experiments were repeated at least three times, and representative experiments are shown. Differences were considered to be significant when P < 0.05.
Results
The biological character of LM-MSCs
LM-MSCs grow in an adherent way, presenting a stretched, short, spindle-shaped and radial arrangement (Fig. 1A). They displayed colony-forming ability and can proliferate during culture (Fig. 1B, C). LM-MSCs express the markers of mesenchymal stromal cell, including CD44, CD90, CD105, and CD73. On the contrary, they were negative for hematopoietic cell marker CD34, monocytes maker CD14, platelet endothelial cell maker CD31, and pan-leukocyte marker CD45 (Fig. 1D).
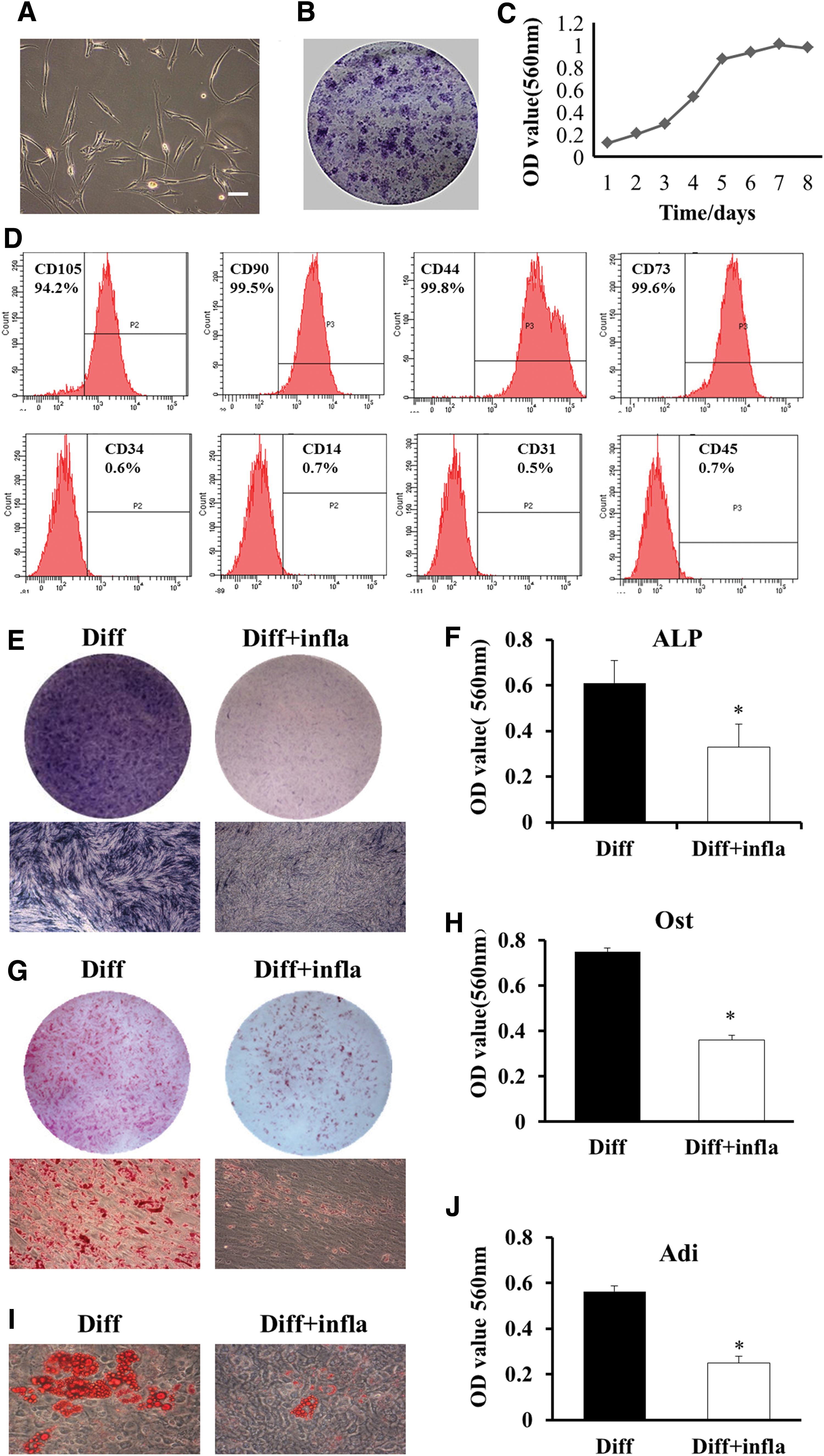
Characterization of LM-MSCs and the effect of inflammatory microenvironment on the differentiation capability of LM-MSCs. Inflammatory microenvironment suppressed the osteogenic and adipogenic differentiation capacity of LM-MSCs. LM-MSCs were obtained and can be cultured and proliferated in vitro
Inflammatory microenvironment suppresses the osteogenesis and adipogenesis of LM-MSCs
Experiments were performed to confirm that laryngeal mucosa mesenchymal stromal cells could differentiate into bone, fat, and cartilage (Supplementary Fig. S1; Supplementary Data are available online at
Inflammatory microenvironment suppressed the myogenic differentiation of LM-MSCs
The transition of myogenic precursors into differentiated cells and of stromal cells into myogenic precursor cells can be confirmed by the expression of several transcription factors, including Myod1, Myf-5, myogenin, and structural proteins, such as myosin heavy chain (MyHC) [29 –33]. Protein expression of Myod1 and myogenin (Myog) was observed at 1 week of induction with control medium or with myogenic inductive medium (Fig. 2A), whereas the mRNA levels of Myod1 and myogenin in myogenic inductive medium were higher than that in the control medium (Fig. 2B), which indicates the initiation of myogenic differentiation. The genes and proteins of MyHC expression was restricted to later induction time points (3 and 6 weeks only), and indicate that the myoblasts have been terminally differentiated (Fig. 2A, B). In addition, western blot was used to detect the related proteins at the corresponding time point. The protein levels of Myod1 and myogenin started to increase from 1 week and lasted to 6 weeks (Fig. 2C), whereas the expression of MyHC increased at later induction time points (3 and 6 weeks) (Fig. 2D).
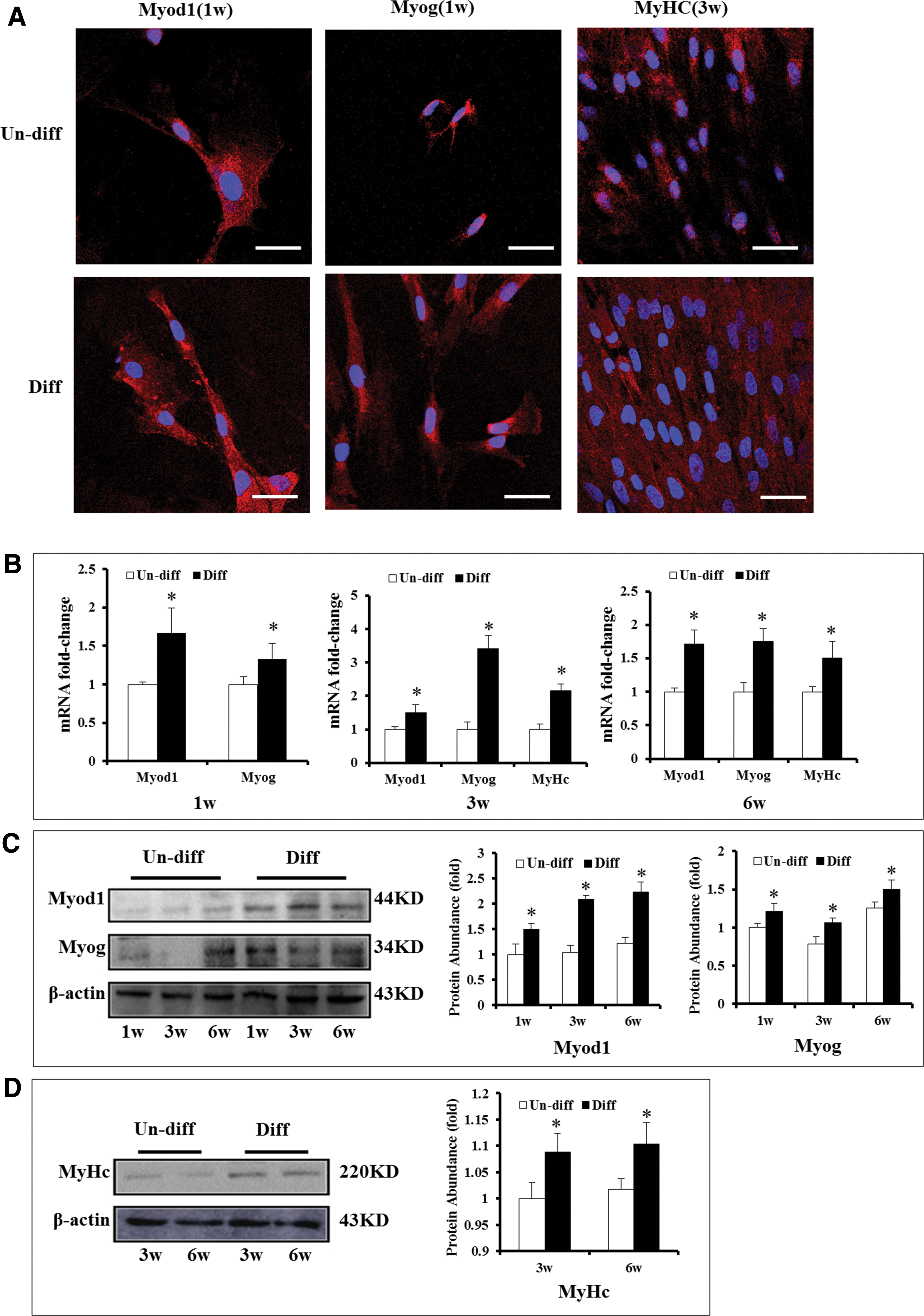
LM-MSCs express the markers of myoblast after culturing in myogenic differentiation medium.
To further identify the influence of inflammatory microenvironment on the myogenic differentiation capability of LM-MSCs, the inflammatory cytokines IL-1β and TNF-α were added to the myogenic induction medium to simulate the inflammatory microenvironment. RNA and protein levels of Myod1, myogenin, and MyHC were detected by RT-PCR and western blot, respectively, at corresponding time points during differentiation. The genes and protein of Myod1 and myogenin, were expressed from 1 to 6 weeks during the myogenic differentiation. Compared with the control conditions, the genes and protein levels were inhibited in the inflammatory microenvironment at 1, 3, and 6 weeks (Fig. 3 A, B, D). The mRNA and protein of MyHC were also affected after myogenic differentiation for 3 weeks in myogenic inductive medium with inflammatory factors (Fig. 3C, E).
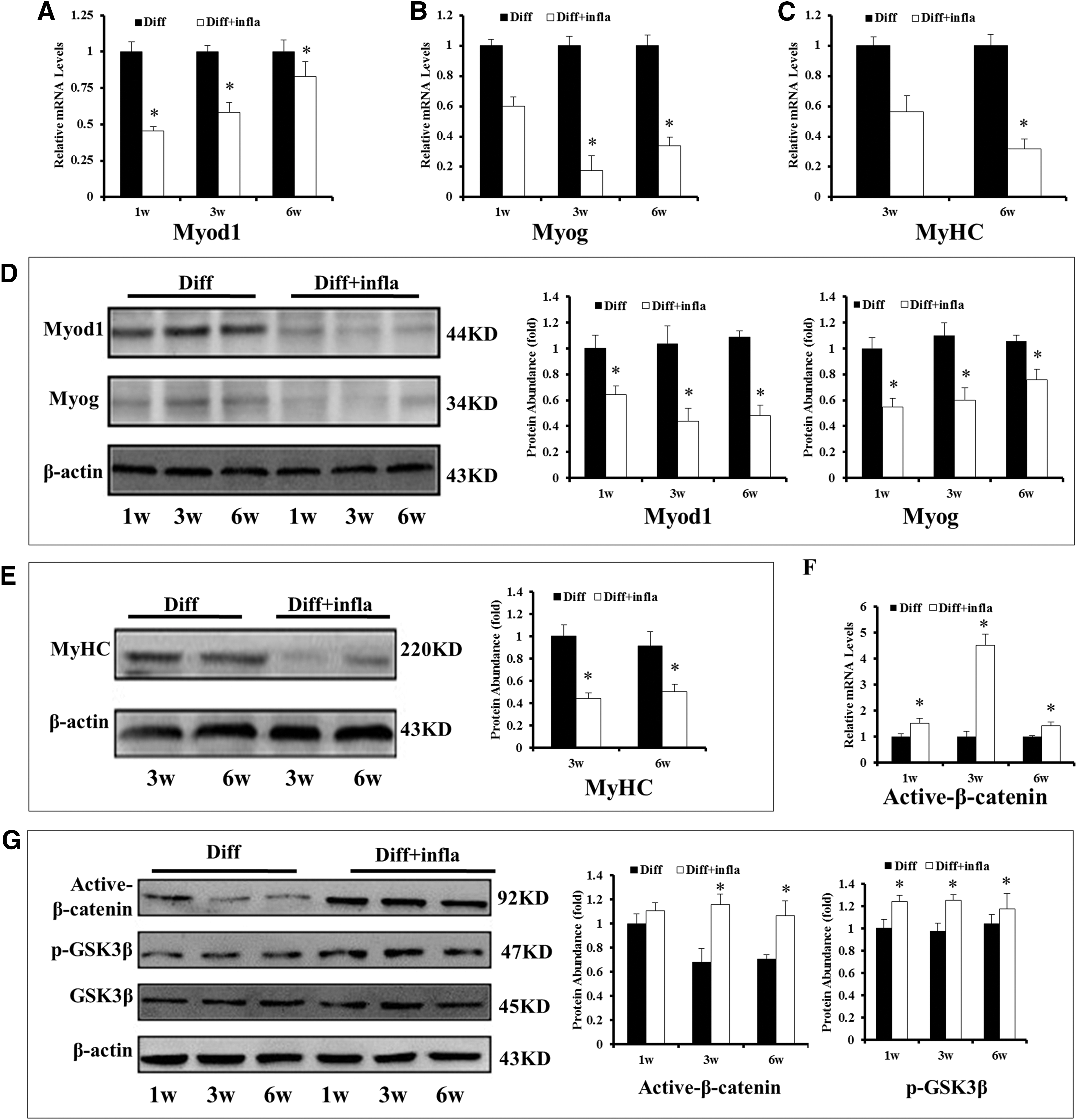
The Wnt/β-catenin signaling pathway was activated under inflammatory microenvironment when LM-MSCs were myogenic differentiated. LM-MSCs treated with or without IL-1β (5 ng/mL) and TNF-α (10 ng/mL) were cultured in myogenic medium for up to 6 weeks, the genes and proteins of Myod1, myogenin, and MyHC were tested, respectively.
Some studies showed that the Wnt/β-catenin pathway could be activated upon inflammatory stimulation [25,34]. Thus, we wonder whether impaired myogenic differentiation was accompanied by the activation of the Wnt/β-catenin pathway, thereby, resulting in the activation of β-catenin and inhibition of GSK3β kinase [35]. To determine the role of Wnt under these conditions, first, we compared the active β-catenin and GSK3β kinase between the control condition and inflammatory microenvironment. Our data showed that the expression of active β-catenin and phosphorylated GSK3β (p-GSK3β) kinase were enhanced in the inflammatory microenvironment, whereas the total levels of GSK3β kinase had no significant change (Fig. 3F, G), and indicate that the Wnt/β-catenin pathway was upregulated during the myogenesis of LM-MSCs in the inflammatory microenvironment.
Inflammatory microenvironment affects the myogenesis of LM-MSCs through the Wnt/β-catenin pathway
To further explore the role of Wnt/β-catenin pathway in the myogenic differentiation of LM-MSCs, wnt3a was used to activate the Wnt/β-catenin pathway at a concentration of 25 ng/mL. Total RNA and protein were extracted and detected at the corresponding time points. The results showed that at 1 week, the genes and proteins expression of Myod1 and myogenin in control group were higher than that in wnt3a group (Fig. 4A, B, I). The genes and protein of MyHC started to express at 3 week, and at this time point, the levels of Myod1, myogenin and MyHC were higher in control group than in wnt3a stimulation group (Fig. 4C–E, J), as well as the results at 6 week (Fig. 4F–H, K). Above, those results indicate that wnt3a stimulation had a similar effect on LM-MSCs as inflammatory microenvironment.
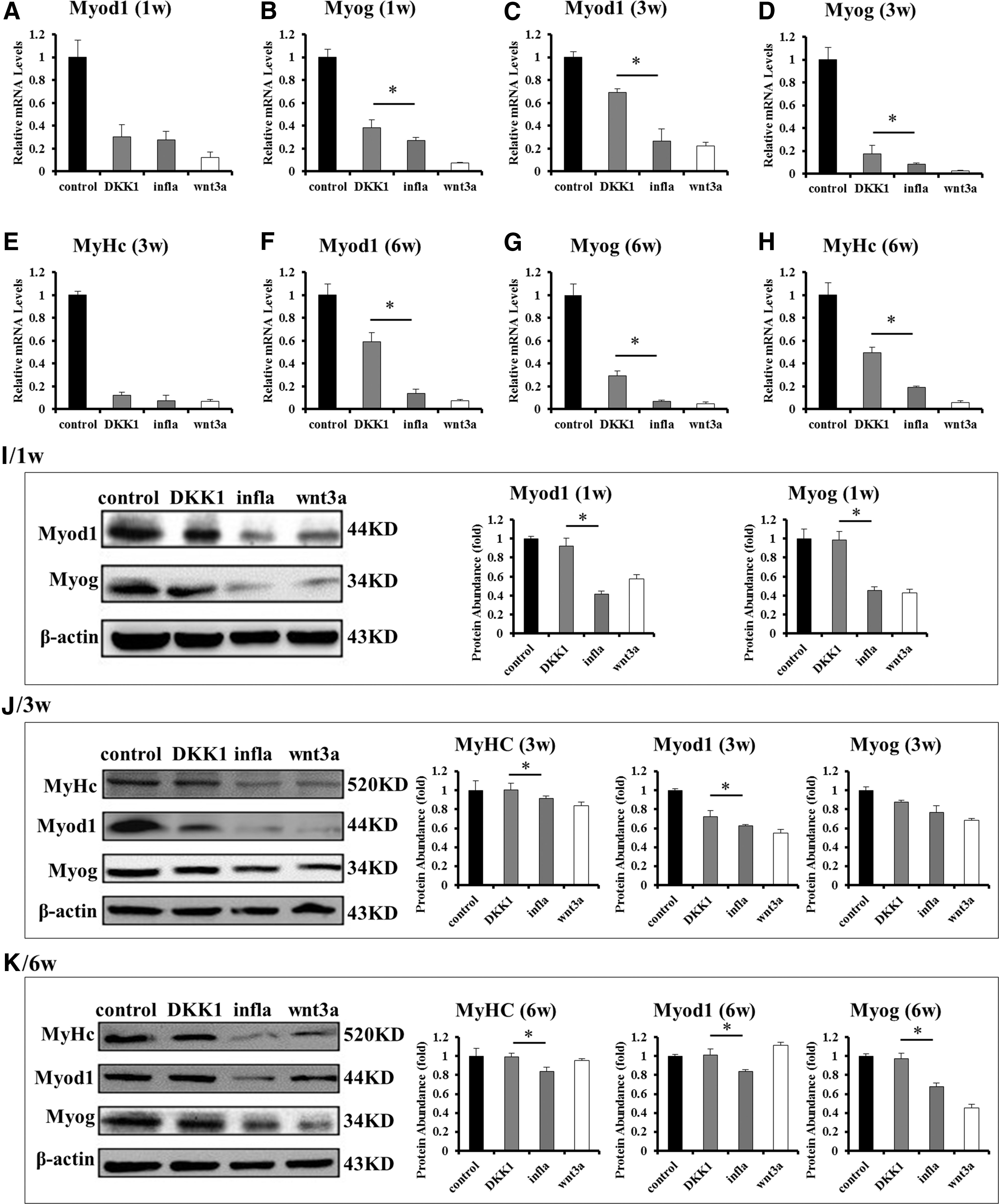
The Wnt/β-catenin signaling pathway mediated the myogenic differentiation of LM-MSCs.
To reverse the inhibitory effect of the IL-1β and TNF-α on the myogenic differentiation of LM-MSCs, we inhibited the Wnt/β-catenin pathway by adding DKK1 to the IL-1β and TNF-α stimulation culture medium. At 1 week, the gene and protein levels of Myod1 and myogenin were higher in DKK1 group compared with the inflammatory group (Fig. 4A, B, I). At 3 weeks, the gene and protein levels of Myod1, myogenin, and MyHC in DKK1 group were higher than the inflammatory group, whereas lower than control (Fig. 4C–E, J). These results were in accordance with the results at 6 weeks (Fig. 4F–H, K). The results mentioned above revealed that the inhibitory effect of inflammatory microenvironment on the myogenic differentiation of LM-MSCs was partially reversed by DKK1.
The Wnt/β-catenin signaling pathway mediates the myogenic differentiation of LM-MSCs in an inflammatory microenvironment in vivo
To assess the effects of inflammatory stimuli and the modulation of Wnt/β-catenin signaling pathway on LM-MSCs in vivo, the myogenic differentiation of LM-MSCs was analyzed in mice. Following the differentiation of LM-MSCs in vitro for 3 weeks, collagen type I was used as a carrier for implantation. The mixture of cells and collagen were injected under the dorsal skin of immunocompromised nude mice. No health deficits were observed in any of the host animals throughout the study period.
The number of myoblasts was decreased both in the inflammatory group and wnt3a group compared with control (Fig. 5A, B). However, the decreased number of myoblasts was partially reversed following the addition of DKK1 (Fig. 5A, B). Furthermore, the levels of myogenic differentiation-associated protein, Myod1, myogenin, and MyHC were also altered under different conditions (Fig. 5C–E).

The Wnt/β-catenin pathway affects the myogenic differentiation of LM-MSCs in an inflammatory microenvironment in vivo. LM-MSCs were cultured in different media for 3 weeks to induce myogenesis. The mixture of cells and collagen were injected subcutaneously under the dorsal skin of immunocompromised nude mice. Two weeks postimplantation, mice were sacrificed and the implant was taken out and processed for Hematoxylin–Eosin staining
Discussion
The aim of this study was to investigate the myogenic differentiation capability of LM-MSCs in an inflammatory microenvironment and alter its effects by addition of the inhibitor of the Wnt/β-catenin signaling pathway, DKK1.
In our previous work, we isolated and cultured the laryngeal mucosa to obtain LM-MSCs. When used for vocal fold injury, the cells were injected into damaged vocal folds and they can help improve the reconstruction microenvironment. LM-MSCs have the potent ability to differentiate into myofibroblasts and fibroblasts, and they can slow down the rapid decrease of elastic fiber and hyaluronic acid, thereby prevent the formation of vocal fold scar. These previous results indicated that LM-MSCs became the cells of the vocal fold muscular layer in vivo, which provided evidence that the LM-MSCs effectively promote healing of the vocal folds [8,35].
During tissue regeneration, the defective or injured tissues are often under the condition of increased inflammatory cytokines [16,36,37]. Extensive evidence has suggested that proinflammatory cytokines affect the multiple differentiation potential of stromal cells and tissue repair [16,38 –40]. Thus, the study of the impact of the microenvironment on the functions of the stromal cells has attracted interests within the field. In this study, the inflammatory factors IL-1β and TNF-α were used to help determine the influence of inflammatory microenvironment on the multilineage differentiation of LM-MSCs. These results indicated that inflammatory microenvironment could suppress the osteogenesis, adipogenesis, and myogenesis of LM-MSCs. The inhibition of myogenesis in LM-MSCs may affect their repair function following vocal fold injury.
We further explored the underlying molecular mechanisms that regulate the suppression of myogenesis in LM-MSCs under inflammatory condition. Our results suggested that, the Wnt/β-catenin signaling pathway was functionally activated in inflammatory microenvironment. Recently, investigations revealed that the Wnt/β-catenin signaling pathway represses various stromal cell differentiation processes, such as odontoblast-like differentiation of dental pulp stromal cells or osteogenic differentiation of adipose-derived MSCs [41,42], which is consistent with the findings of our study. Under certain conditions, the Wnt/β-catenin signaling pathway may be an important target for inhibiting myogenesis in LM-MSCs in an inflammatory microenvironment. β-catenin and p-GSK3β kinase are the key components of a regulatory switch in the Wnt/β-catenin signaling pathway. In our study, we found that the expression of active β-catenin and p-GSK3β kinase in inflammatory microenvironment was significantly increased, which indicated that the Wnt/β-catenin signaling pathway was activated in inflammatory microenvironment.
When the LM-MSCs were treated with myogenic inductive differentiation medium with wnt3a or with DKK1, IL-1β, and TNF-α, the results showed that the capability of myogenic differentiation was weakened by wnt3a in the control conditions, and can be rescued by DKK1 in inflammatory microenvironment. There is a similar trend between the results of the inflammatory cytokine group and wnt3a group both in vivo and in vitro. Both of these facilitated the activation of the Wnt/β-catenin signaling pathway and suppressed the myogenic differentiation of LM-MSCs. Finally, after the addition of DKK1, the myogenic differentiation of LM-MSCs in an inflammatory microenvironment could be reversed.
The current study showed that inflammatory microenvironment could suppress osteogenesis, adipogenesis, and myogenesis of LM-MSCs, and the Wnt/β-catenin signaling pathway was activated during myogenesis in an inflammatory microenvironment. The myogenic differentiation capability of LM-MSCs in the inflammatory microenvironment could be reversed by DKK1. Thus, these results may help improve the efficacy of using LM-MSCs injection therapy for vocal fold scarring.
Footnotes
Acknowledgment
This study was supported by funding support from the National Nature Science Foundation of China (81470679, 31670995, 81670915, and 81271065).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.