
Abstract
To date, little is known about the mechanisms underlying the self-renewal of embryonic stem (ES) cells from fish species. In this study, we report that the leukemia inhibitory factor (LIF; named as OnLif) from a teleost fish, Nile tilapia (Oreochromis niloticus), is essential for the proliferation, survival, and pluripotency maintenance of Nile tilapia ES cells (TES1) by activating the signal transducer and activator of transcription 3 (Stat3). This protein has 221 amino acid residues with similar sequence features to mammalian LIF. By fusing to a small ubiquitin-related modifier and inducing expression at 16°C, the soluble tag-free protein had been successfully obtained. Further investigation indicates that OnLif could significantly enhance the proliferation and survival of TES1. Moreover, it contributed to the pluripotency maintenance of TES1 characteristic of high expression of pluripotency genes, no or low expression of differentiation genes, and strong alkaline phosphatase activity. Notably, it mediated Stat3 phosphorylation, whose inhibitor treatment could lead to apoptosis. In addition, OnLif significantly enhanced the proliferation of ES cells from medaka (Oryzias latipes), suggesting its potential role in other fish ES cells. These data first suggest that Lif/Stat3 signaling has an essential role in the self-renewal of ES cells from fish, just like that in the ground state pluripotency maintenance of mouse and human ES cells. Our study would not only be helpful for the understanding of molecular mechanisms underlying self-renewal of ES cells from the perspective of evolution but also facilitate ES-based biotechnology application in fishery.
Introduction
L
Accumulative studies reveal that LIF is critical for pluripotency maintenance of mouse ES cells and iPSCs by activating the signal transducer and activator of transcription 3 (STAT3) [7,11 –15]. LIF binds to its signaling receptor (LIF-R) and recruits glycoprotein 130 to form a heterodimer; upon dimerization, the signaling receptors recruit and phosphorylate JAKs, which, in turn, phosphorylate STAT3; the phosphorylated STAT3 (pSTAT3) dimerizes, translocates to the nucleus, and regulates the transcription of a wide range of target genes, including pluripotency factors such as KLF4, GBX2, and TFCP2L1, which confer varying degrees of self-renewal [7,12,16]. In humans, it had been perceived that LIF/STAT3 signaling was not required for the pluripotency maintenance in ES cells [13,14,17 –20]. Nevertheless, recent studies reveal that human ES cells in ground state and primed state rely on distinct signaling environments [12,21 –24]. The LIF/STAT3 signaling is dispensable for human ES cells in the primed state, but not in the ground state; furthermore, in the presence of LIF/STAT3 stimulation, human ES cells in the primed state could be conversed to the ground state, suggesting its conservation in ground state pluripotency maintenance among mice and humans [24 –26]. However, whether the fundamental program of pluripotency maintenance is conserved across vertebrates still remains to be elucidated.
To date, our understanding about the role of Lif/Stat3 in the self-renewal of ES cells from ectothermic vertebrates just like fish is lacking, which would be very informative for our understanding of the molecular mechanisms underlying pluripotency maintenance from the perspective of evolution. In fish, the Lif orthologue has been identified in zebrafish (Danio renio); functional experiments reveal that it is critical for neural development [27 –29]. However, whether Lif plays a role in fish stem cells remains unknown. Although a previous study has reported that murine LIF has no effect on medaka fish (Oryzias latipes) ES cells [30], considering that the sequence of LIF varies greatly between species, it is desirable that fish genes are studied in fish stem cells.
In our previous studies, an ES cell line from the Nile tilapia (Oreochromis niloticus) (named as TES1) has been successfully established, which is characteristic of stable growth over 200 days of culture, the potential of self-renewal, and differentiation in vitro and in vivo, providing an excellent model to investigate the mechanisms underlying self-renewal of fish ES cells [31]; furthermore, the key pluripotency regulator Pou5f3 from Nile tilapia and its biological activity in pluripotency maintenance has been identified [32], providing a good tool to monitor cell pluripotency. In this study, we first demonstrate that the Nile tilapia Lif (named as OnLif hereafter) plays a crucial role in the proliferation, survival, and pluripotency maintenance of Nile tilapia ES cells by activating Stat3.
Materials and Methods
Reverse transcription–polymerase chain reaction
Nile tilapias were kept in recirculating aerated water tanks at 26°C under an artificial photoperiod of 12-h light to 12-h darkness as previously described [32]. Embryos at 12, 24, 36, 48, 72, 96, and 120 hpf (hours postfertilization), adult tissues, including brain, pituitary, gill, heart, spleen, liver, intestine, ovary, testis, kidney, muscle and head kidney, and TES1 cells were collected and stored at −80°C until further processing. Total RNAs were extracted from the above-mentioned samples and subsequently reverse-transcribed into complementary DNAs (cDNAs) using the PrimeScript II 1st Strand cDNA Synthesis Kit (Takara) as previously described [33]. Each polymerase chain reaction (PCR) primer set was designed and validated for use by gel analysis (Supplementary Table S1; Supplementary Data are available online at
Sequence analyses
Comprehensive analyses of OnLif were done as previously described [33]. The amino acid identity was calculated with the program Matrix Global Alignment Tool (MatGAT) V2.0.3. The phylogenetic tree was constructed using neighbor-joining algorithm within MEGA version 5.0. Chromosomal synteny was calculated through the Ensembl Genome Browser (
Prokaryotic expression and purification
The cDNA sequence encoding OnLif deleted of the 31-amino acid signal peptide was inserted into the prokaryotic expression vector pET28a-His6-SUMO, which is derived from pET28a containing a fused tag of His6 and a small ubiquitin-related modifier (SUMO), and finally verified by sequencing. The expression conditions were optimized to obtain soluble recombinant protein. Subsequently, the supernatant was collected, purified by Ni-NTA affinity chromatography, and digested with ULP1. The collected samples were detected by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and western blotting using anti-His tag antibody as previously described [34].
Cell culture
TES1 cells at 35–55 passages were incubated as previously described in TESM [31], which is the Dulbecco's modified Eagle's medium supplemented with 20 mM Hepes, 100 U/mL penicillin and streptomycin, 15% fetal bovine serum, the non-protein supplement combination 5 N (2 mM
Cell Counting Kit-8 cell viability assay
The Cell Counting Kit-8 (CCK8) assay was used to quantitatively assess cell viability as previously described [36]. TES1 cells (∼5,000 cells/well) were seeded in 0.1% gelatin-coated 96-well plates with 5 N containing no or 1, 10, and 100 ng/mL of OnLif, five replicates in each group. At the indicated time, CCK8 solution (Dojindo) was added and measured (Thermo Labsystems, Vantaa, Finland). The proliferative activity of each sample is arbitrarily indicated by the relative absorbance value, which is the ratio of (the absorbance value at 450 nm to the absorbance value at 630 nm) at indicated time/(the absorbance value at 450 nm to the absorbance value at 630 nm) at 0 h. The experiment was repeated for at least thrice.
5-Ethynyl-2-deoxyuridine labeling
To further evaluate the proliferating cells, TES1 cells were first incubated in 5 N containing no or 10 ng/mL OnLif for 24 h, and then exposed to 10 μmol EdU (5-ethynyl-2-deoxyuridine; RIBOBIO, Guangzhou, China) for 4 h. EdU staining was carried out as described previously [36]. The fluorescence signals were observed under Nikon Ti–S inverted fluorescence microscope.
Annexin V-FITC/PI staining
To detect the apoptosis of TES1 cells incubated in 5 N containing no or 10 ng/mL OnLif, Annexin V-FITC/PI kit (BD) was used following the manufacturer's instructions. The fluorescence signals were observed under Nikon Ti–S inverted fluorescence microscope.
Immunocytochemistry
TES1 cells were incubated in TESM and 5 N containing no or 10 ng/mL OnLif for 5 days, respectively. The cells were separately collected and subdivided into three groups. One group was used for further PCR analysis and the other two groups were used for the detection of alkaline phosphatase (AP) activity and Pou5f3 expression as previously described [31,32,36]. Briefly, TES1 cells were fixed with 4% paraformaldehyde and subsequently stained with rabbit anti-OnPou5f3 primary antibody (1:200), PE-conjugated goat anti-rabbit IgG (1:500; Vector Laboratories), and finally Hoechst dye solution (Molecular Probes). For AP activity staining, only the bromochloroindolyl phosphate/nitroblue tetrazolium (Beyotime) was used. The fluorescence signals were observed under Nikon Ti–S inverted fluorescence microscope.
Real-time PCR
The messenger RNA (mRNA) levels of TES1 cells from triple treatments were quantified by real-time PCR using delta-delta Ct (threshold crossing value) calculations, four replicates for each treatment. Primer pairs were gene specific and validated by gel analysis (Supplementary Table S1). Real-time PCR was performed on the StepOnePlus™ Real-Time PCR System (Life Technologies) as previously described [36].
Western blotting
TES1 cells were preincubated in 5 N for 12 h and then subdivided into four groups. One group was continually incubated in 5 N as the control. The other three groups were incubated in 5 N supplemented with 10 ng/mL OnLif along with different concentrations of S3I-201 (100, 50, and 0 μM) for 6 h, respectively, which is a chemical inhibitor of STAT3 activity by blocking its phosphorylation and dimerization. The cells were harvested and then total proteins were extracted using a lysis buffer. Western blotting was performed as previously described [34]. The primary antibodies used were as follows: Tyr708 p-STAT3 (MBL;
Statistical analyses
Statistical analyses were performed with Student's t-test for the comparison between two groups. P < 0.05 or P < 0.01 is considered as significant or very significant difference from the control, respectively.
Results
Sequences analyses
A 1,067-bp lif cDNA sequence from Nile tilapia was obtained through bioinformatics analysis and then its open-reading frame was amplified by reverse transcription–polymerase chain reaction (RT-PCR) and verified by sequencing (Supplementary Fig. S1). It displays a three-exon genomic organization, which is just like zebrafish and human Lif (Fig. 1A). The putative protein OnLif has 221 amino acid residues with similar sequence features to mammalian LIF, including the presence of signal peptide, 2 potential N-glycosylation sites, and 4 cysteine residues (Supplementary Fig. S1). Multiple alignment analysis reveals that OnLif shares identity 71% to flounder, 60% to medaka, 59% to rainbow trout, and 35% to zebrafish, while as low as about 20% identity to mammalian homolog (Fig. 1B and Supplementary Fig. S2). In the phylogenetic tree, OnLif forms a fish-specific clade together with other fish homologues and then clusters with tetrapod clade (Fig. 1C). Cross-species comparison of chromosomal location indicates that the Nile tilapia lif-containing region in LG 7 has good synteny to lif-containing region in zebrafish chromosome 5 and Lif-containing region in human chromosome 22 (Fig. 1D). Taken together, these data suggest that OnLif is orthologous to mammalian LIF.
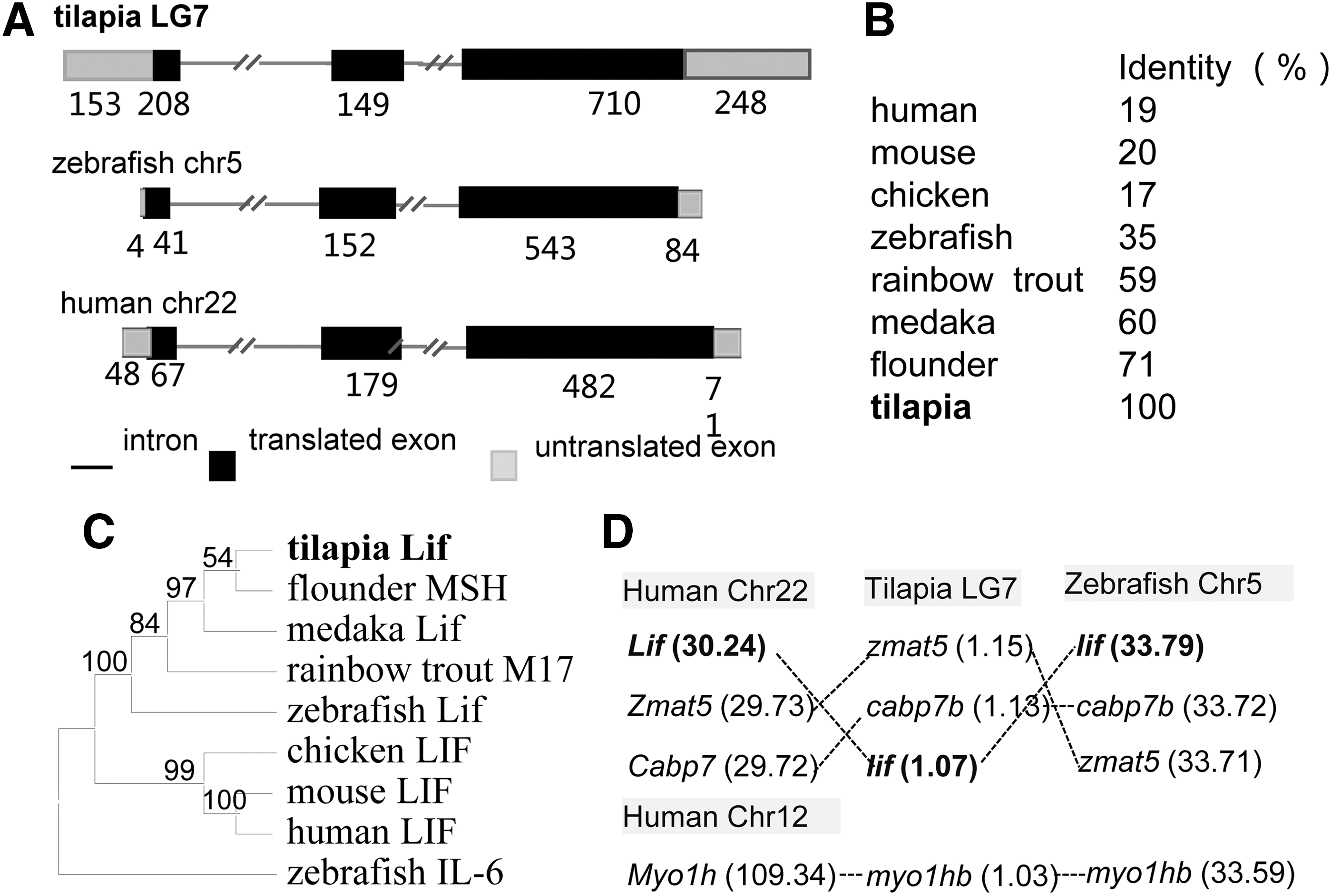
OnLif sequence analyses.
The mRNA expression profile
In mammals, Lif is widely expressed in various tissues, including the brain, heart, liver, gut, kidney, lung, and thymus, while CNTF and CT-1 are preferentially expressed in the nervous system [37]. In zebrafish, the expression of lif occurs in a variety of tissues as well, particularly high in the brain and kidney [27,28]. In this study, the mRNA expression of Onlif was detected in the heart, spleen, intestine, and muscle with high level, in the brain, pituitary, liver, ovary, and head kidney with relative low level, and in the gill, testis, and kidney with barely undetectable level (Fig. 2A). Meanwhile, the mRNA expression of Onlif occurred in developing embryos through 12–120 hpf with relative high level (Fig. 2B), implying its potential roles in embryos from blastula stage onward.

The mRNA expression levels of Onlif in adult tissues
Production of soluble tag-free Onlif protein
The Onlif cDNA sequence deprived of the 31-amino acid signal peptide encoding region was inserted into the prokaryotic expression vector pET28a-His6-SUMO (Fig. 3A). By inducing expression at 16°C, Ni-NTA affinity chromatography, and ULP1 digestion, soluble tag-free OnLif protein (∼21 kDa) had been successfully obtained through the removal of His6-SUMO tag from purified recombinant protein (∼37 kDa) (Fig. 3B). As expected, a strong band compatible with the predicted size was observed in the recombinant protein-containing samples, but not in the purified tag-free OnLif by western blotting detection, consistent with the absence of His6-SUMO tag (Fig. 3C).
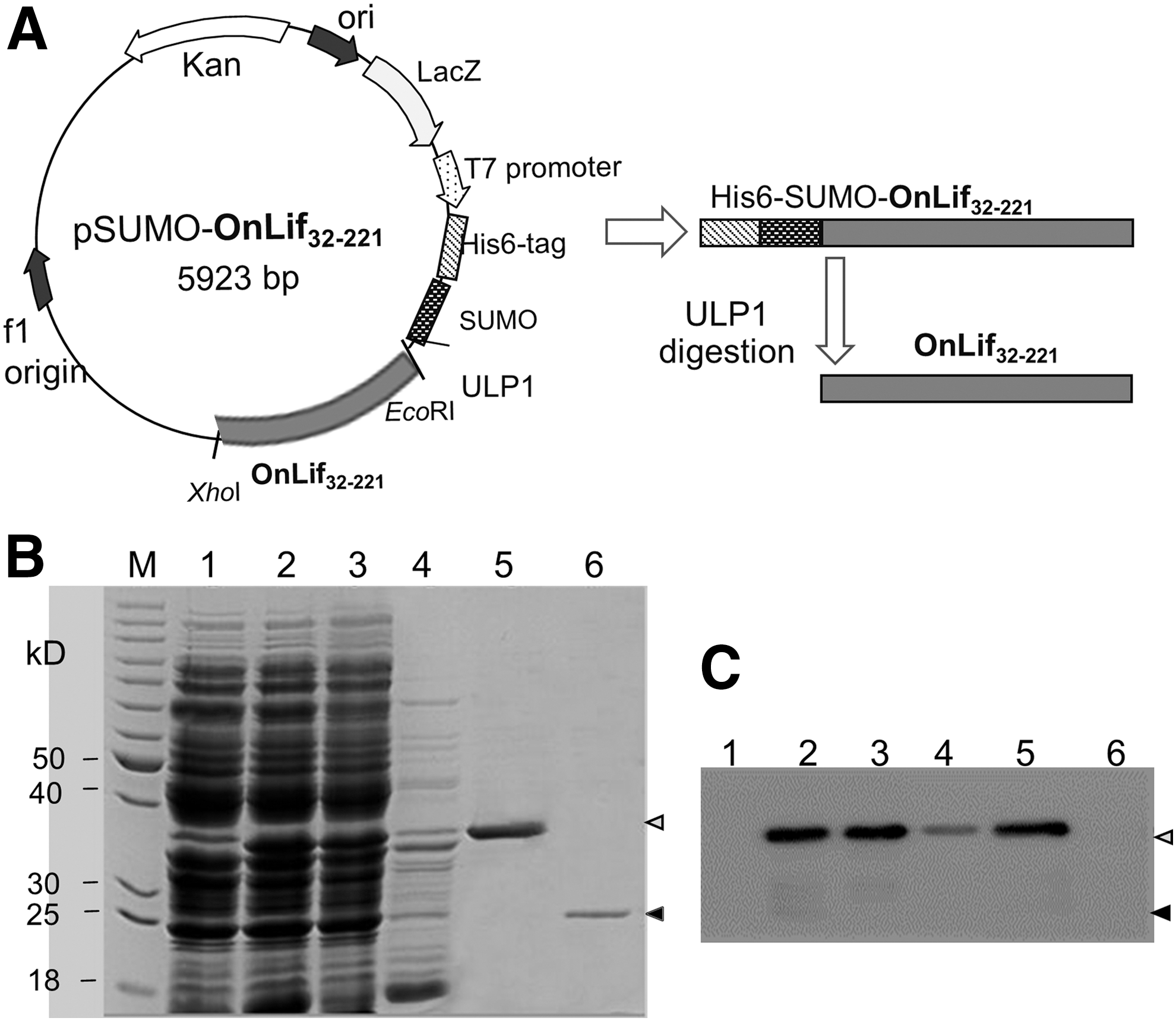
Production and purification of OnLif.
OnLif enhances proliferation of TES1
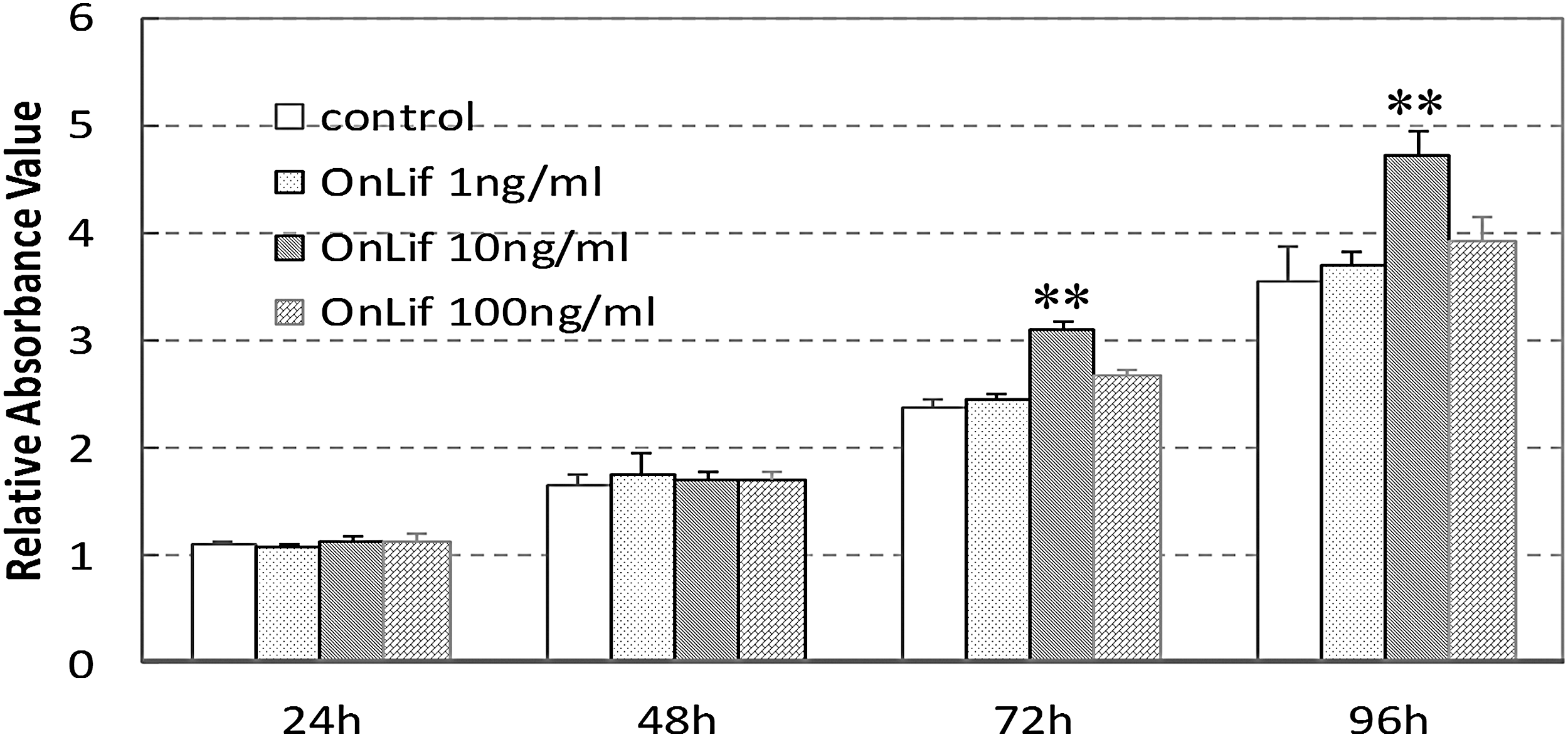
To determine whether OnLif has a significant effect on cell proliferation, the viability of Nile tilapia ES cells (TES1) incubated in 5 N containing no or 1, 10, and 100 ng/mL OnLif was measured by CCK8 assay and EdU staining, respectively. TES1 exhibited no significant proliferative activity in 5 N containing 1 or 100 ng/mL OnLif compared with that in 5 N (P > 0.05), whereas a significant proliferative activity in 5 N containing 10 ng/mL OnLif compared with that in 5 N (P < 0.01) (Fig. 4A). By contrast, commercial murine LIF at any concentration had no proliferative effect on TES1 (data not shown). The proliferative activity of OnLif was further detected by EdU incorporation staining. After a 4-h EdU incubation, the cells displayed about 13% proliferative cells in 5 N, whereas about 24% in 5 N containing 10 ng/mL OnLif (Fig. 4B), consistently indicating the proliferative activity of OnLif on TES1.
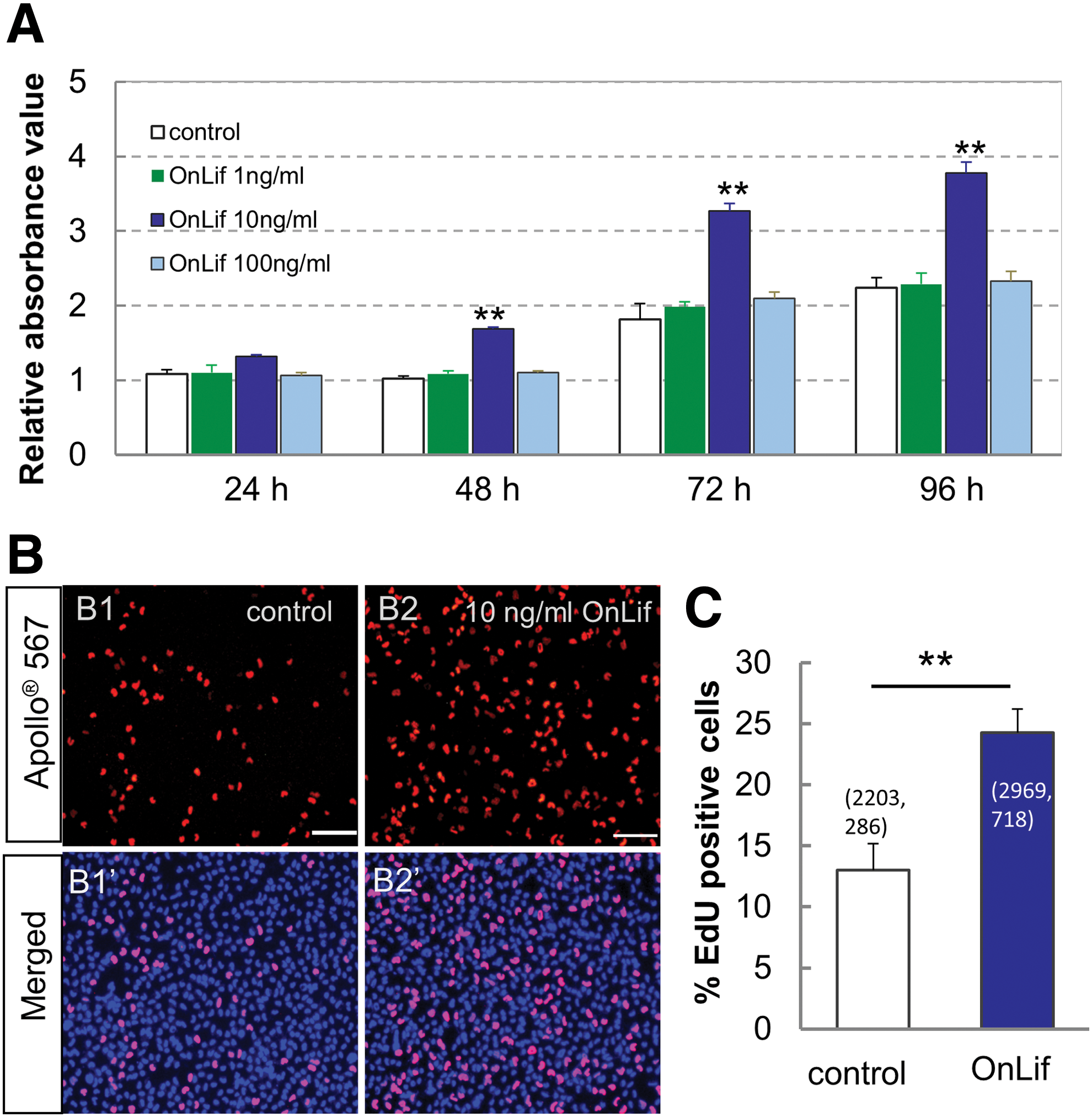
OnLif enhances proliferation of TES1.
OnLif enhances survival of TES1
To test whether OnLif could enhance cell survival, TES1 at low, middle, and high cell density (ie, 1.5 × 105, 3.0 × 105, and 6.0 × 105 cells per 24-well, respectively) were incubated in 5 N containing no or 10 ng/mL OnLif. In the control 5 N, after a 48-h incubation, almost all cells appeared apoptotic at low cell density, and a large quantity of cells appeared apoptotic at middle or high cell density (Fig. 5A). By contrast, in the presence of 10 ng/mL OnLif, very low quantity of cells appeared apoptotic at a different cell density, even at low cell density (Fig. 5A).
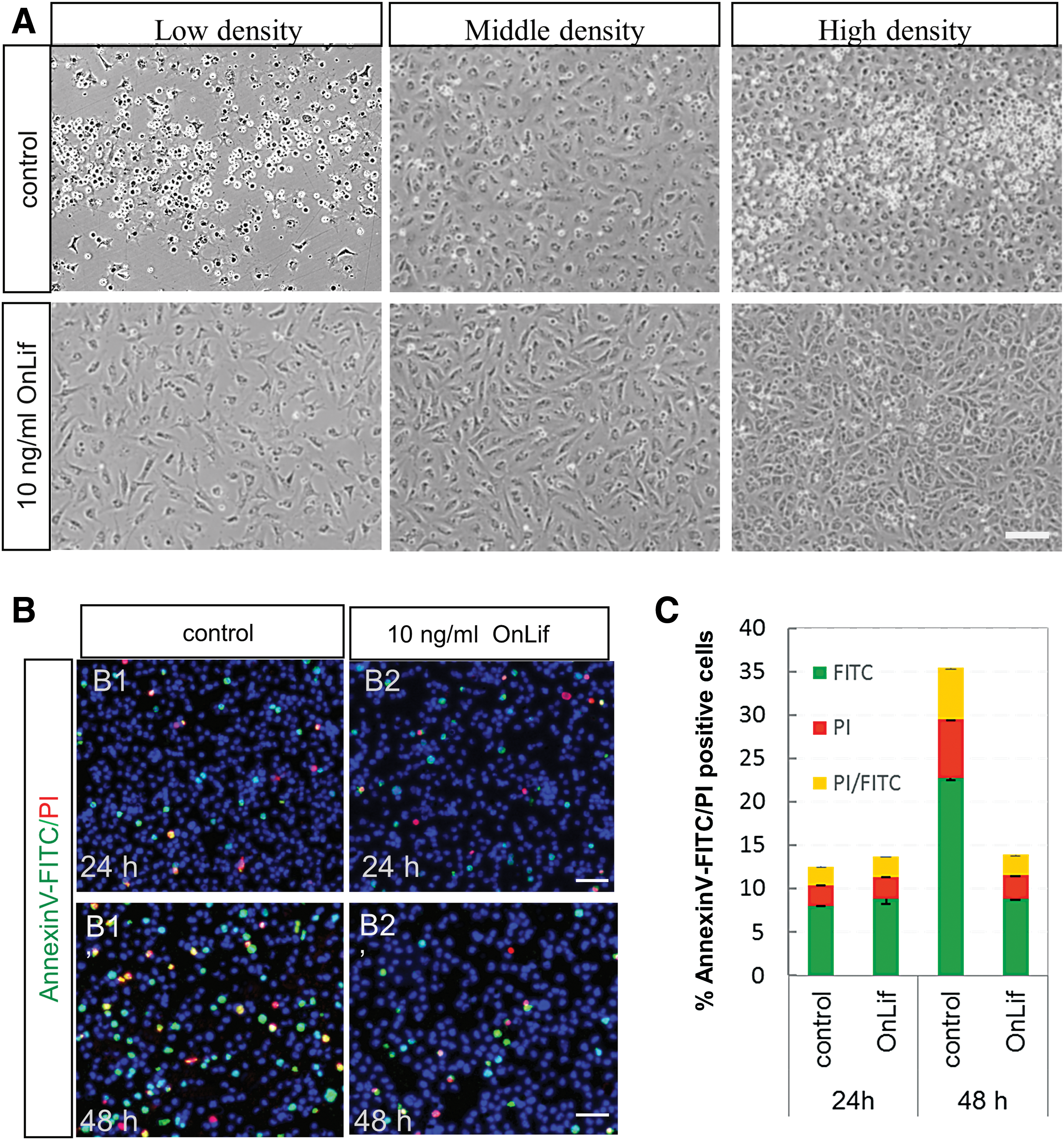
OnLif enhances survival of TES1 by suppressing apoptosis.
To further evaluate the suppressing apoptotic activity of OnLif, TES1 at middle cell density were measured at an indicated time by Annexin V-FITC/PI staining. At 24 h, no matter the presence of OnLif or not, low percentage (about 9%) of apoptotic cells were observed (P > 0.05; Fig. 5B, C). However, at 48 h, the proportion of apoptotic cells (FITC positive) significantly increased in 5 N (23%), but not in 5 N containing 10 ng/mL OnLif (9%) (P < 0 .001; Fig. 5B, C). In addition, among all the apoptotic cells (FITC-positive), about 27% were at late stage (FITC/PI double-positive cells) (Fig. 5B, C). These data suggest that OnLif could enhance survival of TES1 by suppressing apoptosis.
OnLif maintains pluripotency of TES1
TES1 are characteristic of high pluripotency gene expression, including pou5f3, myc, and klf4, low differentiation marker gene expression, including nf2000 and actn2, and a strong AP activity [31]. The mRNA expression levels of these pluripotency and differentiation markers in TES1 were detected using both RT-PCR and real-time PCR. In the presence of OnLif, TES1 showed significantly higher expression levels of pluripotency markers (pou5f3, myc, and klf4) and lower expression levels of differentiation markers (nf2000 and actn2) than that in the control (Fig. 6A, B). Meanwhile, the cells displayed a strong AP activity in the presence of 10 ng/mL OnLif, whereas less in the control (Fig. 6C1, C2).
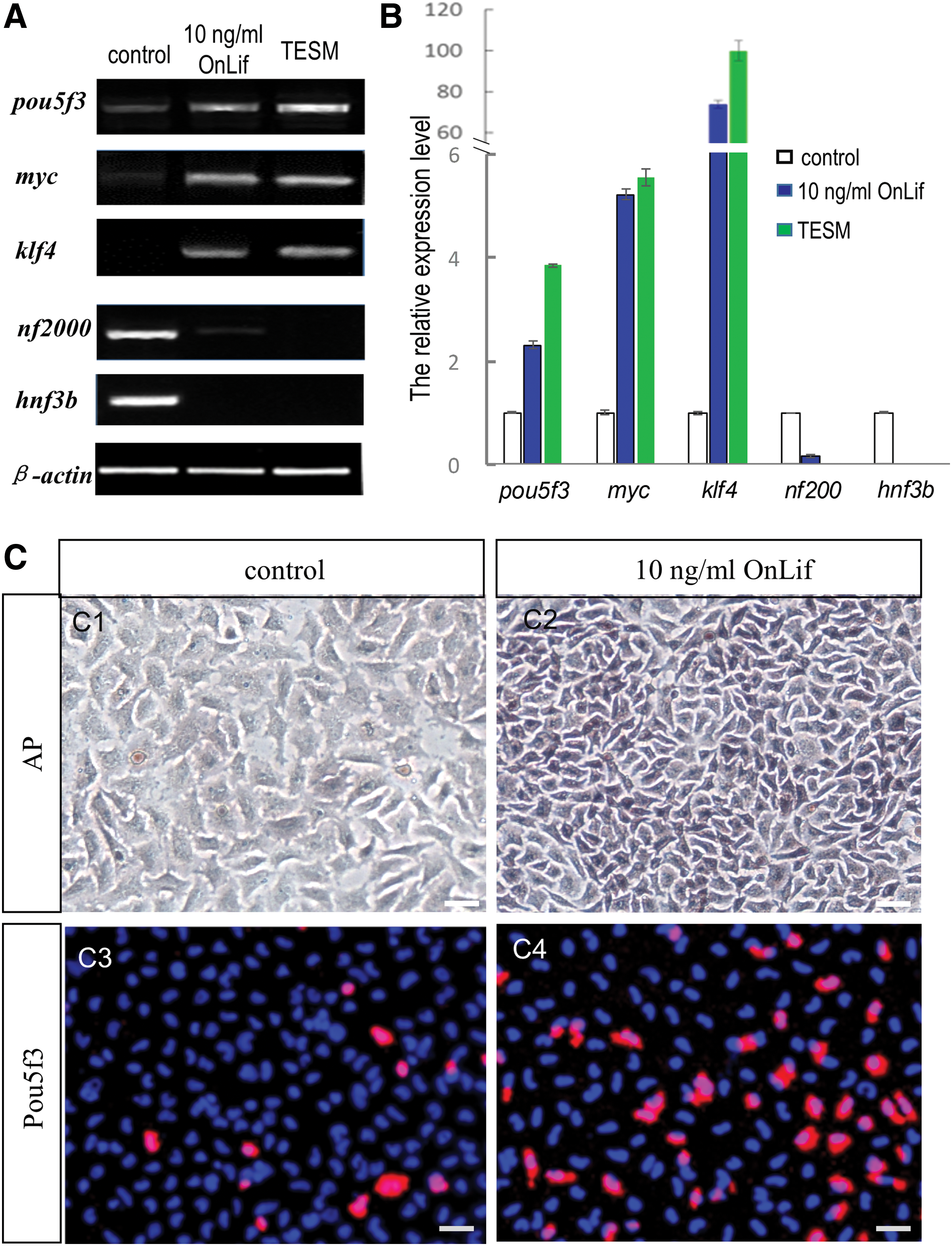
OnLif maintains pluripotency of TES1.
The expression of pluripotency marker Pou5f3 was further determined using immunocytochemistry. Consistent with the results by PCR, in the presence of OnLif, TES1 displayed significantly higher expression of Pou5f3 protein than that in the control (Fig. 6C3, C4). Taken together, OnLif could contribute to pluripotency maintenance of TES1.
OnLif activates Stat3
In mammals, JAK-STAT3 is the major pathway in Lif-mediated self-renewal of ES cells [7]. To determine whether OnLif could activate Stat3, the expression and tyrosine phosphorylation levels of Stat3 were measured by western blotting. Under different culture conditions, TES1 had similar expression levels of Stat3, whereas the tyrosine phosphorylation levels of Stat3 varied greatly (Fig. 7A, B). In the absence of OnLif, TES1 displayed undetectable tyrosine phosphorylation level of Stat3. In the presence of 10 ng/mL OnLif, TES1 displayed significantly high tyrosine phosphorylation level of Stat3, whereas it dramatically decreased to undetectable level after being supplemented with 50 or 100 μM S3I-201 (the inhibitor of Stat3 activity). These data suggest that OnLif might mediate Stat3 activating in TES1 (Fig. 7A, B).
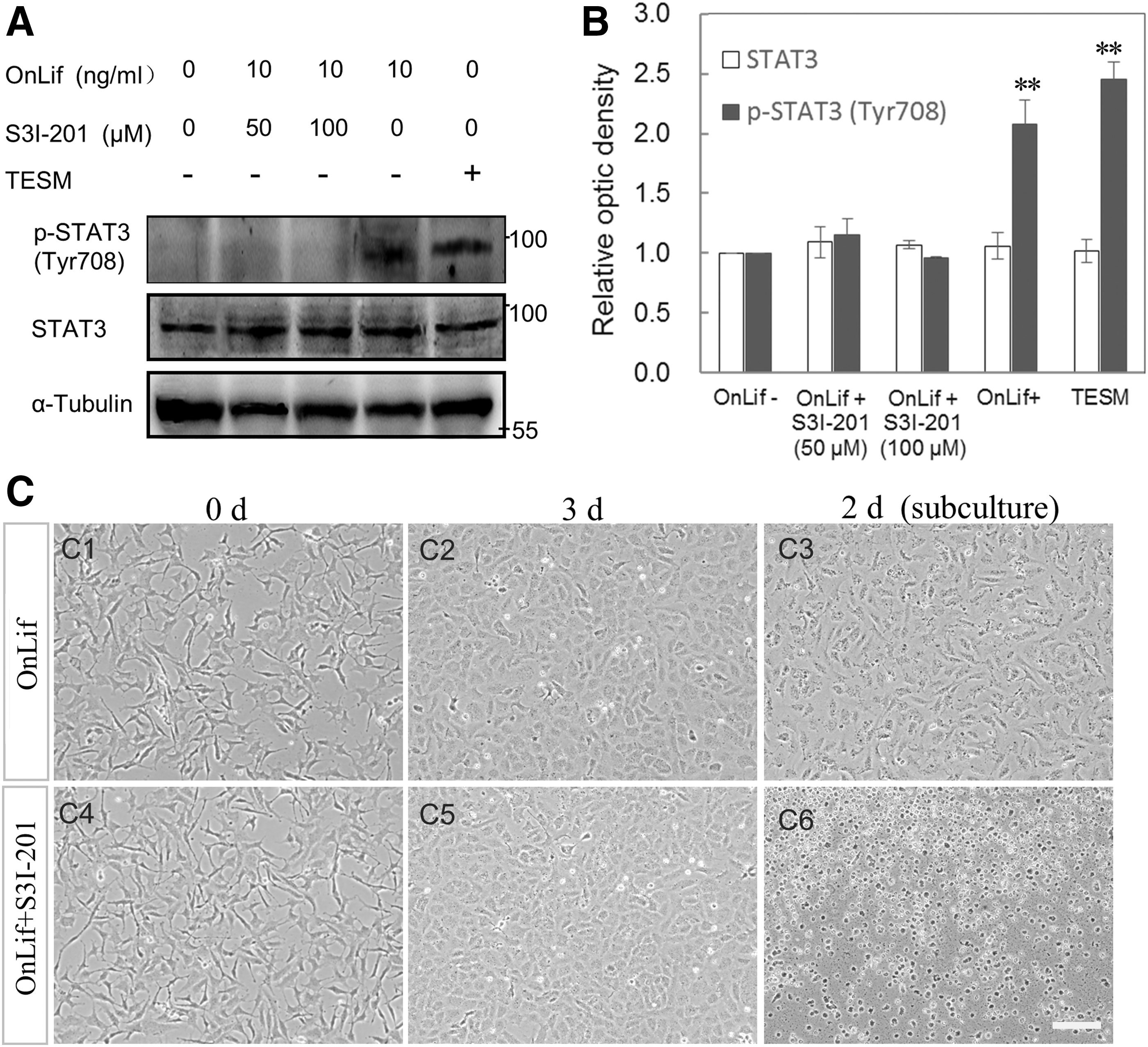
OnLif mediates self-renewal of TES1 cells by Stat3 signaling.
To further test whether OnLif-mediated Stat3 signaling could contribute to the proliferation and/or survival of TES1, the cells incubated in 5 N containing 10 ng/mL OnLif were treated with no (the control) or 50 μM S3I-201. During the incubation period of 1–3 days, TES1 appeared obvious proliferation and developed into a monolayer regardless of S3I-201 treatment or not (Fig. 7C1–C3). However, after a 2-day subculture, almost all cells treated by S3I-201 appeared apoptotic, whereas the control cells adhered and developed into a monolayer (Fig. 7C4–C6). These results suggest that OnLif might mediate self-renewal of TES1 primarily through the Stat3 signaling pathway.
OnLif enhances proliferation of ES cells derived from medaka
To test the species specificity of OnLif, the proliferation activity of ES cells derived from medaka (MES1) in the 5 N containing no or different concentrations of OnLif was measured by the CCK8 kit. Just like TES1, MES1 exhibited a significant proliferative activity in 5 N containing 10 ng/mL OnLif compared with that in 5 N containing no or 1 and 100 ng/mL OnLif (P < 0.01) (Fig. 8). This result suggests that OnLif has conserved biological activity in some other fish ES cells just like medaka.
OnLif enhances proliferation of ES cells from medaka (MES1). The cells were incubated in 5 N containing no or different concentrations of OnLif. These are the representative results from three independent experiments. **Indicate very significant differences (P < 0.01) from the control at the corresponding time. ES, embryonic stem.
Discussion
In this study, we reveal that the LIF orthologue from a teleost fish is essential for the self-renewal of fish ES cells by activation of Stat3. Four independent lines of evidence support this notion. First, OnLif is orthologous to mammalian LIF, which displays similar sequence features like the mammalian LIF, including mRNA sequence trait (instability motifs), the presence of signal peptide, potential N-glycosylation sites, cysteine residues, and 3-exon genomic organization (Fig. 1 and Supplementary Fig. S1) [3,27,38]. Phylogenetic tree and chromosome synteny analyses also support the ortholog to mammalian LIF. Second, the mRNA expression pattern of OnLif is similar to that of mammalian LIF at blastula stage onward, implying its potential role in pluripotency maintenance during embryogenesis. Third and more importantly, OnLif is critical for the proliferation, survival, and pluripotency maintenance of fish ES cells. Finally, OnLif can mediate the Stat3 Tyr phosphorylation, which contributes to the proliferation and/or survival of ES cells. Collectively, these data first suggest that Lif/Stat3 signaling plays an essential role in the self-renewal of fish ES cells.
ES cells are a powerful system for exploring the fundamental program of pluripotency maintenance. In recent years, deep insights have been made about the molecular mechanisms involved in regulation of pluripotency maintenance in mice and humans. Mouse canonical ES cells have the ability of germline transmission, which are considered to be totipotent and termed as ground state (or naïve) ES cells [25]. On the other hand, human conventional ES cells, whether derived from blastocysts or induced by somatic cell reprogramming, differ substantially from canonical mouse ES cells and are considered representing a later phase of epiblast development, and termed as primed state ES cells [25]. It has been elegantly demonstrated that the LIF signaling is essential in the pluripotency maintenance of mouse canonical ES cells, but not required in human conventional ES cells [7,25,39]. Nonetheless, recent studies reveal that in the presence of LIF stimulation, human conventional ES cells could be converted from the primed state into ground state, suggesting the conservation of LIF signaling in ground state pluripotency maintenance among mice and humans [4,7,12,13,22,25,26]. Furthermore, extensive studies suggest that LIF can maintain ES cells from other species such as pig, rabbit, and chicken in an undifferentiated state [38,40 –42]. Combined with our observations in fish ES cells, these data suggest that LIF plays an essential and conserved role in the pluripotent maintenance among vertebrates.
Accumulative studies demonstrate the central role of STAT3 activation in the LIF-mediated pluripotency maintenance of mouse ES cells [7,20,43 –46]. In the absence of STAT3, mouse ES cells are unable to self-renew [46]; conversely, STAT3 activation is sufficient to confer LIF-independent self-renewal of mouse ES cells [45]. In vertebrates other than mouse, such as human, pig, rabbit, and chicken, LIF can mediate pluripotency maintenance of ES cells by STAT3 signaling [22,26,38,40,42]. In this study, OnLif mediated Stat3 Tyr phosphorylation in TES1, which could be specifically inhibited by its activity inhibitor S3I-201 (Fig. 7). Moreover, S3I-201 treatment led to TES1 apoptosis at 2-day subculture, suggesting the critical role of Stat3 signaling in the self-renewal of fish ES cells. These data collectively suggest that LIF/STAT3 signaling plays an essential and conserved role in the pluripotency maintenance among vertebrates.
Functional analyses show that transcription factors, including Klf4, Tfcp2l, and Gbx2 are target genes regulated by LIF/STAT3, which can confer varying degrees of LIF-independent self-renewal [16,47,48]. In this study, in addition to the notable pluripotency transcription factors pou5f3 and myc, klf4 was dominantly upregulated by OnLif stimulation, suggesting the conservation of the core pluripotency regulatory network in fish ES cells. This makes fish ES cells an ideal model system to investigate the mechanisms underlying self-renewal among vertebrates.
Multiple amino acid sequence alignment of LIF from different species reveals low homolog of Lif across vertebrates (Supplementary Fig. S2). The sequence identity of OnLif is below 20% to mammalian homolog. A previous study reports that murine LIF has no effect on medaka fish (O. latipes) ES cells [30]. Considering the high sequence variety across species, we suspect that mammalian LIF might have no function in fish ES cells. Hence, in this study, to evaluate the biological activity of fish Lif in fish ES cells, the OnLif protein was obtained by prokaryotic expression for further study.
The insolubility and low yield is a challenging problem in the production of LIF. Hence, it takes about 90% cost of the culture medium in iPSC/ES cell research [10,30]. To increase the solubility and yield of LIF, different strategies have been attempted, such as codon optimized and synthetic cDNA template, low temperature expression, recombinant protein fused to different tags, including glutathione-S-transferase, thioredoxin, thioredoxin-histidine, and the b′ a′ domain of protein disulfide isomerase [10,31 –37]. In recent years, another solubility tag that has gained popularity is SUMO [38]. After expression, the SUMO moiety can be cleaved using SUMO-specific proteases such as ULP1 [38]. In this study, soluble OnLif protein was obtained fused to His6-SUMO tag and induced at low temperature of 16°C. To avoid the impact of the expression tag on the function of target protein, the His6-SUMO tag was removed by SUMO-specific enzyme ULP1 digestion. Our study indicates that the SUMO fusion protein expression system is simple and efficient for the soluble and native Lif protein production.
Consistent with the previous report in medaka [30], our results indicate that murine LIF has no proliferative effect on TES1, supporting the species specificity of LIF between mammals and fish. Nevertheless, OnLif has a significant proliferation-promoting activity in ES cells from medaka (Fig. 8), suggesting its potential application in some other fish ES derivation and culture. Taken together, we have firstly demonstrated that Lif has a critical role in fish ES cell self-renewal. Our study would promote the availability of ES cells from other fish species and extend the power of ES-based genetic manipulation to commercial animals.
Footnotes
Acknowledgments
This work was supported by grants XDJK2017B007, XDJK2017D102, XDJK2017D103, and XDJK2015A004 from Fundamental Research Funds for the Central Universities (Ministry of Education of China), grants 31502147, 31572597, and 31630082 from the National Natural Science Foundation of China, and grant cstc2016jcyjA0529 from the Natural Science Foundation Project of CQ CSTC.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.