
Abstract
In preclinical studies, the cynomolgus monkey (CM) model is frequently used to predict the pharmacokinetics of drugs in the human small intestine, because of its evolutionary closeness to humans. Intestinal organoids that mimic the intestinal tissue have attracted attention in regenerative medicine and drug development. In this study, we generated intestinal organoids from CM induced pluripotent stem (CMiPS) cells and analyzed their pharmacokinetic functions. CMiPS cells were induced into the hindgut; then, the cells were seeded on microfabricated culture vessel plates to form spheroids. The resulting floating spheroids were differentiated into intestinal organoids in a medium containing small-molecule compounds. The mRNA expression of intestinal markers and pharmacokinetic-related genes was markedly increased in the presence of small-molecule compounds. The organoids possessed a polarized epithelium and contained various cells constituting small intestinal tissues. The intestinal organoids formed functional tight junctions and expressed drug transporter proteins. In addition, in the organoids generated, cytochrome P450 3A8 (CYP3A8) activity was inhibited by the specific inhibitor ketoconazole and was induced by rifampicin. Therefore, in the present work, we successfully generated intestinal organoids, with pharmacokinetic functions, from CMiPS cells using small-molecule compounds.
Introduction
C
The small intestine plays an important role in nutrient absorption and functions as a barrier to prevent the entry of luminal pathogens into the bloodstream. Furthermore, the small intestine plays an important role in the pharmacokinetics of orally administrated drugs as several drug transporters and drug-metabolizing enzymes are expressed in this organ [8 –10]. Specifically, CYP3A8, solute carrier family 15 member 1/peptide transporter 1 (SLC15A1/PEPT1), which acts as a peptide transporter, and the ATP-binding cassette, subfamily B, member 1/multidrug resistance 1 (ABCB1/MDR1), which is an efflux transporter, are expressed at high levels in the small intestines of CM as well as those of humans [11 –13] and regulate the bioavailability of orally administrated drugs. In particular, the CYP3A subfamily, including CYP3A8, is involved in the metabolism of numerous drugs, and its expression is induced by ligands of nuclear receptors, such as rifampicin. Thus, the pharmacokinetic evaluation of drugs in the small intestine is critical in drug development studies. However, there is no appropriate in vitro model for the evaluation of human intestinal pharmacokinetics, including the metabolism and absorption of drugs. In drug development, to accurately predict human intestinal pharmacokinetics in in vitro assays, normal human small intestinal tissue or cells are necessary. Importantly, a large amount of tissues or cells is required for wide use in preclinical studies. However, it is difficult to provide these to such an extent at the present time. Moreover, considerable lot-to-lot variations would be unavoidable in normal human small intestinal tissues, even if we were able to obtain the quantities needed. Furthermore, because the direct evaluation of intestinal pharmacokinetics in humans is challenging, it is necessary to compare the in vivo and in vitro data in animal models [14]. Specifically, the prediction of human intestinal pharmacokinetics in vivo can be performed using in vitro–in vivo correlation data from the CM animal model. Species-related differences may be overcome by determining the difference between the pharmacokinetic profiles of drugs using the same system in both humans and CM. The intrinsic metabolic clearance of drugs in the small intestinal microsomes differs between CM and humans [15,16]; therefore, a system in which both intestinal metabolism and absorption can be evaluated is necessary. Intestinal organoids have been proposed to be useful for the prediction of such pharmacokinetic properties.
Intestinal organoids are three-dimensional (3D) tissue structures that mimic the intestinal tissues; they have attracted attention in the field of regenerative medicine and drug development. Sato et al. reported that intestinal cells cultured in Matrigel support intestinal epithelial growth and self-renewal, and demonstrated that these cells form villus–crypt structures with all differentiated cell types such as enterocytes, goblet cells, Paneth cells, and enteroendocrine cells [17]. Spence et al. reported the generation of 3D gut-like organoids from human induced pluripotent stem (iPS) cells [18] using this method. The organoids generated exhibited polarity, formed villi, and crypt-like structures, and comprised various types of intestinal cells. Mizutani et al. reported that organoids generated from mouse primary small intestinal cells were useful for the evaluation of the dynamics of ABCB1/MDR1-mediated drug transport [19]. However, the organoids generated in these studies were immature. Furthermore, studies on the application of intestinal organoids to the evaluation of drug pharmacokinetics are rare and, to our knowledge, the generation of CM induced pluripotent stem (CMiPS) cell-derived organoids has not been reported to date.
In this study, we report the successful generation of intestinal organoids from CMiPS cells and the analysis of their pharmacokinetic functions. The present system is expected to be of utility in the prediction of drug pharmacokinetics in the human small intestine.
Materials and Methods
Animal tissues
CM skin and intestinal tissues were from Ina Research Co. (Nagano, Japan), an accredited animal facility, complying with the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC) Policies (approval No.: 001107). This study was performed according to the guidelines of the Institutional Animal Care And Use Committee (IACUC) of Ina Research Co.
CM fibroblast culture
CM skin tissues were collected from 4-year-old male CMs weighing 2.85 kg. CM fibroblasts were collected from skin tissues and cultured in Dulbecco's modified Eagle's medium (DMEM; Wako) containing 10% fetal bovine serum (FBS; Nichirei), 2 mM L-glutamine (L-Gln; Nacalai Tesque), 1% nonessential amino acids (NEAA; Nacalai Tesque), 100 U/mL penicillin, and 100 μg/mL streptomycin (Nacalai Tesque).
Generation of CMiPS cells
Three classic episomal vectors, namely pCXLE-hOCT3/4-shp53-F, pCXLE-hSK, and pCXLE-hUL, which encode reprogramming factors, were obtained from Addgene. CM fibroblasts were cotransfected with a mixture of these plasmids by electroporation (Neon Transfection System Kit; Thermo Fisher Scientific). Transfected fibroblasts (6.0 × 105 cells) were seeded onto gelatin-coated 6-well plates and cultured in DMEM containing 10% FBS, 2 mM L-Gln, 1% NEAA, 100 U/mL penicillin, and 100 μg/mL streptomycin. Seven days after electroporation, cells were harvested by trypsinization and reseeded onto mitomycin C-treated mouse embryonic fibroblasts (MEF, 1.0 × 105 cells/100 mm dish). The medium was replaced with DMEM/Ham's F-12 (DMEM/F12; Wako) supplemented with 20% KnockOut Serum Replacement (KSR; Thermo Fisher Scientific), 2 mM L-Gln, 1% NEAA, 100 U/mL penicillin, 100 μg/mL streptomycin, 0.1 mM β-mercaptoethanol (Sigma-Aldrich), and 5 ng/mL FGF2 (PeproTech), and cells were cultured at 37°C in humidified air with 5% CO2. The medium was exchanged every other day. Then, 3–5 weeks after electroporation, three colonies (#1, 2, and 3) were picked and dissociated mechanically into small clumps by pipetting, seeded onto mitomycin C-treated MEFs, and cultured. The undifferentiated colonies were expanded by repeating passage. Thereafter, we used phosphate-buffered saline (PBS; TaKaRa Bio, Inc.) containing 0.25% trypsin (Thermo Fisher Scientific), 0.1 mg/mL collagenase IV (Gibco), 1 mM CaCl2, and 20% KSR as a dissociation solution. To confirm differentiation into the three germ layers, we used a method reported by Okabayashi and Asashima [20], with some modifications. Briefly, CMiPS cells were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Thermo Fisher Scientific) containing 0.5% FBS, 2 mM GlutaMAX (Thermo Fisher Scientific), and Activin A (PeproTech). For differentiation into ectoderm, mesoderm, or endoderm, Activin A was added at 0.5, 10, or 100 ng/mL, respectively. After 3 days of differentiation, we replaced the medium containing 0.5% FBS with that containing 2% KSR, and the cells were subsequently cultured for 2 days.
Differentiation into intestinal organoids
We used the #2 CMiPS cell line for induction into intestinal organoids. CMiPS cells were passaged onto Growth Factor-Reduced Matrigel (Matrigel, Corning)-coated plates and cultured with StemSure® hPSC medium (Wako) supplemented with 35 ng/mL FGF2 before differentiation. After reaching 80% confluence, cells were cultured with RPMI 1640 medium containing FBS, 2 mM L-Gln, 100 U/mL penicillin, 100 μg/mL streptomycin, and 100 ng/mL Activin A for 3 days. FBS concentration was gradually increased from 0% to 0.2% and 2%. Then, the medium was replaced with RPMI 1640 medium containing 2 mM GlutaMAX and 2% FBS with 250 ng/mL FGF2 and 6 μM CHIR99021 (Focus Biomolecules). After 4 days of culture, cells were trypsinized and filtered through a 40 μm nylon mesh cell strainer (BD Biosciences). The cells (3.0 × 106 cells) were then plated on an EZSPHERE plate (AGC Techno Glass) to form spheroids. Y-27632 (10 μM, Focus Biomolecules) was added for 72 h after replating. The spheroids were cultured on an EZSPHERE plate for 3 days, and then transferred to ultralow binding plates (Corning). Next, to generate intestinal organoids, spheroids were cultured by floating with Advanced DMEM/F12 (Thermo Fisher Scientific) containing 3% Matrigel, 200 ng/mL R-Spondin1 (Noves), 100 ng/mL Noggin (GenScript), and 100 ng/mL epidermal growth factor (EGF; GenScript) with 2 mM L-Gln, N2 supplement (Thermo Fisher Scientific), B27 serum-free supplement (Thermo Fisher Scientific), 100 U/mL penicillin, 100 μg/mL streptomycin, and 15 mM HEPES (Millipore) for 27 days; 0.5 μM A-83-01 (AdooQ Bioscience), 20 μM PD98059 (AdooQ Bioscience), 5 μM 5-aza-2′-deoxycytidine (Chem-Impex International), and 5 μM N-[(3,5-difluorophenyl) acetyl]-
RNA extraction and reverse transcription reaction
Small and large intestinal tissues were collected from three male CMs (2.9, 4.4, and 4.9 years of age). Total RNA was extracted from CMiPS cell-derived intestinal organoids, as well as from CM small and large intestinal mucosa, using the Agencourt RNAdvance Tissue Total RNA Purification Kit (Beckman Coulter). First-strand cDNA was prepared from the total RNA. The reverse transcription reaction was performed using the ReverTra Ace qPCR RT Master Mix (TOYOBO) according to the manufacturer's instructions.
Real-time polymerase chain reaction analysis
Relative mRNA expression levels were determined using the Eco Real-Time PCR System with Eco Real-Time PCR System software v5.0 (Illumina). Polymerase chain reaction (PCR) was performed with the primer pairs listed in Table 1 using the KAPA SYBR Fast qPCR Kit (NIPPON Genetics) or TaqMan probes (CYP2C43/2C9; Assay ID: Mf04363679_m1 and CYP2C75/2C19; Assay ID: Mf04363854_m1) (Applied Biosystems). The mRNA expression of CYP1A1 and CYP2D17/2D6 was measured by real-time PCR as described previously [11,16]. mRNA expression levels were normalized to the expression of the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Alkaline phosphatase staining
Alkaline phosphatase staining was performed using the Leukocyte Alkaline Phosphatase Kit (Sigma-Aldrich), according to the manufacturer's instructions. The cells were washed with PBS and fixed to slides by incubating in citrate-buffered acetone for 30 s. Then, cells were gently rinsed in MilliQ water for 45 s and incubated with an alkaline-dye mixture for 60 min. After incubation, the slides were rinsed thoroughly in MilliQ water for 2 min.
Hematoxylin–eosin and Alcian Blue staining
Intestinal organoids and CM small intestinal tissues were fixed with 4% paraformaldehyde. The samples were embedded in optimal cutting temperature (OCT) compound (Sakura), frozen, and cut into 10-μm sections. Hematoxylin–eosin (HE) staining was performed according to conventional methods. Alcian Blue staining was performed at pH 2.5, and Nuclear Fast Red was used for nuclear staining.
Immunofluorescence staining
Intestinal organoids and CM small intestinal tissues were fixed with 4% paraformaldehyde. The samples were embedded in OCT compound, frozen, and cut into 10-μm sections. The sections were subject to antigen retrieval, blocked in PBS containing 5% FBS for 30 min, and incubated with primary antibodies overnight at 4°C. Then, sections were washed and incubated with secondary antibodies for 1 h at 20°C–25°C. The antibodies used and dilution ratios are listed in Table 2. After washing, the sections were incubated with 1 μg/mL 4′,6-diamidino-2-phenylindole (DAPI; Wako) and washed again. The sections were mounted using SlowFade Diamond Antifade Mountant (Thermo Fisher Scientific) and viewed using a Zeiss LSM510 Meta Laser Scanning Confocal Microscope (Carl Zeiss Microscopy).
Transmission electron microscopy
The intestinal organoids were fixed with 2.5% glutaraldehyde and incubated overnight at 4°C. Next, postfixation was carried out with 1% osmium tetroxide for 2 h at 4°C, followed by dehydration with ethanol. After substitution with propylene oxide, the organoids were embedded in resin. The embedded samples were cut into 0.1-μm sections, stained with uranyl acetate, and viewed using a Hitachi H7600 transmission electron microscope (JEOL).
Permeability study of fluorescein isothiocyanate–dextran 4000
The organoids were washed with Hank's Balanced Salt Solution (HBSS, pH 7.4) and incubated with HBSS containing 1 mg/mL fluorescein isothiocyanate–dextran 4000 (FD-4; Sigma-Aldrich) for 1 h at 37°C. After incubation, they were washed with ice-cold HBSS and viewed using an ECLIPSE Ti-S fluorescence microscope (Nikon) [19,21].
Determination of CYP3A8 activity
The organoids were incubated with Advanced DMEM/F12 containing 2 mM L-Gln, N2 supplement, B27 serum-free supplement, 100 U/mL penicillin, 100 μg/mL streptomycin, and 15 mM HEPES containing 5 μM midazolam for 24 h at 37°C. After incubation, 40 μL of the reaction medium was collected, and the reactions were stopped by the addition of 120 μL of ice-cold acetonitrile containing 3.6 μM chlorpropamide as an internal standard; the mixture was stored at −80°C until analysis. The organoids were solubilized with 1 M NaOH, and the lysates were collected. The levels of 1′-hydroxylated metabolites were measured using ultraperformance liquid chromatography–tandem mass spectrometry (UPLC–MS/MS). For UPLC–MS/MS analyses, samples were thawed and centrifuged at 20,600 g for 5 min at room temperature. To measure metabolites, 10 μL aliquots of supernatant were injected into the UPLC-MS/MS apparatus. UPLC analysis was performed using a Water ACQUITY UPLC system (Waters) and a XBridge BEH C18 column (2.1 × 50 mm, 3.5 μm, Waters); the mobile phase consisted of 10 mM ammonium formate (A) and acetonitrile (B) and the temperature of the column and sample was 40°C and 4°C, respectively. The gradient with 5% B started at the flow rate of 0.25 mL/min; 0.5 min later, the gradient increased linearly for 2.0 min until 95% B. Subsequently, the flow rate rose to 0.55 mL/min by 2.1 min, and the gradient was maintained at 95% B until 3.5 min. Then, the gradient linearly decreased to 5% B by 3.6 min, and the flow rate decreased to 0.25 mL/min by 3.7 min, which was maintained until 5.4 min. Multiple reaction monitoring was used for detection of 1′-hydroxymidazolam derivative (m/z 342.03 → 168.40). Chromatographic data were analyzed using the Mass Lynx 4.1 software (Waters). To correct for CYP3A8 activity, the total protein content was measured using the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific), according to the manufacturer's instructions.
Statistical analysis
For mRNA expression analysis of CM small and large intestinal tissues, such as those in Fig. 3, “n” represents the number of animals that were analyzed. For experiments involving CMiPS cell-derived intestinal organoids, “n” represents the number of biological replicates (organoids from three to four wells were collected and pooled for each replicate). Each experiment was repeated on at least two separate occasions (independent experiments). Quantification of data is represented as the mean ± standard deviation (SD). Statistical comparisons between groups were performed with a two-tailed Student's t-test (Fig. 7A), one-way analysis of variance with Bonferroni method (Fig. 3), or a two-tailed Dunnett's test (Fig. 7B) using the PASW statistics 18 software. The α level for all tests was 0.05. P values were as follows: *P < 0.05, **P < 0.01.
Results
Generation of CMiPS cell lines
CM fibroblasts were cotransfected with episomal vectors encoding human OCT3/4-shp-53, SOX2, KLF4, LIN28, and l-MYC (Fig. 1A) [22]. Around day 28 after transfection, three flat and tightly packed CM embryonic stem (ES) cell-like colonies were observed (Fig. 1B). The cells exhibited a morphology similar to that of CMES cells [23], with large nuclei and scanty cytoplasm. The three colonies (#1, 2, and 3) stained positive for alkaline phosphatase activity (Fig. 1C). The mRNA expression levels of OCT3/4 and SOX2 were higher in CMiPS cells than CM fibroblasts (Supplementary Fig. S1A; Supplementary Data are available online at
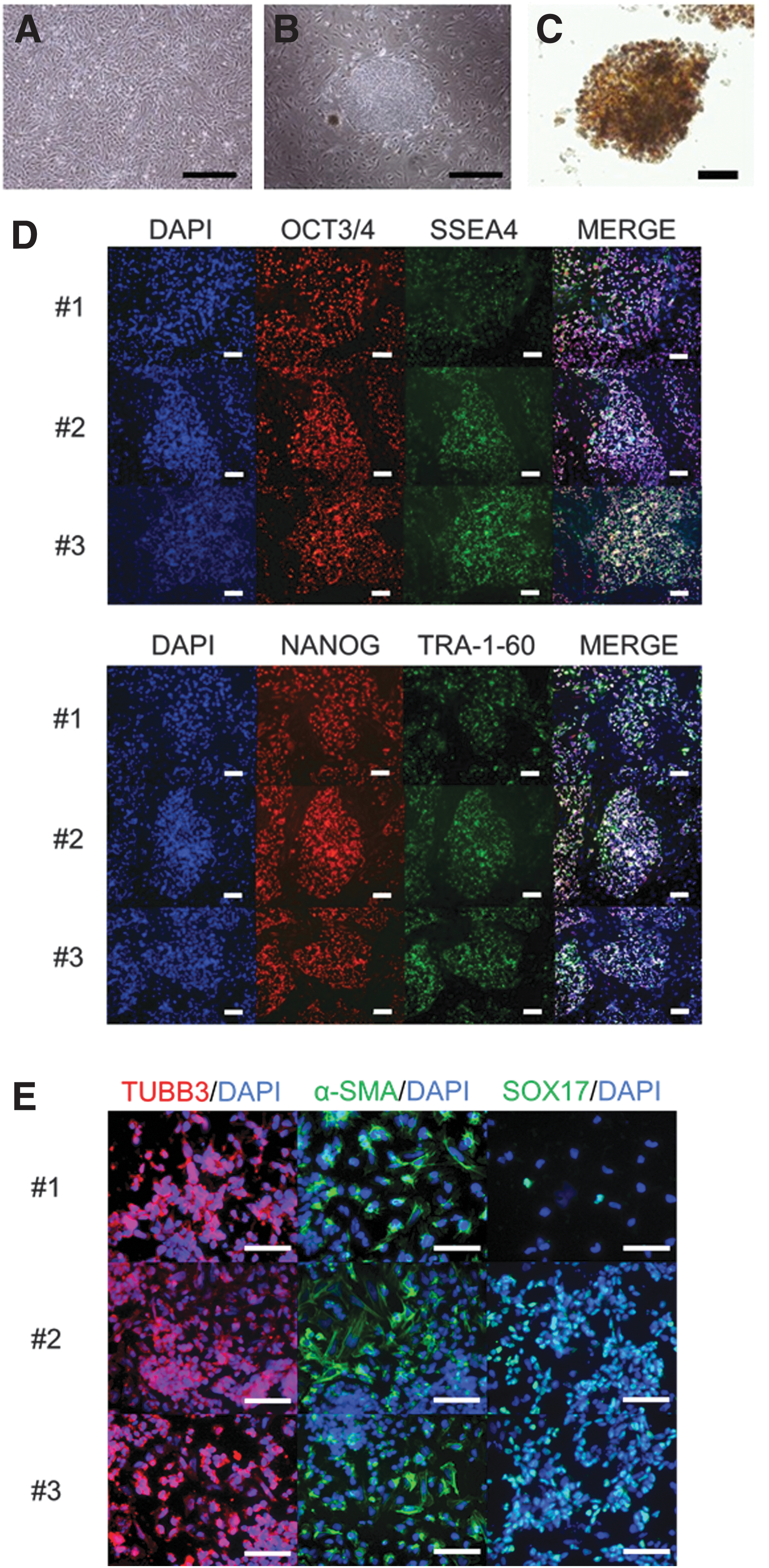
Characterization of the CMiPS cell lines. We generated iPS cell lines from CM fibroblasts by electroporating pCXLE-hOCT3/4-shp53-F, pCXLE-hSK, and pCXLE-hUL.
Generation of uniform spheroids
To generate intestinal organoids of uniform size easily and efficiently, we used a unique type of microfabricated culture vessel, namely the EZSPHERE plate for spheroid formation [24]. At day 7 of differentiation, the cells were seeded into the standard type of EZSPHERE plate (#900, microwell size: 500 μm in diameter and 100 μm in depth), at 500 cells per microwell, and cultured for 3 days (Fig. 2A). The cells spontaneously dropped into each microwell and formed homogeneous spheroids (Fig. 2B). Next, we generated intestinal organoids from uniformly sized spheroids by suspension culture (Fig. 2C) according to the protocol shown in Fig. 2D.
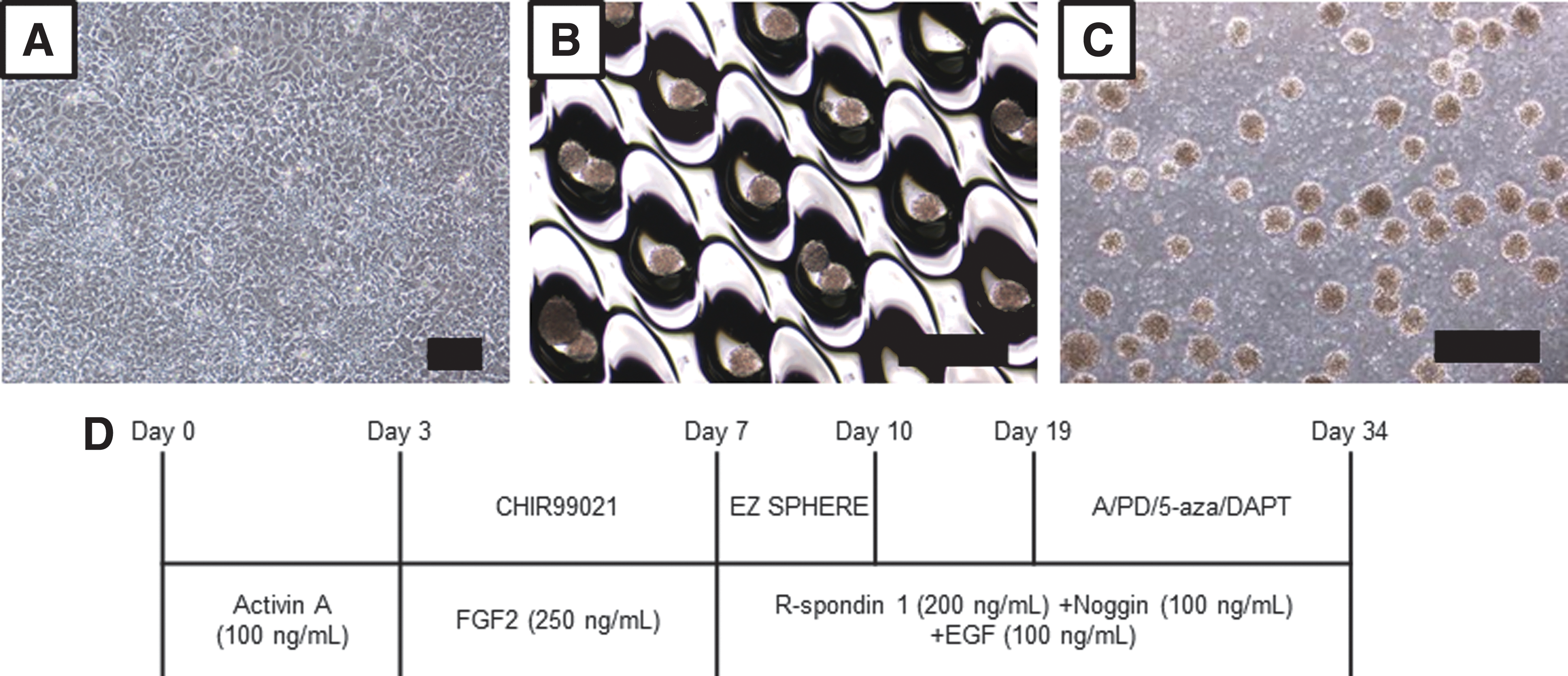
Morphology of CMiPS cell-derived spheroids.
mRNA expression of pharmacokinetic-related genes and intestinal markers in the CMiPS cell-derived intestinal organoids at day 34
Drug metabolism and absorption are important functions of the small intestine. Thus, we investigated the expression of genes involved in pharmacokinetics in the organoids (Fig. 3). The mRNA expression levels of CYP3A8 and pregnane X receptor (PXR), a nuclear receptor involved in CYP3A8 induction [25], were significantly increased following treatment with A-83-01, PD98059, 5-aza-2′-deoxycytidine, and DAPT (A/PD/5-aza/DAPT) compared with those in the other groups. Similarly, the mRNA expression of SLC15A1/PEPT1, solute carrier organic anion transporter family member 2B1/organic anion-transporting polypeptide 2B1 (SLCO2B1/OATP2B1), ABCB1/MDR1, ABCG2/breast cancer resistance protein (BCRP), and ABCC2/multidrug resistance-associated protein 2 (MRP2), which are uptake or efflux drug transporters expressed in the small intestine [11,13], was significantly increased by A/PD/5-aza/DAPT treatment compared with that in all other groups. On the contrary, the levels of CYP1A1 and CYP2C43/2C9 were similar to those in the small intestine, whereas the levels of CYP2C75/2C19 and CYP2D17/2D6 were lower than those in the small intestine (Supplementary Fig. S2). Notably, the expression levels of villin and sucrase–isomaltase, which are enterocyte markers [26,27], markedly increased following treatment with the small-molecule compounds (compare treated and nontreatment group) and the levels of these markers were the highest in the A/PD/5-aza/DAPT treatment group. The expression of the goblet cell marker MUC2 [28], the enteroendocrine marker chromogranin A [29], the Paneth cell marker lysozyme [30], and CDX2 also increased following treatment with the small-molecule compounds. In contrast, mRNA expression levels of LGR5 and olfactomedin 4 (OLFM4), which are intestinal stem cell markers [31,32], decreased. Interestingly, following treatment with the indicated small-molecule compounds, the expression pattern of the genes analyzed resembled the pattern observed in the small intestine more closely than that in the large intestine. Furthermore, we were able to confirm the same gene expression pattern in the #1 CMiPS cell line (Supplementary Fig. S3) upon the addition of A/PD/5-aza/DAPT. Accordingly, we used A-83-01, PD98059, 5-aza-2′-deoxycytidine, and DAPT to induce intestinal differentiation in the subsequent assays.
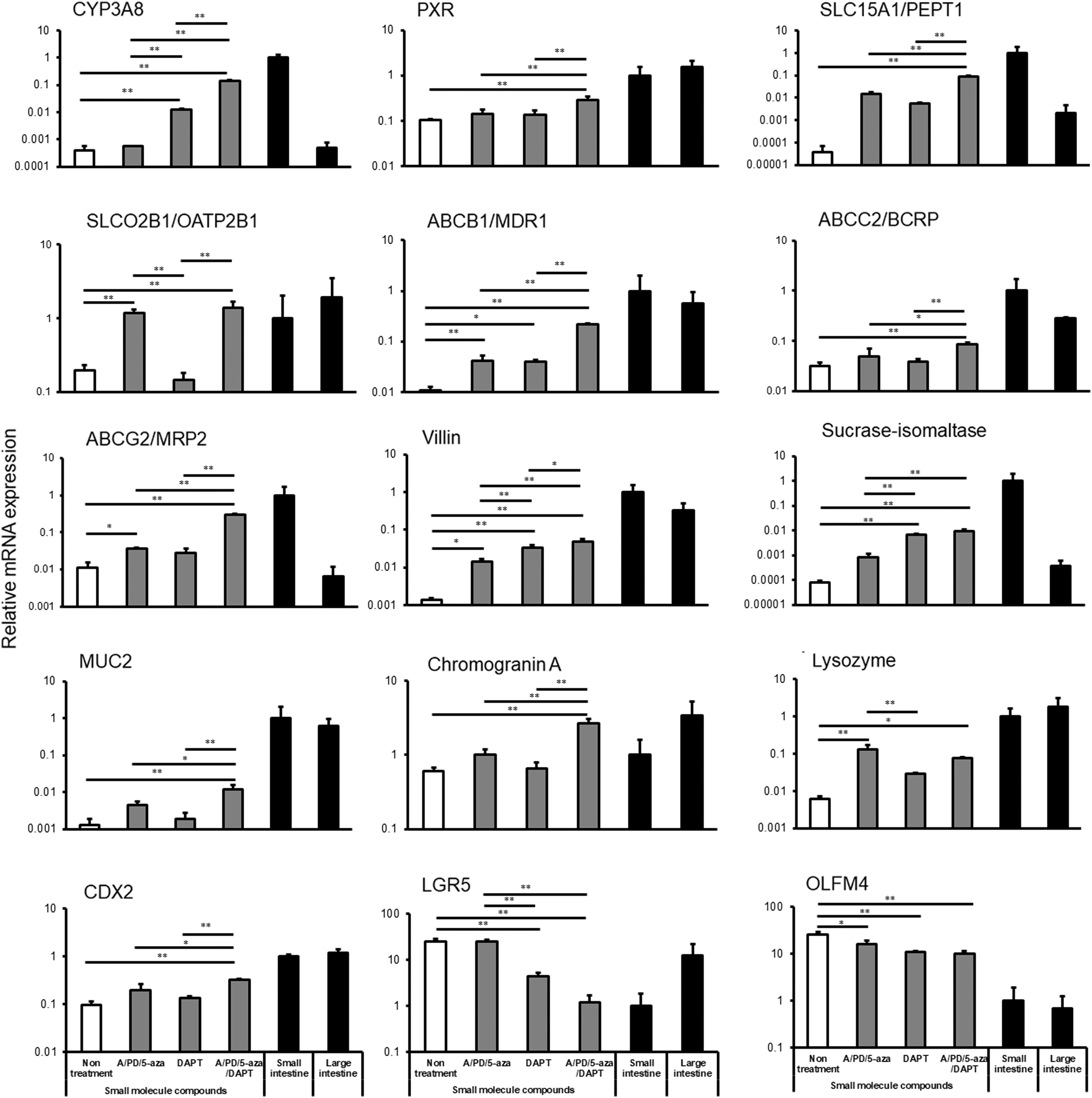
mRNA expression of pharmacokinetic-related genes and intestinal markers in the CMiPS cell-derived intestinal organoids at day 34. Around 0.25% DMSO (vehicle–untreated samples), A-83-01/PD98059/5-aza-2′-deoxycytidine (A/PD/5-aza), DAPT, or A-83-01/PD98059/5-aza-2′-deoxycytidine/DAPT (A/PD/5-aza/DAPT) were added on days 19–34. After 34 days of differentiation, total RNA was extracted, and mRNA expression was analyzed by real-time polymerase chain reaction. Data are represented as the mean ± SD (n = 3). mRNA expression levels were normalized to the expression of GAPDH. Gene expression levels are represented as relative to the levels in the small intestine, set to 1. Statistical analysis was performed using Bonferroni method, *P < 0.05, **P < 0.01. DAPT, N-[(3,5-difluorophenyl) acetyl]-
Morphology of the CMiPS cell-derived intestinal organoids at day 34
The CMiPS cell-derived intestinal organoids appeared as bubble-like structures (Fig. 4A). Microvilli and tight junctions, which are typical features of small intestinal tissues, were observed by transmission electron microscopy, and the interior of the organoids was the apical side (Fig. 4B). HE staining revealed that the organoids consisted of both well-polarized epithelium and epithelium with weaker organization compared with CM small intestinal tissues (Fig. 4C, D). Similar to the CM small intestinal tissues, the organoids were also observed to produce mucous (Fig. 4E, F).

Morphology of CMiPS cell-derived intestinal organoids at day 34.
Immunofluorescence staining of intestinal cell markers in the CMiPS cell-derived intestinal organoids at day 34
Immunofluorescence staining revealed that the CMiPS cell-derived intestinal organoids expressed villin, OLFM4, CDX2, MUC2, chromogranin A, and lysozyme, similar to the CM small intestinal tissues (Fig. 5A–H). In addition to these markers,E-cadherin, an intercellular adhesion molecule [33], was also expressed. Furthermore, the expression of the fibroblast marker vimentin and α-SMA was observed (Fig. 5I, J). Some cells were positive for both vimentin and α-SMA expression, a characteristic of intestinal subepithelial myofibroblasts [34].
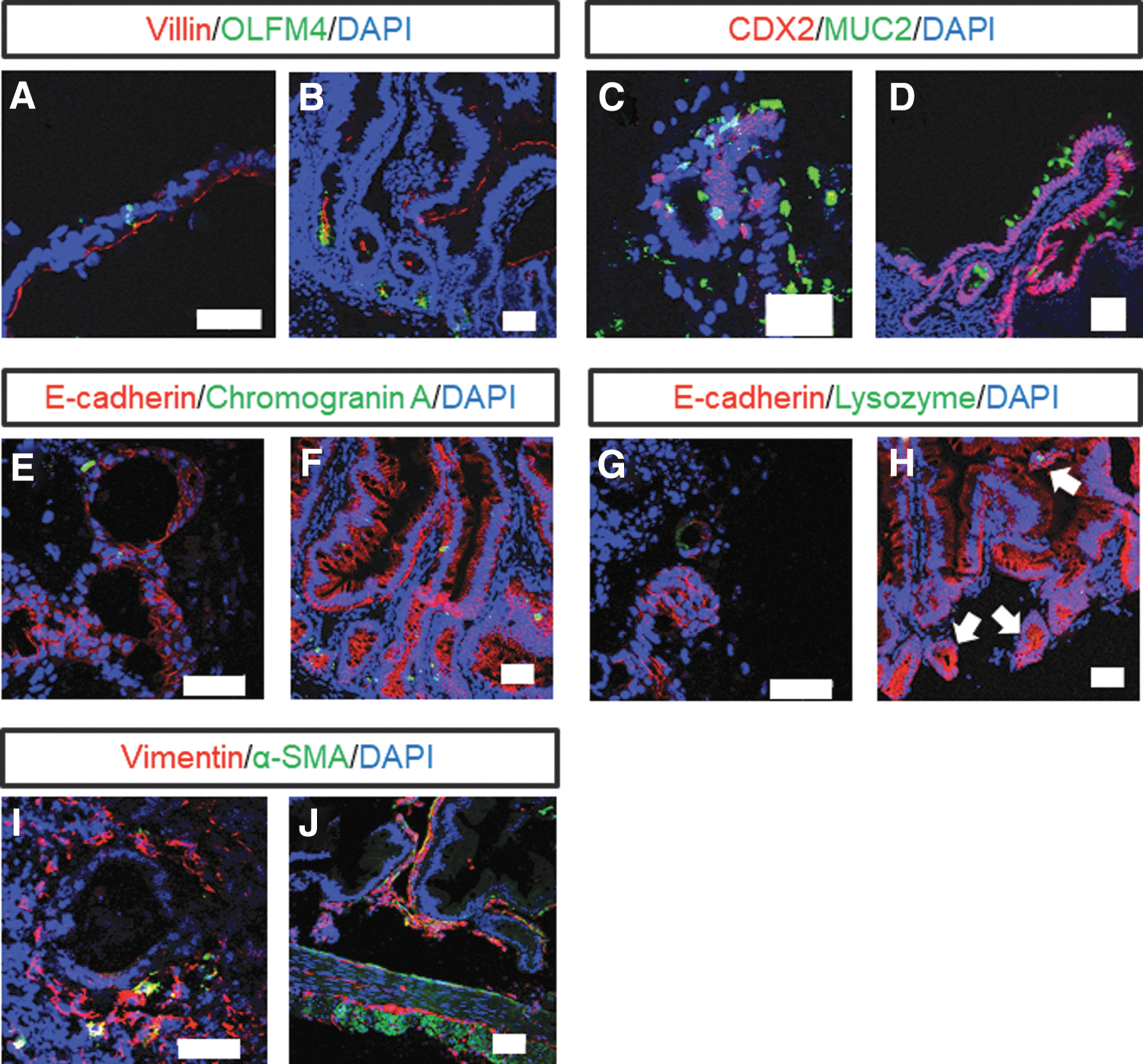
Immunofluorescence staining of intestinal cell markers in CMiPS cell-derived intestinal organoids at day 34. Immunofluorescence staining for villin, OLFM4, CDX2, MUC2, E-cadherin, chromogranin A, lysozyme (white arrows show lysozyme-positive cells), vimentin, and α-SMA in CMiPS cell-derived intestinal organoids
Analysis of the barrier function of the CMiPS cell-derived intestinal organoids at day 34
To investigate whether CMiPS cell-derived organoids formed functional tight junctions, we performed immunofluorescence staining of occludin, which is a tight junction marker [35], and carried out a permeability study of FD-4. The expression of occludin was observed in the lumen of the organoids (Fig. 6A). When CMiPS cell-derived intestinal organoids were incubated with FD-4, which is a nonabsorbable marker, its accumulation in the organoids was not observed (Fig. 6B). SLC15A1/PEPT1 and ABCB1/MDR1 were expressed in the lumen of the CMiPS cell-derived intestinal organoids (Fig. 6C, E), whereas in the CM small intestine, these transporters were expressed on the apical side (Fig. 6D, F).

Analysis of barrier function and drug transporter expression in the CMiPS cell-derived intestinal organoids at day 34.
Activity and induction of CYP3A8 in the CMiPS cell-derived intestinal organoids at day 34
Finally, we investigated whether the activity of CYP3A8 is induced in CMiPS cell-derived intestinal organoids. The metabolic activity of CYP3A8 was evaluated using midazolam and ketoconazole, which constitute a specific substrate and inhibitor of CYP3A8, respectively. The metabolic activity of midazolam was detected in the organoids; this was significantly inhibited by up to ∼5% by ketoconazole (Fig. 7A). In addition, we investigated the induction of CYP3A8 by rifampicin, an inducer of CYP3A8 [25], and 1α,25-dihydroxyvitamin D3, which is reported to induce human intestinal CYP3A4 through the vitamin D receptor [36]. CYP3A8 activity was induced 7.5- and 3.5-fold by rifampicin and 1α,25-dihydroxyvitamin D3, respectively (Fig. 7B).
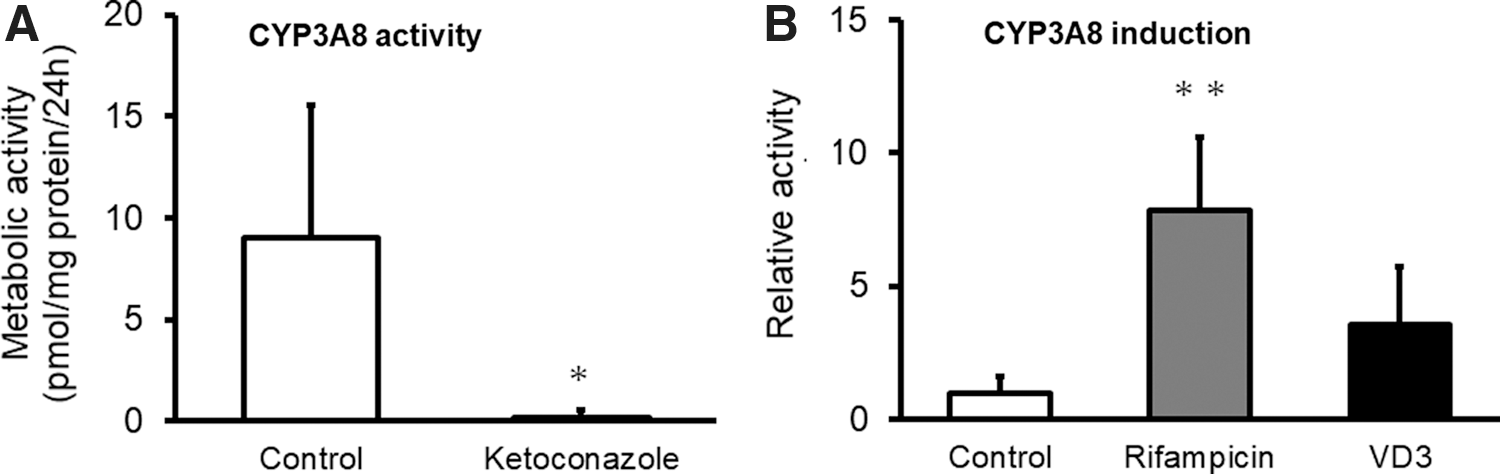
Activity and induction of CYP3A8 in the CMiPS cell-derived intestinal organoids at day 34.
Discussion
In this study, we successfully generated CMiPS cell-derived intestinal organoids with pharmacokinetic functions. It has previously been reported that human intestinal organoids generated by conventional methods are immature [37,38]. Accordingly, we attempted the generation of functional CMiPS cell-derived intestinal organoids by improving the organoid generation method.
In the conventional method, the formed spheroids are embedded into Matrigel and cultured [18,39,40]. However, this method is complex and time consuming; furthermore, the generation of spheroids of uniform size using this method is difficult to achieve. Therefore, to overcome this challenge, we cultured the spheroids in suspension, after their generation in EZSPHERE plates. Our results showed that the method we used allows the generation of a large number of uniform spheroids (Fig. 2B, C). It has been reported that the size of the embryoid body is a crucial parameter controlling the differentiation of ES cells [41]. Thus, if the spheroid size is not uniform, variation of differentiation efficiency and experimental data will likely occur. Moreover, a large number of uniform organoids is required in drug development studies. Therefore, our method is particularly useful because, unlike conventional methods, it allows spheroids of uniform size to be efficiently formed and cultured in suspension.
We next examined whether the CMiPS cell-derived intestinal organoids could mature. We observed that the cotreatment with A-83-01, PD98059, 5-aza-2′-deoxycytidine, and DAPT during the intestinal differentiation stage resulted in the increased expression of various intestinal cell markers, such as villin, sucrase–isomaltase, MUC2, chromogranin A, lysozyme, and CDX2 (Fig. 3). Conversely, the mRNA expression of intestinal stem cell markers, such as LGR5 and OLFM4, was significantly decreased compared with that in the untreated group (Fig. 3). It has previously been suggested that small-molecule compounds promote differentiation into intestinal epithelial cells and we have found that the combination of A-83-01, PD98059, and 5-aza-2′-deoxycytidine promotes the differentiation of human iPS cells into enterocytes [42]. Notably, the mRNA levels of various pharmacokinetic-related genes and intestinal markers in the generated organoids were lower than those in the small intestine. Therefore, it is still necessary to perfect the organoids differentiation protocol, and reach gene expression levels that are closer to those in the small intestine. Ogaki et al. reported that 6-bromoindirubin-3′-oxime and DAPT promote the intestinal differentiation of human ES cells [43]. Our results in CMiPS cells were consistent with these results, indicating that small-molecule compounds can effectively induce the generation of CMiPS cell-derived intestinal organoids in both CM and human iPS cells.
The CMiPS cell-derived organoids exhibited mRNA and protein expression of many intestinal cell markers (Figs. 3 and 5A, C, E, G), suggesting that the organoids comprise absorptive epithelial cells, enteroendocrine cells, goblet cells, Paneth cells, fibroblasts, and smooth muscle cells, among others. We additionally observed vimentin/α-SMA-positive cells indicative of intestinal subepithelial myofibroblasts in the organoids (Fig. 5I, J). Intestinal subepithelial myofibroblasts are particularly important for epithelial differentiation and gut morphogenesis [44,45]; therefore, their presence indicates that the epithelial–mesenchymal crosstalk may be important in the differentiation and growth of CMiPS cell-derived intestinal organoids.
Tight junctions and microvilli, which are morphological features of the small intestine, were observed in the CMiPS cell-derived intestinal organoids (Fig. 4B). E-cadherin and occludin, which are intercellular adhesion molecules, were also found to be expressed (Fig. 6A). Furthermore, the accumulation of nonabsorbable marker FD-4 into the organoids was not detected (Fig. 6B), suggesting that the tight junctions were functional. SLC15A1/PEPT1 is expressed on the apical side of the small intestine and involved in peptide transport from the lumen [46]. ABCB1/MDR1 is located on the apical membrane of intestinal epithelial cells, where it exports drugs and xenobiotics [46]. The CMiPS cell-derived intestinal organoids expressed these transporters in their luminal part (Fig. 6C, E), indicating that the internal part of the organoids corresponds to the apical side of the small intestine.
The mRNA expression analysis of pharmacokinetic-related genes indicated that CYP3A8, PXR, and uptake and efflux transporters were expressed in the CMiPS cell-derived intestinal organoids (Fig. 3). The levels of most of the genes investigated were more similar to those in the small intestine than in the large intestine. Intestinal metabolism is crucial for the prediction of the intestinal pharmacokinetics of drugs, with CYP3A4 contributing significantly to drug metabolism in humans. CM CYP3A8 has high amino acid sequence homology to, and the same substrate specificity as, human CYP3A4, suggesting that CYP3A8 corresponds to CYP3A4 [3]. The CMiPS cell-derived intestinal organoids exhibited CYP3A8 activity, which was almost completely inhibited by ketoconazole (Fig. 7A). Additionally, the activity of CYP3A8 was significantly induced by rifampicin (Fig. 7B), indicating that the organoids possessed functions related to drug metabolism. Besides CYP3A8, the expression of other CYPs, including CYP1A1, CYP2C43/2C9, CYP2C75/2C19, and CYP2D17/2D6, has been detected in the small intestine and several studies have reported that these drug-metabolizing enzymes are involved in the first-pass effects of drugs in CM small intestine [47,48]. The expression of CYP1A1 and CYP2C43/2C9 in the organoids was similar to that in the small intestine. On the contrary, the levels of CYP2C75/2C19 and CYP2D17/2D6 were lower in the organoids (Supplementary Fig. S2). It has been suggested that enzymes other than CYP3A8 could also be evaluated to investigate the first-pass metabolism of drugs in the intestinal organoids. However, in the present work, we only investigated a few pharmacokinetic functions of the organoids. To evaluate intestinal pharmacokinetics, the quantitative analysis of pharmacokinetic data in the organoids and the in vivo scenario is required.
CMs, which are closely related to humans, have been used in pharmaceutical research as preclinical models, to investigate the pharmacokinetic properties of drugs. CM pharmacokinetic parameters are more similar to humans, compared with those of mouse, rat, and dogs [6,7]. Similarly, Kim et al. reported that the CYP3A induction data from both in vitro and in vivo experiments in CMs are predictive of human CYP3A4 induction [25]. Therefore, we believe it is important to generate in vitro CM small intestinal models. On the other hand, the intestinal availability of some drugs in CM is lower than that in humans [15,16], regardless of the genetic similarity between the two species. However, when using CM intestinal organoids, the consideration of the species-related differences should allow the prediction of the intestinal availability of pharmacological drugs in humans. Notably, in this study, we found that the interior of the organoids corresponded to the apical side of the small intestine; therefore, when evaluating the intestinal pharmacokinetics of drugs, including metabolism and absorption, appropriate evaluation methods must be adopted.
We also expect that CMiPS cell-derived intestinal organoids will be useful for the development of cell transplant therapies in inflammatory bowel diseases (IBDs). iPS cells constitute a potential source of autologous intestinal epithelial cells for mucosa repair, and provide a major benefit over other sources of cells: the lack of immune rejection. Some studies have investigated the use of intestinal organoids for transplantation therapy in IBDs [49]. Shiba et al. reported that allogeneic transplantation of CMiPS cells leads to integrated graft survival and improves cardiac contractility in a CM myocardial infarction model [22]. Therefore, we believe that CMiPS cell-derived intestinal organoids will be an important research tool for allogeneic cell transplantation in humans.
In conclusion, we generated functional intestinal organoids from CMiPS cells. The intestinal organoids expressed intestinal markers and pharmacokinetic-related genes, and displayed pharmacokinetic functions, such as metabolic activity and inducibility of CYP3A8. We additionally found that several small-molecule compounds, especially the combination of A-83-01, PD98059, 5-aza-2′-deoxycytidine, and DAPT, were effective in inducing the differentiation of CMiPS cells into intestinal organoids. The intestinal organoids so generated may be useful as an in vitro evaluation system for the prediction of intestinal pharmacokinetics.
Footnotes
Acknowledgments
The authors thank Ina Research Co. for providing CM skin, and small and large intestinal tissues. This study was supported by grants from the Japan Society for the Promotion of Science (grant nos.: 26293036 and 16K15164); the Research on Development of New Drugs from Japan Agency for Medical Research and Development (15mk0101011h0102); and a Grant-in-aid for Research in Nagoya City University in 2016.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.