
Abstract
Due to the limited self-healing capacity of articular cartilage, innovative, regenerative approaches are of particular interest. The use of two-stage procedures utilizing in vitro-expanded mesenchymal stromal cells (MSCs) from various cell sources requires good manufacturing practice-compliant production, a process with high demands on time, staffing, and financial resources. In contrast, one- stage procedures are directly available, but need a safe enrichment of potent MSCs. CD271 is a surface marker known to marking the majority of native MSCs in bone marrow (BM). In this study, the feasibility of generating a single-stage cartilage graft of enriched CD271+ BM-derived mononuclear cells (MNCs) without in vitro monolayer expansion from eight healthy donors was investigated. Cartilage grafts were generated by magnetic-activated cell sorting and separated cells were directly transferred into collagen type I hydrogels, followed by 3D proliferation and differentiation period of CD271+, CD271−, or unseparated MNCs. CD271+ MNCs showed the highest proliferation rate, cell viability, sulfated glycosaminoglycan deposition, and cartilage marker expression compared to the CD271− or unseparated MNC fractions in 3D culture. Analysis according to the minimal criteria of the International Society for Cellular Therapy highlighted a 66.8-fold enrichment of fibroblast colony-forming units in CD271+ MNCs and the only fulfillment of the MSC marker profile compared to unseparated MNCs. In summary, CD271+ MNCs are capable of generating adequate articular cartilage grafts presenting high cell viability and notable chondrogenic matrix deposition in a CE-marked collagen type I hydrogel, which can obviate the need for an initial monolayer expansion.
Introduction
B
In two-stage procedures, monolayer-expanded MSCs are produced under good manufacturing practice (GMP) conditions, similar as performed in a chondrocyte-based therapeutic approach. The quality assurance and quality control, including testing of identity, purity, potency, stability, and sterility of the MSCs, have to be recorded according to European Regulation 1394/2007 [5,6], ensuring the maximum safety for these advanced therapeutic medicinal products. However, cellular expansion was defined as a “substantial manipulation” by the European Medicines Agency [6] as it influences the differentiation capacity of the cells and enhances the risk for contamination and genetic instabilities in these cellular products [7,8]. Apart from the accompanied challenges, the time to treatment is prolonged. In single-stage procedures, the articular cartilage defect can be evaluated during arthroscopy or open surgery intraoperatively, and a subsequent therapeutic intervention by isolation and enrichment of potent MSCs can be simultaneously addressed. Thus, the challenges of the time- and cost-consuming GMP expansion of MSCs and the inherent contamination risk might be reduced.
The common single-stage procedure is the microfracture of the subchondral bone leading to the infiltration of BM, forming a blood clot at the defect site and stimulating the recruitment of local MSCs from subchondral BM and synovial membrane [9,10]. Alternatively, the lesion can be coated with a concentrated BM aspirate containing autologous MSCs and chemokines. Nonetheless, both techniques provide only few potent MSCs contaminated with high numbers of hematopoietic cells in a frequency between 1 and 1.8 × 105 [2,11]. Moreover, only the treatment of small defects is recommended by microfracture (<2.5 cm2) or concentrated BM (<4 cm2) [12], and in the aftermath, the formation of mechanically unstable fibrocartilage in mid- to long-term follow-up is not uncommon [13].
Hitherto, a procedure overcoming time- and cost-consuming GMP expansion of MSCs, reducing the inherent contamination risk, avoids the (substantial) cell manipulation, and may fulfill the requirements for quality control and quality assurance of the cellular product is still absent.
The consideration of both single- and two-stage procedures underlines the advantages and disadvantages and additionally, the need for a single-stage procedure combining the opportunity of the direct treatment and the safety for the formation of a stable cartilage graft. A possible attempt to address this problem might be the intraoperative enrichment of MSCs in BM aspirates using surface markers.
A number of surface markers have been suggested for prospective isolation of MSCs from BM [14]. Among these surface markers, the low-affinity nerve growth factor receptor, CD271+ BM-derived MSCs, yielded excellent proliferation rate in monolayer culture and differentiation potential compared to MSCs selected by plastic adherence [15 –21].
Furthermore, CD271+ cells contain the majority of fibroblast colony-forming units (CFU-F) [16,22] and displayed an enhanced chondrogenic differentiation potential after monolayer expansion in a rat cartilage defect model [19].
Jones et al. in 2010 described CD271 as a suitable surface molecule to enrich MSCs from BM aspirates intraoperatively [21].
This in vitro study investigated the feasibility of generating cartilage grafts from human CD271+ BM cells in a CE-marked collagen type I hydrogel without initial monolayer expansion. Cell viability, DNA content, chondrogenic differentiation capacity, extracellular matrix secretion, and graft properties were monitored for up to 5 weeks to investigate the single-stage therapeutic approach for focal cartilage defects.
Material and Methods
The flow chart in Fig. 1 gives an overview of this study and is described in detail in the following sections:
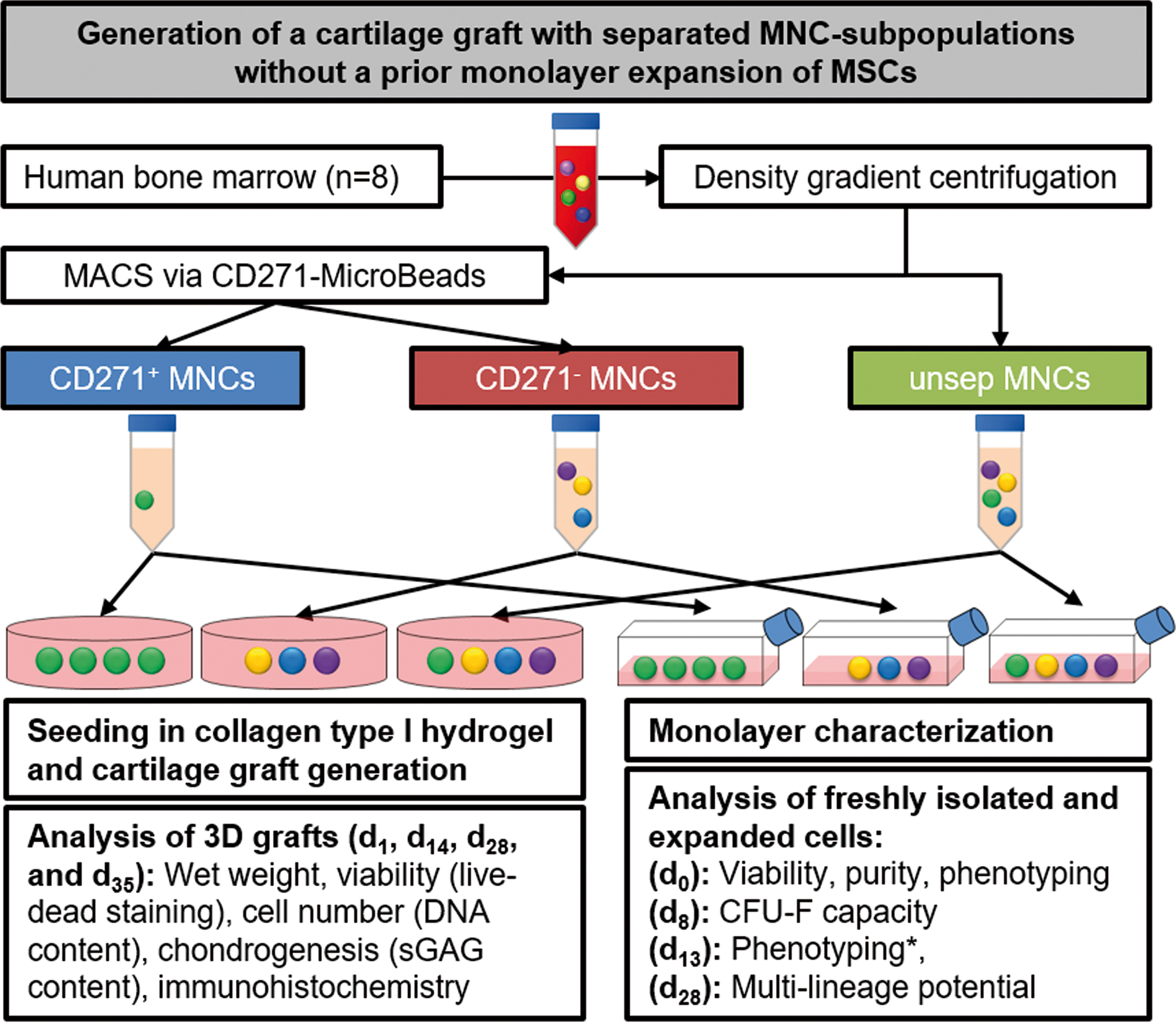
Overview of the study design. Cartilage grafts were generated from CD271
Ethics statement
The BM was aspirated as part of BM donations from eight donors without hematopoietic diseases, approved by the local ethics committee (State directorate of Saxony, EK-BR-86/14-1), and performed in accordance to Declaration of Helsinki. Macroscopically healthy cartilage was obtained during total knee arthroplasty for control immunohistochemistry. Tissue harvesting was approved by the local ethics committee (University of Leipzig, 190-14-02062014) and performed in accordance to Declaration of Helsinki.
All donors gave written informed consent. Baseline data were collected regarding donor's age and gender.
Cell isolation
Thirty to 60 mL of BM from the iliac crest was collected into a sterile heparinized syringe (170 IE/mL BM; Ratiopharm, Ulm, Germany) under anesthesia according to standard surgery protocol. Within 20–24 h after aspiration, whole BM was filtered with 70 μm mesh, diluted 1:3 with phosphate-buffered saline without Ca2+/Mg2+ (phosphate-buffered saline [PBS]; Biochrom, Berlin, Germany), and centrifuged through Ficoll paque premium (ρ = 1.077 g/mL; GE Healthcare, Buckinghamshire, United Kingdom) in Leucosep® tubes (Greiner Bio One) to obtain mononuclear cells (MNCs), including the MSCs. MNCs were washed twice with PBS and resuspended in DMEM (DMEM, high glucose, GlutaMAX™ supplement, pyruvate; Thermo Fisher Scientific, Dreieich, Germany). The number of viable cells was determined at each isolation step by dead cell exclusion by 4′, 6-diamidino-2-phenylindole (DAPI) staining (0.5 μg/mL in PBS; Sigma Aldrich, Munich, Germany) in the MACSQuant Analyzer 10 (Miltenyi Biotec, Bergisch-Gladbach, Germany). 1.2 × 107 MNCs were taken as unseparated (unsep) MNCs, as a reference cell population. The majority of the MNCs were subjected to separation by magnetic cell separation (MACS).
Magnetic cell separation
The MNCs were labeled with CD271 MicroBeads according to the manufacturer's protocol (all reagents Miltenyi Biotec) with minor modifications. First, nonspecific binding sites were blocked with 20 μL FcR-blocking reagent per 107 MNCs for 5 min. Afterward, cells were incubated with 20 μL CD271 MicroBeads per 107 MNCs for 30 min at room temperature in the dark. MNCs were washed with autoMACS Running Buffer and transferred onto the MACS column within a MACS Separator. Unlabeled cells were washed out and represent the group of CD271− MNCs. After removal of the column from the separator, CD271+-enriched MNCs were eluted with an autoMACS Running Buffer.
The cell fractions were used for the generation of 3D cartilage grafts without a prior monolayer expansion (see chapter 3D cultivation and chondrogenic differentiation) as well as expanded (2D) for 12–14 days for the characterization of cells according to the specifications of the International Society for Cellular Therapy (ISCT) [23].
The cells should fulfill “minimal criteria for defining multipotent mesenchymal stromal cells.” MSCs are defined by plastic adherence in monolayer cultivation as well as expression of CD73, CD90, and CD105 and deficiency of CD14, CD34, CD45, or CD11b, CD79α or CD19, and HLA-DR markers. In addition, MSCs must show multilineage potential by differentiating into adipocytes, osteoblasts, and chondroblasts [23]. These monolayer culture-based minimal criteria exclude a prospective isolation of MSCs from BM, as it would be essential for intraoperative single-stage therapy approaches. Nonetheless, these criteria enable a standardized definition of monolayer-expanded MSCs. To highlight the difference of monolayer-expanded MSCs and single-stage optimized enriched cell populations, all used cell fractions are named MNC fractions, as they also contain non-MSCs, for example, lymphocytes and monocytes.
Flow cytometry analysis
To characterize the cells and to control separation, freshly isolated and primary expanded MNCs were examined for the presence of CD271, CD73, CD90, and CD105 and the absence of CD14, CD19, CD34, CD45, HLA-DR, HLA-DP, HLA-DQ, and CD235a using direct labeled antibodies (Supplementary Table S1; Supplementary Data are available online at
Evaluation of CFU-F
Due to the different clonogenic capacity of cell fractions examined in preliminary experiments and reported previously [18], the initial seeding cell numbers were adjusted to 6.7 × 102 CD271+ MNCs, 1.0 × 105 CD271− MNCs, and 2.0 × 104 unsep MNCs/cm2 to determine the number of colony-forming cells from freshly isolated and separated cells. The cells were plated in 12-well plates and incubated with 1 mL DMEM/10% allogeneic AB-serum (Biowest, Nuaillé, France) at 5% pO2 and 5% pCO2. The medium was changed once a week. After 7 days, the cells were washed twice with PBS, fixed with methanol for 5 min, and colonies of more than ∼50 cells were counted using light microscopy (Zeiss Axiovert 200; Zeiss Microscopy, Jena, Germany). The number of CFU-Fs was calculated as mean from 6 to 12 wells.
Evaluation of osteogenic and adipogenic potential
To determine the osteogenic potential by mineralization, cells were seeded in a density of 3.0 × 104 cells/cm2 in 12-well plates in DMEM/10% allogeneic AB serum. The next day, the medium was exchanged to the osteogenic induction medium StemMACS OsteoDiff (Miltenyi Biotec) and renewed every 2–3 days. After 14–21 days, the cells were fixed with 4% formaldehyde, stained by von Kossa, and analyzed as described [24].
To determine the adipogenic potential through lipid deposition, cells after passage 1 were seeded into a well of a 12-well plate at a cell density of 5 × 102 cells/cm2 in 1 mL DMEM/10% allogeneic AB serum. The next day, the medium was exchanged to the adipogenic medium StemMACS AdipoDiff (Miltenyi Biotec) and replaced twice a week. On days 14 and 21, accumulated lipids in the cells were stained by Oil Red O and analyzed as described [24].
3D cultivation and chondrogenic differentiation
MNCs were seeded into a CE-marked, clinically approved 3D collagen type I hydrogel (Arthro Kinetics Biotechnology, Krems, Austria) according to the manufacturer's protocol without an initial in vitro monolayer expansion. Briefly, cells were mixed with a gel neutralization buffer (Arthro Kinetics Biotechnology) on ice and the collagen type I solution (2 mg/mL; Arthro Kinetics Biotechnology) was added. For each gel, 550 μL gel suspension was transferred into a well of a 24-well plate and polymerized for 20 min at 37°C. Due to the higher clonogenic capacity of the CD271+ MNCs determined in preliminary experiments, different cell numbers of each MNC fraction were seeded into the gels: 9.0 × 103 CD271+ MNCs, 1.125 × 106 CD271− MNCs, and 2.25 × 105 unsep MNCs/graft.
In BM of four donors of preliminary experiments, in mean, 2.7 × 106 MNCs per mL BM were obtained after density gradient centrifugation (Supplementary Table S2). Following CD271 MACS separation in this BM, 2.67 × 105 CD271+ MNCs were separated. Based on this observation, the study was conducted, leading to a seeding density of 9,000 CD271+ MNCs per graft due to the different time points, replicates, and the required cells for CFU assay, flow cytometry analysis, and 2D expansion for evaluation of osteogenic and adipogenic potential (1.4 × 105 CD271+ MNCs lost for gel production).
On the basis of previously published reports and approved cell therapeutics for cartilage repair [25] for the control group (unsep MNCs), a seeding density of 2.25 × 105 MNCs (≙ 1 × 106 cells/cm2) was chosen.
The grafts were then initially cultivated for 14 days in DMEM/10% allogeneic AB serum, followed by serum-free chondrogenic differentiation media (Lonza, Basel, Switzerland) supplemented with 10 ng/mL recombinant human transforming growth factor β3 (Miltenyi Biotec) for up to 21 days at 5% pO2 and 5% pCO2. The media were changed twice a week. The grafts were analyzed after 1, 14, 28, and 35 days.
The usage of human AB serum instead of fetal calf serum as supplement increased the proliferative activity of CD271+ MSCs at least in monolayer expansion [26]. Therefore, human AB serum was used in this study.
Analysis of 3D grafts
Cell viability was visualized by staining the grafts with ethidium homodimer and calcein AM (Live/Dead® Viability/Cytotoxicity Kit; Invitrogen, Karlsruhe, Germany) for 15 min at 37°C. After three washing steps with PBS, images of the grafts were taken with a fluorescence microscope (Zeiss Axiovert 200, Zeiss Microscopy) and analyzed.
To determine the DNA and secreted sulfated glycosaminoglycan (sGAG) content, the grafts were weighed and subsequently digested with 50 μg/mL papain (Sigma-Aldrich) at 60°C overnight. The DNA content in the samples was quantified with the Quant-iT PicoGreen assay (Thermo Fisher Scientific). The cell number/graft was calculated as 6.4 pg DNA per cell [27]. The sGAG content was measured with the dimethylmethylene blue (Sigma-Aldrich) assay [28]. All measurements were performed in triplicate according to the manufacturer's instructions.
For immunostaining, the grafts were shock frozen, bisected, and embedded in Tissue-Tek OCT Compound (VWR, Darmstadt, Germany). Eight micrometer cross-sections were stained for classical chondrogenic differentiation markers with monoclonal antibodies for aggrecan and collagen type II (Supplementary Table 1), detected by BrightVision goat-anti-mouse/rabbit/rat horseradish peroxidase (medac, Wedel, Germany) and stained with 3-amino-9-ethylcarbazole (AEC; Sigma Aldrich) substrate. Nuclei were counterstained with hematoxylin (DakoCytomation, Groß Lindow, Germany). Moreover, macroscopically healthy cartilage obtained during total knee arthroplasty was used as positive control for immunohistochemistry. Digital scans of the stained sections were taken with a Pannoramic Slides scanner (Sysmex, Norderstedt, Germany) and analyzed.
Statistical analysis
Data are presented as mean ± standard deviation. Comparative analyses of the non-Gaussian distributed data were performed by Kruskal-Wallis tests of the three cell fractions at the same time point. The level of significance was defined with P value <0.05. If measurements of individual fractions of a donor could not be performed due to the limited cell number, all values were withdrawn from the analysis of these time points. These events are indicated by varying n-values within the respective tables and figures. The statistical analyses were performed with Statistica version 12.7 (
Results
Identity and purity of the MNC fractions
BM aspirates (mean volume 43.3 ± 10.8 mL) were collected from the iliac crest of eight healthy adults (mean age: 28.9 ± 3.4 years; one female and seven male donors) and processed. The yield of unsep MNCs from BM after density gradient centrifugation was 5.24 ± 2.95 × 106 MNC/mL BM (range: 2.01–9.75 × 106 MNC/mL BM) with a viability of >98.5%. The majority of unsep MNCs were CD45+ leukocytes (95.42% ± 2.49%, Table 1). Only 0.05% ± 0.04% of the unsep MNCs were CD45−/CD271+, that is the putative MSC-containing fraction [21].
Mean % viable singlet MNCs ± SD and P values; significant P values are highlighted in bold; n = 8; * n = 6; ** n = 16.
SD, standard deviation; MNC, mononuclear cell.
The MACS separation by CD271 MicroBeads resulted in 1.32 ± 0.82 × 104 CD271+ MNCs/mL BM and 4.14 ± 2.00 × 106 CD271− MNCs/mL BM. The CD45−/CD271+ fraction in CD271+ MNCs was enriched by the factor 165.0 from unsep MNCs, with a purity of 31.72% ± 26.14% CD271+ cells. In parallel, CD45+ cells were reduced in CD271+ MNCs from 95.42% to 89.57%.
In the CFU-F assay, the number of colony-forming units was enhanced 66.8-fold in CD271+ MNCs (4551 ± 2833 CFU-F/106 cells) compared to unsep MNCs (68 ± 33 CFU-F/106 cells). CD271− MNCs contained 58.0 times less CFU-Fs (1 ± 1 CFU-F/106 cells).
Analysis of cell characteristics and multilineage potential
Monolayer expansion was performed to simultaneously characterize the MNC fractions according to the minimal criteria on MSCs reported by the ISCT [23]. Monolayer expansion was calculated as the ratio of gained cell number per 106 MNCs initially plated. After 13 days of monolayer expansion, the CD271+ MNCs contained the majority of plastic-adherent, proliferating cells (Table 2). CD271− MNCs showed no or minimal proliferation, although 25-fold higher initial cell numbers were initially plated compared to CD271+ MNCs. The unsep MNCs showed a reduced cell yield after primary monolayer expansion compared to CD271+ MNCs.
By cultivating monolayer-expanded cells with specific differentiation media, both adipogenic and osteogenic differentiation were seen in each MNC fraction. However, CD271+ cells showed a higher adipogenic lipid and osteogenic matrix deposition than CD271− MNCs, while unsep MNCs were comparable to CD271+ cells (Fig. 2).
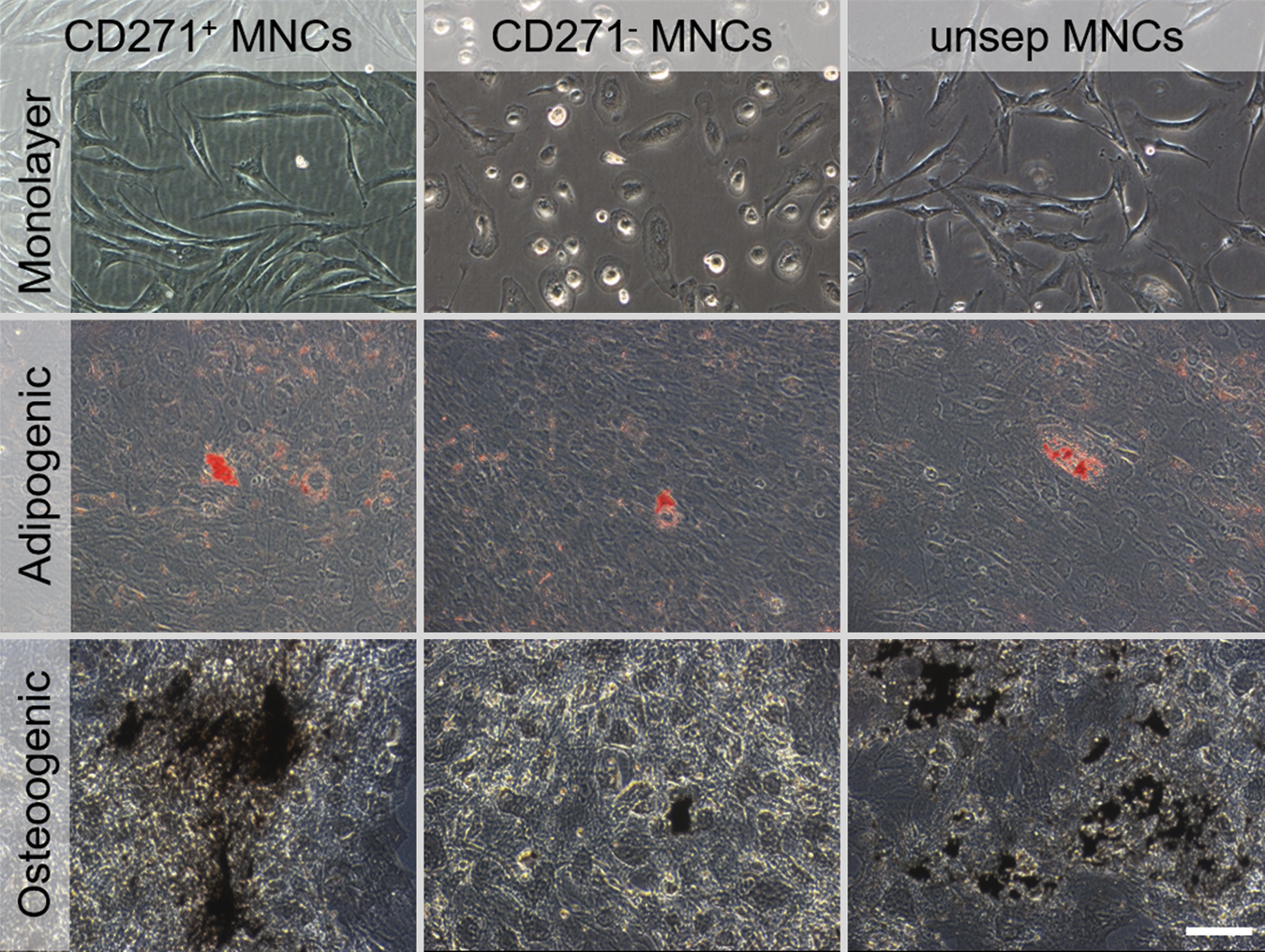
Characterization of monolayer-expanded MNCs. Phase contrast microscopy of all MNC fractions after analysis-related monolayer expansion (day 13). Oil Red O staining of the adipogenic differentiation and von Kossa staining of the osteogenic differentiation of the MNC fractions; scale bar 100 μm. Color images available online at
Flow cytometric characterization after monolayer expansion showed that the CD271+ MNCs fulfilled the ISCT criteria for MSCs, except for the slightly increased percentage of 4.56% ± 0.49% HLA-DR, HLA-DP, and HLA-DQ+ cells (Table 3). In the monolayer-expanded CD271− MNCs, the percentage of CD45+ and HLA-DR, HLA-DP, and HLA-DQ+ cells was increased. The monolayer-expanded unsep MNCs could not fulfill the MSC criteria concerning CD90 expression; however, the percentage of CD90+ cells was below the postulated 95% threshold (Table 3).
For the fulfillment of the criteria, CD73, CD90, and CD105 should be expressed ≥95%, while CD14, CD19, CD34, CD45, and HLA-DR, HLA-DP, and HLA-DQ should be ≤2% (mean % viable singlet MNCs ± SD and P values n = 6; * n = 16; ISCT criteria matches are marked in bold).
ISCT, International Society for Cellular Therapy.
In summary, CD271+ MNCs showed a distinct MSC phenotype with a spindle-shaped cell morphology after primary monolayer expansion regarding to plastic adherence, colony formation, and ISCT marker expression, as well as an adipogenic and osteogenic differentiation potential, thus they can be considered primary MSCs. CD271− MNCs showed a minor proliferation in monolayer culture with a flattened, globular cell morphology and did form no or only marginal colonies. Moreover, they displayed an increased expression of hematopoietic markers and nearly no multilineage potential, and therefore they did not fulfill minimal MSC criteria. Although the properties of the unsep MNCs were comparable to CD271+ MNCs, a higher amount of these cells are necessary to achieve similar effects related to the high amount of non-MSC cells in this fraction.
3D cultivation and chondrogenic differentiation
In preliminary experiments, a rapid cell death within the first 48 h was noticed without a proliferation phase, independent of the used fraction (CD271+ MNCs, CD271− MNCs, and unsep MNCs). Therefore, an initial cultivation period of 14 days in DMEM/10% allogeneic AB serum was necessary and performed in this study.
Generated grafts of CD271+ and unsep, but not CD271− MNCs yielded proliferating cell colonies after 14 days (data not shown), corresponding to the CFU-F formation. After 1 day of culture, the cell numbers corresponded to cell numbers at the time of seeding (CD271+ MNCs: 9.0 × 103; CD271− MNCs 1.12 × 106; and unsep MNCs: 2.25 × 105). The cell number increased within the grafts of CD271+ MNCs during cultivation (Fig. 3A; Supplementary Table S2). After the total cultivation time of 35 days, the cell number was 2.8-fold higher compared to the first day for CD271+ MNCs grafts. In contrast, CD271− MNCs did not proliferate in the grafts and unsep MNCs showed only a slight proliferation. On day 35, the cell number in grafts with CD271+ MNCs was 2.2-fold higher compared to grafts with unsep MNCs. These data were confirmed by qualitative live/dead staining of the cells within the grafts (Fig. 4). After seeding, only few viable CD271+ MNCs were present, whereas CD271− and unsep MNCs showed a high number of viable cells. Proliferation and colony formation could be especially seen in grafts with CD271+ MNCs and at a lower level, also in grafts with unsep MNCs during cultivation. In contrast, most of the CD271− MNCs died; after 35 days, nearly no viable cells were present.

Temporal characteristics of the MNC-seeded collagen I hydrogel grafts regarding cell number
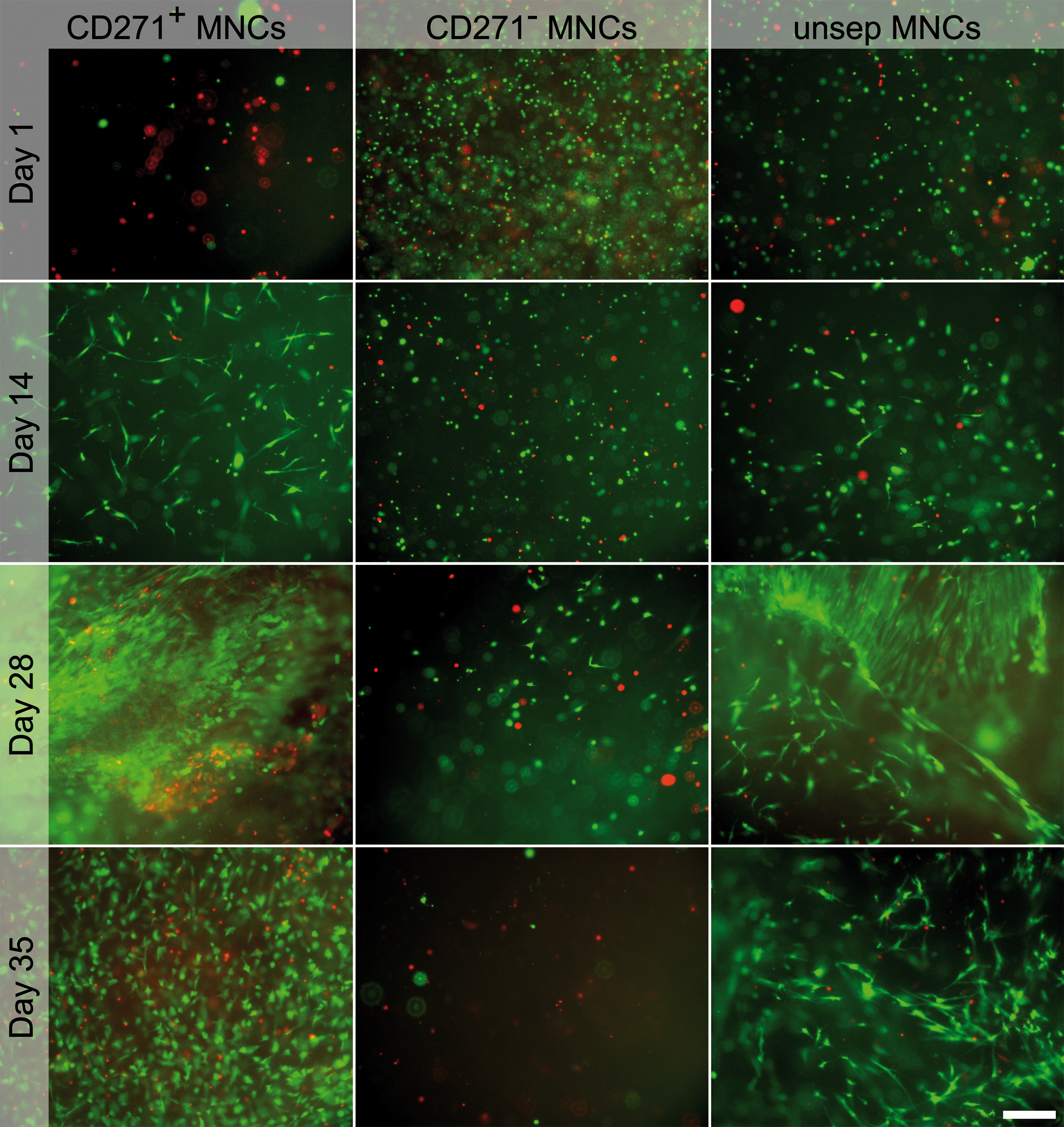
Live/dead staining of CD271+, CD271
To evaluate the chondrogenic differentiation potential and the amount of freshly produced extracellular matrix, the sGAG content was quantified within the grafts (Fig. 3B; Supplementary Table S3). The sGAG content of an acellular graft was 1.5–2.0 μg. In grafts with CD271+ MNCs, sGAG production increased over time and reached its maximum on day 35, whereas grafts with CD271− MNCs showed no measurable sGAG deposition. The amount of sGAG in unsep MNC grafts increased only slightly over the whole cultivation period.
After change to and supplementation with the chondrogenic differentiation medium on day 14, the grafts of CD271+ and unsep MNCs immediately began to shrink, while CD271− MNC grafts remained unchanged with a wet weight of about 241 mg each. In minimum, the CD271+ MNC grafts weighted 17 ± 4 mg/graft, while unsep MNC grafts weighted 31 ± 11 mg each after 35 days (Fig. 3C). Thus, proliferating cells induced shrinkage of collagen-based grafts in vitro.
Cryosections of the grafts after 35 days of culture were stained for the chondrogenic differentiation markers aggrecan and collagen type II (Fig. 5). The cryosections of grafts with CD271+ and unsep MNCs showed a homogenous distribution of the cells within shape of dominant cell colonies. In contrast, CD271− MNC grafts only rarely showed solitary colonies within the graft. Aggrecan and collagen type II staining varied considerably between the MNC donors (Fig. 5). Collagen type II positive staining was observed in CD271+ MNC grafts (5/8 donors) and unsep MNC (2/8) grafts. Due to individual grafts sizes, varying sizes of cryosections were present. In comparison to macroscopically healthy cartilage, 3D grafts of the CD271+ group yielded a proceeding extracellular matrix production (Supplementary Fig. S1).

Immunohistochemical staining for aggrecan and collagen type II of CD271+, CD271
As mentioned in the Material and Methods section, different seeding densities were used, possibly causing varying cell numbers and sGAG contents in the grafts related to differences in CFUs. Thus, donor-specific analyses of CFU contents and their relationship (CD271+ to unsep MNCs) were performed (Table 4). Indeed, higher CFUs in the CD271+ MNCs compared to the unsep MNCs resulted in an obvious higher cell number and sGAG content in the graft of CD271+ MNCs (eg, donor 1, Table 4). However, even if the CFUs in donors for CD271+ MNCs and the unsep MNCs were quite similar (eg, donor 2, Table 4), cell number and sGAG in grafts of CD271+ MNCs were higher compared to the unsep MNCs. Surprisingly, if the CFUs of the unsep MNCs were twofold higher than CD271+ MNCs (eg, donor 4, Table 4), cell number and sGAG content in the grafts of the CD271+ MNCs were still higher compared to the unsep MNCs. Therefore, the different seeding densities and/or resulting CFUs might have only a slight impact on the observed results.
sGAG, sulfated glycosaminoglycan; CFU, colony-forming units.
Discussion
Intraoperative strategies for the application of nonexpanded MSCs or MNC subpopulations from BM already exist [21] and may lead the way to broad clinical application beyond their homologous use for bone regeneration and repair.
Native MSCs are widely available in vivo, for example, from adipose tissue, umbilical cord blood, synovial membrane, synovial fluid, periosteum, skin, trabecular bone, infrapatellar fat pad, and muscle [1,29]. Apart from BM-derived MSCs, adipose tissue-derived MSCs are assumed as a promising alternative for cartilage repair related to their large availability and the easy access [30].
In contrast to other tissues sources, such as the umbilical cord, BM and adipose tissue CD271 can be utilized to isolate MSCs [20]. Moreover, adipose-derived MSCs displayed similar characteristics as BM-MSCs regarding their potential for chondrogenic differentiation in 3D culture.
Nevertheless, their quality in tissue engineering of cartilage is discussed controversial due to differing results regarding their chondrogenic potential in comparison to BM-MSCs [31 –35]. In addition, the outline translational single-stage preparation of MSCs for a cartilage graft might avoid substantial manipulating procedures, for example, enzymatic digestion of adipose tissue.
The utilization of a CD271+ MSC fraction (from BM) instead of unsep MSCs for cartilage repair strategies possibly has several benefits in vivo. The therapeutic superiority of CD271+ MSCs was recently shown in a rat cartilage defect model [19]. In this study, CD271+ MSCs showed a higher histological score in cartilage staining and fewer apoptotic cells when compared to the outcome of plastic-adherent unsep MSCs. In addition, intrinsic cartilage repair mechanism can also be accompanied by proliferating and matrix-deposing CD271+ cells either derived from the surrounding synovial membrane [36] or from the BM. Histological studies of knee and femoral heads from patients undergoing total knee or hip arthroplasty yielded the migration of CD271+ cells from BM through the tidemark in cases of full-thickness cartilage defects [37,38].
In this study, we show the feasibility of generating a cartilage graft from CD271+ BM-derived MNCs from eight donors in a CE-marked collagen type I hydrogel without previous monolayer expansion.
In the BM of the herein investigated eight healthy human donors, 0.01%–0.12% of the unsep MNCs were CD45−/CD271+. These observed frequencies were threefold higher when compared to published data [21,39], probably due to the young and uniform age of all donors in our study. Indeed, the frequency of native MSCs in BM depends on age [40] and various clinical comorbidities such as osteoarthritis [38] or myelodysplastic syndrome [41,42].
The direct MACS enriched the frequency of CD45−/CD271+ cells 165-fold and the number of CFU-F 66.8-fold in the CD271+ MNC fraction compared to unsep MNCs. These data are in line with previous studies reporting a 37-fold and a 54-fold CFU-F enrichment in CD271+ MNCs, respectively, and nearly no CFU-F formation in the CD271− MNC fraction [16,26,43]. As reported previously, CD271+ separation using indirect MACS sorting or clinical-grade CD271 MicroBeads resulted in a purity of 40%–50%, comparable to the here-obtained purity using direct MACS [43,44].
After seeding into collagen type I hydrogels, the CD271+ MNCs showed the highest proliferation rate, cell viability, sGAG deposition, and cartilage marker expression compared to CD271− and unsep MNC fractions. In grafts of CD271− MNCs, the cell number decreased continuously and cartilage matrix was absent. Final cell numbers and cartilage matrix deposition were higher in grafts from the CD271+ MNC fraction compared to grafts from unsep MNCs. For cartilage grafts, a 0.26 μg sGAG/mg wet weight value was determined. Previous studies measured sGAG contents of 20–50 μg sGAG/mg wet weight of the femoral head cartilage in elderly patients with a femoral neck fracture using the Alcian Blue staining [45]. Nevertheless, it should be emphasized that differences occur by differing locations, age groups, and measurement methods [46]. Moreover, the varying sGAG contents and their comparison might be limited comparing in vivo cartilage and in vitro cartilage grafts. In addition, sGAG released into the supernatant was not measured, leading to an underestimation of the sGAG content produced by the cells seeded in collagen type I grafts.
A variety of materials have been utilized as potential scaffolds for cartilage repair. Due to an appropriate biodegradability, biocompatibility, control over cell–matrix interactions, and similar mechanical behavior to articular cartilage, hydrogels are widely used [47,48]. Based on previous obtained preclinical and clinical results for this study, the collagen type I hydrogel (CaReS™, Arthro Kinetics, Esslingen) was chosen [49,50].
Indeed, nonsimilar seeding densities might be a pitfall of this study, but the obtained results yielded similar cell numbers, viability, sGAG content, and histological structure compared to the higher seeded unsep MNCs. These differences might be most plausible caused by the threefold higher CFU content of the CD271+ group leading to the here-presented data. Nevertheless, in donor-specific analyses of CFU contents, in cases of similar CFU contents of one donor, the CD271+ MNCs displayed better results (Table 4). Moreover, the CD271+ cell fraction was capable to produce similar cell numbers, even the CFU content of the unsep MSC in a donor was twofold higher.
In parallel to graft seeding, we expanded the various obtained MNCs by monolayer culture to evaluate them according to the minimal criteria of MSCs of the ISCT guideline [23]. CD271+ and unsep MNCs fulfilled these criteria in almost all points. HLA-DR, HLA-DP, and HLA-DQ expression were slightly higher as the ISCT limit in both cell fractions. However, the CD90 expression was reduced in unsep MNCs. Based on their marker expression, colony-forming and multilineage potential CD271− MNCs showed a non-MSC phenotype.
This study has several experimental limitations besides the above-mentioned variation of differing seeding densities. First, we could not correlate the numbers of CFU-Fs with the CD271+ MNC amount, because of donor-dependent variations with regard to separation frequency, cell purity, and yield. Second, as found in preliminary experiments, culturing gels without a proliferation phase were marked by a massive cell death within the first 48 h. Dexheimer et al. highlighted the close link of proliferation and differentiation in chondrogenesis. Therefore, a proliferation stimulating environment is a crucial basis for an efficient chondrogenic differentiation of MSCs [51].
However, for the use of a single-stage approach in vivo, the question arises to what extent a simultaneous proliferation and differentiation would take place. For this purpose, in vivo tests are necessary in future studies. This effect is well known from standard chondrogenic pellet culture [51] and only proliferating MSC colonies are capable to differentiate into chondrocytes. Third, the presented data focus only on BM-derived MSCs. Further MSC sources are known to give rise to potent MSC populations. Finally, it remains unclear which particular, minimal, dose of CD271+/CD45− cells ensures the optimal generation of a cartilaginous graft, which in turn induces effective and sustainable cartilage regeneration in vivo. However, the identification of the ideal cell dose was typically conducted in a phase 2 clinical trial [52].
In summary, because of the limited self-healing capacity of cartilage defects, the need for potent, safe, and easily applicable cellular therapies is underlined. In this study, MSCs are a promising cell source for cartilage regeneration. An intraoperative procedure to enrich MSCs without a previous monolayer expansion as shown here, will overcome a cost-, time-, and personnel-intensive GMP production of cartilage grafts. This is the first study demonstrating that CD271+ MNCs are capable of forming appropriate cartilage grafts with high cell viability and chondrogenic matrix deposition, in combination with a clinically approved collagen type I hydrogel without a monolayer expansion.
Footnotes
Acknowledgment
This study was supported by the Saxon Ministry of Science and Fine Arts (SMWK, grant no. 100124286 and 1315883) to O.P., C.S., and R.M.S. The authors like to thank Frank Peinemann and Christian Thuemmler for their technical assistance during the experiments.
Author Disclosure statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.