
Abstract
Obesity-associated erectile dysfunction (ED) involves pathologic change that may be related to deficit of the penile endogenous stem/progenitor cells. Therefore, an in-depth study of the penile stem/progenitor cells in the pathogenesis of ED is warranted. For this study, eight Zucker Lean (ZUC-Leprfa 186; ZL group) and 16 Zucker Fatty (ZUC-Leprfa 185; ZF) male rats received an intraperitoneal injection of 5-ethynyl-2-deoxyuridine (EdU) to track endogenous stem cells. Twelve weeks later, the ZF rats were randomized to gavage feeding with 1.5 mg/kg/day of icariside II (ZF + ICA II group) or the solvent (ZF group). Treatment lasted 4 weeks and was followed by a 1-week washout period. ZF rats had impaired erectile function with related pathologic changes compared with ZL rats. ICA II treatment restored erectile function and prevented smooth muscle atrophy, endothelial dysfunction, and lipid accumulation compared with no treatment. EdU label-retaining cell levels were higher in the ZF + ICA II group compared with the ZF group. Histone 3 phosphorylation at Ser 10, a specific mitotic cell marker, was additionally used to identify dividing cells. ICA II activated more penile stem cells to proliferate in ZF rats compared with ZL rats. These results suggest that ZF rats can be used as a model for obesity-associated ED and that ICA II improves erectile function and pathologic changes through endogenous progenitor cell preservation and proliferation.
Introduction
A
Most previous studies of OAED have used animal models with diet-induced obesity [2]. However, such a model is not ideal because most of those animals also develop diabetes, which itself can cause ED. However, Zucker Fatty (ZF) rats have a leptin receptor gene mutation that leads to glucose intolerance without diabetes mellitus [7]. Because these ZF rats are used to study genetic obesity, insulin resistance, glucose intolerance, and the metabolic syndrome, they may be a better animal model of OAED. Several reports have demonstrated ED in these animals, but have not described the underlying mechanism [8,9].
Herba Epimedii, a traditional Chinese medicine, has been used for centuries in East Asia to treat a variety of human illnesses, including sexual concerns. Icariin (ICA), a flavonol glycoside, is the most metabolically active extract obtained from the Herba Epimedii plant. Our previous studies indicate that ICA produces erectogenic effects in vivo. Icariside II (ICA II), the bioactive form of ICA, has higher bioavailability compared with ICA. The ICA II compound has been demonstrated to improve erectile function in rodents with type 1 diabetes mellitus through preservation of smooth muscle, endothelial function, and nitric oxide synthase activity [10]. Furthermore, ICA II has been used to enhance endogenous stem cell differentiation in rats with bilateral cavernous nerve injury [11]. To our knowledge, the utility of ICA II to potentiate erection in an animal model of obesity has not yet been explored.
In this study, we administered ICA II to ZF rats as a daily supplement. Rats underwent functional erectile hemodynamic testing and histological and molecular assessment of corpus cavernosum tissues to determine how obesity affects penile structure and function. We also investigated the feasibility of ICA II treatment and the mechanisms underlying its therapeutic effect in rats with OAED.
Materials and Methods
Animals
Experiments were approved by the Institutional Animal Care and Use Committee at the University of California, San Francisco. Eight newborn male Zucker Lean (ZL) rats (ZUC-Leprfa 186) and 16 ZF rats (ZUC-Leprfa 185) were used in this study. To track putative endogenous stem cells, each pup received an intraperitoneal injection of 5-ethynyl-2-deoxyuridine (EdU; Invitrogen, Carlsbad, CA) (150 mg/kg) at birth, as previously described [12]. Body weight was measured every 2 weeks. At 10 weeks, all rats were tested for insulin tolerance. At 12 weeks, ZF rats were given ICA II (Suzhu Guang'ao Healthcare Co., Ltd) (1.5 mg/kg/day; ZF + ICA II group) or solvent (vehicle only; ZF group) by gavage feeding for 4 weeks (n = 8 per group). This dose was chosen based on our previous work. Following a 1-week washout period without ICA II, erectile function was evaluated by measuring intracavernosal pressure (ICP) and mean arterial pressure (MAP). After functional testing, the animals were sacrificed and penile tissues were preserved for histological study.
Intraperitoneal insulin tolerance
All rats were fasted for 6 h with access to drinking water. Basal blood glucose levels were measured before insulin injection. Fast-acting insulin (Sigma-Aldrich, St. Louis, MO) was given at a dose of 0.5 IU/kg by intraperitoneal injection. Blood was obtained by tail puncture at 30, 60, 90, and 120 min postinsulin administration. Blood glucose was measured using an Accu-Chek Active blood glucose meter (Roche Diagnostics, Indianapolis, IN). Glucose levels are presented as percentages of baseline plasma glucose.
Erectile function
The ICP was measured as previously described. Briefly, using ketamine (100 mg/kg) and midazolam (5 mg/kg) anesthesia, the major pelvic ganglion and cavernous nerve were exposed through midline laparotomy. A 25-gauge butterfly needle was inserted into the left penile crus to monitor ICP. A PE-50 tube filled with heparinized saline (200 IU/mL) was cannulated into the left common carotid artery and connected to a pressure transducer (Utah Medical Products, Midvale, UT) to continuously monitor MAP. Erectile response was elicited by electrical stimulation using a bipolar electrode hooked to the cavernous nerve. Stimulation parameters were 1.5 mA current, 20 Hz frequency, 0.2 ms pulse width, and 50 s duration. The maximum ICP increase measured from three stimuli applied to each side of every rat was used for statistical analysis. The ICP to MAP ratio (ICP/MAP) was calculated to standardize results for systemic blood pressure variability. Total ICP was calculated as the area under the curve during stimulation.
Immunofluorescence and lipid droplet staining
Cryosections of penile tissue samples were prepared according to a previously described protocol [13]. Tissue sections were incubated with primary antibodies, including anti-actin, α-smooth muscle (α-SMA) antibody (1:1,000; Sigma-Aldrich), mouse anti-rat RECA-1 antibody, clone HIS52 (1: 500; Bio-Rad, Hercules, CA), and anti-phospho-Histone H3 (Ser10) antibody (H3P; 1:100; Merck Millipore, Billerica, MA). Smooth muscle was stained using Alexa Fluor 488 phalloidin (1:500; Invitrogen). Sections were assessed for EdU labeling by using the Click-iT reaction cocktail (Invitrogen) containing Alexa Fluor 594 (1:500) for 30 min at room temperature, then subjected to nuclear staining with 4′,6-diamidino-2-phenylindole, dihydrochloride (DAPI; 1 μg/mL; Invitrogen).
To identify the location of lipid deposition in penile tissue, phalloidin staining was followed by LipidTOX neutral lipid stain (1:200; Invitrogen) at room temperature for 30 min. Nuclear staining was performed by incubation with DAPI.
Masson's trichrome staining
Penile tissue sections were prepared as described above. Masson's Trichrome staining was used according to the standard protocol [14]. Semiquantitative analysis was performed to evaluate intensity using Image-Pro plus software (Media Cybernetics, Silver Spring, MD).
Image analysis and quantification
For image analysis, four randomly selected fields per tissue sample for each experimental and control animal were photographed and recorded using a Retiga Q Image digital still camera and ACT-1 software (Nikon Instruments, Inc., Melville, NY). To evaluate penile lipid accumulation, photographs were taken of five fields in the sections at 200 × magnification. To quantify lipid accumulation, the red-colored LipidTOX-positive area was measured and expressed as a percentage of the total area.
Statistical analyses
Results were analyzed using GraphPad Prism version 6.02 (GraphPad Software, San Diego, CA) and expressed as mean ± standard deviations. Statistical analyses were performed using one-way analysis of variance followed by the Tukey's multiple comparisons test. Differences among groups were considered significant for P values less than 0.05.
Results
Metabolic variables
Body weight of ZF rats was higher than that of ZL rats starting at 6 weeks of age (P < 0.001; Fig. 1A). During the insulin tolerance test at 10 weeks of age, blood glucose levels dropped sharply in ZL rats, whereas they declined slowly and rebounded at 60 min in ZF rats.

Growth chart and insulin tolerance test.
At 12 weeks of age, ZF rats had a higher mean body weight and blood glucose level compared with ZL rats. At both the week 12 randomization and after a 4-week treatment, ZF and ZF + ICA II groups did not differ with respect to body weight or blood glucose level (Table 1).
Data are presented as mean ± standard deviations.
P < 0.05 compared with the Zucker Lean rats.
ZF, Zucker Fatty rats; ZF + ICA II, Zucker Fatty rats treated with icariin; ZL, Zucker Lean rats.
ICA II treatment improves erectile function in ZF rats
As illustrated in Fig. 2, the ZL group had normal ICP curves and high ICP/MAP ratios. In contrast, the ZF group had lower ICP with a curve fluctuation at its plateau (0.78 ± 0.07 vs. 0.42 ± 0.11, P < 0.001). ICA II treatment increased the ICP/MAP ratio in response to cavernous nerve electrical stimulation (0.68 ± 0.08, P < 0.001).
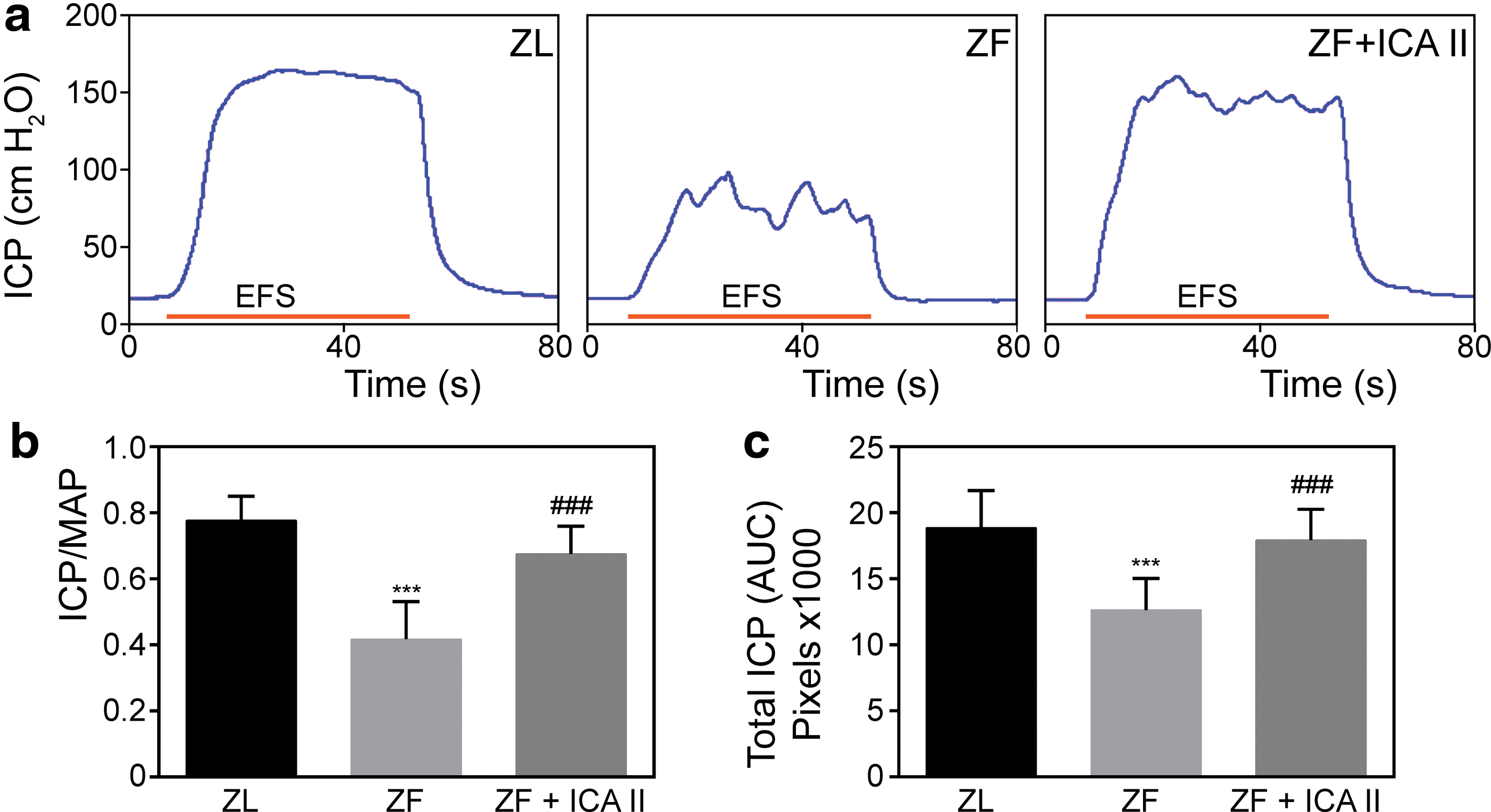
ICA II improved erectile function upon cavernous nerve electrostimulation in ZF rats.
ICA II treatment improves smooth muscle content and the smooth muscle to collagen ratio in the corpus cavernosum
Obesity was associated with cavernous smooth muscle atrophy, as evidenced by decreased smooth muscle content in penile tissue from ZF rats compared with ZL rats. The ICA II treatment group experienced smooth muscle regeneration and reduced cavernous fibrosis. The smooth muscle/collagen ratio was 0.227 ± 0.039 for the ZL group, 0.140 ± 0.017 for the ZF group, and 0.208 ± 0.022 for the ZF + ICA II group (Fig. 3).
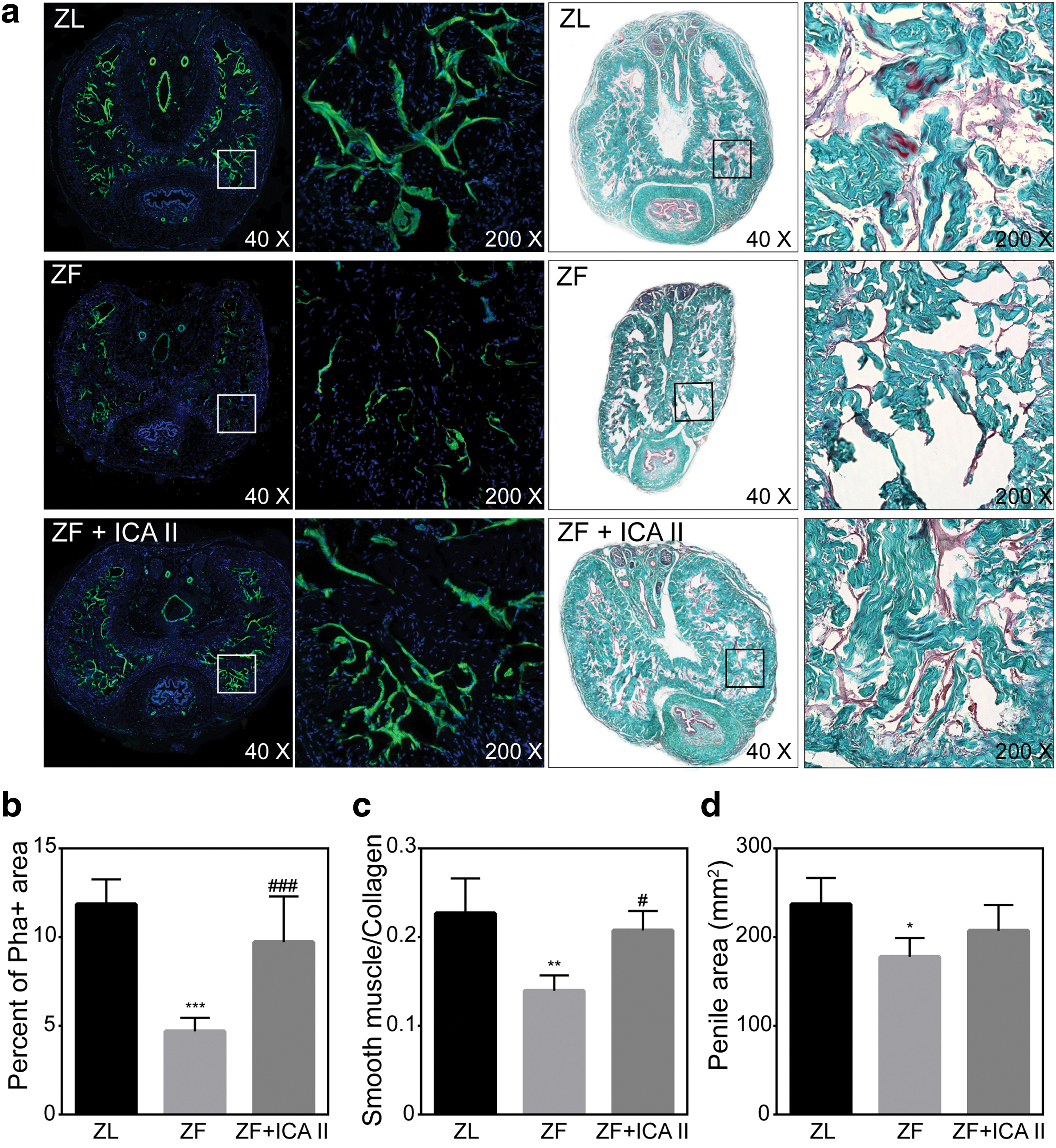
Phalloidin staining and Masson's Trichrome staining of penile mid-shaft specimens.
ICA II treatment reverses endothelial damage in ZF rats
As shown in Fig. 4, RECA-1 (a specific endothelial marker in rats) markedly decreased in the ZF group compared with the ZL group. At high magnification, ZF samples had evidence of endothelial monolayer disruption, whereas ZF + ICA II samples had preserved endothelial sinusoid lining.

RECA-1 staining of cavernosal sinusoid.
ICA II treatment suppresses lipid droplet accumulation in the corpus cavernosum of ZF rats
Abundant lipid droplets accumulated within the corpora cavernosa of ZF rats, but few droplets were observed in ZL rats. Compared with ZL rats, ZF rats had a higher percentage of LipidTOX+ areas in the penile tissue (4.85 ± 0.64 vs. 25.62 ± 2.49; P < 0.001). Figure 5 compares lipid droplet infiltration in the cavernous sinusoids for the ZL, ZF, and ZF + ICA II groups. ICA II treatment was associated with decreased lipid droplet accumulation in penile tissue (11.66 ± 2.29).
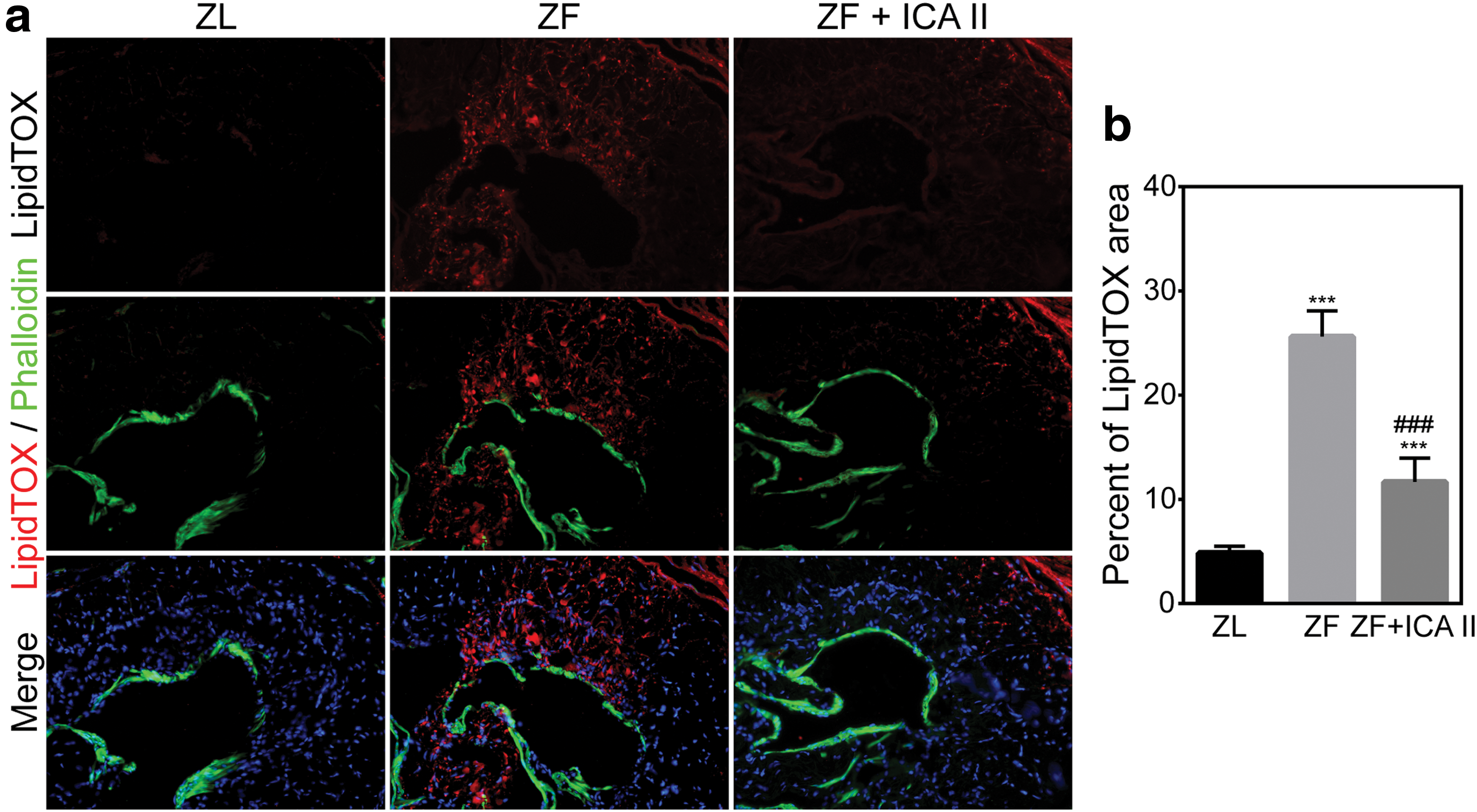
Lipid accumulation in the corpus cavernosum for ZL, ZF, and ZF + ICA II groups.
ICA II treatment activates EdU+ progenitor/stem cells in the corpus cavernosum
Progenitor cells and those with stem cell properties are characterized by their ability to retain nucleotide analogs, such as BrdU or EdU, for prolonged periods. Therefore, we examined EdU+ cells to track progenitor cells in the penis. Compared with ZL rats, ZF rats had a decreased total number of EdU+ cells (P < 0.05). Treatment with ICA II dramatically increased the total number of EdU+ cells. However, the EdU+ cell distribution pattern differed from that of ZL rats. As shown in Fig. 6, rats in the ZF + ICA II group had a larger proportion of EdU+ cells localized to the parasinusoidal area, although most EdU+ cells were localized to the subtunical region in all three groups.
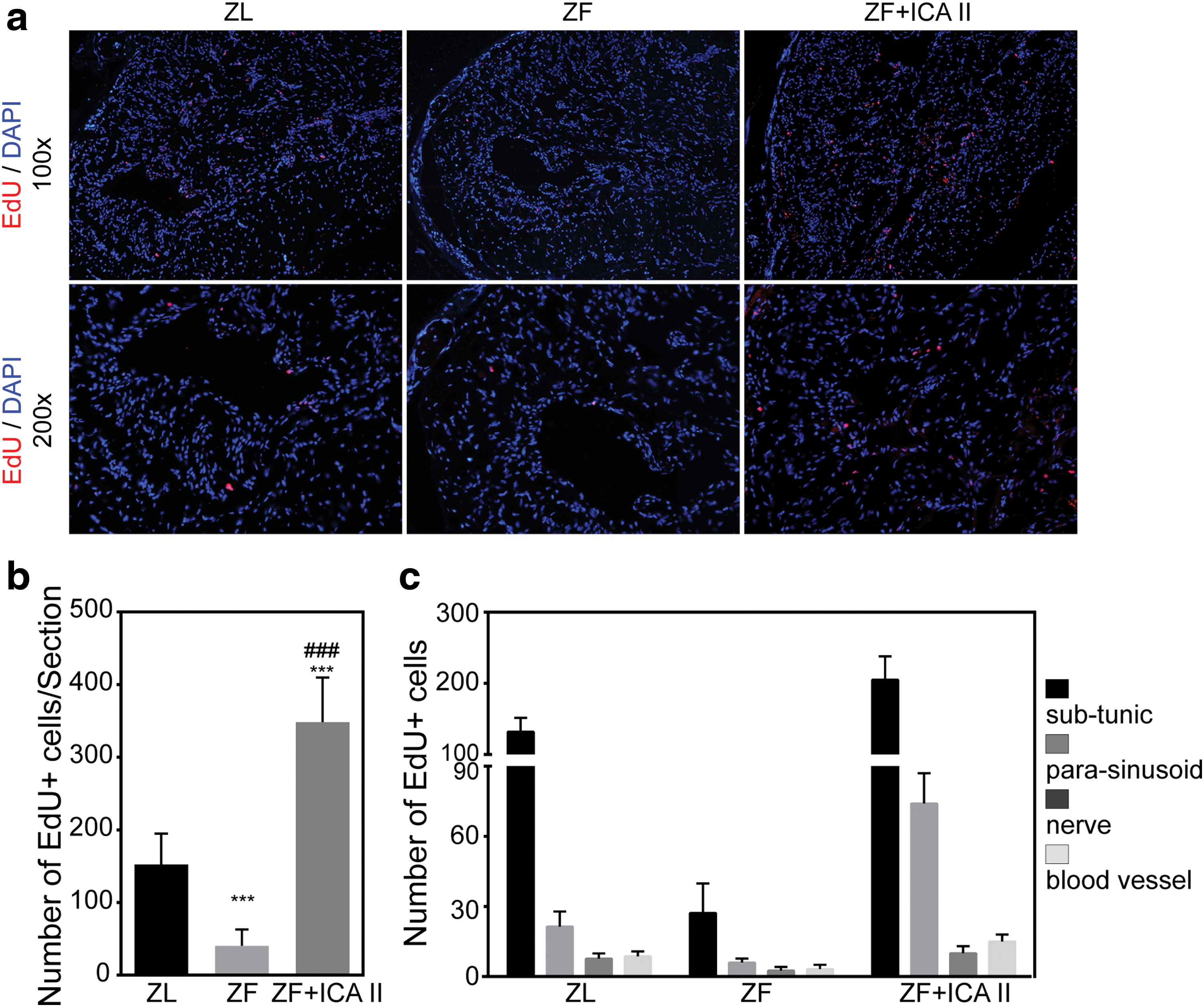
Evaluation of LRCs.
Activated EdU+ cells were tracked by staining with differentiated cell markers. Label-retaining cells (LRCs) coexpressing α-SMA or RECA-1 were found in the cavernosal sinusoid, and sinus lining, respectively (Supplementary Fig. S1; Supplementary Data are available online at
ICA II treatment promotes endogenous stem/progenitor cell proliferation in ZF rats
To test whether the increase in EdU+ cells resulted from residential stem cell proliferation, we determined the number of cells staining positive for H3P (a nuclear protein indicative of cell proliferation). Although ZL and ZF rats did not differ in this regard, H3P+ cells were increased in ZF + ICA II rats (Fig. 7a, b). Only 4.17% of the H3P+ cells were EdU+, and the distribution pattern of these two markers is shown in the partial magnification of Fig. 7c. The image suggests that two cells are undergoing mitosis, with the one retaining the EdU marker likely to be the mother cell and the other one likely to be the daughter cell.
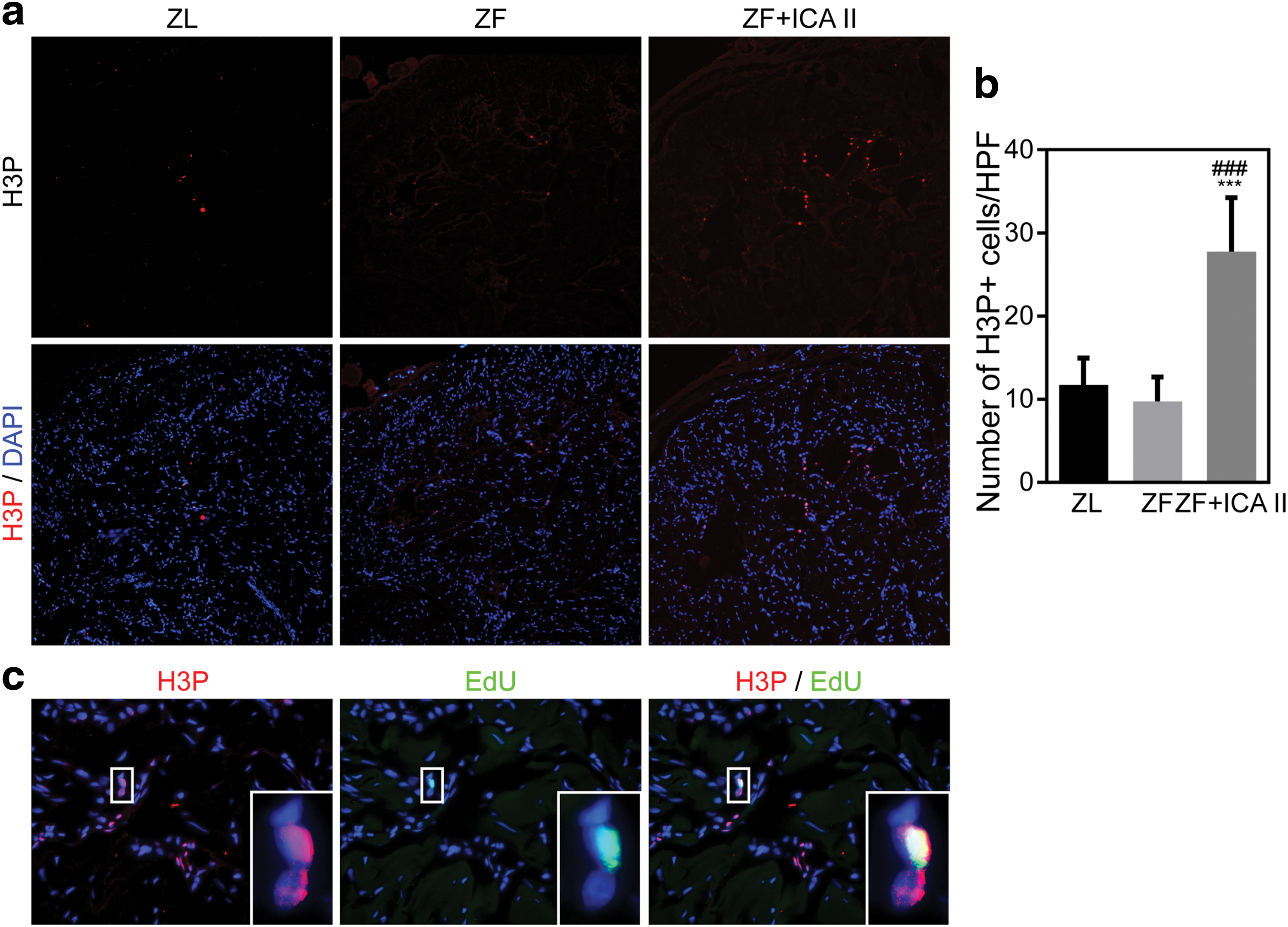
ICA II promoted endogenous stem/progenitor cells proliferation.
Discussion
In recent years, various pathologic subtypes of ED (eg, diabetic, neurogenic, and vasculogenic) have been intensively investigated [11,14 –16]. Multiple epidemiological studies have shown strong associations between obesity and ED across populations [17 –20]. Because of the lack of a suitable animal model, the mechanisms underlying OAED pathology are not well understood, and no targeted therapies have been developed. Furthermore, currently available pharmacologic treatments are less effective in men with obesity, and their use cannot prevent progression to irreversible ED [2]. It is, therefore, essential to determine the ideal animal model to study this common medical condition. In this study, we used ZF rats, which are commonly used to study genetic obesity without spontaneous diabetes, to demonstrate ED as measured by decreased maximal ICP and total ICP.
Cavernous smooth muscle, the predominant component of erectile tissue, is responsible for blood flow control into the corpus cavernosum. In ED, smooth muscle atrophy and penile fibrosis are associated with increased apoptosis and reduced smooth muscle content. In the clinic, many obese men complain of reduced penis size, but physicians generally believe that the small appearance is related to the presence of the suprapubic fat pad covering a portion of the penis. However, Sanchez et al. [9] showed that ZF rats have smaller penile arteries compared with ZL rats (because of obesity-associated testosterone deficiency), which leads to decreased blood supply to the penile tissue. Our results show a reduced penis cross-sectional area in ZF rats that is explained by cavernosal smooth muscle atrophy. Thus, a small penis in obese men may be partially due to penile smooth muscle atrophy.
Obesity is associated with endothelial dysfunction, which occurs with elevated free fatty acid levels produced by adipose tissue, and with insulin resistance [21]. Endothelium plays a vital role in the initiation/maintenance of penile erection [3]; thus, morphologic endothelial changes could be associated with dysfunction. In the present study, ZF rats had “patch-like” endothelial cells, whereas ZL rats had intact endothelium lining the cavernous sinuses. This difference may be attributable to elevated free fatty acid levels and insulin resistance, which increase reactive oxygen species levels and cause pathological inflammation of the vascular endothelium.
ZF rats also had profound lipid accumulation in the penile tissue, which may impair veno-occlusion that is necessary for erection [22]. In ZF rats, this lipid infiltration was present from the tunica albuginea to the central cavernosal smooth muscle, with further lipid droplets located in the subtunical region (Supplementary Fig. S2). Indeed, this structure-related dysfunction may lead to the fluctuating plateau phase of the erection in the ICP recording (Fig. 2a).
ICA and ICA II have garnered attention as potential treatments for sexual dysfunction because of their inhibitory effects on PDE5. The current study provides functional evidence that ICA II can improve erectile function in obese rats. The observed effects may extend beyond PDE5 inhibition, as we detected significantly higher smooth muscle content and fibrosis regression in ZF + ICA II rats compared with ZF controls. It has also been suggested that RECA-1 is upregulated after ICA II treatment, indicating that ICA II may promote endothelial regeneration. Prior work has supported a role for ICA and its derivatives in enhancing erectile function both in vitro and in various animal models. Li et al. found that ICA II preserves cavernous endothelial cells' proliferative capacity under high glucose conditions and restores endothelial nitric oxide synthase expression [23]. Furthermore, an in vivo study showed that ICA II restores endothelial and smooth muscle components and decreases lipid peroxidation levels in diabetic rats with ED [12]. Our previous study finding that ICA II can improve erectile function in a rodent model with neurogenic ED by enhancing endogenous stem cell differentiation [11] seems to be a promising therapy for ED. Consistent with previous studies, the present study confirmed the rehabilitative role of ICA II for endothelial and smooth muscle components in penile tissue. More notably, we found that ICA II decreased lipid accumulation in penile tissue of obese rats. ICA II has shown its ability to inhibit adipogenetic stem cell differentiation [24]; thus, it reduces the source of fat-containing cells and preserves residential stem/progenitor cells for tissue repair.
Although exogenous stem cell treatment for ED has been studied for decades, undesirable side effects in off-target tissues have limited researchers' enthusiasm in this approach. Since penile stem/progenitor cells were first identified in 2015 [13], several studies have been designed to boost repair activity in penile tissue through activation of local progenitor cells [11,25]. In the present study, the label-retaining EdU technique demonstrated that penile tissue treated with ICA II had more stem/progenitor cells, which is consistent with the process of tissue repair [16].
It has been reported that even young animals may experience stem cell niche remodeling that impacts tissue regenerative potential [26], particularly when exposed to prolonged metabolic disruption (eg, obesity) [27]. In this study, the ZF group had fewer EdU+ cells present in the penile tissue, and the injured organ failed to regenerate without adequate stem cells. Several studies have demonstrated that ICA II extends the healthy lifespan through the insulin/IGF-1 pathway that is related to endogenous stem cell activation [28]. We found that the number of LRCs was significantly higher in penile tissue in the ZF + ICA II group compared with the ZF group. More importantly, the proportion of LRCs located in parasinusoidal areas was much higher following ICA II treatment. We theorize that activated stem/progenitor cells may differentiate into various functional parasinusoidal cells. This hypothesis is supported by the finding that α-SMA and RECA-1 are coexpressed with EdU in penile tissue, which indicates that the increased LRCs were committed to differentiate into functional cells.
Histone 3 phosphorylation at Ser10, a conserved event that occurs during mitosis [29], is regarded as a specific marker for mitotically active cells [30]. Our study finding of more H3P+ cells in the ZF + ICA II group indicates that ICA II activates more cells and promotes cell proliferation [31]. Double labeling for H3P and EdU, which was intended to identify endogenous stem/progenitor cell proliferation, showed that only 4.17% of H3P+ cells were EdU+. Intriguingly, we found a pair of H3P+ cells in the ICA II-treated rats, in which only one retained EdU positivity. Initial stem cells in the penis may generate two types of cells after asymmetric cell division: EdU+ mother cells that incorporate EdU during S-phase at birth and H3P+ EdU- daughter cells that are generated by the stem cells. The daughter cells maintain their proliferative ability and can serve as progenitor cells for tissue regeneration. This explanation may account for the low proportion of EdU positivity in H3P+ cells.
In summary, obesity impairs erectile function by consuming endogenous stem cells, which leads to smooth muscle atrophy, endothelial dysfunction, and lipid accumulation in the corpus cavernosum. ZF rats can serve as a novel animal model in which to study OAED. ICA II restores normal penile hemodynamic parameters and improves functional outcomes in association with preserved muscle content and endothelium. The underlying mechanisms of ICA II therapy are activation of endogenous stem/progenitor cells, promotion of cellular proliferation, and acceleration of penile tissue regeneration. Our ongoing research aims to determine how ICA II activates stem cells using molecular biology techniques. Our current findings are of interest not just to validate this traditional treatment for ED, but to introduce a new method by which endogenous stem/progenitor cells may be modulated to treat other diseases.
Footnotes
Acknowledgment
This work is supported in part by the China Scholarship Council under the grant CSC 20160616110.
Author Disclosure Statement
Zhongcheng Xin and Guiting Lin are consultants to Suzhu Guang'ao Healthcare Co., Ltd. All other authors have no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.