
Abstract
Continuous spermatogenesis from puberty to old age in males relies on spermatogonial stem cells (SSCs) that possess the property of self-renewal and differentiation. The delicate balance between self-renewal and differentiation is of great importance. In mice, SSCs exist as a subpopulation of undifferentiated spermatogonia. SSCs are controlled by intrinsic molecular pathways that can be activated by extrinsic signals. Our results here first show that the expression of forkhead box C2 (FOXC2) is restricted to GFRα1-positive spermatogonia in the testis. Whole-mount immunofluorescence results reveal that FOXC2 is expressed predominately in As and Apr spermatogonia. Reduction of Foxc2 gene expression by shRNA lentivirus treatment significantly impairs the maintenance of SSCs in vitro. Furthermore, knock-down of Foxc2 decreases SSC colonization to only 10.42% compared to the control by transplantation. Reverse transcription and real-time quantitative PCR gene analyses following knock-down of Foxc2 indicate that Foxc2 may act as a suppressor for SSC differentiation. Extrinsic stimuli treatments show that glial cell line-derived neurotrophic factor and retinoic acid act in opposite ways to regulate FOXC2 expression and subsequent SSC property. These results suggest that FOXC2 is a critical intrinsic regulator of SSC self-renewal and differentiation.
Introduction
C
The study of SSC biology was difficult initially because of their rarity in the testis, estimated to be 0.02%–0.03% of all germ cells in mouse testis and the complex niche microenvironment [4]. Fortunately, a robust transplantation technique developed in 1994 made it possible to unequivocally quantify and characterize SSCs based on their functional ability to reestablish spermatogenesis [5,6]. In 2003, a long-term in vitro culture system that supports self-renewal and differentiation of mouse SSCs was established [7]. Although the number of SSCs in testes is very low, this culture system allows SSCs to proliferate rapidly in vitro and to be collected for molecular and biochemical analyses.
The prevailing mouse SSC model assumes that the stem cell population resides in the As population and that cyst length reflects the extent of differentiation [3,8,9]. A corollary of this “As model” is that all As spermatogonia are homogeneous. This “As model,” proposed in 1971, has provided the framework of SSC research in mice and other animals for many years. However, several recent studies show that undifferentiated spermatogonia were heterogeneous in the expression of several genes such as NANOS2, GFRα1, NANOS3, and NGN3 [10 –12]. GFRα1 and NANOS2 are expressed in large subsets of As and Apr and in small subsets of Aal. Conversely, NGN3 and NANOS3 are expressed mainly in Aal while rarely in As and Apr. The As spermatogonia are not homogeneous. Thus, As, Apr, and Aal populations can be stratified both morphologically (cyst length) and by gene expression. NANOS2+ or GFRα1+ SSCs are called “actual stem cells,” while NANOS3+ or NGN3+ SSCs are referred to as “potential stem cells” [12 –15]. Potential stem cells possess the potential to self-renew but do not do so in undisturbed testes. However, during regeneration after damage, they retain latent self-renewing potential.
Forkhead box (FOX) proteins are a family of transcription factors present in various organs and tissues during development and play crucial roles in a variety of processes such as cell growth, proliferation, differentiation, and longevity [16 –18]. FOX proteins are characterized by the FOX, a sequence of around 100 amino acids forming a motif that binds to DNA [19,20]. Because of the butterfly-like appearance of the loops in the protein structure of the domain, this forkhead motif is also known as the winged helix [21]. Forkhead box C2 (FOXC2), also known as mesenchyme forkhead protein (MFH1), belongs to the forkhead “C” subfamily [22]. FOXC2 is required for cardiovascular development, early organogenesis of the kidney and adipocyte differentiation [23 –25]. Recently, FOXC2 was found to be associated with epithelial–mesenchymal transition and stem cell properties in breast cancer [26]. FOXO1, another FOX family transcription factor, plays a critical role in control of SSC self-maintenance and spermatogenesis initiation [27]. Foxc2 was found to be overexpressed in GFRα1-positive cells of the 6-day-old mouse testis [28]. However, the role of FOXC2 in SSCs has yet to be explored. In this study, we investigated the expression and function of FOXC2 in SSCs and demonstrated that FOXC2 is required for maintaining murine SSCs.
Materials and Methods
Animals
DBA/2, C57BL/6 and C57BL/6-Tg (CAG-EGFP) 131Osb mice were used in this study. All animals were kept in a specific pathogen-free environment. All animal procedures were approved by State Key Laboratory of Agrobiotechnology, China Agricultural University.
Immunofluorescence
Mouse testes were fixed in 4% PFA for 2–12 h according to their size at 4°C, incubated in 30% sucrose in phosphate-buffer saline (PBS) overnight at 4°C and embedded in O.C.T. compound. Ten micrometers sections were washed in PBS and blocked in 10% normal donkey serum in PBS for 1 h at room temperature (RT). Then, the sections were incubated with primary antibodies against FOXC2 (1:100; AF6989; R&D) and PLZF (1:100; sc-22839; Santa Cruz) in PBS overnight at 4°C. After washing, the sections were incubated with secondary antibodies of NL557-conjugated donkey anti-sheep IgG (1:200; NL010; R&D) and Alexa Fluor 488-conjugated donkey anti-rabbit IgG (1:200; A-21206; Invitrogen) in PBS for 2 h at RT. After washing, the sections were counterstained with DAPI (1:500; 10236276001; Roche) for 15 min and observed using a fluorescence microscope photograph system (IX73; Olympus). Immunofluorescence of other cell markers was performed according to the same protocol using primary antibodies against GFRα1 (1:50; MAB560; R&D), STRA8 (1:150; ab-49405; Abcam), KI67 (1:400; 9129; Cell Signaling), and G3G4-anti-BrdU (1:200; AB 2314035; DSHB). For 5-Bromo-2′-deoxyuridine (BrdU) labeling experiment, mice were injected intraperitoneally with 100 mg/kg BrdU (B5002; Sigma) 4 h before they were killed.
Whole-mount immunofluorescence
Seminiferous tubules of 35-day-old mouse testis were removed from tunica albuginea, untangled mechanically and fixed in 4% PFA for 2 h at 4°C. The seminiferous tubules were incubated in 0.3% Triton X-100 in PBS for 30 min and blocked in 10% normal donkey serum in PBS for 1 h at RT. Then, the seminiferous tubules were incubated with primary antibodies against FOXC2 (1:100; AF6989; R&D) and PLZF (1:100; sc-22839; Santa Cruz) in 0.3% Triton X-100 in PBS overnight at 4°C. After washing, the samples were incubated with secondary antibodies of NL557-conjugated donkey anti-sheep IgG (1:200; NL010; R&D) and Alexa Fluor 488-conjugated donkey anti-rabbit IgG (1:200; A-21206; Invitrogen) in 0.3% Triton X-100 in PBS for 2 h at RT. The seminiferous tubules were attached to slide glass and observed using laser scanning microscopy (LSM710; Zeiss).
Cell culture
SSCs cultured in vitro are designated as germ-line stem (GS) cells [7]. Mouse GS cell cultures were established from THY1+-enriched germ cells. Briefly, mouse testes were harvested from 5 to 7 day-postpartum (dpp) C57BL/6♀ × DBA/2♂ F1 (BDF1) or C57BL/6-Tg (CAG-EGFP) 131Osb♀ × DBA/2♂ F1 (BDF1-GFP) pups and tunica albuginea was removed. Single-cell suspension was prepared by digestion with 0.125% Trypsin (25200072; Gibco) for 10 min and 5 μg/mL of DNase I (DN25; Sigma) for 5 min at 37°C. CD90.2 (THY1.2) MicroBeads (130-049-101; Miltenyi Biotec) were used to enrich the THY1+ germ cells by magnetic activated cell sorting (130-042-201; Miltenyi Biotec). THY1+ cells were seeded at a density of 2 × 105 cells/well on 12-well plate containing mitomycin C-inactivated mouse embryonic fibroblasts (MEFs) [29]. Culture medium for the long-term GS cells self-renewal and proliferation was StemPro-34 SFM (10640019; Gibco) supplemented with StemPro-34 Nutrient Supplement (10641025; Gibco), N-2 Supplement (17502048; Gibco), 20 μg/mL insulin (I5500; Sigma), 5 mg/mL bovine serum albumin (A3803; Sigma), 6 mg/mL D-(+)-glucose, 2 mM
Lentivirus infection
Lentivirus-mediated short hairpin RNA (shRNA) was used to permanently knockdown Foxc2 gene expression in GS cells. To create lentivirus particles, pLKO.1-puro vector (Addgene plasmid # 8453) containing Foxc2 shRNA or pLKO.1-scramble shRNA vector (Addgene plasmid # 1864) was co-transfected with packaging vector psPAX2 (Addgene plasmid # 12260) and envelope vector pMD2.G (Addgene plasmid # 12259) into 293FT cells (R70007; Invitrogen) using Lipofectamine 2000 (11668019; Invitrogen) [31,32]. The virus supernatant was concentrated by ultracentrifuge (Optima L-100XP; Beckman). Five Foxc2 shRNAs were designed using BLOCK-iT RNAi Designer (Invitrogen). The shRNA with the highest knock-down efficiency was used in this study and the sequence was
Transplantation
Transplantation procedures were performed as described in [34]. Six-week-old BDF1 mice were injected intraperitoneally with 45 mg/kg busulfan (B2635; Sigma). Four weeks later, endogenous germ cells were depleted from the seminiferous tubules and the mice were used for transplantation (Supplementary Fig. S2A). BDF1-GFP GS cells were collected by trypsin digestion and counted under fluorescence microscope to distinguish them from MEFs. Cells were resuspended at a concentration of 2 × 106 GS cells/mL in culture medium containing 0.4% trypan blue. Approximately 10 μL of cell suspension (2 × 104 GS cells) was injected into the seminiferous tubules of a recipient testis (Supplementary Fig. S2B). GS cells infected with control shRNA lentivirus and GS cells infected with Foxc2 shRNA lentivirus were injected into the left and right testis of the same mouse, respectively. Recipient mouse testes were recovered 2 months after GS cell transplantation and donor GS cells regenerated GFP-labeled germ cells (Supplementary Fig. S2C). To count colonies, recipient testes were analyzed by observing fluorescence under a laser dissecting microscope (Olympus SZX16) [35,36]. Donor germ cells were identified specifically because recipient testis cells had no fluorescence. A cluster of germ cells was defined as a colony when it occupied the entire circumference of the tubule and was at least 0.1 mm long [7,36,37].
Reverse transcription and real-time quantitative PCR
GS cells were separated from MEFs by gentle pipetting and total RNA was extracted from GS cells using RNAiso Plus (9109; Takara) according to the manufacturer's instructions. Reverse transcription was performed using M-MLV Reverse Transcriptase (M1701; Promega). qPCR was performed using FastSYBR Mixture (CW2621 M; CWBIO) in the ABI 7500 Real-Time PCR System (Applied Biosystems). qPCR conditions were 95°C for 5 min, followed by 40 cycles of 95°C for 5 s and 60°C for 34 s. Transcript levels were normalized to endogenous control Gapdh. qPCR primers used in this study are listed in Supplementary Table S1.
Western blot
GS cells were separated from MEFs by gentle pipetting and lysed with RIPA buffer (C1053; Applygen) containing 10 mM DTT (7016; Cell Signaling) and 1 mM PMSF (8553; Cell Signaling). The protein concentration was determined using BCA Protein Assay Kit (P001; Vigorous) according to the manufacturer's instructions. Total protein was separated by 12% SDS-PAGE and transferred onto polyvinylidene difluoride (PVDF) membranes (IPVH00010; Millipore). The membranes were blocked in 5% (w/v) nonfat dry milk for 3 h and incubated with primary antibodies against FOXC2 (1:2,000; AF6989; R&D) or GAPDH (1:5,000; AM4300; Ambion) in 0.1% Tween-20 in tris-buffered saline (TBST) overnight at 4°C. After washing, the membranes were incubated with secondary antibodies of HRP-conjugated donkey anti-sheep IgG (1:5,000; HAF016; R&D) or HRP-conjugated goat anti-mouse IgG (1:10,000; ZB-2305; Zhongshan) in TBST for 2 h at RT. After washing, the membranes were treated with West Pico Chemiluminescent Substrate (34080; Thermo) and photographed using a chemiluminescent imaging system (5200; Tanon). ImageJ Software was used to assay the relative intensity of each blot. The intensity values of each group were normalized to endogenous control GAPDH in the same group.
Extrinsic stimuli treatment
For growth factor treatment, GS cells were cultured without growth factors for 24 h. Then, PBS, GDNF (15 ng/mL) or FGF2 (10 ng/mL) was supplemented for 24 h and samples were collected for reverse transcription and real-time quantitative PCR (RT-qPCR) or western blot analysis [38]. For retinoic acid (RA) treatment, GS cells were treated with DMSO or RA (100 nM) for 24 h and samples were collected for RT-qPCR or western blot analysis [39]. For inhibitor of signaling pathways treatment, GS cells were cultured without growth factors for 24 h. Then, GS cells were pretreated with DMSO, PD98059 (25 μM; 9900; Cell Signaling) or LY294002 (33 μM; 9901; Cell Signaling) for 1 h, and PBS or GDNF (15 ng/mL) was supplemented for 24 h [40]. Samples were collected for RT-qPCR or western blot analysis.
Statistical analysis
Data are presented as means ± SEM. Statistical analysis was performed using SPSS Statistics. Comparisons between two groups were analyzed by Student's t-test. A value of P < 0.05 was considered to be statistically significant.
Results
The expression of FOXC2 in testis is restricted to GFRα1-positive spermatogonia and is mainly in As and Apr spermatogonia
We examined the expression of FOXC2 in testes of 1-, 4-, 7-, 14-, 21-, and 35-day (d)-old mice. In 1-d-old testis, FOXC2 was expressed in the gonocytes in the luminal compartment of seminiferous tubules. Gonocytes resume proliferation around postnatal day (P) 3, migrate to the seminiferous tubule basement membrane and differentiate into SSCs [2]. In 4-, 7-, 14-, 21-, and 35-d-old testis, FOXC2 was expressed in the spermatogonia on the basal membrane of seminiferous tubules. To detect FOXC2 expressing spermatogonial cell types, double immunofluorescence was performed. The expression of PLZF is restricted to undifferentiated spermatogonia [2]. We found that FOXC2 was expressed in a subpopulation of PLZF-expressing-undifferentiated spermatogonia (Fig. 1A and Supplementary Fig. S3). FOXC2 was expressed in 90.17%, 78.64%, 51.56%, 41.77%, 29.97%, and 26.62% of undifferentiated spermatogonia in 1-, 4-, 7-, 14-, 21, and 35-d-old testes, respectively (Fig. 1B). With the development of testis, the proportion of FOXC2-expressing spermatogonia decreased in the undifferentiated spermatogonia. GFRα1 is expressed in large subsets of As and Apr spermatogonia and in small subsets of Aal spermatogonia [11]. STRA8 is expressed in differentiating spermatogonia and spermatocytes, and its expression begins shortly before spermatogonial differentiation [41]. Interestingly, we found that FOXC2 was expressed in the same cells with GFRα1 and in the different cells with STRA8 (Fig. 1C, D and Supplementary Fig. S4). These results confirm the undifferentiated state of FOXC2-expressing spermatogonia.
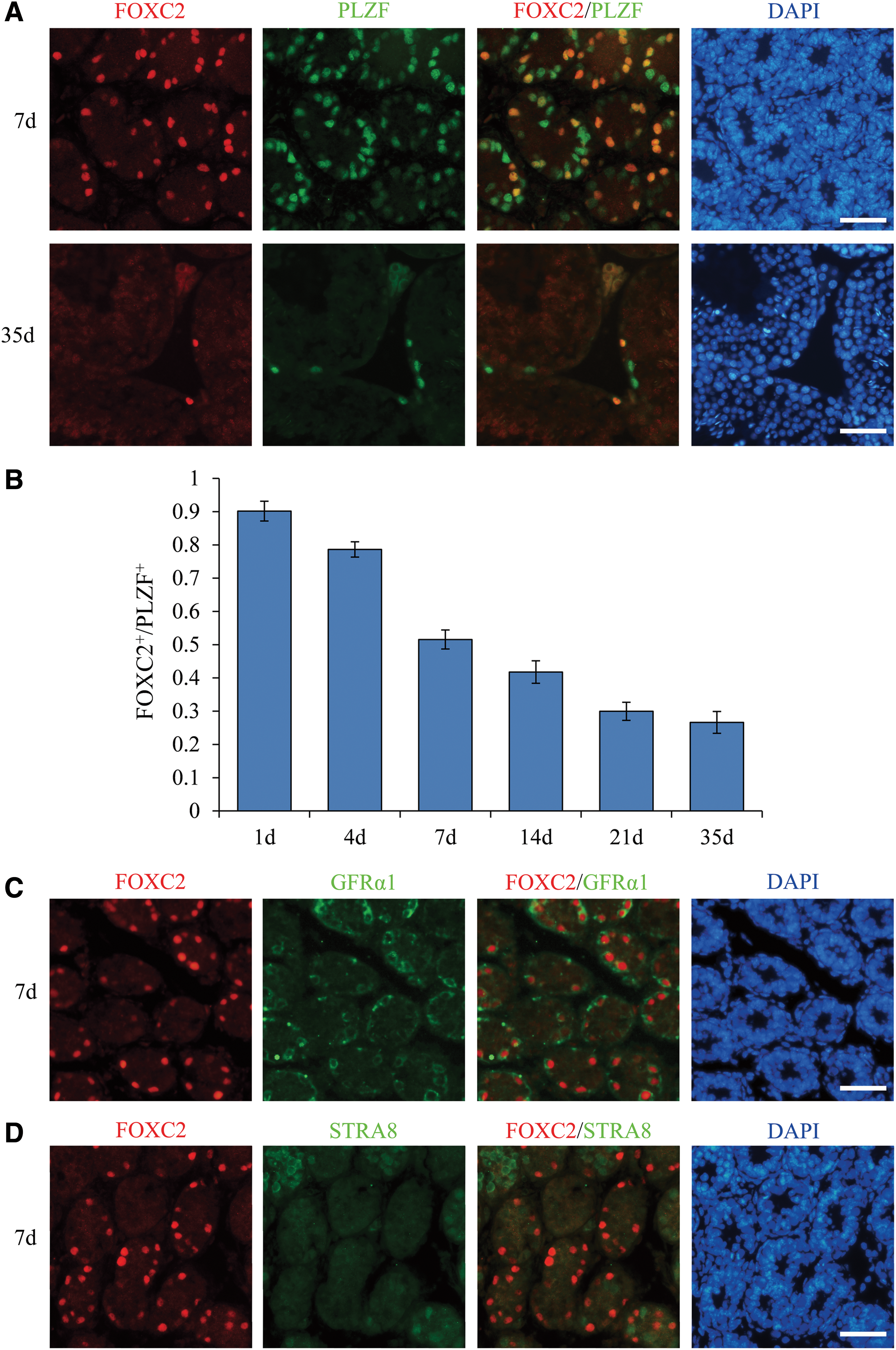
Expression of FOXC2 and PLZF/GFRα1/STRA8 within the seminiferous epithelium of testes from postnatal mice.
To detect the proliferation status of FOXC2-expressing spermatogonia, double immunofluorescence of FOXC2 and KI67 was performed. However, no obvious relationship was found between FOXC2 and KI67. Some of the FOXC2-expressing spermatogonia were KI67 positive, but others were KI67 negative (Supplementary Fig. S5A). Further, we performed BrdU labeling experiments to label cells in the S phase. Double immunofluorescence of FOXC2 and BrdU showed that most of the FOXC2-expressing spermatogonia were BrdU negative (Supplementary Fig. S5B). These results suggest that FOXC2 may not affect the proliferation of SSCs.
Undifferentiated spermatogonia are classified as Asingle (As), Apaired (Apr), and Aaligned (Aal) according to the number of cells in the clones. Whole-mount double immunofluorescent staining of seminiferous tubules was performed to investigate the expression of FOXC2 in undifferentiated spermatogonia. We found that FOXC2 was expressed preferentially in As and Apr spermatogonia (Fig. 2A). A small amount of Aal-4 spermatogonia also expressed FOXC2 (Fig. 2A). FOXC2 was very rarely expressed in Aal-8 spermatogonia and the expression level was much lower than those of As and Apr spermatogonia (Fig. 2B). In Aal-16 spermatogonia, FOXC2 expression was not detected (Fig. 2C). FOXC2 was expressed in 85.41% of As spermatogonia, 68.48% of Apr spermatogonia, 22.11% of Aal-4 spermatogonia, 11.67% of Aal-8 spermatogonia, and 0% Aal-16 spermatogonia (Fig. 2D). These results suggest that FOXC2-expressing spermatogonia are even further undifferentiated than other undifferentiated spermatogonia.

Expression of FOXC2 and PLZF within the seminiferous tubules of testes from 35-d-old mice.
Reduction of the Foxc2 transcript level affects SSC clump maintenance in vitro and decreases SSC colonization
SSC culture provides an excellent method for studying SSC function in vitro. These cells are designated as germ-line stem (GS) cells [7]. To investigate the function of FOXC2, we used GS cells from BDF1 and BDF1-GFP mice. Before beginning functional studies, immunocytochemistry was performed to obtain the makeup of the germ cells present in BDF1 GS cells [42]. Immunocytochemistry staining of the GS cells showed that all GS cells expressed PLZF while only a portion of them expressed FOXC2, which was consistent with the testis immunofluorescence and whole-mount results (Fig. 3A).
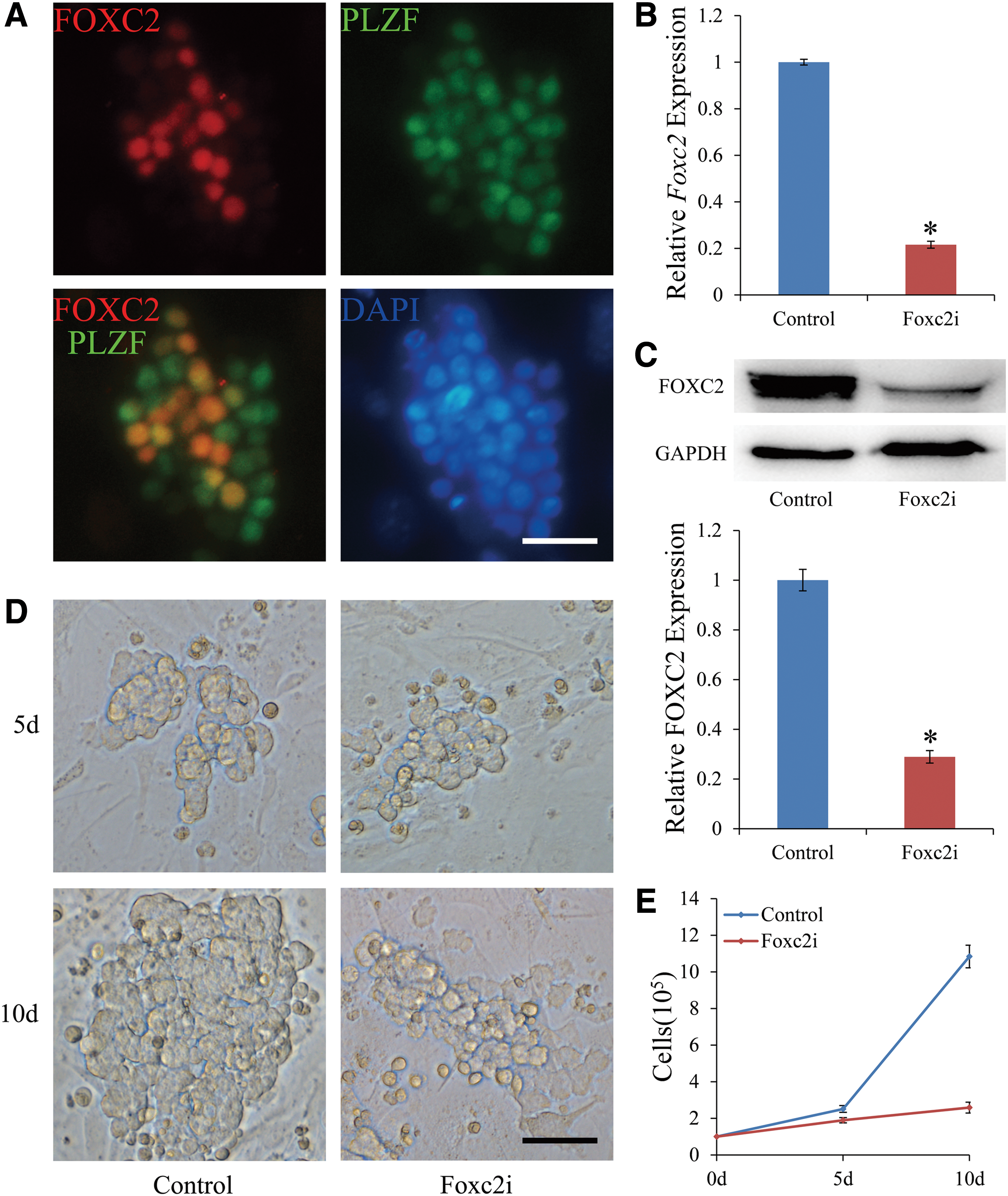
Expression of FOXC2 and PLZF in the GS cells and the evaluation of FOXC2 function in GS cell maintenance in vitro.
To demonstrate its biological significance, we examined the effects of FOXC2 on SSC maintenance in vitro. BDF1-GFP GS cells allow for readily distinguishable GS cells by means of GFP, which allows them to be easily distinguished from MEFs when doing cell counts. To suppress the mRNA level of Foxc2, cultured GS cells were stably transduced by lentivirus containing a Foxc2 shRNA transgene. The mRNA level of Foxc2 in cultures treated with Foxc2 shRNA lentivirus (designated as Foxc2i) was significantly reduced to only 21.58% of that in control cultures treated with nontargeting shRNA lentivirus (designated as control) as measured by qRT-PCR after 72 h (Fig. 3B).The FOXC2 protein level of Foxc2i was reduced to 28.93% compared with control, similarly (Fig. 3C). Next, puromycin (1 μg/mL) was supplemented for approximately 48 h to select cells that had stably incorporated the transgene. At 10 days after transduction with the Foxc2 shRNA lentivirus, there was a significant reduction in both the clump size and number of GS cells compared with cells transduced with the negative control shRNA lentivirus. As a result, viable GS cells in Foxc2 shRNA-treated cultures proliferated much more slowly compared with negative control shRNA-treated cultures (Fig. 3D). The number of GS cells in the negative control shRNA-treated cultures and Foxc2 shRNA-treated cultures was 2.51 × 105 and 1.89 × 105 at 5 days after transduction, and 10.84 × 105 and 2.59 × 105 at 10 days after transduction, respectively (Fig. 3E). These data demonstrate that inhibiting of Foxc2 has a significant impact on SSC maintenance in vitro and suggest that FOXC2 is an important transcription factor for SSC self-renewal and survival.
To determine the effects that Foxc2 knock-down has on the SSC population, we used BDF1-GFP GS cells and performed transplantation experiments. Foxc2 shRNA-treated GS cells and control shRNA-treated GS cells were selected by puromycin (1 μg/mL) for 48 h and were transplanted into the testes of busulfan-treated recipient BDF1 mice (approximately 2 × 104 GS cells per testis). As previously reported, this approach enables quantification of SSCs in vivo and allows us to determine the reduction in the number of SSCs that occur in the GS cell cultures following knock-down of Foxc2. We performed six transplantation experiments and four busulfan-treated mice were used in each experiment. Approximately 2 months following transplantation, the mice were killed and the testes were observed using a laser dissecting microscope to determine the number of donor-cell-derived colonies. Knock-down of Foxc2 within GS cell cultures significantly decreased the formation of donor-cell-derived colonies (Fig. 4A). The control shRNA-treated GS cells formed 103.96 colonies per 105 cells while the Foxc2 shRNA-treated GS cells formed only 10.83 colonies per 105 cells, about 10.42% of the control (Fig. 4B and Table 1). The findings presented here further validate the importance of FOXC2 in maintaining the SSC population and demonstrate that the loss of FOXC2 reduces the number of SSCs in GS cell cultures in vitro.
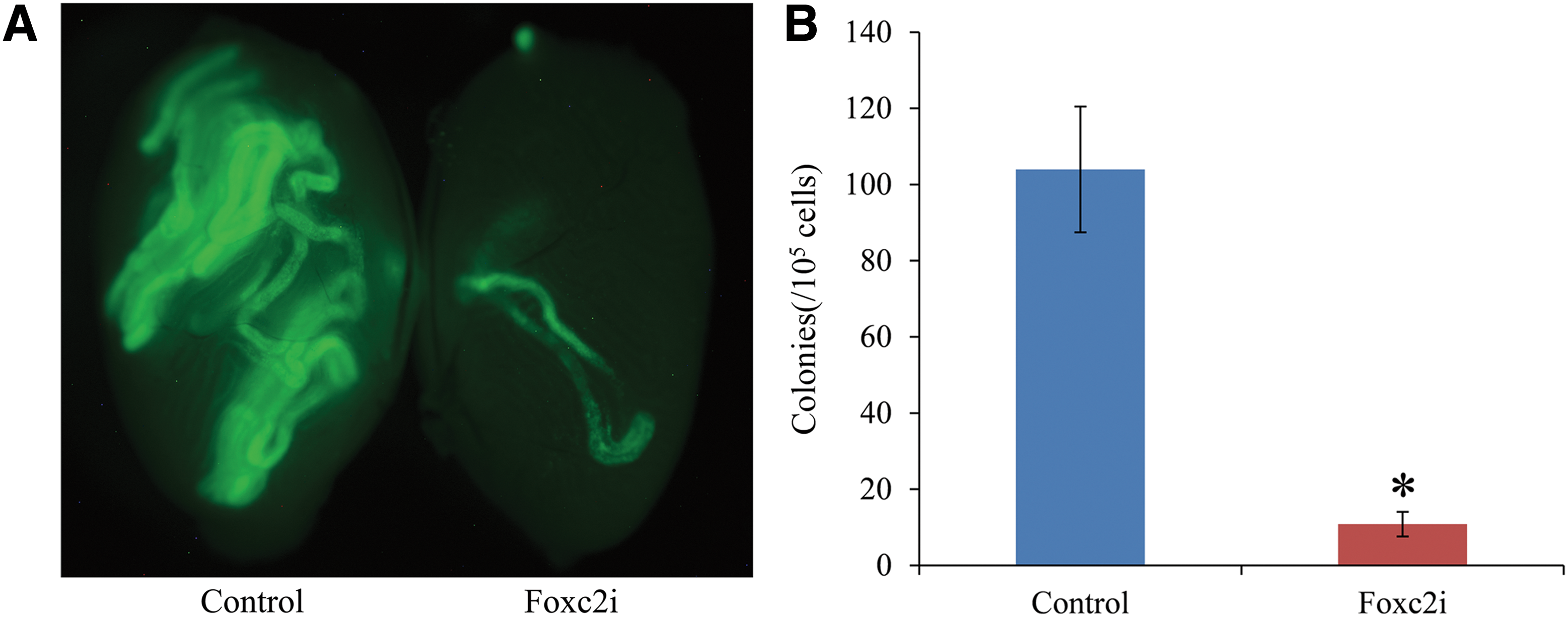
Colonization of recipient testes by GS cells.
Further, we did the overexpression experiments using lentivirus. FOXC2 was overexpressed in both mRNA and protein level in GS cells (Supplementary Fig. S6A, B). However, the number of Foxc2-OE GS cells was almost the same compared to control-OE at 15 days after infection (Supplementary Fig. S6C, D). The colonization ability of Foxc2-OE GS cells slightly improved, but the statistical analysis showed no significant difference (Supplementary Fig. S6E, F).
Gene expression changes following knock-down of Foxc2
Knock-down of Foxc2 impaired SSC maintenance and reduced SSC colonization. This may be caused by a decrease in self-renewal, a decline in homing ability, differentiation of SSCs, or a combination of all three. Since a number of factors, including Plzf, Etv5, c-Ret, Gfrα1, Bcl6b, Pin1, Oct4, Foxo1, Atm, Csf1r, Hras, Fbxw7, Itgb1, Cdh1, Cxcr4, Rac1, Ngn3, Nanos2, Nanos3, Id4, Dazl, Dmc1, Sycp3, Sohlh1, Sohlh2, Sox3, Rarg, Stat3, Dmrt1, Sall4, Stra8, and c-Kit [2,27,30,38,41,43 –68], have been reported to be involved in regulating SSC self-renewal, homing, or differentiation, we examined the effects of Foxc2 knock-down on the expression of these genes in GS cell cultures. The reason for the small degree of change of the indicated genes is that the GS cells in the culture were heterogeneous and only a small portion of them expressed Foxc2. RT-qPCR results showed that there were no significant changes in the genes associated with SSC self-renewal (Fig. 5A) and homing (Fig. 5B). However, the levels of Stra8 and c-Kit that are involved in SSC differentiation increased, especially Stra8 (Fig. 5C). Stra8 and its inducer, RA, have been reported to promote spermatogonia differentiation [41]. c-Kit is also known as a spermatogonia differentiation marker [68]. Of the genes whose levels were reduced, Nanos2 decreased most (Fig. 5C). Nanos2 has been reported to act downstream of GDNF signaling to suppress differentiation of SSCs [56]. These results indicate that Foxc2 may play an important role in suppressing SSC differentiation. Stra8 had the biggest change degree and may have a close relationship with Foxc2.
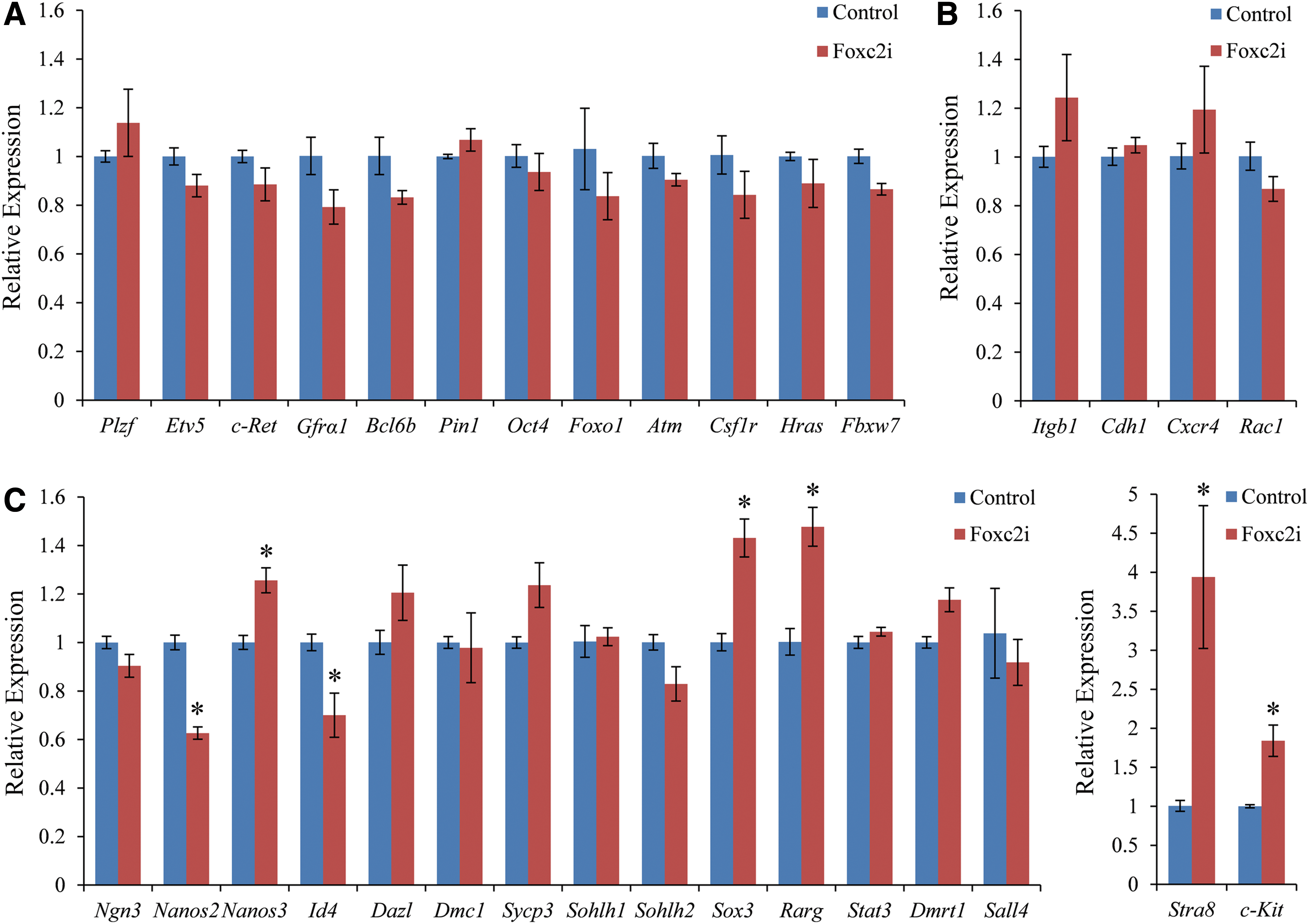
RT-qPCR analysis of the indicated genes following knock-down of Foxc2.
FOXC2 is upregulated by GDNF and downregulated by RA
GDNF and basic fibroblast growth factor (FGF2) are critical regulators of SSC self-renewal and survival. In particular, GS cells do not survive and proliferate in vitro without GDNF [69]. GS cells were cultured without cytokines for 24 h, and samples were collected 24 h after GDNF or FGF2 supplementation. RT-qPCR and western blot results showed that FOXC2 was significantly upregulated by GDNF and slightly upregulated by FGF2 (Fig. 6A, C). It is also known that RA and its target gene Stra8 regulate the initiation of spermatogonia differentiation in mice [41]. We therefore treated cultured GS cells with 100 nM RA for 24 h. RT-qPCR and western blot results showed that FOXC2 was downregulated by RA (Fig. 6B, D).
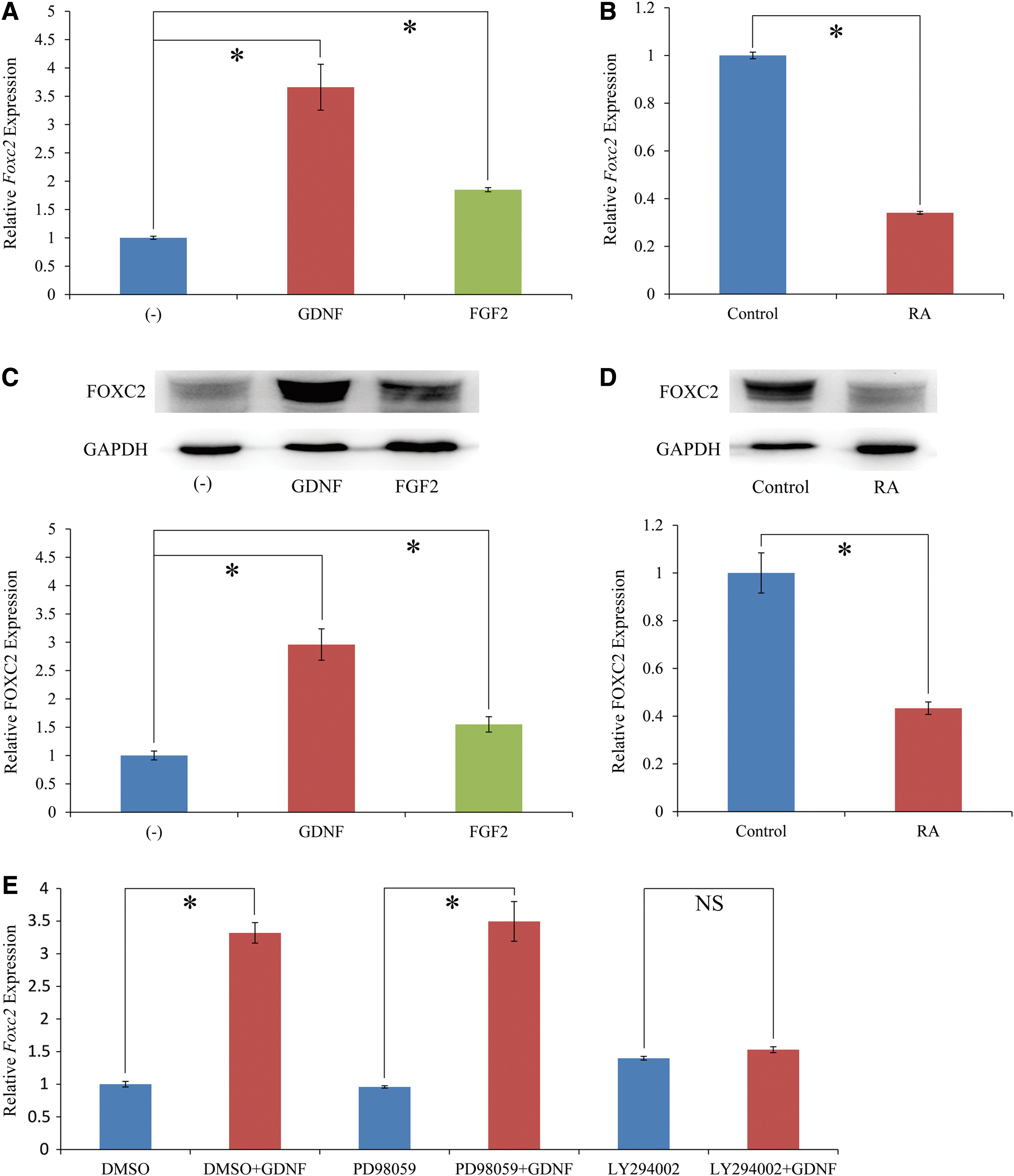
Regulation of FOXC2 expression in GS cells by extrinsic stimuli.
To identify the intracellular signaling pathway of GDNF that regulates Foxc2 expression in GS cells, GS cells cultured without cytokines for 24 h were pretreated with DMSO, PD98059 (MEK inhibitor), or LY294002 (PI3K inhibitor) for 1 h, followed by treatment with PBS or GDNF for 24 h. The Foxc2 mRNA levels were detected by RT-qPCR and the results showed that upregulation of Foxc2 by GDNF stimulation was blocked by LY294002 (Fig. 6E). These results suggest that GDNF upregulates Foxc2 expression through PI3K-Akt pathway.
Discussion
The prevailing SSC model for many years is the “As model” [3,8,9]. This model assumes that the stem cell population resides in the As spermatogonia, whereas Apr and Aal do not have the stem cell function of self-renewal and are irreversibly committed to differentiation. However, this theory was challenged. Nakagawa et al. showed that Ngn3+ spermatogonia could revert to SSCs [13]. When Ngn3/CreER™; CAG-CAT-Z mice are injected with tamoxifen for lineage tracing most of the labeled colonies disappear within 3 months. However, a small fraction of them still exist and are maintained for 14 months, which suggests that Ngn3-expressing cells revert to SSCs. Similarly, Barroca et al. showed that KIT-expressing spermatogonia appear to retain some stem cell potential [70]. Several genes were found to be heterogeneously expressed in undifferentiated spermatogonia. GFRα1 and NANOS2 are expressed in large subsets of As and Apr and in small subsets of Aal [10,11]. Conversely, NGN3 and NANOS3 are expressed mainly in Aal while rarely in As and Apr [10,11]. OCT4 is heterogeneously expressed in GS cells, but in adult mice, OCT4 is also present in Sertoli cells [48,71]. Even the As spermatogonia is not a homogeneous stem cell population and it can be classified by gene expression. GFRα1+ spermatogonia are shown to function as stem cells that possess the ability to self-renew in steady state and can therefore be referred to as “actual stem cells.” NGN3+ spermatogonia exit from the stem cell compartment and tend to differentiate. This population possesses the potential to self-renew but does not normally do so in steady-state spermatogenesis. During regeneration after damage, NGN3+ cells reverse to GFRα1+ and restore their ability to self-renew. This functional compartment is referred to as “potential stem cells” [13 –15].
In our research, we found that FOXC2 is expressed mainly in As and Apr spermatogonia. A small amount of Aal-4 spermatogonia also express FOXC2. FOXC2 is very rarely expressed in Aal-8 spermatogonia and the expression level is much lower than those of As and Apr spermatogonia. In Aal-16 spermatogonia, FOXC2 expression was not detected. Additionally, the proportion of FOXC2-expressing spermatogonia decreases in the undifferentiated spermatogonia with testicular postnatal development. When gonocytes convert to SSCs around postnatal day (P) 3, most germ cells are SSCs. With testicular development and SSC differentiation, the proportion of SSCs decreases to 0.02%–0.03% of all germ cells in adult testis [4]. The decreasing proportion of FOXC2-expressing spermatogonia is consistent with this phenomenon. In GS cells cultured in vitro, FOXC2 is expressed in only some of the GS cells. The expression pattern of FOXC2 is the same as that of GFRα1, so it can also be considered as a marker for actual stem cells in testis. To our knowledge, FOXC2 is the first transcription factor that was found expressing in this pattern and is therefore thought to play an important role in SSCs.
To demonstrate the functional importance of FOXC2 on SSC survival and self-renewal, further analysis was done. Using lentivirus-mediated reduction of Foxc2 transcript levels in self-renewing GS cells, we found a dramatic effect on SSC maintenance in vitro. Because of the heterogeneous nature of cultured GS cells, treatments in vitro must be coupled with transplantation assays to unequivocally draw conclusions about effects on SSCs since the capability to reestablish and support spermatogenesis defines SSCs. The transplantation results show that there are fewer SSCs in the Foxc2 shRNA-treated GS cells than those in the negative control shRNA-treated GS cells. These results suggest that FOXC2 plays an important role in maintaining the SSC population.
To indicate the mechanism by which Foxc2 affects GS cell biology, we analyzed genes associated with SSC self-renewal, homing and differentiation. The RT-qPCR results show that Foxc2 may play an important role in suppressing SSC differentiation and may have a close relationship with Stra8. An important feature of GS cells in culture is that there is a considerable level of cell death [48]. The main reason is that the current conditions cannot support the viability of differentiating spermatogonia. Therefore, the impaired SSC maintentance in vitro following knock-down of Foxc2 may be caused by increased differentiation of SSCs and these differentiating spermatogonia die due to current culture conditions.
The results of withdrawal and supplementation of the growth factors show that FOXC2 levels are significantly upregulated by GDNF and slightly upregulated by FGF2. GDNF has been established as the primary regulator of SSC cell fate. Undifferentiated spermatogonia accumulate in the seminiferous tubules of mice overexpressing GDNF, whereas spermatogenesis gradually declines in the heterozygous knockout (KO) mice, which leads to male infertility [72]. GFRα1 is the membrane receptor of GDNF and FOXC2 is expressed in the same cells with GFRα1. The dramatic alteration in FOXC2 expression levels resulting from withdrawal and supplementation of these growth factors indicates that FOXC2 acts as an important molecule in SSCs. The signaling pathway experiments show that GDNF signals via the GFRα1/RET co-receptor through activation of PI3K-Akt pathway and subsequently induces several target genes in SSCs, with Foxc2 being one of them [40,73 –75]. RA is known to trigger the differentiation of spermatogonia [76]. Vitamin A-deficient mice and rats are infertile because the differentiation of spermatogonia is inhibited, and treating them with either vitamin A or RA results in the recovery of spermatogonia differentiation [77 –79]. FOXC2 is also downregulated by RA.
In conclusion, our results can be interpreted as schematic depicted in Fig. 7. SSCs are controlled by intrinsic molecular pathways that can be activated by extrinsic signals such as growth factors. FOXC2 is a transcription factor that plays a critical role in the survival and fate decision of SSCs. FOXC2 is predominately expressed in As and Apr spermatogonia to safeguard the delicate balance between actual and potential stem cells. When FOXC2 expression level is high, SSCs possess the stem cell properties and self-renew to maintain their population. When FOXC2 is not expressed or the expression level is very low, SSCs differentiate to produce daughter cells that go through several steps of mitosis and meiosis and eventually become spermatozoa. GDNF and RA, locally produced by Sertoli cells of testis, act in opposite ways to regulate FOXC2 expression and SSC property in postnatal germ cells. GDNF upregulates FOXC2 expression and promotes SSCs to self-renew, while RA downregulates FOXC2 expression and induces SSCs to differentiate.

Model of FOXC2 function in SSC fate decision. When FOXC2 expression level is high, SSCs act as actual stem cells and self-renew to maintain their population. When FOXC2 is not expressed or the expression level is very low, SSCs act as potential stem cells and differentiate to produce daughter cells in normal steady-state spermatogenesis. GDNF and RA locally produced by testis Sertoli cells act in opposite ways to regulate FOXC2 expression and SSC property in postnatal germ cells. GDNF upregulates FOXC2 expression and promotes SSCs to self-renew, while RA downregulates FOXC2 expression and induces SSCs to differentiate. Color images available online at
Footnotes
Acknowledgments
This work was supported by the National Basic Research Program of China (2014CB138505) and the Natural Science Foundation of China (31172287 and 31372393).
Author Disclosure Statement
The authors indicate no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.