
Abstract
Mesenchymal stem cells (MSCs) have a superior immunomodulatory capacity compared to other cells of the immune system, and they hold great promise for treating various immune disorders. However, their regulatory effects on the maturation of immature dendritic cells (imDCs) are not fully understood. In this study, we show that Sca-1+Lin−CD117−MSCs restrain the lipopolysaccharide-stimulated maturation transition of imDCs cocultured without exogenous cytokines. The Notch signaling pathway plays a critical role in the process by controlling interferon regulatory factor 8 (IRF8) expression in an RBP-J-dependent manner. We observed a high degree of H3K27me3 modification mediated by SUZ12 and a relatively low degree of H3K4me3 modification regulated by WDR5 at the IRF8 promoter during coculture. These data reveal a possible mechanism by which Sca-1+Lin−CD117−MSCs modulate imDC maturation and further support the role of MSCs in treating immune disorders.
Introduction
M
However, the detailed mechanisms responsible for their immunoregulatory effects on DCs remain unclear. Untangling the complicated interactions between MSCs and DCs is critical for understanding the diverse mechanisms involved in the pathogenesis of immune-related diseases and for discovering new therapeutic targets for advanced treatment.
DCs, which form a heterogeneous group of functionally specialized antigen-presenting cells (APCs), direct the type and specificity of immune responses [8]. DCs have been reported to exist in distinct activation states, often termed “immature” and “mature.” “Maturation” is used to denote DC differentiation in response to environmental stimuli, which allows DCs to adapt the type of immunity to the challenge at hand [9]. Toll-like receptors (TLRs) are pattern recognition receptors located on the surface of DCs that recognize pathogens to activate innate immunity and regulate the transition from innate to acquired immune responses. DCs mature during this process.
Mature DCs express high levels of the surface molecule MHC II and the costimulatory molecules CD80 and CD86 and secrete higher levels of cytokines such as interleukin (IL)-12. This transition is indispensable for triggering an immune response because immature DCs (imDCs) not only fail to effectively prime T cells but also serve to promote tolerance induction [10 –12]. Therefore, the precise regulation of TLR signaling pathways is important for controlling the inflammatory response. Until now, various intracellular signaling molecules have been shown to be involved in the regulation of the TLR pathway to maintain the immunological balance. However, whether signals via cell membrane receptors are involved in cross-talk with TLR signaling to regulate the inflammatory response needs to be clarified [13,14].
Notch is an evolutionarily conserved transmembrane protein that plays a key role in the development of a variety of immune cells. Four Notch receptors (Notch1-4) and five ligands (Jagged1 and 2, Delta 1, 3, and 4) have been identified. Notch signaling is initiated through the binding of Notch ligands. After ligand activation, the Notch intracellular domain (NICD) is released and translocates to the nucleus. Notch signaling is classified into canonical and noncanonical pathways based on whether NICD interacts with a CBF1/RBP-J, Su(H), or Lag-1 (CSL) transcription factor [15].
In mice, CSL is known as a recombination signal binding protein for the immunoglobulin kappa J region (RBP-J) and functions as a transcriptional repressor. Canonical Notch signaling involves NICD binding to RBP-J and converting it from a repressor to an activator, resulting in the transcription of Notch-dependent genes that influence developmental and differentiation systems. Evidence of NICD binding to RBP-J to maintain a repressor status has been recently reported and involves the dislocation and recruitment of coactivators and corepressors, respectively [16,17]. Recent evidence suggests that there is cross-talk between Notch and TLR signaling pathways; however, the detailed relationship and underlying mechanisms between Notch and TLR signaling are far from being understood.
Specific gene expression patterns are controlled by epigenetic alterations, and modifications to histone proteins on chromatin play crucial roles in the development and differentiation of various cell types [18 –20]. Increased acetylation (ac) of histone tails at promoters is typically associated with transcriptional activation, and the functional consequences of methylation (me) depend on the number of methyl groups, the residue itself, and its location within the histone tail. For example, H3K4me3, which is catalyzed by the Trithorax family of proteins, is correlated with an open chromatin structure and active gene expression, whereas H3K27me3 catalyzed by polycomb repressive complex 2 (PRC2) is associated with an inactive chromatin structure and gene repression [21,22]. Based on accumulating evidence, epigenetic alterations are involved in regulating the expression of various genes, but little is known about their diversification during the imDC maturation transition [23 –25].
Although MSCs have been shown to adjust the development and function of DCs both in vitro and in vivo [26 –28], researchers have not yet determined whether they regulate the fate of imDCs and the underlying mechanisms. In this study, mouse BM-derived Sca-1+Lin−CD117−MSCs influenced the maturation transition of imDCs by suppressing TLR4-stimulated interferon regulatory factor 8 (IRF8) expression via the Notch-RBP-J signaling pathway, revealing that a hierarchical molecular network leads to the maturation transition blockade. Our results have led us to hypothesize that the modulation of imDC maturation by MSCs could be used as a potential therapeutic approach for some pathological conditions.
Materials and Methods
Mice
Five- to six-week-old BALB/C and C57BL/6 mice were purchased from the Laboratory Animal Center of Chinese Academy of Medical Sciences (Beijing, China). All mice were bred and maintained under specific pathogen-free conditions. Animal use and experimental procedures were approved by the Animal Care and Use Committee of the Chinese Academy of Medical Sciences.
Culture of mouse BM-derived MSCs
MSCs were prepared from mouse BM cells as previously described [29], with minor modifications. In brief, the tibia and femora were isolated and crushed using a mortar and pestle. The crushed bones were then chopped and incubated for 1.5–2 h at 37°C in 15 mL of phosphate-buffered saline containing 0.2% (wt/vol) collagenase type 1 (Sigma-Aldrich) and 20% fetal bovine serum (FBS). The cell suspension was filtered through a 100-μm cell strainer placed in a 50-mL conical tube and then centrifuged twice. The isolated cells were then cultured in MSC basal medium (StemCell) supplemented with 10% MSC stimulatory supplements (StemCell) at 37°C with 5% CO2.
The nonadherent cells were removed after 48 h, and the adherent cells were harvested by trypsinization when they reached 90% confluence and were designated BM-MSCs. Scal+CD117− cells were sorted using an EasySep CD117-Positive Selection Kit for CD117− cells and an EasySep Sca-1-Positive Selection Kit (StemCell) for Sca-1+ cells. The purity of Sca-1+CD117−Lin− cells was confirmed by flow cytometry.
Preparation of imDCs and mature DC from BM
Mouse BM-derived DCs were generated from BM progenitors according to an established protocol [30], with minor modifications. BM mononuclear cells were prepared from BALB/C mouse femur BM suspensions by depleting red cells and then culturing at a density of 2 × 106 cells/mL in six-well plates in RPMI-1640 medium supplemented with 10% fetal calf serum, 10 ng/mL granulocyte-macrophage colony-stimulating factor and 5 ng/mL IL-4 (R&D Systems). Nonadherent cells were gently washed out on day 6, and the remaining loosely adherent cell clusters were collected as imDCs. The imDCs cultured for one additional day under the stimulation of 1 μg/mL lipopolysaccharide (LPS; Sigma-Aldrich) were used as maDCs.
Coculture experiment
The ratio of MSCs/imDCs was 1:10. After 1 day of culture, MSC-DCs [imDCs were termed MSC-DCs after coculturing with BM-MSCs in the presence of LPS (1 μg/mL) for 1 day] were harvested. In some cases, the experiments were divided into two groups: MSC-DCs treated with dimethyl sulfoxide (DMSO) and MSC-DCs plus the Notch inhibitor N-[N-(3,5-difluorophenacetyl)-
Flow cytometry analysis
Flow cytometry analyses were performed using a previously described method [29]. The fluorescent antibodies (Abs) used in the study included fluorescein isothiocyanate-conjugated anti-mouse Sca-1, CD9, CD90, CD31, CD44, CD45, CD11c, CD80, CD86, CD40, and phycoerythrin-conjugated anti-mouse CD73, CD11b, and Ia (BD Biosciences). For each Ab, immuonoglobulin G (IgG) of the same isotype from the same species was used as an isotype control (BD Biosciences). Analyses were performed using Accuri C6 flow cytometers and CFlow software (Accuri Cytometers, Ann Arbor, MI). Detailed information on the antibodies used is presented in Supplementary Table S1 (Supplementary Data are available online at
Enzyme-linked immunosorbent assay for cytokines
The supernatant of the MSC-DCs cultured in RPMI 1640 medium without FBS was harvested after 4 h. The concentrations of IL-10, IL-12, and tumor necrosis factor-α (TNF-α) in the supernatant were determined using ELISA kits (BD Biosciences) according to the manufacturer's instructions.
Analysis of DC phagocytic ability
An endocytosis assay was performed as previously described [29].
Western blot analysis
In brief, equal quantities of extract (30 μg of total protein) were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and the proteins were transferred to polyvinylidene difluoride membranes. After blocking, the membranes were sequentially incubated with primary and secondary Abs diluted in 3% bovine serum albumin. Horseradish peroxidase-labeled IgG was used as the secondary Ab for detection. Abs were obtained from Cell Signaling Technology. Detailed information on the antibodies used is presented in Supplementary Table S1.
Lentiviral vector preparation and infection and small interfering RNA knockdown assay
Lentiviruses were produced according to the manufacturer's instructions from GenePharma (
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) was performed using an EZ-Magna ChIP™ Kit (Millipore) according to the manufacturer's instructions. ChIP-grade Abs against H3K4me3, H3K27me3, and EED were obtained from Millipore; Abs against SIRT1 and IRF8 were obtained from Cell Signaling Technology; anti-WDR5 and anti-SUZ12 Abs were obtained from Abcam, and anti-ASH2, anti-RbBP5, and anti-MLL1 Abs were obtained from Bethyl Laboratories.
The promoters were selected from eukaryotic promoter database (
Coimmunoprecipitation
Cells were harvested and lysed in RIPA buffer. The lysate was incubated with protein A/G agarose beads (Millipore); the RBP-J or IRF8 Ab (10 μg) was then added, and the resulting mixture was incubated overnight at 4°C. Beads that were conjugated with the lysate, and Abs were precipitated, washed three times with RIPA buffer, and then analyzed by western blotting. Detailed information on the antibodies used is presented in Supplementary Table S1.
Statistical analyses
All experiments were performed at least three times. All statistical analyses were performed using SPSS software version 17.0. The P-values were calculated using Student's t test, and P-values <0.05 were considered statistically significant.
Results
Sca-1+CD117−Lin− MSCs prevent the TLR4-induced maturation of imDCs
We isolated Sca-1+CD117−Lin− MSCs from BM (Supplementary Fig. S1), seeded imDCs on MSC monolayers at a ratio of 10:1 (imDCs:MSCs), and then treated the cells with 1 μg/mL LPS to investigate the mechanism by which Sca-1+CD117−Lin− MSCs regulate imDC maturation. In contrast to the maturation of imDCs that exhibited longer and a greater number of dendrites, which are important morphological markers of maDCs, imDCs cocultured with Sca-1+CD117−Lin− MSCs gradually became larger, but produced fewer and shorter dendrites (Fig. 1a). In addition, according to the phenotype analysis (Fig. 1b and Supplementary Fig. S2), these DCs controlled by Sca-1+CD117−Lin− MSCs, which were termed MSC-DCs, expressed lower levels of the myeloid lineage marker CD11b and the functional markers Ia, CD80, CD86, and CD40 than maDCs.
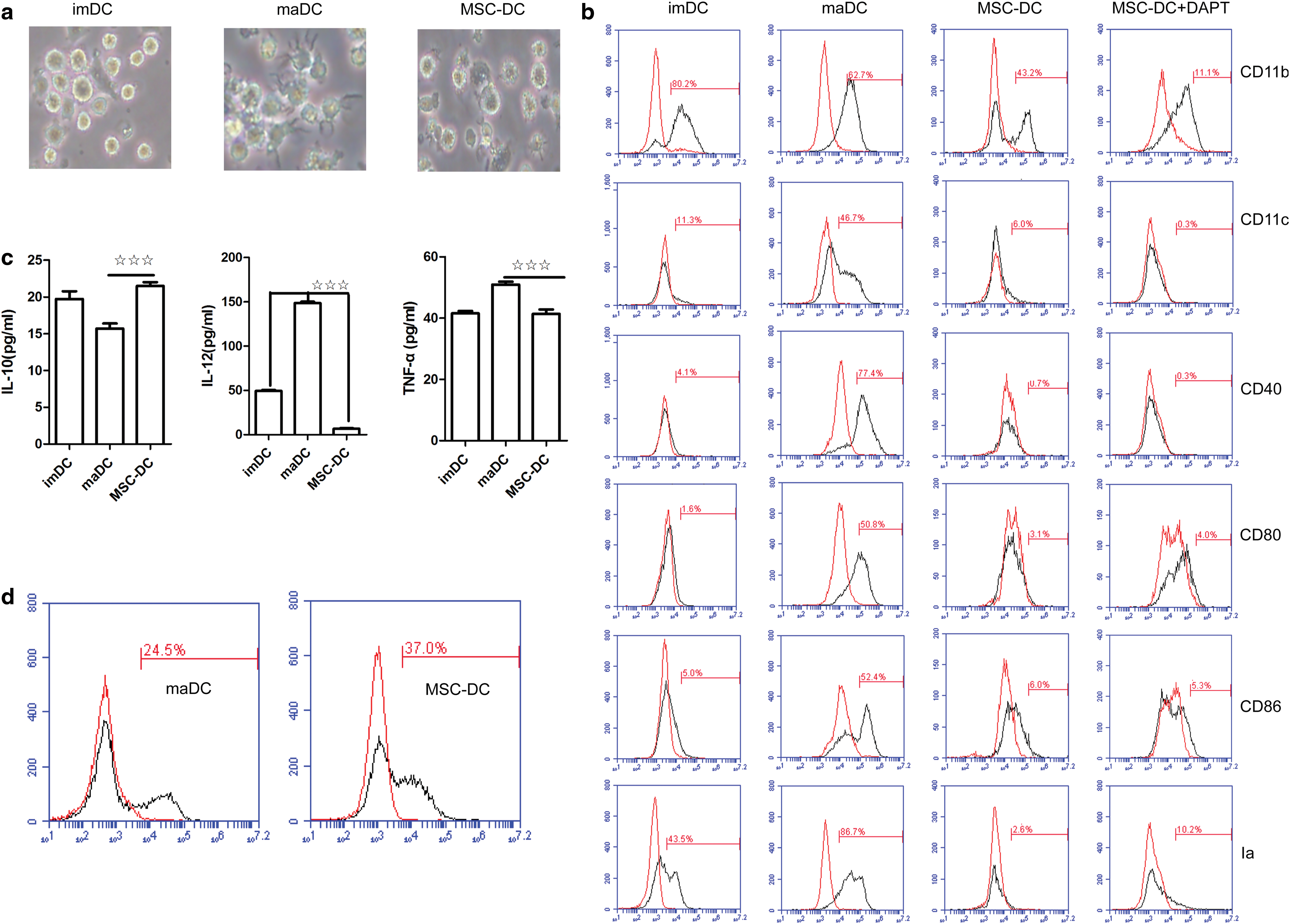
Sca-1+CD117−Lin− MSCs regulate TLR4-induced imDC maturation.
Secretion of inflammatory factors, such as IL-12, is a vital function of maDCs; therefore, we further determined their cytokine expression patterns by enzyme linked immunosorbent assay (Fig. 1c). In contrast to maDCs, MSC-DCs spontaneously secreted more IL-10 but less IL-12 and TNF-α, suggesting that MSC-DCs may be involved in immune regulation. Moreover, MSC-DCs exhibited a greater phagocytic capacity (Fig. 1d and Supplementary Fig. S2). These results (including: morphology, phenotype, levels of cytokine secretion, and expression patterns of characteristic transcription factors) suggest that mouse BM-derived Sca-1+CD117−Lin− MSCs block the maturation of imDCs and induce imDCs to differentiate into a unique population of MSC-DCs.
TLR4-triggered IRF8 expression and inflammatory cytokine production are suppressed by Notch signaling
Notch signaling has been shown to play an important role in the inflammatory response. We sought to determine whether the Notch signaling pathway influences the maturation of imDCs cocultured with Sca-1+CD117−Lin− MSCs. Reduced IL-10 and increased IL-12 and TNF-α secretion by MSC-DCs were observed when DAPT (a Notch inhibitor) was added to the coculture (Fig. 2a). As shown in the western blot analysis, the MSC-DCs expressed lower levels of PU.1, TRAF6, IRF8, Batf3, Id2, and Ikaros but similar levels of IRF4, RBP-J, RelB, and TCF4 compared with maDCs. However, after DAPT was added, these cells expressed more PU.1, TRAF6, IRF4, IRF8, RelB, Batf3, and Id2 (Fig. 2b and Supplementary Fig. S3), indicating that Notch signaling plays an important role in the generation of MSC-DCs.

IRF8 expression and TLR4-triggered inflammatory cytokine production are suppressed by Notch signaling.
IRF8, which is also known as interferon consensus sequence-binding protein, is a component of the IRF transcription factor family, and is one of the downstream signaling molecules in the TLR4 pathway. As shown in Fig. 2a and b, Notch inhibits both IL-12 secretion and IRF8 expression in MSC-DCs. Thus, we next sought to examine whether IL-12 secretion and IRF8 expression were related in MSC-DCs. A ChIP assay was used to ascertain whether IL-12 expression is directly controlled by IRF8 in MSC-DCs. We also used RNA silencing to knock down IRF8 expression and then detected the level of IL-12 in MSC-DCs. The results of the ChIP assay revealed that IRF8 directly controls IL-12 expression by binding to the promoter region of the IL-12 gene, and silencing IRF8 in MSC-DCs markedly decreased the level of IL-12 (Fig. 2c, d). Thus, Notch inhibits IRF8 expression, and IRF8 regulates IL-12 secretion in MSC-DCs.
Notch-controlled IRF8 expression depends on RBP-J
Canonically, the ability of Notch to control downstream gene expression depends on RBP-J, the major nuclear transducer of the Notch signaling that plays a key role in signal transduction [31,32]. However, Notch-independent RBP-J activities have also been reported [33]. Further studies are required to determine whether Notch-controlled IRF8 expression depends on RBP-J.
We used RNA interference to knock down RBP-J expression in imDCs and then cocultured these cells with Sca-1+CD117−Lin− MSCs. Compared with the control group, IRF8 expression was significantly reduced in the siRNA-RBP-J-transfected imDCs, suggesting that IRF8 is the target of RBP-J-mediated regulation (Fig. 3a). Furthermore, the level of IL-12 was consistent with IRF8 expression (Fig. 3b). Based on these data, Notch regulates IRF8 expression in an RBP-J-dependent manner in MSC-DCs. We next detected the acetylation state of RBP-J in both MSC-DCs and DAPT-treated MSC-DCs using coimmunoprecipitation (Co-IP) to further explore the underlying mechanisms and found that RBP-J bound to less SIRT1 (a deacetylase) in DAPT-treated MSC-DCs where Notch signaling was blocked compared with MSC-DCs (as control) (Fig. 3c).
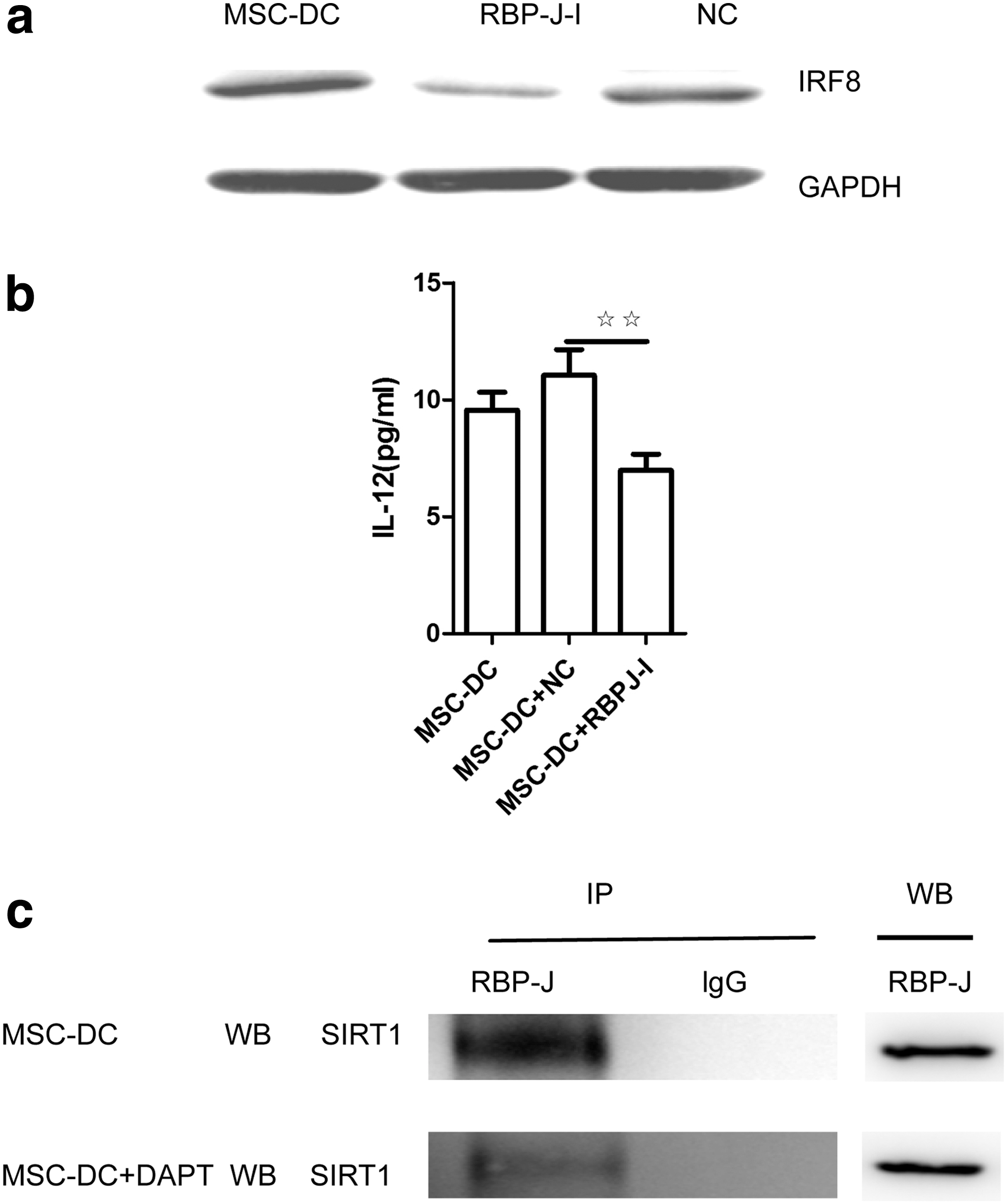
Notch signaling-controlled IRF8 expression was dependent on RBP-J. imDCs were transfected with a negative control siRNA (NC) or siRNA-RBP-J (RBP-J-I) and then cocultured with Sca-1+CD117−Lin− MSCs for 1 day.
Notch signaling regulates IRF8 expression through histone modifications
Epigenetic alterations are often involved in regulating gene expression. H3K4me3 and H3K27me3 are mediated by the histone methyltransferase complexes MLL (including MLL1, WDR5, RbBP5, and ASH2L) and PRC2 (including SUZ12 and EED), respectively. We examined the levels of H3K4me3 and H3K27me3 at the IRF8 promoter region by ChIP to verify the hypothesis that Notch signaling also regulates IRF8 via epigenetic modifications.
The H3K27me3 level at the IRF8 promoter was increased in MSC-DCs compared with maDCs, whereas DAPT-treated MSC-DCs showed a relative increase in the level of H3K27me3 and a decrease in the level of H3K4me3 (Fig. 4a), indicating that the chromatin structure of the IRF8 gene is maintained in a repressive state in MSC-DCs but might convert into an active state in the presence of DAPT. WDR5 of the MLL complex and SUZ12 of the PRC2 complex are primarily responsible for the H3K4me3 and H3K27me3 modifications, respectively, of IRF8 in MSC-DCs (Fig. 4b). Furthermore, based on the Co-IP results, IRF8 combined less SIRT1, a deacetylase, in DAPT-treated MSC-DCs than in MSC-DCs (as control) (Fig. 4c). Taken together, Notch signaling strongly downregulates IRF8 in MSC-DCs through the Notch-RBP-J signaling pathway and via changes in histone modification.
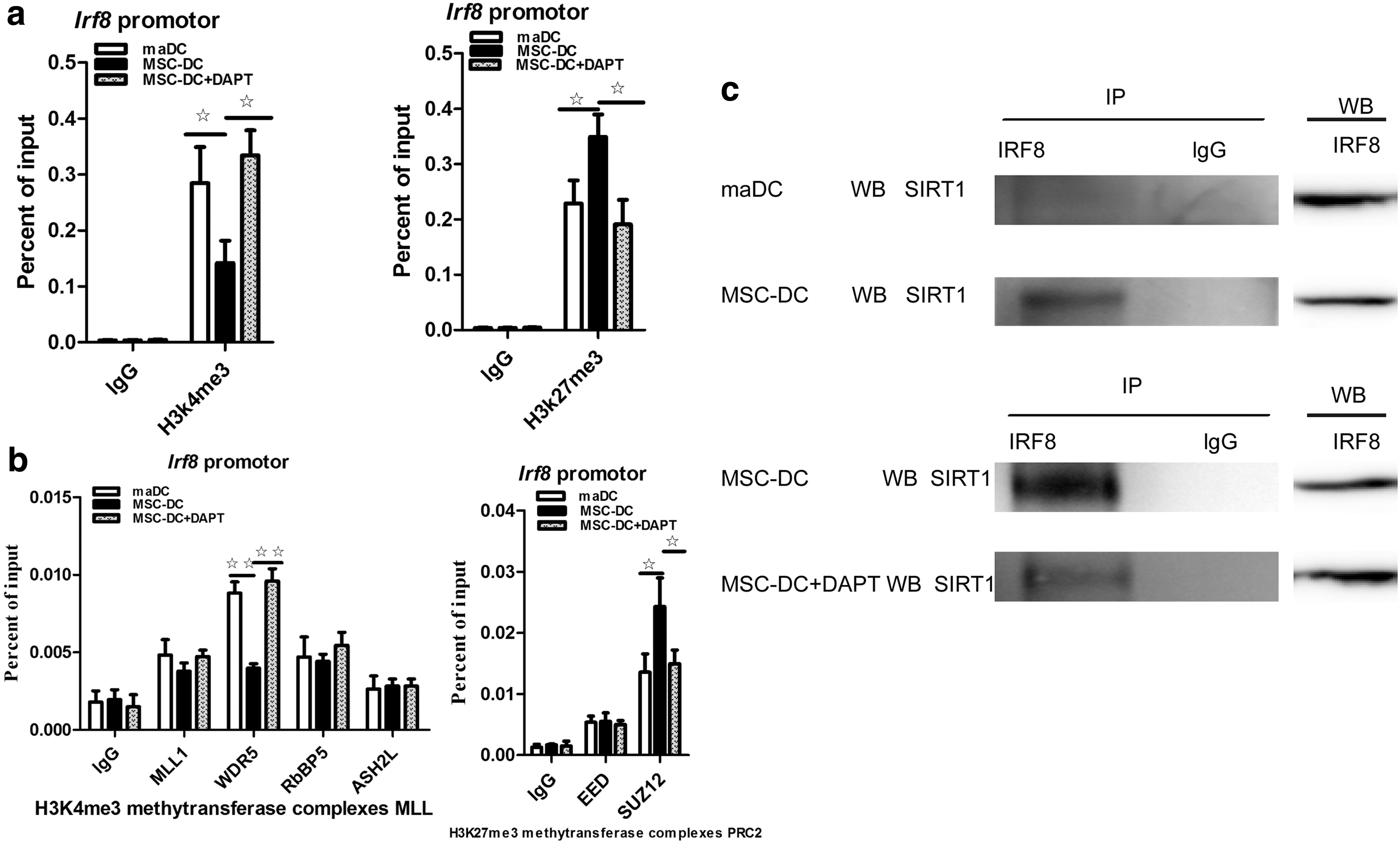
Notch signaling regulates IRF8 expression through histone modifications.
In summary, these experiments revealed a key role for Notch-RBP-J signaling in controlling imDC maturation. Collectively, these observations suggest that MSCs could be used in treatments of various immune dysregulatory diseases and that they represent a potential therapeutic agent for controlling the immune response.
Discussion
MSCs are immunoregulatory cells that migrate to sites of inflammation and exert potent immunosuppressive and anti-inflammatory effects. However, the mechanisms responsible for their inhibitory effects are not fully understood. IL-10, transforming growth factor-β, hematopoietic growth factor, prostaglandin E2, indoleamine 2,3-dioxygenase, and nitric oxide are some of the soluble molecules associated with the immune-suppressive effect of MSCs [34,35]. In this report, Sca-1+CD117−Lin− MSCs unexpectedly interfered with imDC maturation by suppressing TLR4-induced IRF8 expression through the Notch-RBP-J signaling pathway and histone modifications, thus revealing a new mechanism underlying their immunoregulatory effects. According to our data, Sca-1+CD117−Lin− MSCs significantly downregulated the expression of the myeloid lineage molecule CD11c, the presentation molecule Ia, and the costimulatory molecules CD80, CD86, and CD40 in imDCs upon LPS stimulation and largely altered the expression patterns of DC-specific transcription factors. Activated Notch-RBP-J signaling in imDCs might restrain TLR4-stimulated IRF8 expression, and IRF8 further controls the levels of secreted IL-12 and TNF-α.
DCs are important APCs, and they initiate immune reactions or induce tolerance according to their maturation state or subset. DCs recognize invading pathogens through TLRs and are activated by binding to TLR ligands, which leads to the production of inflammatory cytokines such as TNF-α and IL-1 and cytokines of the IL-6 and IL-12 family that regulate T cell differentiation [36]. In this study, the activation of imDCs by TLR4 not only increased the production of inflammatory cytokines, including TNF-α and IL-12, and decreased the secretion of the anti-inflammatory cytokine IL-10 but also changed the expression pattern of DC-specific transcription factors. As shown in Fig. 2b, the expression levels of PU.1, TRAF6, and TCF4 were decreased and the levels of IRF4, IRF8, RelB, Id2, and Ikaros were increased upon stimulation with LPS. However, MSCs reversed these changes in cytokine secretion by imDCs and altered the expression of several DC-specific transcription factors in MSC-imDC cocultures (Figs. 1c and 2b), suggesting that MSCs influence the fate decisions of maturing imDCs and induce the generation of MSC-DCs instead of maDCs.
Among TLRs, TLR4 recognizes bacterial lipopeptides and LPSs and induces cytokine production via the downstream signaling molecules IκB kinases (IKKs), mitogen-activated protein kinases and IRF [37]. We detected increased expression levels of IRF8, P-IKKa/b, P-p38, and P-JNK and higher levels of IL-12 and TNF-α secretion in imDCs following LPS stimulation, but only IRF8 expression and the levels of secreted IL-12 and TNF-α were restored to normal levels in the presence of MSCs. No changes in the expression levels of P-IKKa/b, P-p38, or P-JNK were detected in the presence or absence of MSCs (data not shown). Taken together, we postulate that MSCs influence the imDC maturation transition by controlling IRF8 expression. Therefore, we primarily focused on the role of IRF8 in the present study.
The Notch signaling pathway has been implicated in the development and differentiation of DCs. As shown in the study by Lewis et al., Notch signaling is required for DC differentiation and exerts DC subset-specific effects on DC fate specification [38]. According to the study by Zhang et al. [13], Notch signaling suppresses TLR-triggered inflammatory responses in macrophages by inhibiting extracellular signal-regulated kinase 1/2-mediated nuclear factor κB activation. TLR responses are regulated to achieve coordinated responses that are appropriate for the cellular environment. The unrestrained activation of TLR responses leads to excessive inflammation and tissue damage and contributes to the pathogenesis of inflammatory disorders. Therefore, TLR signaling is subject to negative regulation and feedback inhibition [39].
However, the molecular mechanisms that are regulated by Notch signaling in MSC-imDC cocultures remain largely unknown. Determining the mechanisms by which specific aspects of TLR responses are regulated is important for understanding the balance between the beneficial and potentially toxic effects of TLR stimulation. In this study, Sca-1+CD117−Lin− MSCs influenced imDC maturation through the Notch signaling pathway and which suppressed TLR4-triggered IRF8 expression and IL-12 and TNF-α secretion by imDCs. Additional miRNA interference assays and ChIP analysis revealed that IL-12 secretion depends on IRF8 in an RBP-J-dependent manner in MSC-DCs.
Strikingly, IRF8 was also reported to play a critical role in regulating several proinflammatory genes (including IL-12) that function during macrophage activation [40 –42]. Further studies are required to determine whether these pathways are related. Our findings highlight the importance of Notch signaling in MSC immunoregulatory functions. Similarly, the Notch signaling pathway is activated in DCs in vivo by Notch ligands expressed on BM stromal cells [43,44]. However, this report is the first to define the effects of MSCs on the fate of imDC maturation via the Notch signaling pathway.
Based on emerging evidence, TLRs also induce the remodeling and posttranslational modification of chromatin (histone “marks”) that either activates or silences gene expression [45,46]. The redundant combination of multiple active/suppressive histone modifications and the existence of bivalent domains indicates the complex nature of gene regulation by histone modifications. In this study, epigenetic mechanisms and transcriptional signaling pathways were both involved in controlling the imDC maturation transition. Blockade of Notch signaling increased H3K4me3 levels and decreased H3K27me3 levels at the IRF8 promoter region via mechanisms mediated by WDR5 and SUZ12, respectively, resulting in the upregulation of IRF8 expression in the MSC-DC + DAPT group. Thus, these data support our hypothesis that IRF8 expression is coregulated by Notch-RBP-J signaling and histone modifications.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (no. 81370466 and 81600087), the Young Teachers Training Program of Peking Union Medical College (2014zlgc0712), the Natural Science Foundation of Shandong Province (ZR2016HB45 and ZR2016HB05), and the open project of Shandong Collaborative Innovation Center for Antibody Drugs, No.CIC-AD1827.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.