
Abstract
Pluripotent stem cell-derived endothelial cells (ECs) have great potential to be used in vascular therapy or tissue engineering. It is also much desired to obtain arterial or venous ECs for specific applications. Factors that are critical for the proper arterial or venous differentiation from pluripotent stem cells still need to be understood. Here, we aim at investigating this problem deeper by examining neuropilin-1 (Nrp1), an early arterial marker that may be critical for arterial cell fate commitment. Using murine embryonic stem cells as the model system, this study investigates the neuropilin-1 (Nrp1) expression during the differentiation of pluripotent stem cells toward a vascular progenitor population. We hypothesize that Nrp1, an early arterial marker present in a developing embryo, may be more responsive when further induced in vitro toward an arterial fate. We developed a two-step differentiation approach that yielded a large percentage of Nrp1+ vascular progenitor cells (VPCs) and investigated their potential to become arterial ECs. We have defined the culture parameters that contribute greatly to the emergence of Nrp1+ VPCs: certain soluble factors, especially Wnt and BMP4, early cell-cell contact, and hypoxia. Subsequent isolation of this population demonstrated a highly proliferative and network-forming behavior. The Nrp1+ VPCs exhibited increased gene expression of several Notch pathway-related arterial markers compared with Nrp1− VPCs. Most importantly, Nrp1+ VPCs demonstrated a dramatically greater response to hemodynamic stimuli by upregulating many arterial markers whereas Nrp1− VPCs have very little response. Surprisingly, these differences between Nrp1+ and Nrp1− VPCs are not evident with vascular endothelial growth factor (VEGF) treatment. Our data suggest that Nrp1+ VPCs may serve as the arterial progenitor by enhanced response to hemodynamic flow but not to VEGF, whereas Nrp1− VPCs lack the plasticity to become arterial ECs. The findings of this research indicate that Nrp1+ VPCs in the murine model act as an important step in the arterial differentiation process.
Introduction
P
To facilitate this process, much can be learned from the developmental biology. During normal vascular development, arterial-venous specification appears earlier than the onset of blood flow in the embryo [16], demonstrating that arteriovenous cell fate decision is genetically predetermined in the heterogeneous cell population [17]. For example, preflow segregation of Neuropilin-1 (Nrp1) and Neuropilin-2 (Nrp2) to the arterial and venous plexus regions, respectively, was observed in chick embryos before EphrinB2 appeared [18]. Moreover, lineage-tracking studies revealed that chosen groups of angioblasts from the posterior lateral mesoderm migrate solely to either the dorsal aorta or the cardinal vein, never both, implicating some predetermination before or during the migration [19]. Following the earlier arteriovenous specification, environmental factors such as blood flow are indispensable driving forces that control and maintain the cell identity later on [20], suggesting a plasticity of earlier vascular progenitors.
Nrp1 is one of the earliest markers in distinguishing arteries from veins in a developing embryo. When chick embryos were examined at an early somatic stage, Nrp1 preferentially localized to the dorsal aorta, cardiac region, and vitelline arteries of the yolk sac; whereas Nrp2 and Tie2 became restricted to the vitelline veins and the cardinal vein [21,22]. Mukouyama et al. also noticed their importance in arteriogenesis while studying the embryonic mouse limb skin. They found that in primitive capillary plexus EC progenitors adjacent to the vascular endothelial growth factor (VEGF)-emitting nerve express Nrp1, a co-receptor for VEGF, with the outcome of arterial differentiation biased by a VEGF-Nrp1 positive-feedback loop [23,24]. In contrast, cells lacking Nrp1 expression develop into venous ECs [23,24]. Based on these findings, we hypothesize that stem cell-derived Flk1+Nrp1+ cells serve as arterial EC progenitors. We believe that this subset cell population is predisposed to arterial differentiation and can be selected to guide arterial differentiation in combination with environmental cues. As Nrp1 is present before and at the start of embryonic blood flow, isolating and examining this vascular progenitor population may elucidate the period of arteriovenous plasticity of stem cell-derived ECs in vitro.
Here, we conducted an in-depth study on various factors that may influence Nrp1 expression during early embryonic differentiation, and we have discovered key factors that contribute toward an Nrp1-enriched vascular progenitor cells (VPC) population in vitro. We then compared Nrp1+ with Nrp1− VPCs in their ability to undergo arterial differentiation.
Materials and Methods
The detailed materials and methods are described in detail in Supplementary Materials (Supplementary Data are available online at
Results
Nrp1 expression in monolayer differentiation
There are few studies done in an in vitro setting that investigate the differentiation of Nrp1+ vascular cells. Cimato et al. established that Nrp1 expression emerges before CD34 during endothelial differentiation and Flk1+Nrp1+ cells form endothelial-like colonies [25]. Prasain et al. showed that CD31+Nrp1+ cells differentiated from human embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) liken to cord-blood endothelial colony-forming cells [26]. Whether these progenitor cells also exhibit arterial EC potential is unknown. It is also unknown which in vitro conditions would lead to high Nrp1+ VPCs. We first tested a basic protocol in differentiating mouse ESCs (mESCs) into VPCs. The mESCs were cultured as a monolayer on 0.1% gelatin with serum-free, chemically defined basal medium containing 30 ng/mL VEGF and 5 ng/mL BMP4 (VB medium). They were seeded at an equivalent of 50,000 cells/35 mm dish. The medium was applied from day 0 onward (Fig. 1A). The mRNA expression for Nrp1 and the early/late vascular markers, Flk1 and VEcad, was investigated from day 3 to 10 (Fig. 1B). Nrp1 peaks on day 5 and decreases sharply the next 2 days before rising more consistently after day 7. Nrp1 expression is the highest on day 10 of this timeline. In contrast, Flk1 mRNA expression peaks the highest on day 4 at around 30-fold and then disappears at a later time point. This early rise of Flk1 on day 2 or 3 of vascular differentiation has been noted in past studies [1,12]. VEcad, a more mature endothelial marker, fluctuates at low levels but exhibits an upward trend on later days as expected. Flow cytometry analysis of Nrp1 and Flk1 protein expression confirms the gene expression trend observed (Fig. 1C).

Investigation of Nrp1 expression in mESCs differentiated in a monolayer culture.
This noncoinciding expression of Flk1 and Nrp1 in monolayer culture was also observed when cells were cultured in different medium and extracellular matrix (ECM) coating. In this study, fibronectin (1 μg/cm2) was used as coating and a medium containing 30 ng/mL VEGF, 5 ng/mL BMP4, and 12.5 ng/mL FGF2 was added from the beginning to the time of flow cytometry analysis (Supplementary Fig. S1A, B). Flk1 appeared early on day 4 (∼18%) and decreased quickly the next day to ∼7%. Meanwhile, Nrp1 did not appear in appreciable levels until day 6 (∼16%). Flk1 protein expression did not rise again during this period. There seems to be a very narrow window in these conditions to isolate the already low percentage of Flk1+Nrp1+ VPCs, and it is proved very difficult to find a good time point to sort out a sufficient number of Flk1+Nrp1+ cells using this culture condition.
FGF2 is a critical angiogenesis inducer and is found to be an essential component in maintaining Flk1+ cells [7,27]. To test whether FGF2 can improve Flk1+Nrp1+ vascular differentiation, we used the same gelatin coating and added 12.5 ng/mL FGF2 into the VB medium (VBF medium). Gene expression was analyzed between day 4 and 7 (Fig. 1D). Nrp1 expression was consistently higher when compared with VB medium, with levels reaching ∼1,000-fold on day 6. Flk1 also demonstrated more steady levels of expression, with fold change remaining between ∼18- and ∼32-fold. However, in both media, Flk1 dropped by day 7.
Using the VBF medium, we also checked whether different ECMs significantly influenced Nrp1 and vascular markers (Supplementary Fig. S2A). The ECMs investigated were the following: 0.1% gelatin, 1 μg/cm2 fibronectin, and 2 μg/cm2 laminin. The differences were not large. Overall, Nrp1 expression on day 10 was higher with the gelatin or laminin coating. No significant improvement in Flk1 and VEcad expression was observed across the different ECMs. We also examined the gene expression levels for different ECMs (0.1% gelatin, 2 μg/cm2 collagen type IV, and 1 μg/cm2 fibronectin) at earlier time points from day 3 to 5 (Supplementary Fig. S2B). Again, it was difficult to conclude which ECM was ideal for Nrp1+ vascular differentiation as the differences were minor. These data suggest that although it is possible to derive overall Nrp1+ cells at early time points, vascular differentiation is not very efficient, as demonstrated by the low percentage of Flk1+ or VEcad+ cells. Thus, it is difficult to derive a sufficient amount of Flk1+Nrp1+ or VEcad+Nrp1+ cells under this condition.
Early cell-cell contact during vascular differentiation is critical for Nrp1 expression
Apart from 2D differentiation, some protocols rely on the formation of embryoid bodies (EBs) before differentiation as they mimic more closely a 3D interaction of cells undergoing vasculogenesis in a developing embryo. We investigated whether EB culture would lead to improved Nrp1+ VPC induction. To control size of EBs and keep them largely consistent, we used the Aggrewell™ 400 24-well plates (Stem Cell Technologies) to form the EBs (Fig. 2B). Using the original VB medium, EBs were formed at ∼500 cells per EB. This culture method led to a ∼394-fold and ∼914-fold increase in Nrp1 at later time points, day 9 and 10, respectively, when compared with that of gelatin monolayer culture (∼194- and 213-fold, respectively) (Fig. 2A). Similarly, Flk1 levels reached up to ∼44-fold on day 10 under EB culture, whereas Flk1 expression remained below ∼15-fold for the entire observed period under monolayer culture. In terms of VEcad expression, gene expression became comparable for both culture methods by day 9 and 10 (∼22- and ∼61-fold for gelatin culture, whereas it was ∼31- and ∼77-fold for EB culture). Overall, the EB culture is more consistent in driving the differentiation toward both Flk1+ and Nrp1+ compared with the monolayer culture.
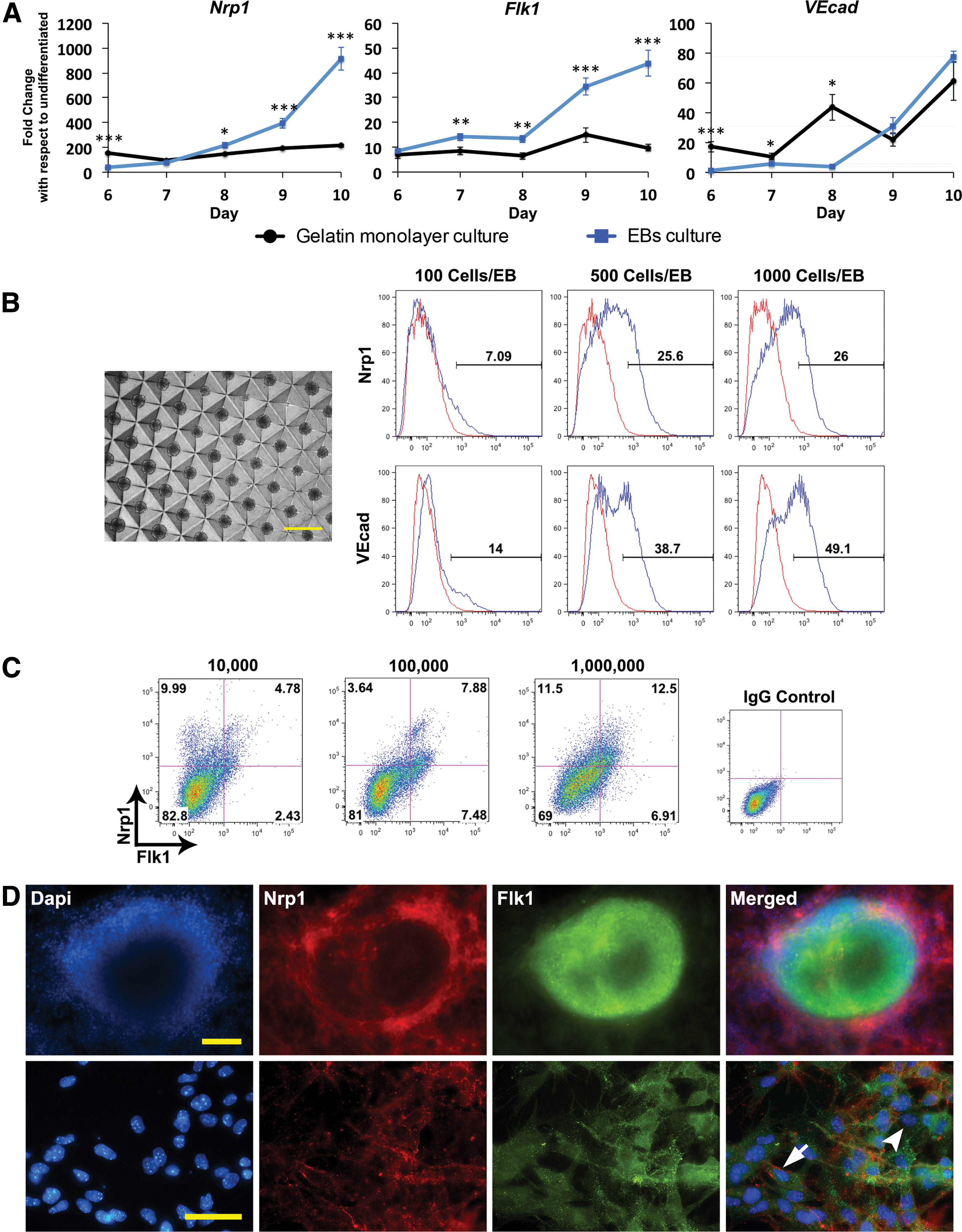
Nrp1 expression requires early cell-cell contact induced by EB culture or a high seeding density.
To determine whether the diameter of the EB influences Nrp1 expression, we tested EBs composed of 100 cells, 500 cells, and 1,000 cells on day 15 (Fig. 2B). Small EBs of 100 cells yielded significantly less Nrp1 (∼7%) than those of 500 and 1,000 cells (∼25–26%). Similarly, VEcad protein expression increased notably with 500- and 1000-cell EBs. It appears that there is a minimum EB size that must be maintained to achieve a robust percentage of Nrp1+ VPC population. To examine the pattern of Nrp1 and Flk1 expression, immunofluorescent staining of EB on day 10 of differentiation showed that Nrp1 is expressed in cells within the EBs and also in cells that outgrow from the EBs, whereas Flk1 is expressed mostly in cells within the EBs. Close examination of the staining confirmed that both Flk1+Nrp1+ (arrow) and Flk1+Nrp1− (arrowhead) cell populations co-exist in the culture (Fig. 2D). EBs also stained positive for CD34 and VEcad (Supplementary Fig. S3).
EBs promote 3D intercellular interaction immediately at the onset of differentiation, which may lead to more efficient differentiation. To determine whether early contact among cells contributes to the increase in Nrp1+ VPCs, we investigated seeding density in 2D monolayer culture. Cells were plated on a 35-mm well at an initial seeding density ranging from 10,000 to 1 million cells. On day 10, Flk1+Nrp1+ VPCs were the highest (∼13%) when differentiation began with 1 million cells, suggesting that initial cell-cell contact is beneficial for differentiation, as this is also the case in EBs differentiation.
A multi-step approach of adding soluble factors improves Flk1+Nrp1+ expression
We used a simplified protocol of adding soluble factors in one step from beginning to end. However, this differentiation system is not as reflective of vasculogenesis in the embryo as developmental signals were introduced in a stepwise transition. It is unknown as to what extent a multi-step differentiation method affects the emergence of Nrp1+ VPCs. To start with, we investigated a 2-step EB culture approach where only basal medium was used for the first 4 days before transitioning to 30 ng/mL VEGF and 12 ng/mL BMP4 (Fig. 3A). Simply delaying the addition of these two growth factors led to an overall increase in gene expression for Nrp1, Flk1, and VEcad (Fig. 3B). Flk1 expression became more upregulated in the two-step method, increasing gradually from ∼59 on day 6 to 100-folds on day 10. By contrast, the one-step method leads to only ∼16- to ∼38-fold increase for Flk1. Flow cytometry analysis confirmed that using a two-step differentiation system enhanced the presence of Flk1+Nrp1+ subpopulation (Fig. 3C). VEcad expression for both methods remained statistically not significant until day 10 when there is a two-fold increase for the two-step method.
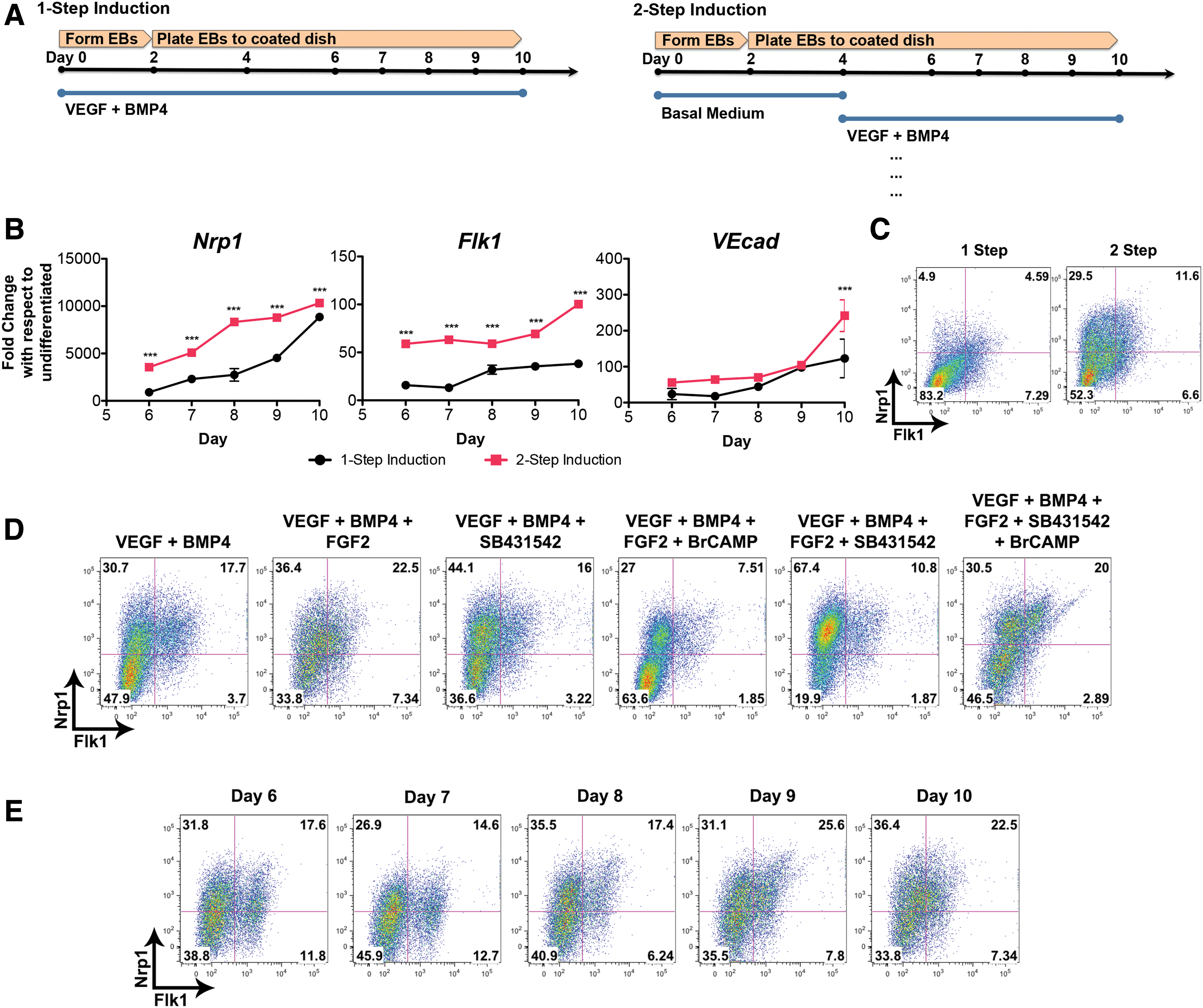
Multi-step approach to adding soluble factors improves both Nrp1 and Flk1 expression.
Soluble factor combinations can have an impact on the expression of Nrp1+ VPC differentiation. Using the EB culture approach, we examined different combinations and examined the percentage of Nrp1+ VPCs on day 10 by FACs analysis (Fig. 3D). From this study, a medium containing VEGF, BMP4, and FGF2 (VBF) gave one of the highest fractions of Flk1+Nrp1+ cells (∼23%). The medium containing VEGF, BMP4, FGF2, SB431542, and BrcAMP also exhibited a high percentage of Flk1+Nrp1+ cells (∼20%). This medium was used in a study by Chiang and Wong to induce hemogenic endothelium from mESCs [7]. These two media, especially VBF, along with VB medium also led to an improved percentage of CD34 and VEcad protein expression (Supplementary Fig. S4A).
Using the VBF medium, we observed the emergence of Flk1+Nrp1+ cells from day 6 to 10 by flow cytometry analysis (Fig. 3E). For this medium, day 9 or 10 was the most optimal time to isolate Flk1+Nrp1+ cells for further study. VEcad+Nrp1+ expression did not improve over the time course; however, CD34+Nrp1+ expression increased gradually from ∼19% on day 6 to ∼35% on day 10 (Supplementary Fig. S4B).
BMP4 promotes Nrp1 expression, whereas CHIR99021 encourages Flk1 expression
It is known that early BMP4 exposure stimulates mesodermal differentiation [28 –31]; how it affected Nrp1+ VPC expression was unknown. CHIR99021 (CHIR or C), a GSK3β inhibitor that ultimately activates the canonical Wnt pathway, has also been reported to enhance vascular differentiation when introduced briefly and early in the first 2 or 3 days of induction [32]. Meanwhile, Naito et al. observed that later Wnt activation in EB differentiation led to hematopoietic/vascular differentiation [33]. CHIR's effect on the differentiation of Nrp1+ VPCs has not yet been investigated. Therefore, we expanded on our study of the soluble factor combinations by looking at whether BMP4 and CHIR affect Nrp1+ VPCs.
We utilized the two-step EB culture system and analyzed by flow cytometry Flk1 and Nrp1 protein expression (Fig. 4A, B). When compared with VF medium, the addition of either 3uM CHIR or 12 ng/mL BMP4 (VFC or VBF) led to an increase in Flk1+Nrp1+ expression. Noticeably, BMP4 clearly upregulated Nrp1 expression as the overall cell population exhibits an upward shift toward Nrp1+. Conversely, CHIR markedly improved overall Flk1 expression as the cell population shifts toward more Flk1+. These data suggest that BMP4 promotes Nrp1 expression whereas CHIR promotes vascular differentiation. The addition of both BMP4 and CHIR (VBFC) led to a significant increase in the Flk1+Nrp1+ subpopulation (∼40%). Interestingly, phase-contrast images revealed that there were distinct morphological differences caused by the combinations of soluble factors used. For example, VF and VFC medium resulted in aggregates of floating EBs even in the presence of the 0.1% gelatin coating. Both VBF and VBFC conditions yielded better cell attachment and spreading on tissue culture plastic, implying that BMP4 may be responsible for this morphology (Fig. 4C). To confirm the vascular progenitors in the differentiation protocol, cells sorted for Flk1 were confirmed for endothelial markers, VEcad and CD34, and demonstrated uptake of acetylated LDL (Fig. 4D).
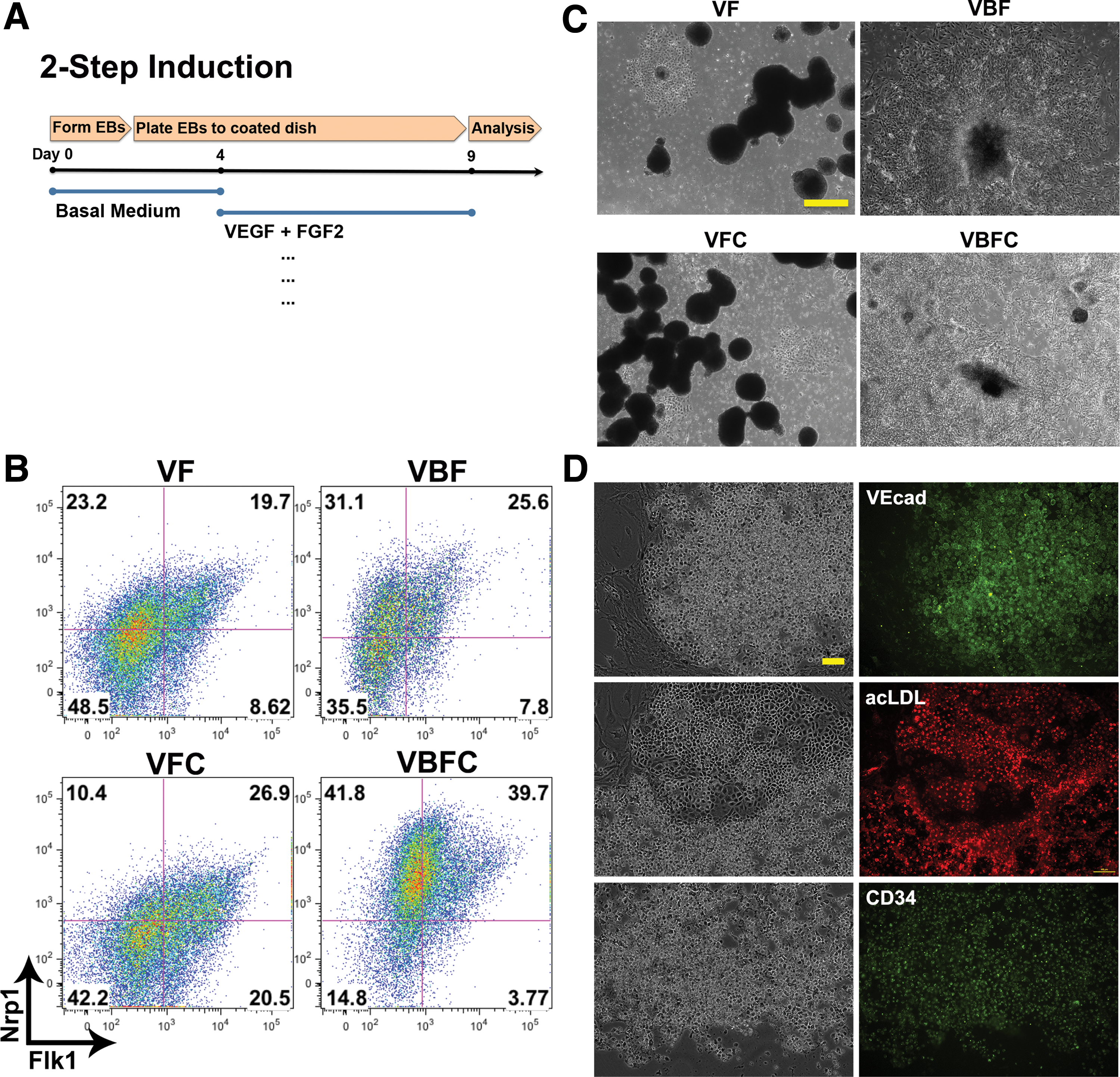
BMP4 promotes Nrp1 expression, whereas CHIR encourages Flk1 expression.
During development, the timing of BMP4 and Wnt signaling is critical for vascular differentiation. To test whether this also affects the in vitro stem cell differentiation, we investigated adding BMP4 and CHIR at earlier time points in differentiation. When BMP4 (12 ng/mL) and/or CHIR (3 μM) were added from day 2 and 4 instead of the basal medium, it initially increased the Flk1+Nrp1+ population on day 9 (Supplementary Fig. S5A, B). However, by day 10, we observed comparable percentages of Nrp1+ VPCs in both basal medium and BMP4+CHIR conditions (Supplementary Fig. S5C, D). In addition, other factors such as serum, BMP4 alone, CHIR alone, or VEGF plus BMP4 and CHIR did not significantly influence the percentage of Flk1+Nrp1+ cells, suggesting that these factors have little effect when added too early (day 2–4) during the differentiation.
To determine whether CHIR and BMP4 are required more in the early stage or in the late stage of vascular differentiation, we added CHIR or BMP4 either from day 2 to 8 or from day 8 to 14 (Supplementary Fig. S6). It appears that the longer and earlier exposure to CHIR during differentiation led to more upregulation of VEcad and CD34. However, it did not necessarily lead to improved Nrp1+ VPC differentiation as most increases in VPCs are Nrp1−.
Nrp1 expression increases under hypoxic differentiation conditions
Hypoxia drives arterial differentiation through the Adrenomedullin and the Notch pathway [34,35]. We evaluated whether a hypoxic environment is a necessary component in the differentiation of Nrp1+ VPCs. For this study, mESCs were induced toward the vascular fate by using the two-step VBFC system. A low oxygen level of 5% was introduced from day 0 onward. Nrp1 and endothelial markers all exhibited upregulation in response to hypoxia (Supplementary Fig. S7A). Nrp1 showed a maximum fold change of ∼23,000-fold in 5% oxygen; in contrast, 20% oxygen conditions only yielded ∼12,000-fold. Similarly, Flk1 upregulated significantly from day 6 to 8 but the difference decreased after day 9. CD34 also showed increased expression under hypoxic conditions. An overall increase in Nrp1 protein expression was confirmed by the upward shift of the cell population toward Nrp1+ by flow cytometry when observed on day 9 (Supplementary Fig. S7B). Interestingly, hypoxia increased both EphrinB2, an arterial marker, and EphB4, a venous marker, in the Flk1+ population (Supplementary Fig. S7C). These data suggest that hypoxia alone may not be a specific inducer of arterial differentiation. It may promote overall vascular progenitors in an uncommitted state by increasing both arterial and venous markers.
Nrp1+ VPC subpopulations exhibit some but not all arterial-related markers, and have limited response to VEGF exposure
After developing a differentiation method that yields a robust percentage of Nrp1+ and Nrp1− vascular progenitors, we next characterized them by comparing their potential to become arterial or venous ECs. Using the two-step VFC differentiation, we cultured the mESCs until day 10 and purified the Flk1+Nrp1+ and Flk1+Nrp1− populations through FACS for mRNA analysis (Fig. 5A). Endothelial markers, such as VEcad and CD34, showed no statistical difference. Venous markers, such as Nrp2, EphB4, and Coup-TFII, also demonstrated no statistical difference. There was also not much change in gene expression for arterial markers, except for Nrp1 and Notch4. It is likely that these populations express earlier markers but have not yet committed to an arteriovenous identity. This hypothesis is also consistent with the observation of cells differentiated under hypoxic conditions until day 9 (Supplementary Fig. S7C), where the Flk1+ cells expressed both EphrinB2 and EphB4. Since Nrp1 expression appears to be sustained in later stages of differentiation, we induced differentiation by using the same medium for a longer time until day 15 where we believed the cells were more committed. VEcad+Nrp1+ and VEcad+Nrp1− populations were sorted and analyzed (Fig. 5B). More clear differences in some arterial and venous markers were observed in this study. Meanwhile, venous marker EphB4 was decreased in the Nrp1+ population. For arterial markers, more Notch-related markers increased in the VEcad+Nrp1+ population: Dll4, Hey1, Notch1, Cxcr4, and EphrinB2. Despite these results, a clear arterial and venous distinction between Nrp1+ and Nrp1− cells is still in question, as the venous marker, Coup-TFII, actually increased, and the arterial marker, Notch4, decreased for the VEcad+Nrp1+ population. Interestingly, in VEcad+ sorting, cells lacking Nrp1 expression did not gain Nrp1 expression when re-plated in a culture dish for some time (8 days), whereas Nrp1+ cells maintained Nrp1 expression throughout the re-plated culture (Fig. 5C), demonstrating a possibility that Nrp1 expression is stable by this stage.
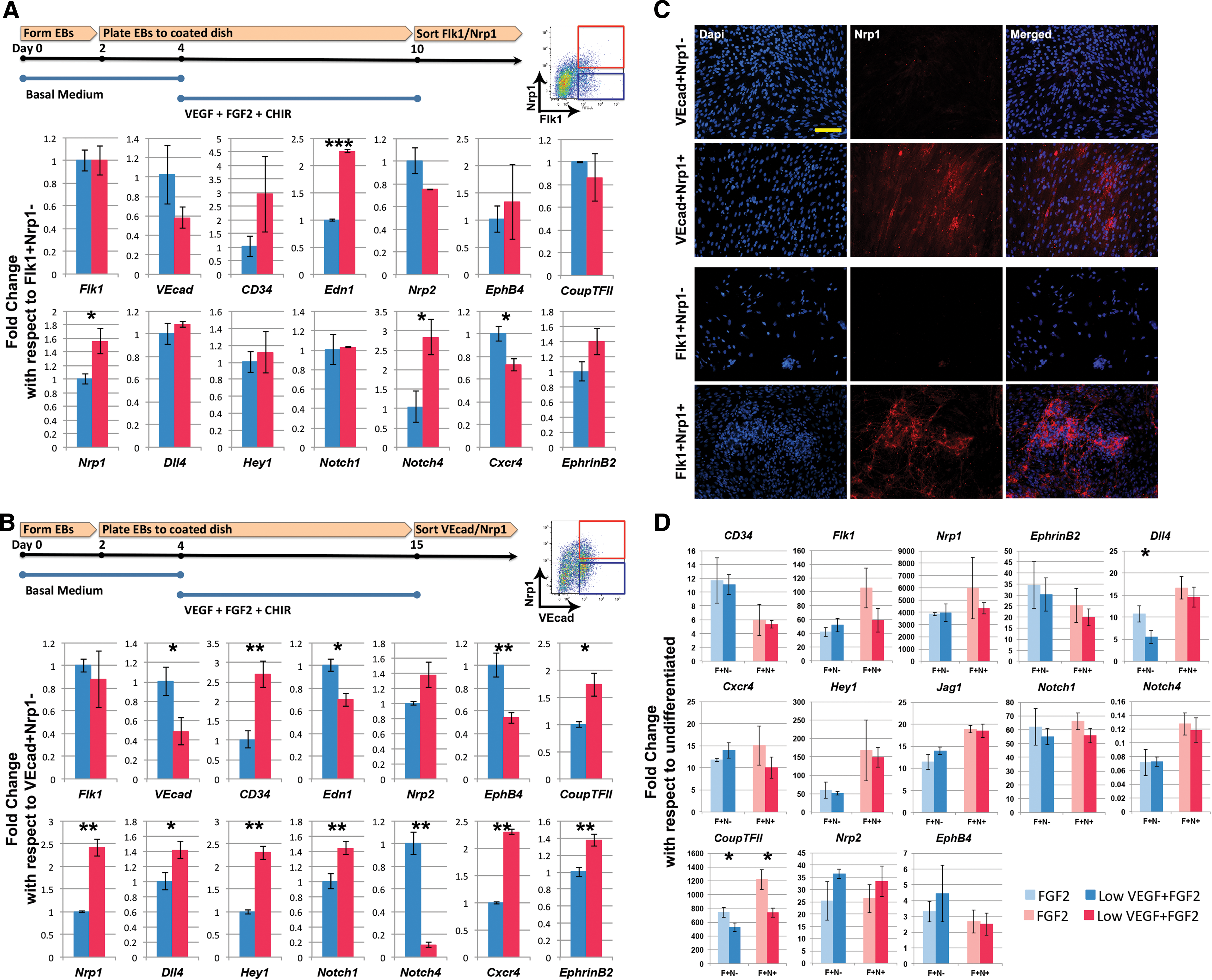
Sorted Nrp1+ VPCs demonstrate an incomplete preference toward an arterial phenotype.
We next investigated the two populations' response to VEGF. Flk1+Nrp1+ and Flk1+Nrp1− populations were sorted and exposed to 5 ng/mL VEGF and 12.5 ng/mL FGF2 for 3 days (Fig. 5D). Gene expression was then analyzed. Under VEGF+FGF2 condition, certain Notch-related markers slightly increased in Nrp1+ versus Nrp1−: Dll4, Hey1, Jag1, and Notch4. Under FGF2 condition, Hey1 and Notch4 were slightly higher in Nrp1+ versus Nrp1−. Nevertheless, these differences were small and not statistically significant. In addition, when comparing VEGF+FGF2 with FGF2 condition, there was not much difference for both cell populations, suggesting that VEGF treatment does not influence arterial differentiation, and Nrp1+ cells do not respond more to VEGF than the Nrp1− cells. Similarly, the VEGF or FGF2 treatment does not show a clear direction in the VEcad+Nrp1+ and VEcad+Nrp1− populations (Supplementary Fig. S8A). Again, in this instance, there is no clear indication of arteriovenous commitment.
Nrp1+ VPCs demonstrate enhanced sprouting
Nrp1 is associated with tip cell/stalk cell biology. We observed spontaneous sprouting of Flk1+Nrp1+ cells when re-seeded onto gelatin-coated plates, whereas Flk1+Nrp1− cells did not exhibit cord formation (Fig. 6A). When measured, the average number of nodes per mm2 area for Flk1+Nrp1+ cells was ∼52, whereas that of Flk1+Nrp1− cells was ∼18. In addition, the average branch length per mm2 area for the Nrp1+ vascular cells was ∼2.78 mm, whereas that of the Nrp1− cells was ∼0.85 mm. To confirm this observation, we performed a Matrigel tube formation assay on the VEcad+Nrp1+ and VEcad+Nrp1− populations as well (Fig. 6B). When seeded onto Matrigel with no growth factors and no serum, VEcad+Nrp1+ demonstrated spontaneous network formation, whereas VEcad+Nrp1− showed minimal sprouting. The average number of nodes for the Nrp1+ VPCs was ∼28, whereas that of Nrp1− VPCs was ∼9. Moreover, the average branching length was ∼2.24 mm for the Nrp1+ VPCs, whereas that of the Nrp1− VPCs was ∼0.87 mm. Despite these obvious morphological differences in the two populations, gene expression was less clear when examining those related to tip cells/stalk cells. For the Flk1-sorted VPCs, Nrp1-positive cells demonstrated higher levels of Pdgfb, Tek, and Hlx (Fig. 6C). However, for the VEcad-sorted cells, Nrp1-positive cells exhibited less Pdgfb, Nid2, and Tek until VEcad+Nrp1+ cells were exposed to 30 ng/mL VEGF for 3 days (Fig. 6D, E). It was then the trend reversed with an upregulation of Pdgfb, Nid2, and Hlx. It is likely that this variation is due to the very nature of tip cells and stalk cells being so dynamic that gene expression analysis before cells undergo a Matrigel assay is not a reliable indicator of tip/stalk behaviors.

Nrp1+ VPCs demonstrate increased sprouting over their Nrp1− counterpart.
VEcad+Nrp1+ cells respond more to flow shear stress than VEcad+Nrp1− cells by increasing gene expression of more arterial markers and decreasing EphB4
Fluid shear stress enhances both differentiation toward the vascular endothelial lineage and arterial identity [12,36 –39]. We next examined whether the VEcad+Nrp1+ and VEcad+Nrp1− populations respond differently to shear stress. Cells were seeded at equal densities onto an Ibidi flow chamber and integrated into a peristaltic pump system aseptically. Murine embryos aged 8.5–10.5 days postcoitum (dpc) experience laminar shear stress ranging from 0 to 5.5 dyn/cm2 [40]. By this time, vessels have already adopted an arterial or venous identity. To mimic the range of shear stress as close as possible, shear stress no higher than 5.5 dyn/cm2 was applied. We found that both VEcad+Nrp1+ and VEcad+Nrp1− populations respond to shear stress by changing their morphologies (Supplementary Fig. S8B). We then examined the gene expression of the selected arterial venous markers. VEcad+Nrp1+ populations were more responsive to fluid shear stress than the VEcad+Nrp1− populations, by upregulating many markers related to the Notch pathway: Notch1, Notch4, Hey1, and Jag1. Other arterial markers Nrp1, EphrinB2, and Cxcr4 as well as tip cell-related markers, Pdgfb and Unc5b, were also higher (Fig. 7A). The degree of upregulation was also higher: in VEcad+Nrp1+ cells, flow induced significant arterial marker upregulation compared with that of static. In contrast, for VEcad+Nrp1− cells, flow induced very little increase in most of the genes (Fig. 7A, table). Again, the arteriovenous distinction was not fully achieved as venous markers showed an upward trend (although not statistically significant) as well. However, after 48 h of shear stress, the VEcad+Nrp1+ cells showed a continued increase in EphrinB2, whereas EphB4, which was increased at 24 h, finally decreased, demonstrating a trend toward a more arterial distinction (EphrinB2+EphB4−). In contrast, the VEcad+Nrp1− population did not show this trend, suggesting that this population is unlikely to undergo arterial differentiation, even under the flow condition (Fig. 7B).
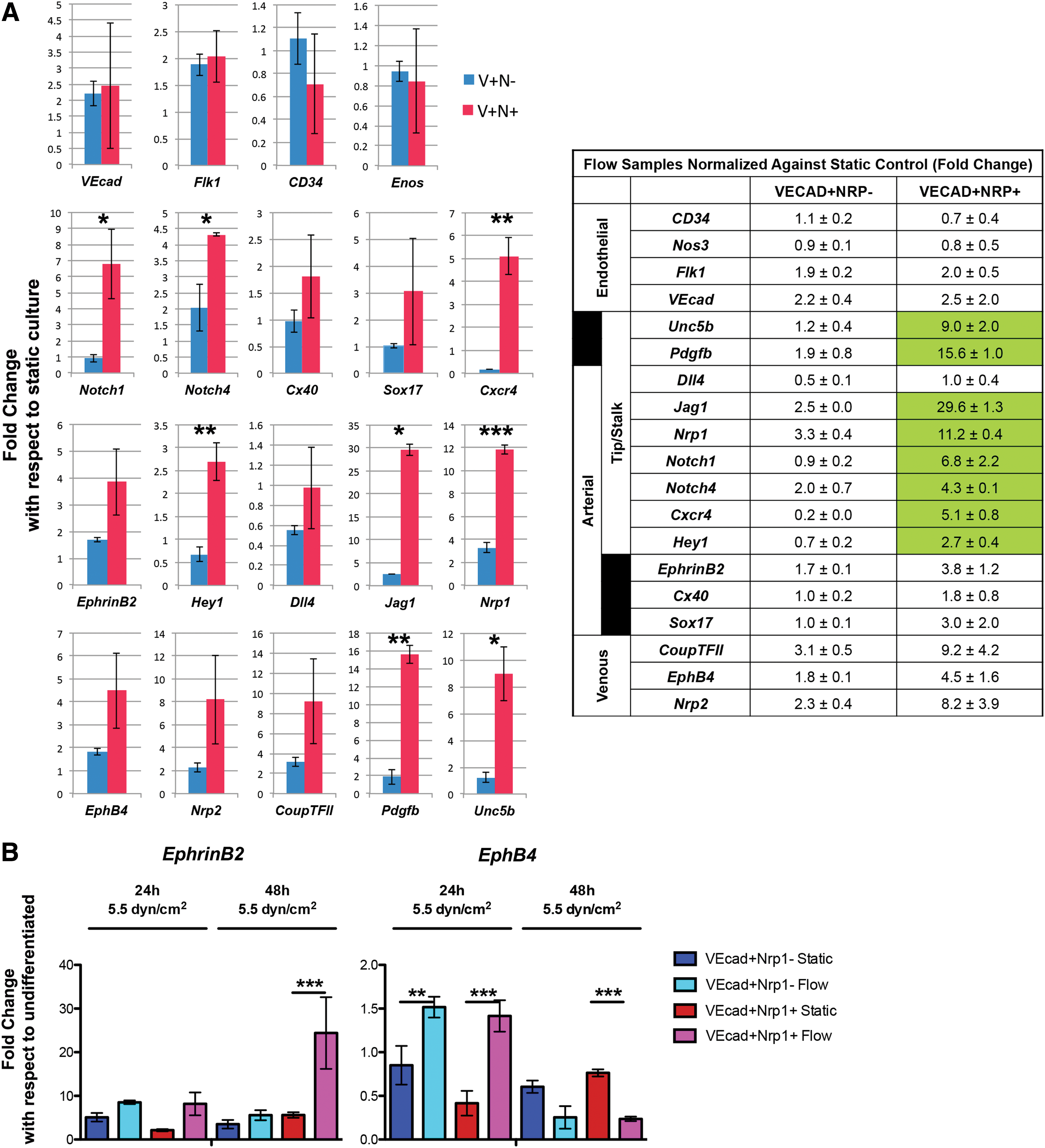
The VEcad+Nrp1+ population demonstrated heightened arterial- and tip cell-related response to shear stress compared with the VEcad+Nrp1− population.
Discussion
In this study. we found that several key factors are critical to the development of Nrp1+ VPCs from murine ESCs. Based on our findings, we have optimized the differentiation system to increase the emergence of these cells. Importantly, our study also suggests that Nrp1+ vascular cells are more responsive to hemodynamic flow to upregulate arterial markers than their Nrp1− counterpart.
Early intercellular interaction, that is, direct cell-cell contact and/or paracrine signaling, is crucial to obtaining a high yield of Nrp1+ VPCs. This feature can be attained by turning to 3D EB formation with >1,000 cells/EB or by seeding cells at a high density from the onset. There is another advantage to using the EB culture system: Monolayer culture requires soluble factors in the basal medium from day 0 to avoid anoikis, whereas EBs do not rely on them and can be cultured in simple basal medium for several days. In fact, delaying the introduction of exogenous factors until day 4 improved the yield of Nrp1+ VPCs. We, therefore, developed a two-step approach to encourage more Nrp1+ VPCs. It is possible, however, that monolayers can exhibit the same independence from exogenous factors if a higher seeding density than what was investigated in this study is used. In conclusion, early intercellular contact encourages cell survival and differentiation even without exogenous factors.
In this study, we observed in monolayer culture an early peak of Flk1 expression on day 2 or 3, which did not coincide with the later emergence of Nrp1 (around day 6 onward). However, we were able to obtain a later Flk1 expression that co-expressed with Nrp1. These observations are consistent with cell-tracking studies that demonstrated that there are two waves of Flk1+ progenitors: the first early wave directing toward a hemangioblast lineage with little cardiac fate potential and the second wave leading to cardiac, endothelial, and vascular smooth muscle lineages [41]. It is possible that the populations we isolated reflect this second wave of vascular progenitors.
We observed that activation of the Wnt signaling with CHIR throughout the differentiation process led to improved endothelial progenitor yield. The timing of the CHIR addition seems flexible as we saw comparable percentages of general Flk1+ vascular progenitors whether CHIR was implemented from day 2 onward or day 4 onward (Fig. 4 and Supplementary Fig. S6). These results coincide with previous studies where lateral mesoderm cells, except for those cardiac fated that required a specifically timed biphasic Wnt signaling, simply required a uniform Wnt/β-catenin manipulation [42].
BMP4 is another cytokine that seems to have a significant impact on Nrp1+ VPC differentiation. This growth factor is critical toward the differentiation of the mesoderm derived from the posterior primitive streak as well as the formation of blood islands in the yolk sac [43]. BMP4 has also been implicated in cell survival that is specific toward arterial ECs, but its role appears to be context dependent. Mature arterial ECs are resistant to BMP4-induced apoptosis, whereas venous and microvascular ECs undergo cell death [44]. It has been determined that high expression of SMAD6/7 in the arterial ECs contributed to the cell survival. Conversely, the BMP receptor-2 (BMPR2), a receptor that binds BMP2/4/7, promotes cell survival of pulmonary arterial ECs [45]. In this study, BMP4 led to upregulation of Nrp1, but this growth factor alone is not sufficient for co-expression of both vascular markers and Nrp1. Its overall effect on the yield of Flk1+Nrp1+ or VEcad+Nrp1+ populations seems to be dependent on other exogenous factors present.
In our differentiation study, continuous hypoxia enhanced the generation of Nrp1+ vascular progenitors in culture. Previous studies suggest that hypoxia increases arterial gene expression through two molecular pathways: Notch and adrenomedullin/cAMP. Low oxygen conditions upregulate Notch4, Hey1, Connexin40, and EphrinB2 whereas they downregulate COUP-TFII through the Notch canonical signaling [35]. Meanwhile, adrenomedullin/cAMP, also activated by hypoxia, causes an increase in Dll4 independent of Notch signaling. Yurugi-Kobayashi et al. observed that cAMP stimulation of stem cell-derived ECs led to a decrease in Coup-TFII and also a substantial increase in Nrp1, Notch1/4, EphrinB2, and Alk1 [13]. However, in our study, we observed that hypoxia also leads to the increased expression of general EC markers Flk1 and CD34, as well as both arterial marker EphrinB2 and venous marker EphB4. Therefore, we interpret this result as hypoxia enhancing the overall vascular differentiation efficiency to all lineages, but not necessarily specific to arterial differentiation.
This study demonstrated that both Flk1+Nrp1+ and VEcad+Nrp1+ VPCs exhibit more sprouting potential than their Nrp1− counterparts. It was believed that this was due to some connection to tip/stalk dynamics as Nrp1, along with Flk1, is expressed in tip cells [46]. However, after sorting the populations and immediately examining the genetic markers for tip/stalk phenotype, we found that the answer was less straightforward. Certainly, for Flk1+Nrp1+ cells, they had higher levels of tip cell markers than the Flk1+Nrp− cells. For the VEcad+Nrp1+ population, however, tip cell markers were not more upregulated than the VEcad+Nrp1− cells. It was only after treating these two populations with a high concentration of VEGF that we did observe that tip cell markers are more upregulated in the VEcad+Nrp1+ population. These observations confirm that the tip cell phenotype arises due to interaction between VEGF and the co-complex of Flk1 and Nrp1 [46 –48]. This study also implies that the presence of Nrp1 is critical to retaining sprouting ability as both Flk1+Nrp1− and VEcad+Nrp1− populations showed little or no sprouting and remained negative for Nrp1 protein expression. Meanwhile, the Nrp1+ VPCs showed retention of sprouting ability. Again, Nrp1 expression was present well after sorting (up to 8 days), indicating a certain stability of this marker in vitro, which is intriguing considering that tip cell/stalk cell plexus formation is a very dynamic process in which Nrp1 expression can quickly switch on and off between tip and stalk cells. Prasain et al. also established that Nrp1 expression remains robust in long-term culture in human PSC-derived ECFCs [26]. ECFCs remained ∼90% positive for Nrp1 at Passage 4 and ∼80% positive at Passage 14. These findings suggest that there is some level of fate commitment for the Nrp1+ & Nrp1− VPCs in culture, although it is possible that other unknown factors might dynamically change it during the tip/stalk cell formation in angiogenesis.
Nrp1 is a transmembrane glycoprotein that acts as a co-receptor for several classes of ligands: specific VEGF isoforms, class 3 semaphorins, and transforming growth factor β-1 (TGFβ1) [49 –51]. The presence of Nrp1/VEGFR2 co-receptor will enhance the growth factor signaling and promote proliferation [52]. Prasain et al. found Nrp1 to be crucial to the proliferative potential of cord-blood endothelial colony-forming cells [26]. In terms of arterial differentiation, it was found that in the early development stage, the expression of Nrp1 on EC progenitors and its interaction with nerve-derived VEGF are critical for EC differentiation toward arterial blood vessels [23,24]. Another study also showed that a high concentration of VEGF can direct endothelial precursors toward an arterial identity, whereas a low concentration can push them toward a venous fate [11]. Based on these data, we would predict that the Nrp1+ VPCs will have a stronger response to VEGF than the Nrp1− VPCs. However, our data showed that both populations have very little response to VEGF treatment, which does not seem to influence arterial differentiation. Recent studies have also confirmed that the exogenously added VEGF is not necessary for proper vascular differentiation in vitro, although the VEGF gene is absolutely essential for vascular development in vivo based on VEGF knockout experiments [32,53]. This is likely to be the case in our experiments, where exogenously added VEGF has very little influence on arterial differentiation.
The most striking finding is that the Nrp1+ and Nrp1− VPCs demonstrate dramatically different responses to hemodynamic stimuli: Nrp1+ cells are drastically more responsive to fluid shear stress by upregulating many arterial markers, whereas Nrp1− cells show very little response (Fig. 7A). More importantly, when Nrp1+ cells are exposed to shear stress for a longer period, arterial marker EphrinB2 consistently increases whereas EphB4 increases initially but then decreases below static level after 48 h, demonstrating a clear direction toward arterial differentiation (Fig. 7B). In contrast, for Nrp1− cells, EphrinB2 does not change and EphB4 decreases after 48 h. This type of response mimics the outcomes observed by Kudo et al. [54]. They studied vein grafts in both human patients and rat models. In both cases, EphB4 decreased but EphrinB2 did not increase, suggesting that mature venous ECs lack the plasticity to adapt fully to arterial hemodynamics. In this regard, the Nrp1− VPCs also lack the plasticity and cannot adapt to arterial flow condition. Our data point out the importance of examining Nrp1 for arterial EC differentiation. As there are many vascular differentiation protocols that never looked at Nrp1, it is difficult to know which one is suitable for arterial EC differentiation. If a protocol generates mostly Nrp1− VPCs, our data suggest that it is unlikely to make them arterial ECs by exposing them to flow.
Despite the enhancement of arterial differentiation via Nrp1, our data indicate that fluid shear stress alone still cannot derive the distinct arteriovenous phenotypes, as flow does not suppress but instead increases some venous markers at the same time. These studies suggest that current methods of soluble factors and hemodynamic flows are insufficient to obtain distinct arteriovenous differentiation. Further, a lack of arterial markers is often taken as the default phenotypes for venous ECs in some other studies. However, recent studies have shown that the definitive venous EC phenotype is not the default pathway of just lacking Notch activation but needs the active involvement of COUP-TFII [55]. Studies have identified COUP-TFII as the critical regulator of the EC phenotypic profile related to anti-thrombotic, pro-atherogenic, and osteogenic functions [56]. How to control COUP-TFII transcriptional regulation during stem cell differentiation to allow distinct venous EC differentiation is still unknown and warrants further studies.
In summary, this study presents some novel findings about ESC-derived Nrp1+ VPCs. We have determined the critical factors that are necessary for induction of high Nrp1 expression in vitro. Nrp1+ VPCs show some characteristics of tip cells, proliferate faster, and are enriched for several arterial markers. Nrp1+ VPCs have greater potential to become arterial ECs by responding better to hemodynamic flow, but not to VEGF stimuli. This study also revealed some level of fate commitment in Nrp1− VPCs as they do not re-gain Nrp1 expression in culture and also have limited response to hemodynamic stimuli; thus, they are unlikely to become arterial ECs. Taken together, our study suggests that Nrp1 is critically important for arterial differentiation and supports the notion of step-wise differentiation via Nrp1 and hemodynamic stimulation for better arterial EC derivation from stem cells.
Footnotes
Acknowledgments
This study was supported mainly by grants from the American Heart Association Scientist Development Grant (12SDG12050083 to G.D.) and the National Heart Lung and Blood Institute (R21HL102773, R01HL118245 to G.D.).
Author Disclosure Statement
The authors indicated no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.