
Abstract
Experimental cell research studying three-dimensional (3D) tissues in space and on Earth using new techniques to simulate microgravity is currently a hot topic in Gravitational Biology and Biomedicine. This review will focus on the current knowledge of the use of stem cells and specialized cells for tissue engineering under simulated microgravity conditions. We will report on recent advancements in the ability to construct 3D aggregates from various cell types using devices originally created to prepare for spaceflights such as the random positioning machine (RPM), the clinostat, or the NASA-developed rotating wall vessel (RWV) bioreactor, to engineer various tissues such as preliminary vessels, eye tissue, bone, cartilage, multicellular cancer spheroids, and others from different cells. In addition, stem cells had been investigated under microgravity for the purpose to engineer adipose tissue, cartilage, or bone. Recent publications have discussed different changes of stem cells when exposed to microgravity and the relevant pathways involved in these biological processes. Tissue engineering in microgravity is a new technique to produce organoids, spheroids, or tissues with and without scaffolds. These 3D aggregates can be used for drug testing studies or for coculture models. Multicellular tumor spheroids may be interesting for radiation experiments in the future and to reduce the need for in vivo experiments. Current achievements using cells from patients engineered on the RWV or on the RPM represent an important step in the advancement of techniques that may be applied in translational Regenerative Medicine.
Introduction
F
Astronauts and cosmonauts returning from long-term space missions exhibited various health problems, among them changes of the immune system, bone loss, muscle atrophy, ocular problems, and cardiovascular changes [12,13]. More than 30 years of biomedical studies in real or simulated microgravity (s-μg) have demonstrated important changes in biological processes of single cells. Space biologists investigated various cell types in space to find the molecular mechanisms responsible for the observed immune disorders, for example, references [14,15].
To prepare for future spaceflight missions, several devices aiming to simulate microgravity were developed to examine different cell types and their behavior when cultured under these conditions. A special interest in 3D culture arose when “National Aeronautics and Space Administration” (NASA) scientists introduced the rotating bioreactors [16,17]. MCS formation occurred within a few hours after the start of the culture in the HARV (high-aspect rotating vessel) bioreactor [16]. Similar results were obtained when thyroid cancer cells and normal thyrocytes were grown in a scaffold-free culture in the 2D fast rotating clinostat or the 3D random positioning machine (RPM) [18 –20]. Within 24 h compact 3D MCSs were detectable. The cytokines IL-6 and IL-8 seem to be involved in the initiation of MCS formation [6,19].
The rotating wall vessel (RWV) bioreactor has been used for long-term tissue engineering studies, using stem cells for the production of cartilage [21], bone [22], and other tissues, such as colon carcinoma cell growth within liver tumor organoids, serving as a model for metastasis growth [23]. In addition, the RPM has shown to be suitable for culturing MCS and tissue constructs of various cell types in long-term studies, which would be much harder or even impossible to generate under static conditions.
A recent publication showed that s-μg using the RPM promotes the osteogenic differentiation of bone mesenchymal stem cells (bMSCs) in the presence of an osteogenic cocktail [24]. In addition, the RPM has been used to engineer cartilage from human chondrocytes or intima constructs from human endothelial cells (ECs) [25].
This review will report on the current knowledge in tissue engineering using stem cells and specialized cells under microgravity conditions (Tables 1 and 2). The special focus of this article is to discuss tissue engineering of multicellular (tumor) spheroids, cartilage and bone constructs, tubular structures (preliminary vessels), as well as ocular tissue, using devices created to simulate microgravity on Earth and to report about spaceflights that allow cell biology in real microgravity.
3D, three-dimensional; AD, adherent cells; AKT, PKB, protein kinase B; CAV1, caveolin 1; CTGF, connective tissue growth factor; EGF, epidermal growth factor; ERK, extracellular signal-regulated kinase; FRC, fast rotating clinostat; fT3, free triiodothyronine; fT4, free thyroxine; HMVEC, human microvascular endothelial cells; IL-6/-8, interleukin 6/-8; ISS, International Space Station; MCS, multicellular spheroid; NF-κB, nuclear factor ‘kappa-light-chain-enhancer’ of activated B-cells; OPN, osteopontin; RCCS, rotating cell culture system; RPM, random positioning machine; RWV, rotating wall vessel; r-μg, real microgravity; s-μg, simulated microgravity; TLN-1, talin 1.
2D, two-dimensional; 5-FU, fluorouracil; ADSC, human adipose derived stem cell; bFGF, basic fibroblast growth factor; BMSC, bone mesenchymal stem cell; CPC, cartilage progenitor cell; ECM, extracellular matrix; EPC, endothelial progenitor cell; GFP, green fluorescent protein; HARV, high-aspect rotating vessel; MSC, mesenchymal stem cells; NGF, nerve growth factor; SCAP, stem cell from apical papilla; STLV, slow-turning lateral vessel; TGF, transforming growth factor; VEGF, vascular endothelial growth factor.
Research in Microgravity
Generating a microgravity environment to exploit the beneficial effect cells are showing under such conditions is not a trivial task. Ideally, experiments are conducted in weightlessness of space, accommodated in space laboratories. Accessing such platforms, however, is demanding, costly, and only a few flight opportunities open up each year. Therefore, alternative solutions for conducting experiments in a low-gravity environment are needed. During the last decades, several kinds of platforms, in-flight, as well as ground based, have evolved and are now available to scientists for their studies.
In-flight microgravity research platforms
Most prominent and intensively used are the in-flight research facilities of the International Space Station (ISS), provided in the “Columbus” module. By focusing on the cell biological experiments only, 25 investigations have been carried out in the “KUBIK” hardware and 6 investigations in the “BIOLAB” hardware so far (more details on the ISS experiments are available at the European Space Agency (ESA) Erasmus Experiment Archive at
Ground-based microgravity research platforms
Because Earth's gravity is an omnipresent force which cannot be removed or shielded off, multiple approaches are being used to create a microgravity or s-μg-like environment on ground. The main three options are: (a) establishing a “free fall” condition, (b) counteracting gravity by suspension, buoyancy, or magnetic levitation, for example, or (c) spatially distributing the gravity vector so that its action averages to zero. Depending on the intended experiments, not every method is applicable to create “artificial” microgravity.
Clinostats negate the effect of gravity by continuously rotating, for example, culture chambers around their horizontal axis. Such a treatment leads to constant free fall of suspended cells through the culture medium inside the chambers. To achieve such an effect, however, a rotation speed of 50–100 rpm around the horizontal axis is needed (Fig. 1). Modern pipette- or cuvette clinostats are implementing this principle. To study the behavior of adherent cells to a s-μg-like environment slide-flask clinostats are the instrument of choice. Photomultiplier clinostats are able to record luminescence measurements of cells during clinorotation and even clinostat microscopes are in use nowadays for in situ observations of subcellular structures [26].

A modification of the original clinostat principle is implemented in the rotating cell culture system (RCCS) or RWV bioreactor (Fig. 1), which has been developed by the US National Aeronautics and Space Administration (NASA) [27]. The system is frequently used to cultivate mammalian cells under s-μg-conditions as ground control either by a HARV or by a slow-turning lateral vessel (STLV) [28]. HARV oxygenates culture media through a flat gas transfer membrane and allows the 3D cultivation of suspended, as well as adherent, cells. In contrast, STLV oxygenates the media through a central core gas transfer membrane and, thus, provides an ideal environment for cell cultures that utilize microcarriers or scaffolds and for explant tissue cultures.
A different approach of generating a s-μg-like environment was proposed by Scano et al. in 1963, applying a so-called “random positioning machine” (RPM, Fig. 1) [29]. The RPM rotates samples not only around one axis like it is realized by clinorotation, but two. The motion pattern is defined in a way so that the gravity pull on the samples is distributed equally, spatially and temporally. In such a situation the gravity vector averages mathematically to zero [30,31]. The constant reorientation of the gravity vector on a RPM furthermore does not allow biological systems to adjust to gravity, so they respond similar like being exposed to actual microgravity. The results achieved by RPM exposure are indeed in a good-to-fair agreement to actual space-flown experiments of multiple cell types (reviewed in Wuest et al. [32]).
Parabolic- or sounding rocket flights are taking advantage of a “free fall” situation to generate a microgravity environment. The μg-periods on these platforms are, however, rather short (seconds to minutes). Depending on the initial speed of the aircraft, the μg period during a parabolic flight can be extended up to almost 1 min [33 –35]. Sounding rockets in contrast offer various microgravity periods. Depending on the apogee, 1 up to 12.5 min can be reached.
Formation of MCS
MCSs are cellular round 3D complex structures or aggregates formed by different cell types. When adherent cells are cultured in spinner flasks or on 96-well plates coated with agarose (liquid-overlay technique), or are engineered with the hanging drop technique and other methods, the cells are able to aggregate to round small spheroids which are floating in the culture medium [5,6]. These spheroids show dynamic cell–cell/cell–matrix interactions; the cells are connected through anchoring junctions and can communicate through gap junctions [1]. Spheroids show differentiated functions such as the synthesis of extracellular matrix (ECM) proteins, cell adhesion molecules, and cytoskeletal filaments [4,36]. It has been known for a while that a microgravity environment also induces human cells to aggregate and to grow three dimensionally. Several studies (Table 1) have demonstrated this in space and on Earth using the RWV, RPM, clinostat, and other devices [2,16 –18,25,37,38].
The 3D cell culture technique using stem cells is capable of maintaining the pluripotent capabilities of these cells and can thus improve the potential of developing methods triggering the induction of differentiation and, ultimately, the formation of organoids.
Kawahara et al. [39] showed that mouse embryonic stem cells (ESCs) could be maintained in feeder-free and serum-free culture conditions without leukemia-inhibitory factor through exposure to a 3D-clinostat. ES cells of the clinostat group aggregated to multiple small spheres after a 3-day exposure (Fig. 2) [39]. These free-floating spheroids grew larger in size over time (Fig. 2).
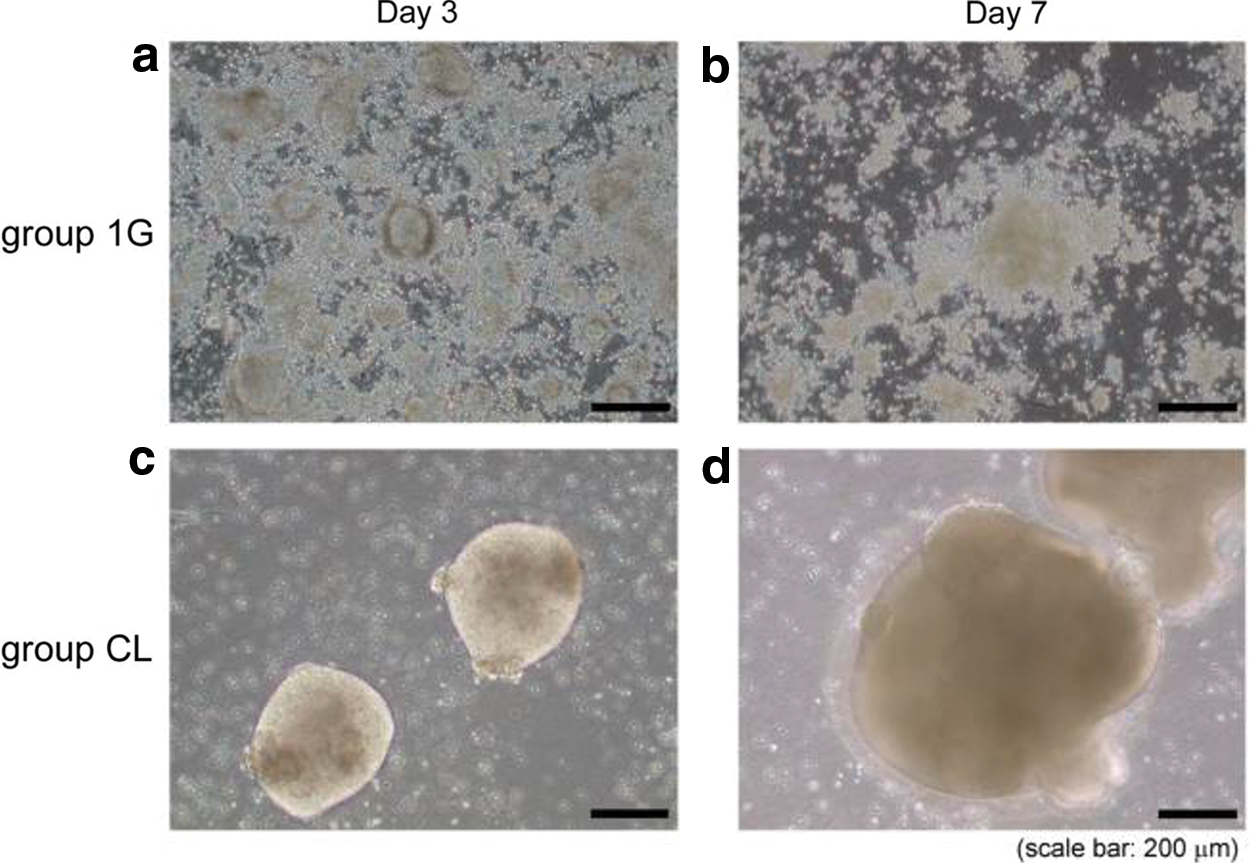
Morphological changes of cultured mouse ES cells on day 3 and 7. All cells elongated to an oval cell shape and flattened, displaying the phenotype of differentiated ES cells in group 1G
Stem cells cultured in s-μg were used for coculture studies in cancer research, for example (Table 2). Host-liver colorectal-tumor spheroids composed of primary human hepatocytes, MSC, and colon carcinoma HCT116 cells were formed using the RWV [23]. These engineered 3D organoids revealed a lower sensitivity to 5-Fluorouracil treatment [23].
Other scientists [40] studied the effects of s-μg created by a fast rotating clinostat on neuronal differentiation of human adipose-derived stem cells. The authors reported that microgravity had a positive effect on the neural differentiation potential of stem cells [40].
The 3D technology is widely used in Dentistry; there stem and progenitor cells can be established from periodontal tissues. For the differentiation of dental stem cells toward the osteoblastic/cementoblastic and neural lineage, the 3D technology is applicable in combination with an advanced protocol [41]. A recent publication reports about engineering of 3D micro nerve tissue using postnatal stem cells from human dental apical papilla [42]. Kim et al. [42] described a stem cell culture which was able to differentiate into neural cell lineages. They applied a modified protocol to engineer a 3D culture [42]. The 3D structures were formed by application of polyethylene glycol (PEG) microwell-mediated cell spheroid formation and subsequent dynamic culture in a HARV bioreactor [42].
Another study reported from murine bone marrow-derived stromal cells, grown as static suspension and 3D scaffold-free MCS, as well as aggregates on poly(lactic) acid microspheres [43]. Increased adipogenesis was found in 3D MCS cultured in a RCCS [43].
ES cells are an important in vitro model in the investigation of development or can be used to study Regenerative Medicine. Aggregation of ES cells is a requisite for the formation of 3D embryoid bodies (EBs) [44]. An STLV type bioreactor was used to engineer cardiac tissue. The authors demonstrated that EB efficiency in STLV was twofold higher than those of liquid suspension cultures [44]. The cardiomyocyte differentiation process of EBs was enhanced during the dynamic culture in STLV [44]. After 1 month of STLV exposure, EBs exhibited no large necrotic areas [45]. In contrast, after HARV-exposure massive EB aggregation was detectable, together with enormous cell death and large necrotic areas. In addition, indicators of differentiation were detectable at the edges of the cell clumps in HARV cultures [45].
The differentiation process of the EBs increased under the dynamic STLV exposure [45]. STLV delivers a fine technique for the initial control of the aggregation rate in human stem cells and induces the formation of small EBs with efficient productivity. At week 2, different tissues deriving from the three germ layers, vascular tissue, neuronal tubes, as well as glandular tissues, were detected in the differentiating EBs [45].
A recent publication reported about the generation of microscale progenitor cardiac spheres from human pluripotent stem cells [46]. The spheres were exposed to a RPM for 3 days during their differentiation to cardiomyocytes [46]. The authors showed that a combination of 3D culture and RPM exposure of the spheres is suitable for the efficient generation of highly enriched cardiomyocytes [46]. This technique greatly enhanced the proliferation and viability of cardiac progenitors [46].
A very interesting approach using RWV technology is to engineer a tubular structure (1 mm i.d.) from aligned human mesenchymal cell sheets as the wall and human endothelial progenitor cell (hEPC) coating as the lumen [47]. The tubular constructs demonstrated a comparable functionality to an animal vessel [47].
Tissue Engineering of Cartilage
Cartilage is a tissue with very limited potential for repair after sustaining damage in conditions such as arthrosis. It is therefore of utmost interest to produce cartilage in vitro, ideally from autologous patient cells, suitable for transplantation into the affected joints. Furthermore, it has been shown in human bed-rest studies, as well as in animal hind limb unloading experiments on Earth, that reduced biomechanical forces over a longer period of time can lead to proteoglycan loss in articular cartilage and to an average loss of cartilage thickness of about 8% [48 –51].
Over the years, several μg-based approaches of tissue engineering have been carried out to produce viable stable cartilage with comparable properties to in vivo tissue. Not surprisingly, only very few experiments were conducted under real μg (r-μg), during rare and costly spaceflight opportunities.
In one of these experiments, bovine chondrocytes were seeded into 3D polyglycolic acid matrices and cultured in rotating bioreactors in 1 g over the course of 3 months. One of the reactors was then transported to the MIR space station, where cultivation continued for 4 more months, while the remaining samples were incubated on Earth. Both conditions yielded functional cartilaginous constructs; however, those grown in space exhibited reduced mechanical stability and size [52 –55]. Similar results were found in a scaffold-free experiment using porcine chondrocytes. Cells were grown in special cylindrical culture chambers and exposed to r-μg on the ISS, to s-μg on a RPM, and to 1 g on Earth. The ISS samples showed less ECM with higher collagen II/I ratio, less aggrecan and versican expression, as well as lower cell density in general compared to the 1 g-controls [56,57].
Many more studies have been conducted on the different s-μg devices. Since the emphasis of this review lies on the use of stem cells, the following discussion will focus on experiments using these cells to engineer cartilage tissue.
Generally, it has been demonstrated that s-μg exposure is promoting the chondrogenic differentiation of bone marrow mesenchymal cells [58]. Ohyabu et al. have shown that rabbit bone marrow MSCs, first cultured in DMEM and then transferred into chondrogenic medium in an RWV bioreactor, were forming large cylindrical (1.25 × 0.6 cm height × diameter) cartilaginous tissue constructs without the necessity of a scaffold. The resulting tissue constructs exhibited collagen I and II, aggrecan, and glycosaminoglycan gene expression and collagen I/II, safranin-O, and toluidine stains similar to the control cartilage, whereas static cultures showed an inferior differentiation rate and cartilaginous properties [59]. Later, the same group refined their method using a collagen sponge as a scaffold to better control the shape of the generated tissue. Furthermore, they were able to show that this scaffold increased the tissue's mechanical stability and glycosaminoglycan content and considerably decreased the culture time necessary to obtain a fragment suitable for transplantation. This study did not include static controls, as they have been shown to be less effective before [60].
Human bone-marrow derived cells isolated from the iliac crest have yielded similar results when cultured in s-μg. First, cells were grown in a monolayer in DMEM for 2–3 weeks and then transferred into a RWV with chondrogenic medium for 2 additional weeks. A stationary pellet culture served as control. The RWV samples were generally larger than the control tissues (4.5 mm vs. about 1 mm in diameter) and yielded about 2.5-fold higher DNA content and about fourfold more glycosaminoglycans. In addition, histological and immunohistological analysis confirmed their hyaline cartilage [61].
Hyaline cartilage was also produced in a 3D clinostat experiment. Human MSCs cultured under s-μg conditions showed a higher proliferation rate, increased numbers of CD44/CD29 or CD90/CD29 positive cells, and increased expression levels of collagen II and aggrecan, compared to static controls. In addition, only the s-μg samples were able to form hyaline cartilage when transplanted into a cartilage defective mouse model [62].
In an effort to produce cartilage tissue for the reconstruction of complicated structures such as craniofacial defects, Takebe et al. used the recently developed pC-HAp/ChS scaffold, a porous material consisting of collagen, hydroxyapatite, and chondroitin sulfate. Cartilage progenitor cells from human elastic cartilage, obtained from microtia patients, were first grown as monolayer cultures, then seeded into the scaffold and cultured in a RWV in differentiation medium for 6 weeks. The resulting products retained the shape of the scaffold, revealed cartilage-like appearance and morphology, and histochemical analyses detected proteoglycan and elastic fibers unique to elastic cartilage [21].
Another interesting scaffold was introduced by Yin et al., when cartilage ECM-derived particles (CEDPs) were cocultured with bone mesenchymal stem cells in a RWV bioreactor. It was shown that the cells were of high viability and rapidly proliferated on the particle surface. Moreover, after 21 days of culture the cells had matured into chondrocytes, and the resulting tissue constructs were superior in their joint recovery and cartilage repair potential in a rat compared to CEDPs alone on fibrin glue. It is also noteworthy that no exogenous TGFβ was necessary to initiate stem cell differentiation in the presence of CEDPs [63]. This finding was also described by Luo et al., who differentiated MSCs into a nucleus pulposus–like phenotype using pellet culture in a HARV bioreactor. They, too, observed that spontaneous cell differentiation in s-μg occurred in absence of external TGFβ [64]. However, controversy over the role of TGFβ still remains. Other researchers have reported that TGFβ increases the collagen II and proteoglycan synthesis and promotes chondrogenesis in a synergistic manner together with μg [65,66].
Overall, s-μg seems to be a promising condition to produce viable functional cartilage, of generally better quality than tissues grown under static 1 g-conditions. Furthermore, s-μg facilitated the even colonization of scaffolds with MSCs during cell culture, ensuring the production of transplants with defined 3D shapes. Due to the tendency of the cells to sediment, comparable results cannot be achieved with conventional static 1 g culture conditions, which make the use of s-μg a promising technique for cartilage tissue engineering. Finally, for applications not requiring cartilage tissue of predetermined conformation, further research might lead to the refinement of scaffold-free approaches, avoiding potential pitfalls associated with nonautologous biomaterials.
Tissue Engineering of Bone
As cited in preceding chapters, microgravity influences several human organ systems. Depending on the affected organ system, alterations, as well as pathologies, are caused by prolonged and short-term exposure to a microgravity environment [67]. Bone loss in space or microgravity-induced osteopenia is especially problematic for extended space missions with included planet exploration. Astronauts lose between 1% and 2% of weight-bearing bone mass during long-term missions [68]. Therefore, deterioration of bone structure due to exposure to microgravity is an essential concern for space agencies today [69,70], as bone loss and radiation are the most substantial health risks astronauts face [71].
Despite this knowledge, it is interesting that the effects of simulated microgravity on bone cells rather lead to the construction of bone. The research area of bone tissue engineering is currently a hot topic in gravitational biology and space medicine. The first results in this field were demonstrated in 1998 [72]. Rat marrow stromal cells on Cytodex-3 microcarrier beads revealed after a 14-day RWV exposure spherical aggregates showing mineralization, alkaline phosphatase activity, as well as collagen type I and osteopontin expression.
The NASA-developed RWV was used in various investigations for bone tissue engineering purposes. Osteoblasts or MSCs were cultured on several scaffolds like bioactive glass-polymer composites, bioderived bone scaffolds, alginate or gelatin, and others. The RWV-derived tissue constructs were similar to natural bone and improved compared to tissue engineered constructs deriving from 1 g approaches. RWV-derived tissue constructs revealed a better in vivo effectiveness in bone repair in animal studies [73 –82].
Therefore, several studies investigating the influence of microgravity have been conducted on this account [25,82]. As reviewed by Han and Dai, microgravity could support future organ or tissue engineering. The authors note stem cell proliferation and differentiation as most critical issues. A large-scale stem cell expansion, which would subsequently allow for determined differentiation is preferable [83]. As published by Huang et al., MSC differentiates differently depending on shifts in gravity. Accordingly, hypergravity leads to a commitment toward osteoblasts and myocardial cells, whereas microgravity causes differentiation toward an adipose cell fate instead [84].
In addition, Zayzafoon et al. observed increased production of PPARy2, that is, a marker for adipogenic differentiation, as well as decreased markers for osteogenic differentiation, for example, RUNX2 under s-μg [85]. In addition, human MSCs (hMSCs) cultured for 7 days on a 3D clinostat showed increased proliferation and an expanded hMSC population in comparison with 1 g controls. Furthermore, cells cultured under s-μg from the same study were xenotransplantated successfully and showed hyaline cartilage differentiation [62].
Finally, Clarke MSF, Sundaresan A, and Pellis NR developed a method to obtain bone constructs by culturing osteoblasts and osteoclast precursors under randomized gravity vector conditions. Generally, the randomized gravity vector conditions are obtained using a low-shear stress rotating bioreactor, such as a HARV culture system [86,87]. Tissue engineering of bone constructs on the RPM and on the RWV using fetal osteoblasts is demonstrated in Fig. 3. Moreover, initial preliminary approaches of exposing hMSC to the RPM for 7 days revealed the formation of small aggregates (Fig. 3). These studies are currently ongoing.
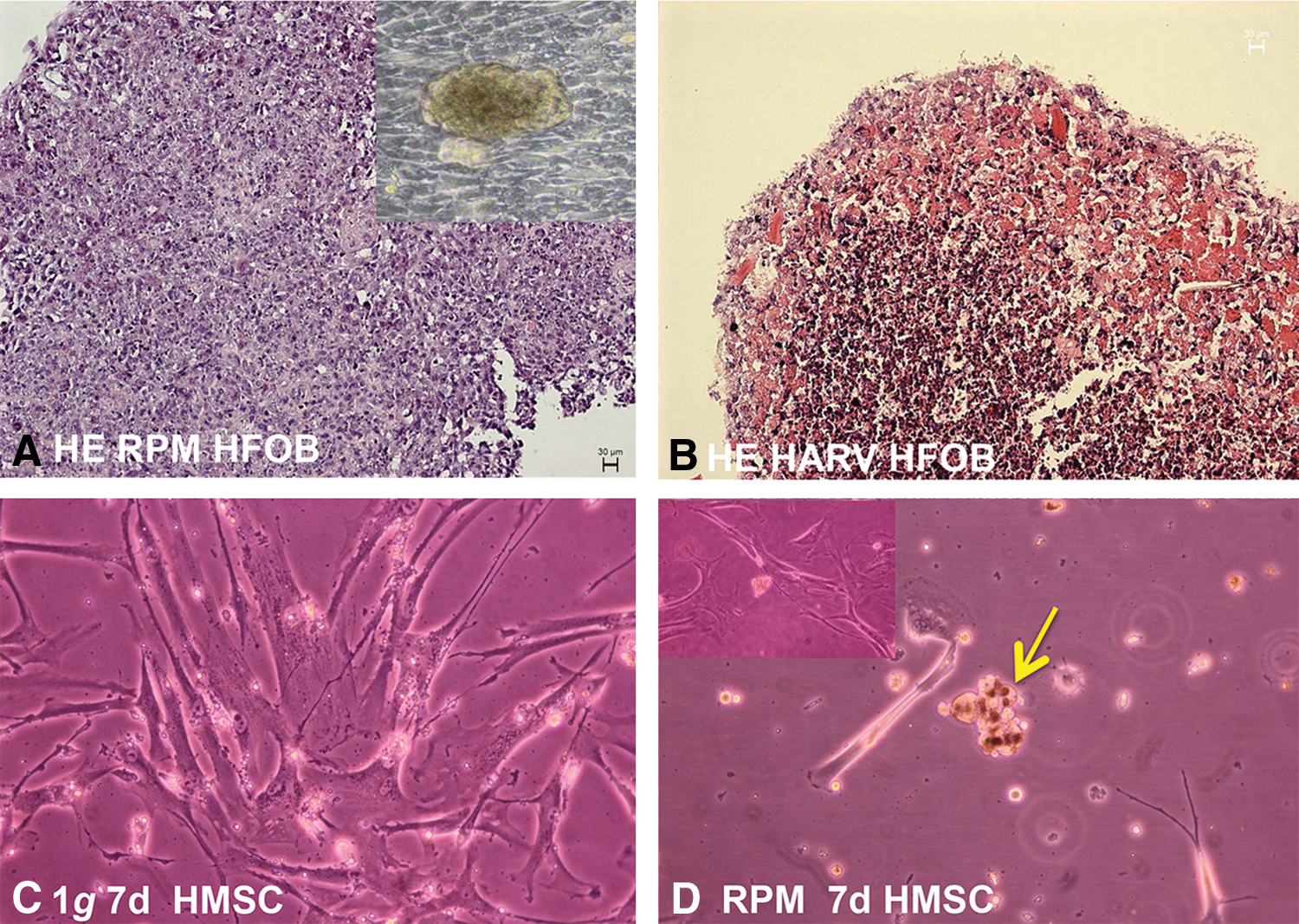
As displayed above, intriguing new developments have been observed in the field of bone tissue engineering in microgravity, and controversial findings have to be assessed by new studies. Due to the huge impact of musculoskeletal medical conditions on Earth and for future space exploration, further studies on the influence of microgravity in bMSCs are of paramount importance for this promising subject.
Construction of Artificial Vessels
Tissue engineering has become a promising approach for generating biocompatible vessels, which can be used in Regenerative Medicine, that is, for vascular reconstruction in transplantation medicine or for plastic surgery purposes (Fig. 4). A key benefit is not to have to introduce any foreign materials to the human body (Table 1).
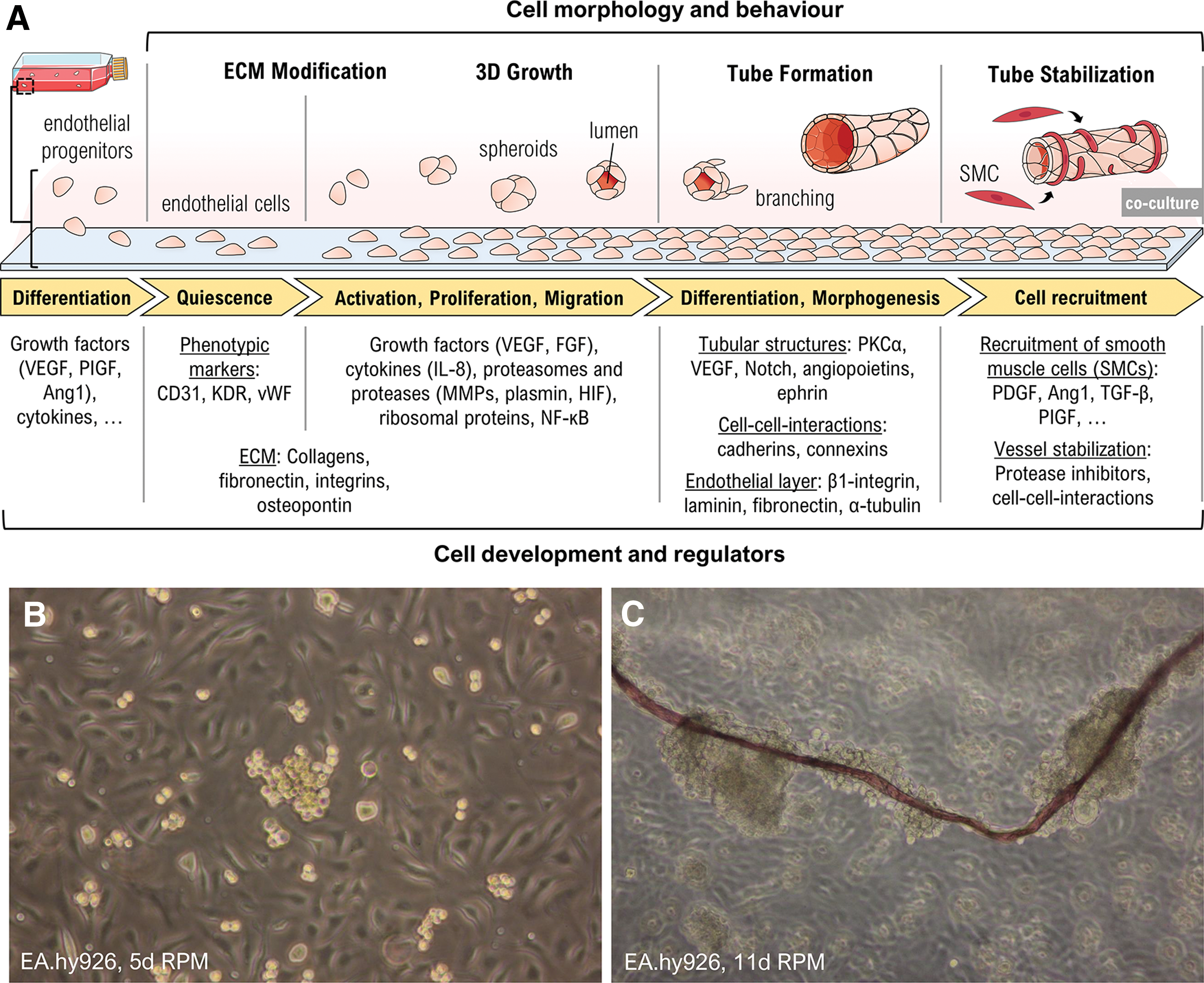
Engineering of vessel-like tissue constructs using the RPM.
Under normal gravity (1 g) conditions, ECs forming the inner surface of blood vessels grow in monolayers, which do not fully exhibit the vascular characteristics found in vivo. Culturing ECs with other cells was an initial promising approach [88]. A breakthrough for vascular engineering was reached later, when seeding functional cells on biodegradable scaffolds, which determine the shape and contribute to the mechanical stability of the generated vessels [89 –92]. However, choosing the right scaffold material is crucial for efficient vascularization, as well as to avoid immunologic problems, a distorted structure of the newly formed vessel or an altered mechanical resilience compared to natural tissues [2].
In the search for the “Holy Grail” of vascular engineering, the de novo formation of scaffold-free, functional, organo-typic tissue constructs, gravitational biology offers a big advantage, as ECs grow as spheroids and form 3D tubular structures under s-μg conditions without requiring any artificial matrix or scaffolds [2,93]. Such aggregates were first generated by Sanford et al. within 30 days in an RWV [94]. In this study, ECs grew as multilayer sheets of cells around Cytodex-3 microcarrier beads and showed increased nitric oxide production and enhanced barrier properties, possibly caused by an upregulation of junctional complex proteins.
Based on this experiment, Chiu et al. cultured CD34-positive human cord blood stem cells, which are involved in vascularization, in RWVs with or without Cytodex-3 microcarrier beads and supplemented them with vascular endothelial growth factor (VEGF) [95]. While after 4 days the cells growing on beads only formed amorphic cell clusters, the cells cultured without beads developed vascular tubular assemblies and exhibited endothelial phenotypic markers such as CD31 and KDR (Flk1). Although this observation was unexpected, the data suggested that CD34+ human umbilical cord blood progenitors are capable of differentiation into a vascular endothelial phenotype and able to assemble into 3D tissue structures in s-μg. Some years later Zhang et al. reported that clinorotation of MSCs also leads to enhanced transdifferentiation into an endothelial phenotype by upregulating the expression of endothelial-specific molecules (KDR and vWF) and enables cells to form a capillary network [96]. In cardiovascular progenitors, these effects of s-μg are apparently age dependent. Adult cells showed elevated expression of endothelial markers, whereas neonatal cells acquired characteristics of dedifferentiation [97] (Table 2).
An often used model for vascular engineering is the immortalized EC line EA.hy926, a fusion of human umbilical vein ECs with a thioguanine-resistant clone of A549 adenocarcinoma cells [98]. Infanger et al. used the RPM for culturing EA.hy926 cells with and without supplemented VEGF in the growth medium [93]. During the 72-h experiment they observed an initial upregulation of ECM proteins induced by both, RPM exposure and VEGF alone. They also found that RPM exposure induced apoptosis in some of the cells beginning from 4 h culture time and increasing until 72 h, while VEGF could reduce the apoptosis rate. After 72 h many nonapoptotic cells had formed tube-like structures. To characterize this tube formation it could be observed that adherently growing EA.hy926 cells cultured on an RPM began to form small colonies by spreading over neighboring cells. These colonies were the origins from where tube-like structures developed and detached after 2 weeks of cultivation. They formed a defined lumen and continued to elongate during 2 more weeks on the RPM.
Interestingly, the tube walls seemed to resemble vascular intimas (Fig. 4): they consisted of a single cell layer, which produced more β1-integrin, laminin, fibronectin, and α-tubulin than cells cultured under 1 g conditions [99]. The alterations of cell physiology are thought to be mediated through cytoskeletal changes induced by gravitational unloading, which also impacts viscoelastic properties of the cells [100 –102].
In a first attempt to elucidate the mechanism of tube formation, transcriptome and proteome analyses were performed. Based on the gene and protein expression patterns, the authors deduced that phosphokinase Cα seems to play a key role in vasculogenesis [103]. Increased levels of angiogenesis are mediated through PI3K-Akt-eNOS signal pathways [104]. Whole genome expression analyses have identified an interaction network of further 27 genes whose expression levels were changed in RPM-treated cells. These genes are involved in signal transduction, angiogenesis, cell adhesion, membrane transport, or serine biosynthesis [105] and could be promising targets for future research on endothelial tube formation.
Protein comparison of adherent EA.hy926 cells and their 3D aggregates, both harvested from the RPM, revealed elevated ribosomal proteins during tube formation [106]. The analysis of EA.hy926 cells together with human microvascular ECs showing fast-developing tubular structures during RPM exposure finally confirmed that the regulation of proteasomes and ribosomal proteins is involved in the tube formation and that NF-κB activity, as well as IL-8, may play a decisive role for 3D growth of ECs [106].
In summary, the specific s-μg culture conditions by ground-based facilities offer a unique opportunity to study the mechanisms of 3D tissue development and provide a very promising method for vessel engineering [25]. First coculture experiments of ECs with vascular smooth muscle cells and fibroblasts on an RPM made the first step in engineering a complete functional vessel [2].
Eye Tissue Engineering
To restore and replace damaged ocular tissue and, hence, improve treatment of serious eye diseases, development of ocular tissue engineering applications is vital. Even though tissue engineering approaches for ocular tissues have indeed come a long way over the last decades, scientists are still not able to provide answers to overcome principal hurdles, strongly advocating for further expansion of eye tissue engineering.
In support of this conception “The National Eye Institute of the US National Institutes of Health” (NIH) has recently launched a competition to develop a working model of the human retina from stem cells—entitled, the 3D Retina Organoid Challenge [107]. As disturbances of visual function are considered to be a major complication of spaceflights, NASA has in parallel announced the eye as an important focus area. Further to this, microgravity conditions aboard the ISS are now investigated as an optimal environment for studying and expanding, for example, stem cells. Collectively, these interesting initiatives will help to clarify the mechanisms of retinal disease, stimulate new technologies based on, for example, tissue engineering and develop more effective therapies.
While a growing body of knowledge and experience has been obtained from experiments in cells, animal models, and surgical procedures in humans (see recent review by Karamichos [108]), only a limited number of studies have focused on investigating ocular tissue engineering approaches in microgravity. Hence, to provide a status, including future directions of ocular tissue engineering in microgravity, the recent findings and challenges in this field—both at normal and annulled gravity—will be discussed.
Ocular tissue engineering in normal gravity
Until now tissue engineering for the retina involves mainly cell- and scaffold-based approaches in animal models. Transplantation of autologous, but not homologous, retinal pigment epithelial (RPE) cells results in improved vision [108,109]. Engineering of primary pigment epithelial cells (PECs), expressing antiangiogenic factors, such as pigment epithelium derived factor (PEDF), has been pursued. Recently, Thuman et al. demonstrated stable integration and overexpression of PEDF in RPE cells using a Sleeping Beauty transposon system [110]. Transplantation of such engineered RPE cells may be an important contribution toward development of novel treatment strategies fighting RPE cell death and vision loss related to exudative age-related macular degeneration [110].
Other researchers elegantly generated and isolated photoreceptor precursors for transplantation from mouse ESCs and showed that rod precursors integrate within degenerated retinas of adult mice and mature into outer segment-bearing photoreceptors [111]. These studies show conclusively that clever isolation of ESCs can provide an important source of photoreceptors for retinal cell transplantation (Fig. 5). Although these findings have verified the potential of cell-based therapies to renew the degenerating retina, they also revealed that introduction of stem and/or progenitor cells to the subretinal space may result in both disorganized and poorly localized grafts [112]. As an alternative, application of biocompatible polymers that gradually degrade has successfully shown that donor cells were able to produce their own ECM, and subsequently, cell-polymer grafts generated healthy donor tissue (Fig. 5).
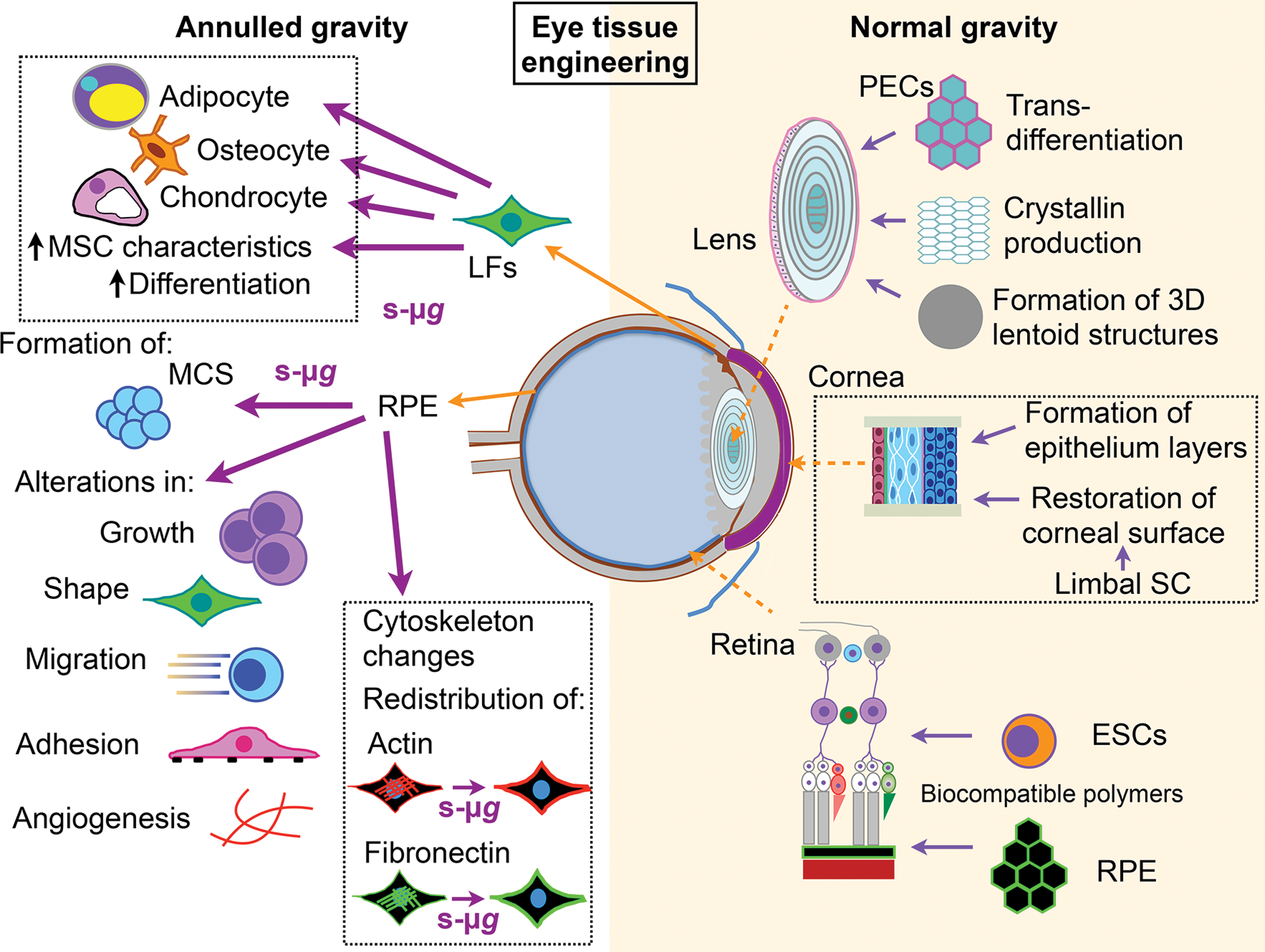
Eye tissue engineering: schematic representation of current attempts to engineer ocular tissue under simulated micro- (left side) and normal gravity (right side). Under reduced gravitational conditions, important information regarding enhanced differentiation and efficient formation of cells with MSC characteristics was obtained by subjecting LFs to s-μg. Such experiments resulted in generation of chondrocytes, osteocytes, and adipocytes. Regarding RPE cells, formation of MCS and alteration of the cytoskeleton, including significant cellular redistribution of actin filaments and fibronectin, were demonstrated following incubation at s-μg. Notably, s-μg results in an alteration of growth, shape, migration, adhesion, and angiogenesis. Under 1 g conditions efforts have been focused on lens, cornea, and retina. Some of the highlights are the lens transdifferentiation into 3D structures resembling lentoids and the formation of crystallin. In corneal tissues, formation of epithelium layers and restoration of corneal surface have been obtained. In retinal tissue engineering ingenious isolation of ESCs may provide an important source of photoreceptors for retinal cell transplantation. Similarly, transplantation of engineered RPE cells may be an important contribution toward the development of novel treatment strategies for fighting RPE cell death. For details see text. LFs, limbal fibroblasts; MCS, multicellular spheroid; PECs, pigmented epithelial cells, RPE, retinal pigment epithelial; SC, stem cell; s-μg, simulated microgravity. Color images available online at
Development of alternatives for cataract surgeries may in some case be beneficial. One of the most compelling findings was presented by Tsonis et al. [113]. In this study lens differentiation was investigated by analyzing transdifferentiation in PECs [113]. Notably the authors showed the ability of these cells to induce synthesis of crystallin and to morphologically differentiate into 3D structures resembling lentoids in vitro, thereby paving the way to define crucial elements involved in lens transdifferentiation from PECs [113] (Fig. 5).
Corneal epithelium layers have also been established in vitro with great success using cell-based [114,115], membrane-based [116,117], or scaffold-supported approaches [116,118] (Fig. 5). Furthermore, engineered epithelial tissue sheets, established by the use of polymeric substrates, have been clinically evaluated using autologous oral mucosal epithelium [114]. However, a major challenge for corneal epithelium layers established in vitro is the lack of structural stability. Studies of limbal stem cells also provide hope for cell-base restoration of corneal surfaces [115]. In addition, tissue grafts, mainly based on amniotic membranes, have been established and applied with some success in humans [119]. Notably, the utilization of a 3D tissue engineering cornea in the context of in vitro tissue models has been extensively studied to recapitulate the three layers of the cornea [116] (Fig. 5).
Ocular tissue engineering in microgravity
Various medical therapies rely on human stem cells. The success of such treatment strategies requires efficient expansion of human stem cells. Presently, scientists have no efficient way to do this, but stem cell expansion might accelerate in microgravity.
A recently commenced initiative by NASA—entitled “Microgravity Expanded Stem Cells”—will investigate the possibility of cultivating human stem cells aboard the ISS for use in clinical trials to evaluate their use in treating disease. The results may also advance future studies by providing new knowledge on how to scale up expansion of stem cells (
MSCs are characterized by self-renewal, as well as by multidirectional differentiation [120,121]. As mentioned above, stem cells originating from the corneal limbus show promise in treating corneal blindness [115]. Interestingly, limbal fibroblasts (LFs) have been shown to hold MSC characteristics [122] and can differentiate into other cell types [123]. In a recent study by Pao et al. the effect of microgravity on the proliferation and differentiation of LFs was investigated with the aim of identifying in vitro culture condition providing high proportion of LFs possessing MSC characteristics [124] (Fig. 5). Cells were cultured in a rotary culture system to simulate microgravity. Using this system, the Pao group [124] showed significantly reduced proliferation rates at s-μg compared to 1 g. In parallel, higher proportions of the cells expressed CD90, CD105, and SSEA4 under s-μg than under 1 g, and the differentiation potential was more distinct at s-μg conditions. Collectively, these findings show that during s-μg conditions, LFs displayed a higher fraction of MSC characteristics and were easily induced to different lineage cells. Hence, the obtained results from this study may help to further optimize conditions for eye tissue engineering based on corneal stem cells.
It is a well-known fact that exposure to microgravity conditions, for example, during a long-term spaceflight, has huge impact on ocular changes, including choroidal folds, optic disc edema, and globe flattening, and hyperopic shifts are considered as a high human health risk of spaceflights [125 –127]. Notably, two recent articles have focused on explaining the etiology of these spaceflight-induced effects [128,129]. In addition, previous findings have found evidence showing that the microgravity environment disturbs retinal development, including loss of RPE cells [130], and induces inflammatory response [131]. Another recent article further investigated the effect of annulling gravity in RPE cells by exposing human adult retinal epithelial cells (ARPE-19) to a RPM [132]. The findings from this study clearly show that RPM exposure induces significant changes in the F-actin cytoskeleton and cytoskeleton-related proteins in ARPE-19, in addition to cell growth behavior and gene expression patterns involved in cell structure, growth, shape, migration, adhesion, and angiogenesis (Fig. 5). Surprisingly, a minor fraction of the ARPE-19 produced MCS following incubation in s-μg conditions implying that annulling gravity promotes 3D growth of ARPE-19 in vitro. The authors hypothesized that weakening of interior and exterior cellular structures of the RPE cells following exposure to microgravity may affect the function of the retinal cells [132].
Even though it is clear that microgravity conditions improve obtainability of cells with stem cell characteristics and promote 3D growth of corneal cells, further studies are clearly required to investigate whether reduced gravity—either in form of real or simulated conditions—is the missing link that will further improve conditions for reconstructive therapy for patients with corneal disease, as well as improve our knowledge of eye tissue engineering based on corneal stem cells.
In summary, these studies are the first steps toward the development of human tissue constructed by the use of microgravity simulation techniques. Future studies will determine the impact of microgravity on applications that may benefit the field of Regenerative Medicine.
Conclusions
The field of tissue engineering has achieved great advances in constructing 3D tissues and spheroid or organoid structures from specialized cells or stem cells with and without biomaterials using microgravity. Growing tissues in space and on Earth using microgravity conditions is currently a hot topic in Biomedicine and Regenerative Medicine [2,25]. Stem cells have been investigated in space on the MIR and the ISS. The ISS project “Space Tissue Loss - Stem Cell Regeneration (STL-Regeneration)” examined the effects of spaceflight on the biology of murine stem cell differentiation (Principal Investigator (PI) Eduardo A. C Almeida, Ph.D., Moffett Field, CA, United States). The results of this mission were published in reference [133] and showed that exposing stem cells to microgravity inhibits their differentiation and preserves stemness [133]. Another study “Microgravity Expanded Stem Cells” is currently under investigation (PI Abba Zubair, Mayo Clinic in Jacksonville, Florida). The investigators report that stem cell expansion may accelerate in microgravity [
In addition, a Japanese ISS project “Study on the Effect of Space Environment to Embryonic Stem Cells to Their Development (Stem Cells)” (PI Takashi Morita, Osaka City University, Osaka, Japan) was performed. The scientists examine the effects of space radiation on murine ESCs [134].
As space missions are rare and costly, ground-based studies are of hightened interest. In addition, it has been known for a long time that many types of differentiated cells grow three dimensionally on the RWV [16].
Furthermore, the ideal media compositions (growth factors and cytokines, for example) to initiate tissue structures and cellular identities remain to be studied. Stem cell differentiation and the development of 3D tissues are dependent on how gases, nutrients, and signaling factors diffuse within cultured tissue constructs [135] and on the best method for 3D aggregation.
This review describes various attempts to use real microgravity and devices constructed to simulate microgravity conditions on Earth for purposes of tissue engineering. We addressed the suitability of the use of stem cells to engineer 3D constructs under low-gravity conditions. In general, stem cells have the huge advantage over matured cells that they can differentiate into a multitude of different cell types, thus allowing the production of a large variety of patient autologous tissues even for those that cannot be directly obtained from an organ source. Furthermore, also in contrast to differentiated cells, they can be cultured for a very long (theoretically indefinite) time, which makes them the ideal tools for Regenerative Medicine. Promising results were found for the construction of 3D structures from CD34+ human umbilical cord blood progenitors in s-μg [95]. Interesting findings have been published about cartilage and bone tissue engineering using the RWV.
Controversial findings concerning bMSC in microgravity have to be assessed by subsequent studies. The dynamic STLV technique was revealed to be suitable for the initial aggregation rate control in human stem cells and for the induction of small EBs with efficient productivity [45]. Compared to real microgravity in space, simulated microgravity seems to offer some advantages for stem cell culture and differentiation. Due to the fact that—especially in the RWV-type bioreactors—the cells are in permanent free fall through a constantly flowing medium, nutrient and growth factor availability is not limited by diffusion rates. Furthermore, the associated mechanical challenge also seems to have a positive influence on cell differentiation rates and tissue stability, as described, for example, for cartilage. Collectively, these different protocols and s-μg studies will therefore increase the current knowledge in Regenerative Medicine and Space Medicine. They project significant impact on various diseases, in particular, on cancer, cardiovascular diseases, and ocular, cartilage, and bone disorders. However, more research in this field is required to bring this technology closer to clinical application.
Footnotes
Author Disclosure Statement
The authors declare no potential conflicts of interest with respect to the research, authorship, or publication of this article. No grant funding was provided for this work.