
Abstract
Human parthenogenetic embryonic stem cells (hPESCs) represent a source of histocompatible tissues for transplantation and carry two copies of the maternal genome, but lack the paternal genome. In this study, we selected 63 known human imprinted genes to investigate the imprinting status of hPESC. The expression level of these genes, including 27 maternally and 36 paternally imprinted were illustrated in hPESC and human embryonic stem cells (hESCs) derived from fertilized embryo lines. The expression activity changes of these genes were analyzed in undifferentiated and early differentiated hPESC lines. In addition, the methylation status of four differentially methylated regions (DMRs) of the imprinted genes was analyzed in undifferentiated and early differentiated hPESC and hESC lines. As a result, we found that all the maternally imprinted genes were expressed at similar levels in the undifferentiated hPESC lines and the hESC lines, except ZNF264 and ATP10A. Twenty-one analyzed paternal imprinted genes were expressed at the same level in two separated hPESC lines as well as compared with the hESC lines, whereas 15 other paternal imprinted genes were significantly downregulated or inactivated in hPESC lines as compared with the hESC line. During prolonged passage, the expression levels of the majority of imprinted genes remained stable in two hPESC lines. The four DMRs, including PEG3/ZIM2 (DMRs), SNURF/SNRPN DMRs, and KVDMR1 DMRs are highly methylated in the genes of two undifferentiated hPESCs and its embryonic bodies (EBs), whereas the genes of the undifferentiated hESCs and its EBs are half methylated. During the early differentiation stage, the imprinted genes showed the same expression trend and the expression levels of H19, IGF2, SLC22A2, SLC22A3/SLC22A18, and CPA4 were significantly upregulated in both hPESC lines. As conclusion, hPESCs show a substantial degree of epigenetic stability with respect to some imprinted genes.
Background
H
The main concern about the future application of hPESCs is their imprinting status. hPESCs carry two copies of the maternal genome, but lack the paternal genome. This phenomenon might lead to abnormal expression of the imprinted genes. Genomic imprinting, which is an epigenetic phenomenon, describes the monoallelic expression of a small subset of genes according to their parental origin [3]. Inappropriate or disrupted expression of the human imprinted genes might alter the amount of the corresponding gene products and involved in the development of some clinical syndromes, including tumorigenesis [4,5].
Although PESCs could be isolated from parthenogenetic blastocysts of mice [6], rabbits [7], buffalo [8], nonhuman primates [9], and humans [10,11], the knowledge about the imprinting status of the PESCs is still limited. In mouse PESCs, the expressions of imprinted genes have shown enormous variation during prolonged passages [12].
The allele-specific expression of imprinted genes is generally regulated by DNA methylation. Most DNA methylation occurs within differentially methylated regions (DMRs). The methylation status of the DNA derived from specific parental origin within DMRs controls the transcription of the imprinted genes and allows discrimination between the paternal and the maternal alleles [13]. It has been reported that some paternal imprinted genes are activated due to the loss of methylation imprints within the DMRs of mouse PESCs or their derivatives [14]. Horii et al. showed epigenetic changes of the imprinted genes that lead to increased expression of U2af1 and SNRPN. Analysis of the rhesus monkey PES, all maternally and at least some paternally imprinted genes have been shown to be activated [15].
In this study, we used undifferentiated hPESC lines at different passages, early differentiated hPESCs, and two human embryonic stem cells (hESCs) from fertilized embryo lines. The expression levels of 63 selected human genes that are known to be imprinted within these cells were measured and compared with each other. In addition, we investigated the DNA methylation patterns of these genes to provide insights into the regulation mechanism of the expression of the imprinted gene.
Materials and Methods
Human material and ethics statement
The design and conduct of this study conformed to the ethical standards of the Helsinki Declaration and our national legislation. This study was approved by the Medical Ethics Committee of The First Affiliated Hospital, Sun Yat-sen University.
Culture and early differentiation of hPESCs
hPESCs and hESCs were plated on mitomycin-C (Sigma) irradiated mouse embryonic fibroblast cells and cultured in hESC culture medium consisting of KnockOut Dulbecco's modified Eagle's medium (Gibco), 20% Serum Replacement (Gibco), 1 mM glutamine (Sigma), 1% nonessential amino acid (Gibco), 0.1 mM 2-mercaptoethanol (Sigma), 50 IU/mL penicillin (Sigma), and 50 IU/mL streptomycin (Sigma) mixture as previously described1. Both cell lines were cultured in 5% CO2 at 37°C in humid conditions and the culture medium was changed daily. Embryonic stem cells were passaged by enzymatic dissociation using Collagenase IV (1 mg/mL) (Gibco).
For the collection of embryonic bodies (EBs), undifferentiated hPESCs were cultured in the medium consisting of Dulbecco's modified Eagle's medium (Gibco) supplemented with 20% KnockOut Serum Replacement (Gibco) and 0.1 mM nonessential amino acids (Gibco), 1 mM
Selection of imprinted genes for detection
For the selection of imprinted genes, we analyzed the public online data, including those from
Quantitative real-time reverse transcription–PCR
Total RNA was extracted and digested with DNase using the RNeasy Mini Kit (Qiagen). Aliquots of 1.5 μg of total RNA were reverse transcribed (RT) into cDNA using SuperScript III Reverse Transcriptase (Invitrogen) and oligo(dT)18 (Promega). For quantitative real-time PCR (qPCR), RT2 Profiler Custom PCR Array (SuperArray Bioscience), containing 63 custom-imprinted genes and 4 housekeeping genes, was used to analyze the global profile simultaneously following the instructions of the manufacturer.
Amplification data were collected by an ABI PRISM 7900 Sequence Detection System (Applied Biosystems) and analyzed with the Sequence Detection System 2.0 software (ABI). Data were calibrated to the average mRNA expression levels of glyceraldehyde 3-phosphate dehydrogenase (GAPDH), beta 2-microglobulin (B2M), hypoxanthine ribosyltransferase (HPRT1), and ribosomal protein L13A (RPL13A). RT-qPCR was also carried out using total RNA extracted from the mouse feeder cells to confirm the absence of reactivity with mouse cDNA.
Methylation analysis by bisulfite sequencing
Genomic DNA (gDNA) was extracted from the hESC lines and EBs using the DNA Extraction Kit (Invitrogen) in accordance with the manufacturer's instructions. Aliquots of 200 ng of gDNA were modified by bisulfite treatment using the MethylCode Bisulfite Conversion Kit (Invitrogen) in accordance with the manufacturer's instructions. The used primers are listed in Table 1. The PCR was carried out in a volume of 50 μL, which contained 1.5 mM MgCl2, 0.2 mM dNTP mixture, 0.2 M of each primer, 200 ng of template DNA, and 3 U of Platinum Taq DNA polymerase (Invitrogen).
DMR, differentially methylated region; F, forward; R, reverse.
Amplicons were electrophoresed through 1.6% Tris–Borate–EDTA agarose gels, stained with Ethidium Bromide, and visualized on a UV transilluminator. PCR products were recovered from stained gels using the PureLink Quick Gel Extraction Kit (Invitrogen), ligated with the vector pMD-18T (TaKaRa), subsequently transfected into the Escherichia coli according to the manufacturer's instructions. Individual colonies were transferred to LB/Amp medium and cultured overnight with shaking.
The cultures were processed with the PureLink Quick Plasmid Miniprep Kit (Invitrogen) in accordance with the manufacturer's protocol and each culture yielded plasmids containing a single PCR product. Using BigDye Terminator, individual clones were then directly sequenced with the M13R(−48) primer on an ABI 3730 Genetic Analyzer (Applied Biosystems). The sequencing results were analyzed using Sequencher software (Gene Codes Corp, Ann Arbor, MI).
Statistical analyses
Student's t-test or analysis of variance was used for the analysis of gene expression. Pearson's or Spearman's correlation coefficient test was used to analyze the correlations among levels of gene expression and passage number. Pearson's chi-square test was used for the analysis of expression ratios. A P value of <0.05 was considered significant. All statistical analyses were performed with Excel X and SPSS15.0.
Results
Selection of maternally imprinted genes and paternally imprinted genes
A total of 63 human imprinted genes were selected to represent the imprinting status of hPESC lines in this research. Among these genes, 27 were maternally imprinted genes and 36 were paternally imprinted genes (Supplementary Table S1; Supplementary Data are available online at
Summary of the expression status of imprinted genes in undifferentiated hPESCs and hESCs
Comparison of globally imprinted gene expression in undifferentiated hPESCs and hESCs
All 63 imprinted genes could be detected in two undifferentiated hESC lines and no differences were found for any of these genes between the two hESC lines (P > 0.05).
For further comparison between hPESCs and hESCs, the average expression level of the imprinted genes in two hESC lines was used. The expression pattern of the imprinted genes within the two hPESC lines was almost the same. Expression of all 27 maternal genes could be detected in both hPESC lines. Twenty-five maternal genes maintain normal expression levels, whereas 2 maternal genes showed a significant increased expression level as compared with hESCs.
The expressive proportions of the paternally imprinted genes in hPES1 and hPES2 were similar: the expression of 29 genes was detected (80.5%), whereas 7 (19.4%) were not detectable in the hPESC. Among the 29 imprinted genes, the expression level of 8 (22.2%) in hPES1 and 9 (25.0%) in hPES2 was dramatically reduced as compared with the expression level in hESCs. The remaining 21/20 (58.3%/55.6%) paternally imprinted genes showed a similar expression pattern in hPES1 and hPES2 cell lines (Fig. 1).
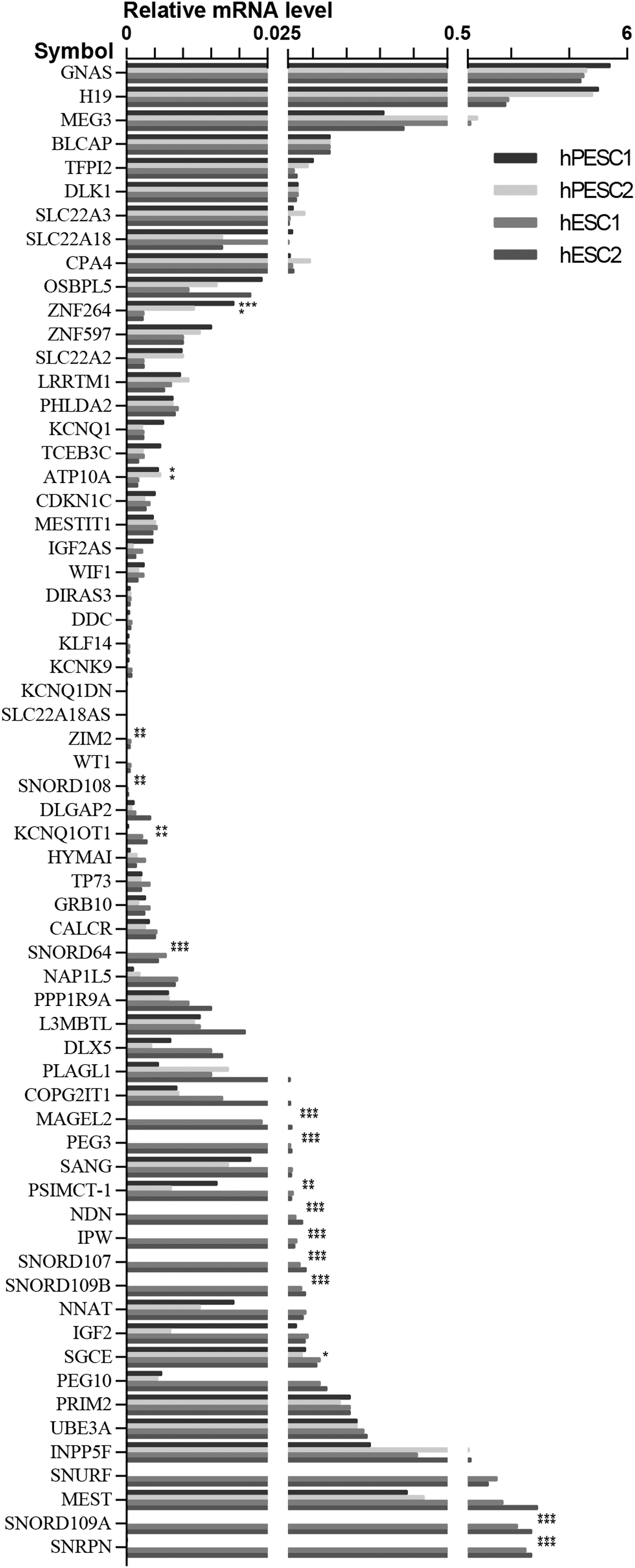
Comparison of globally imprinted gene expression in undifferentiated hPESCs and hESCs; no differences were found for 63 imprinted genes between the two hESC lines (P > 0.05). Expression level of all 27 maternal genes and expression of 29 paternal genes were compared in undifferentiated hPESCs and hESCs. X-axis is imprinted genes, Y-axis is the relative expressed level of imprinted genes *P < 0.05, **P < 0.01, ***P < 0.001. hESCs, human embryonic stem cells from fertilized embryos; hPESCs, human parthenogenetic embryonic stem cells.
Comparison of the expression level of maternal genes in undifferentiated hPESCs and hESCs
The majority of the 25 maternally imprinted genes showed a similar transcription activity between undifferentiated hPESCs and hESCs, and only ATP10a and ZNF264 demonstrated a modulated expression activity in hPESCs as compared with hESCs. The expression of the ATP10A was significantly upregulated by 2.7- and 2.9-folds in hPES1 and hPES2, respectively, whereas the expression of the ZNF264 was induced by 6.51- and 4.19-folds in hPES1 and hPES2, respectively (Fig. 2). No significant difference was found in the levels of expression of the maternally expressed genes between the two hPESC lines (Supplementary Table S2).
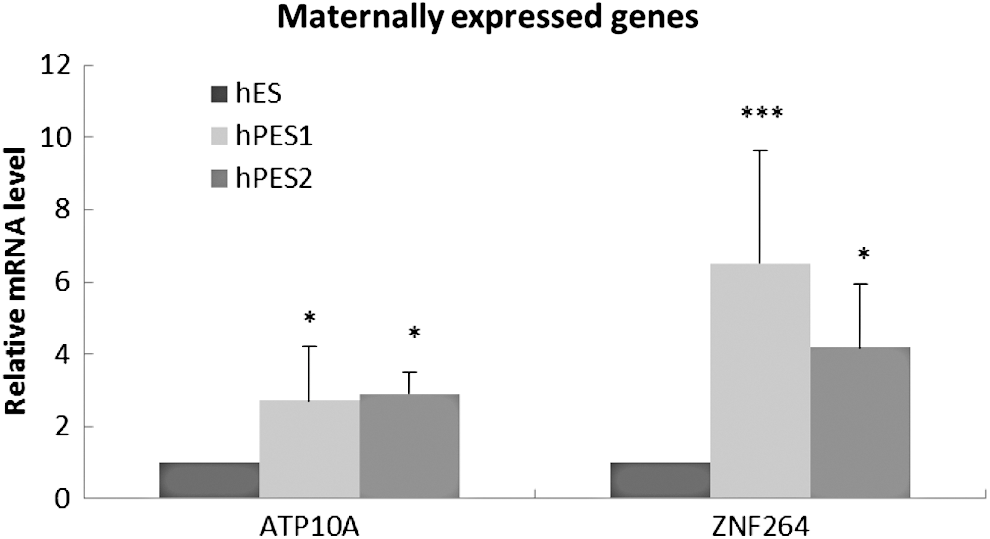
Expression change of 15 maternally expressed genes between hPESC and hESC. X-axis is maternally expressed genes, and Y-axis is the relative expressed level of imprinted genes. *P < 0.05, ***P < 0.001.
Comparison of the expression level of paternal genes in undifferentiated hPESCs and hESCs
The expression of 36 paternally imprinted genes was analyzed, 80.6% (29/36) of the genes was activated and more than the half of the genes [hPES1: 55.6% (20/36), hPES2: 58.3% (21/36)] showed a similar expression level as compared with the hESC lines (Supplementary Table S2). The expression of 15 paternally imprinted genes in both hPESC lines were downregulated as compared with hESC lines. Eight of these genes, including PEG10, SNUR, SNORD109A, ZIM2, PSIMCT-1, KCNQ1OT1, SNORD109B, SNRPN, were significantly reduced as compared with the hESC lines (Fig. 3). The expression level of other seven genes, including MAGEL2, NDN, SNORD107, SNORD64, SNORD108, PEG3, and IPW was not detectable in both hPESC lines.
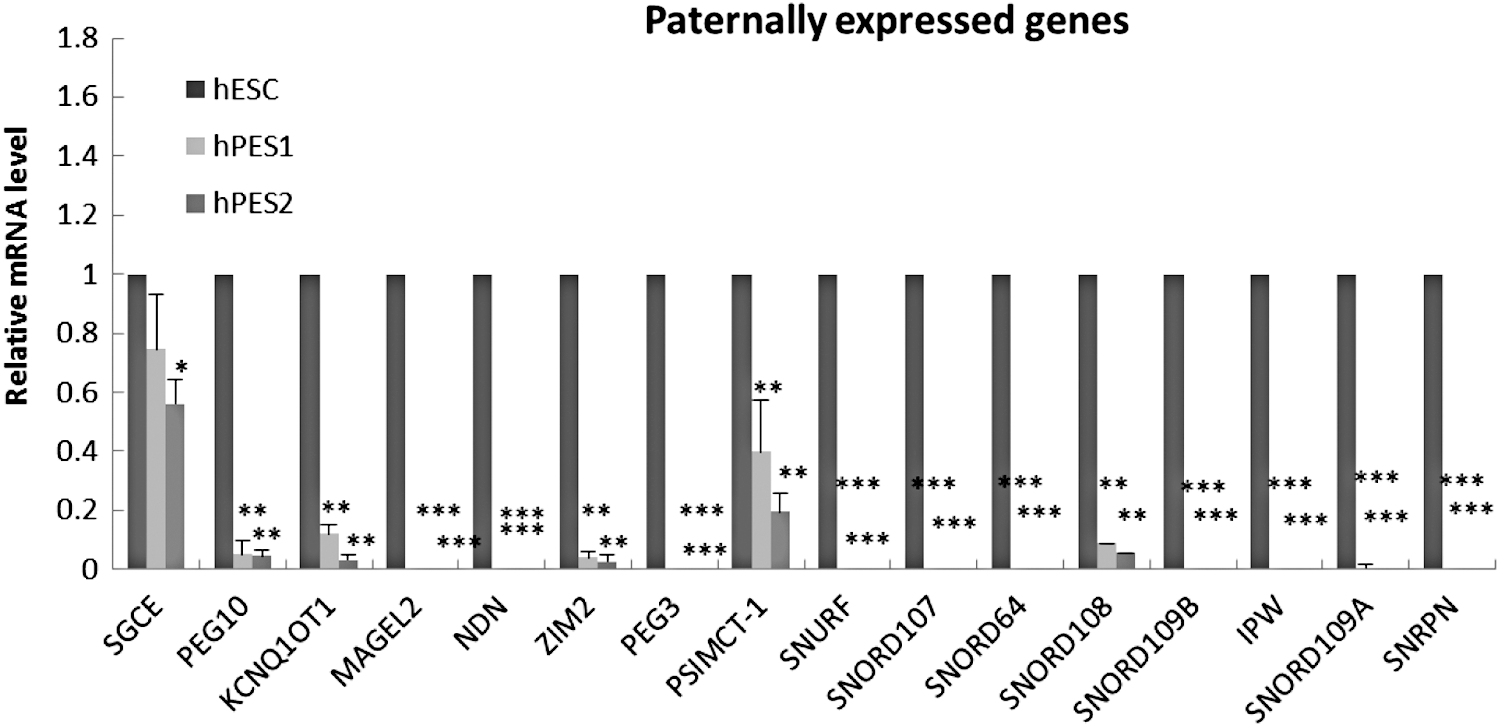
Expression change of 15 paternally expressed genes between hPESCs and hESC. X-axis is paternal expressed genes, and Y-axis is the relative expressed level of imprinted genes. *P < 0.05, **P < 0.01, ***P < 0.001.
To illustrate the expression change of the eight downregulated genes in hPESC lines, the reduction fold of transcription activity was listed below as compared with the undifferentiated hESC lines: the expression level of PEG10 was 19 (P < 0.01) and 22-folds (P < 0.01); SNURF was 25,000 (P < 0.001) and 41,000-folds (P < 0.001); SNORD109B was 2,500 (P < 0.001) and 4,900-folds (P < 0.001); ZIM2 was 26 (P < 0.01) and 36-folds (P < 0.01); PSIMCT-1 was 2.5 (P < 0.01) and 5.1-folds (P < 0.01); KCNQ1OT1 was 8.4 (P < 0.001) and 31-folds (P < 0.001); SNORD109B was 23,000 (P < 0.001) and 47,000 (P < 0.001) folds, and SNRPN was 21,000 (P < 0.001) and 32,000-folds (P < 0.001) reduced in undifferentiated hPESC1 and hPESC2, respectively.
The expression changes of the maternally and paternally imprinted genes in hPESCs during prolonged in vitro culture
During the in vitro culture of hPESC1 line, the expression of SNURF showed a positive correlation with passage number (r 2 = 0.8497, P < 0.05), whereas the other 62 imprinted genes did not show any correlation. This positive correlation was also found between the expression of CDKN1C and the passage number during in vitro culture of undifferentiated hPESC2 (r 2 = 0.8796, P < 0.05) (Fig. 4).
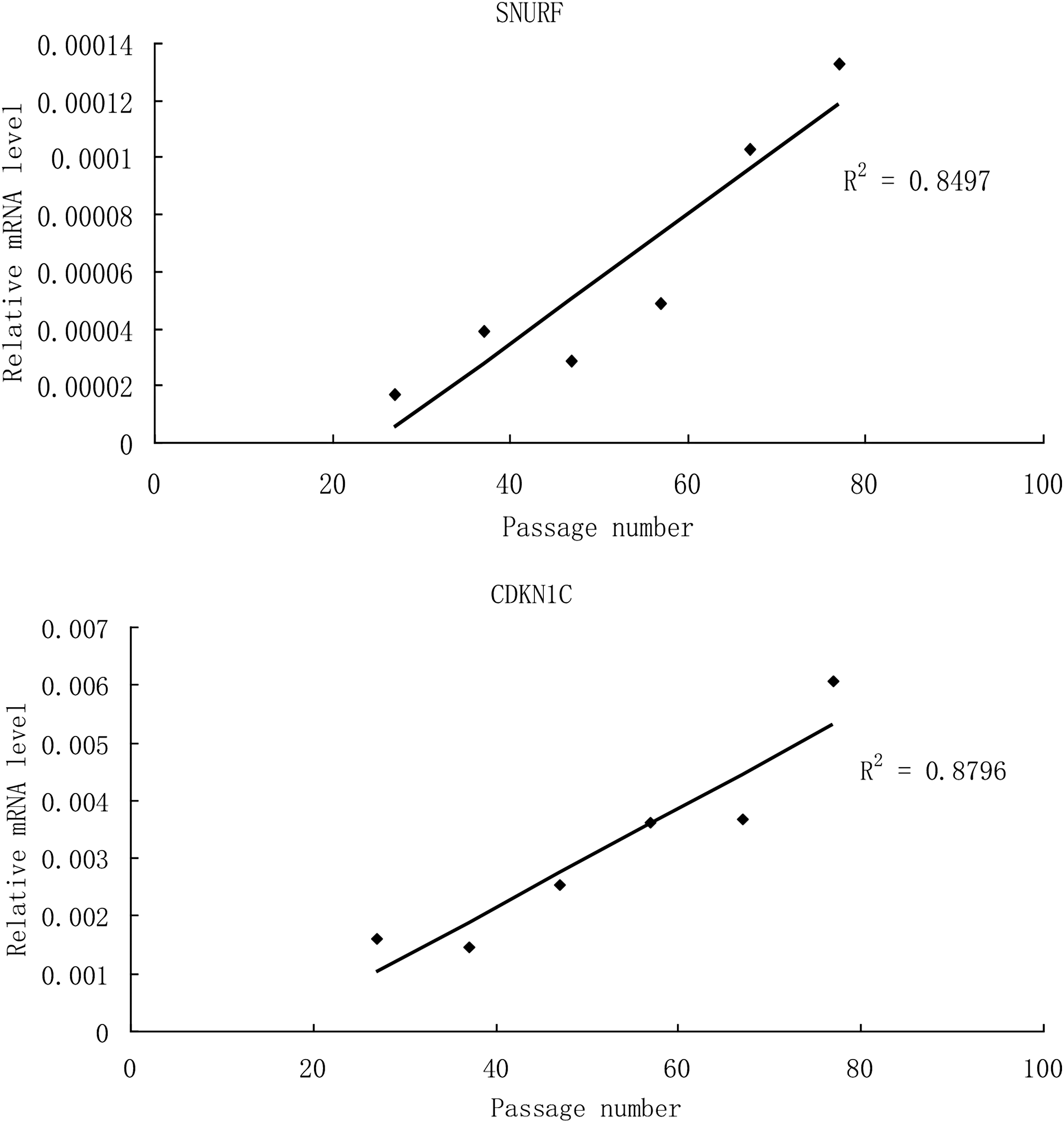
SNURF showed a positive correlation with passage number in hPES1 (r 2 = 0.8479, P < 0.05). CDKN1C showed a positive correlation with passage number in hPES2 (r 2 = 0.8796, P < 0.01).
The correlation between the expression change and the function of the imprinted genes in hPESCs during the differentiation
The expression of the imprinted genes in EBs from the hPESC1 line was analyzed (Supplementary Table S3). During the differentiation, the expression of four maternally (SLC22A2, SLC22A18, CPA4, and H19) and three paternally (IGF2, DLGAP2, and WT1) imprinted genes was significantly changed as compared with undifferentiated hPESC1. The expression of the remaining imprinted genes was not altered by differentiation of the hPESC1 cells. The expression of the imprinted genes, including SLC22A2, SLC22A18, CPA4, H19, and WT1 was upregulated and the increase in folds was 6.83-fold (P < 0.05), 4.19-fold (P < 0.01), 63.5-fold (P < 0.01), 10.3-fold (P < 0.01), and 3.31-fold (P < 0.05), respectively. On the other hand, only the expression of the imprinted gene DLGAP2 was downregulated by 33.3-fold (Fig. 5A).
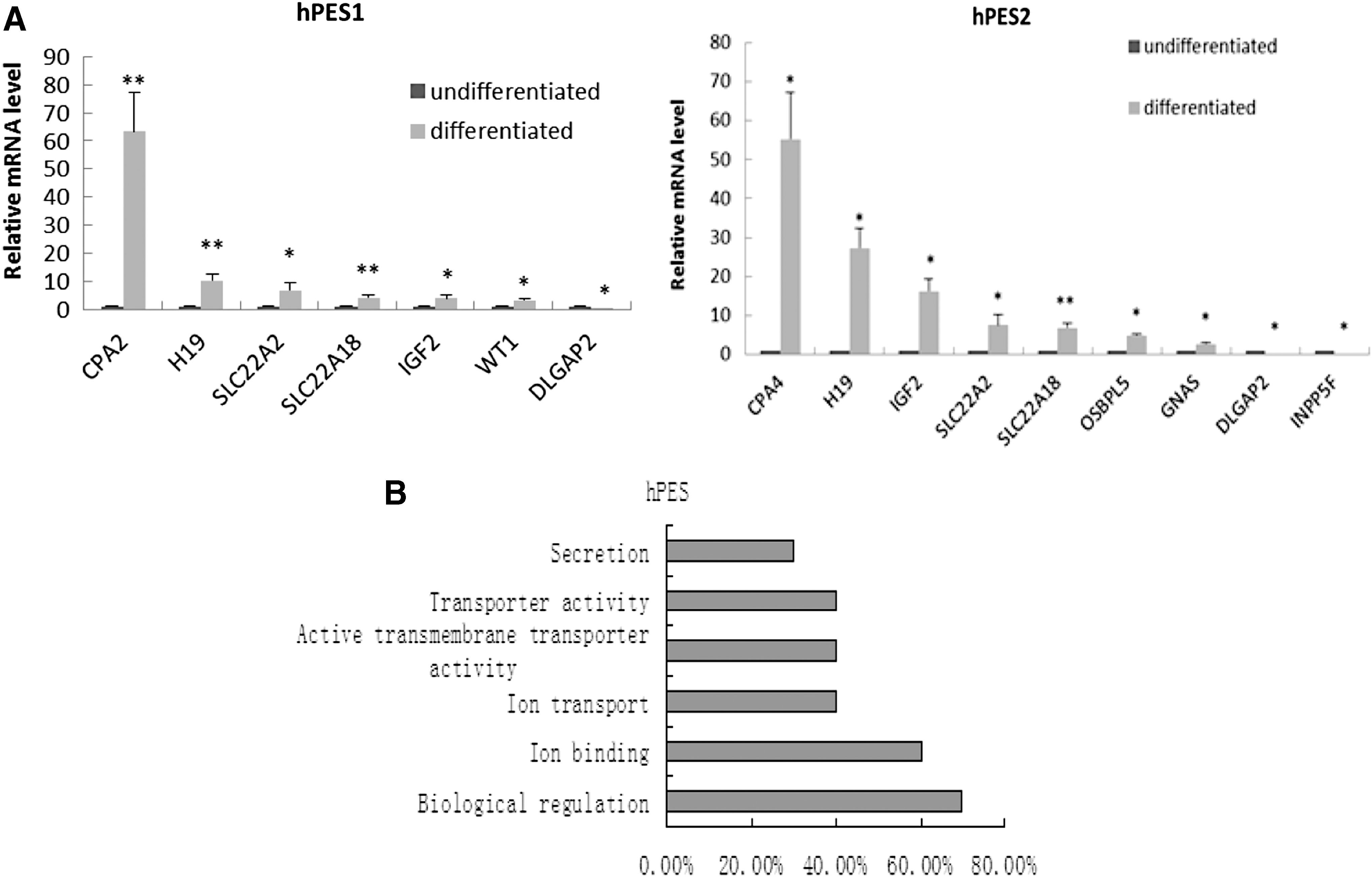
Expression change and analysis of function of imprinted genes in hPESCs in the undifferentiated and differentiated status.
In addition, the expression of the imprinted genes in the hPESC2-derived EBs was also analyzed. During the differentiation, the expression of six maternally (SLC22A2, CPA4, H19, SLC22A18, OSBPL5, and GNAS) and three paternally (DLGAP2, INPP5F, and IGF2) imprinted genes was significantly changed. The expression of the remaining imprinted genes was not altered in the hPESC2 line during the differentiation. The expression of the imprinted genes, including SLC22A2, CPA4, H19, SLC22A18, OSBPL5, GNAS, and IGF2 was upregulated and the increase in folds was 6.4-fold (P < 0.05), 43.7-fold (P < 0.05), 23.6-fold (P < 0.05), 6.1-fold (P < 0.01), 4.3-fold (P < 0.05), 2.2-fold (P < 0.05), and 18.6-fold (P < 0.05), respectively. The expression of the imprinted genes, DLGAP2 and INPP5F, was downregulated and the reduction folds were 18.2-fold (P < 0.05) and 22.2-fold (P < 0.05), respectively (Fig. 5A).
The functional analysis results suggest that the upregulated imprinted genes were markedly involved in ion binding, sodium ion transportation, and chromatin modification during early differentiated stage in hPESCs (Fig. 5B).
Comparison of the DNA methylation status between hPESC and hESC lines
The DNA methylation locations are major centers for regulation of transcription activity of the genes and may affect the 15 silenced or downregulated paternal genes in hPESC lines. Therefore, the methylation status of four DNA methylation locations, including KVDMR1 DMR, SGCE/PEG10 DMR, PEG3/ZIM2 DMR, and SNRPN/SNURF DMR was analyzed.
In the earlier passages (P27), a significant change of the methylation level within the four DMRs was observed between undifferentiated hESC lines and hPESC lines, P < 0.001 (Fig. 5). The methylation levels of KVDMR1, SGCE/PEG10 DMR, PEG3/ZIM2 DMR, and SNRPN/SNURF are listed as follows: undifferentiated hESC lines (58.2%, 52.4%, 49.2%, and 49.5%), undifferentiated hPESC1 line (95.9%, 55.9%, 91.6%, and 96.2%), and undifferentiated hPESC2 line (97.5%, 88.8%, 90.8%, and 97.1%).
The methylation level of four DMRs was analyzed in the genes of the EBs of hESC lines and hPESC lines. A significant change of the methylation level was also observed between these cell lines on day 14, P < 0.001 (Fig. 6). The methylation levels of KVDMR1, SGCE/PEG10 DMR, PEG3/ZIM2 DMR, and SNRPN/SNURF are listed as follows: differentiated hESCs (51.2%, 48.3%, 47.5%, and 50.5%), differentiated hPESC1 cells (99.4%, 60%, 80%, and 99%), and 96.3%, 90%, 90% and 95.7% in hPESC2 cells (Fig. 7).
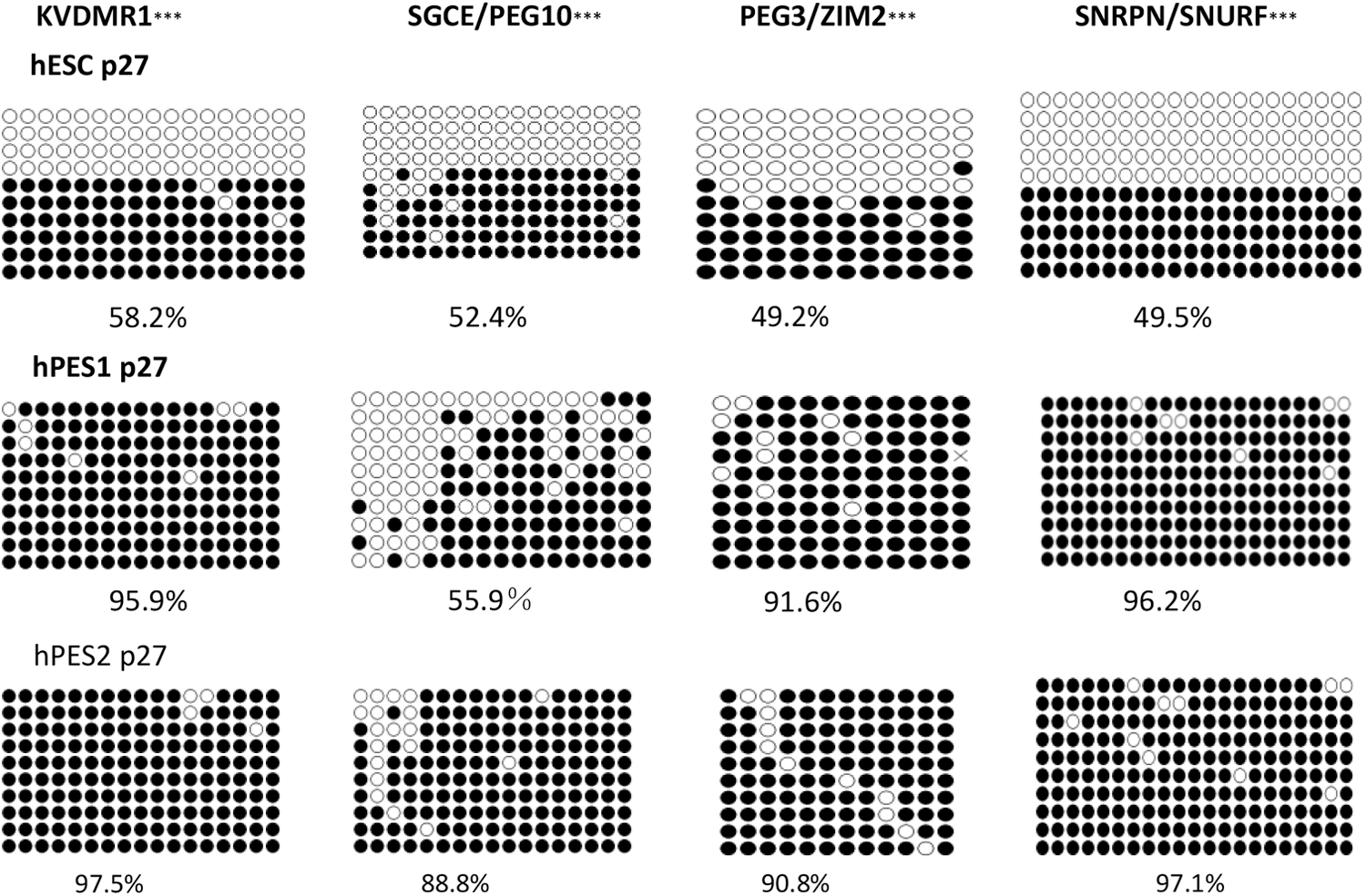
Comparison of the methylation levels of KVDMR1, SGCE/PEG10 DMR, PEG3/ZIM2 DMR, and SNRPN/SNURF in the earlier passages of undifferentiated hESC, hPES1 cells, and hPES2 cells. ***P < 0.001. DMR, differentially methylated region.
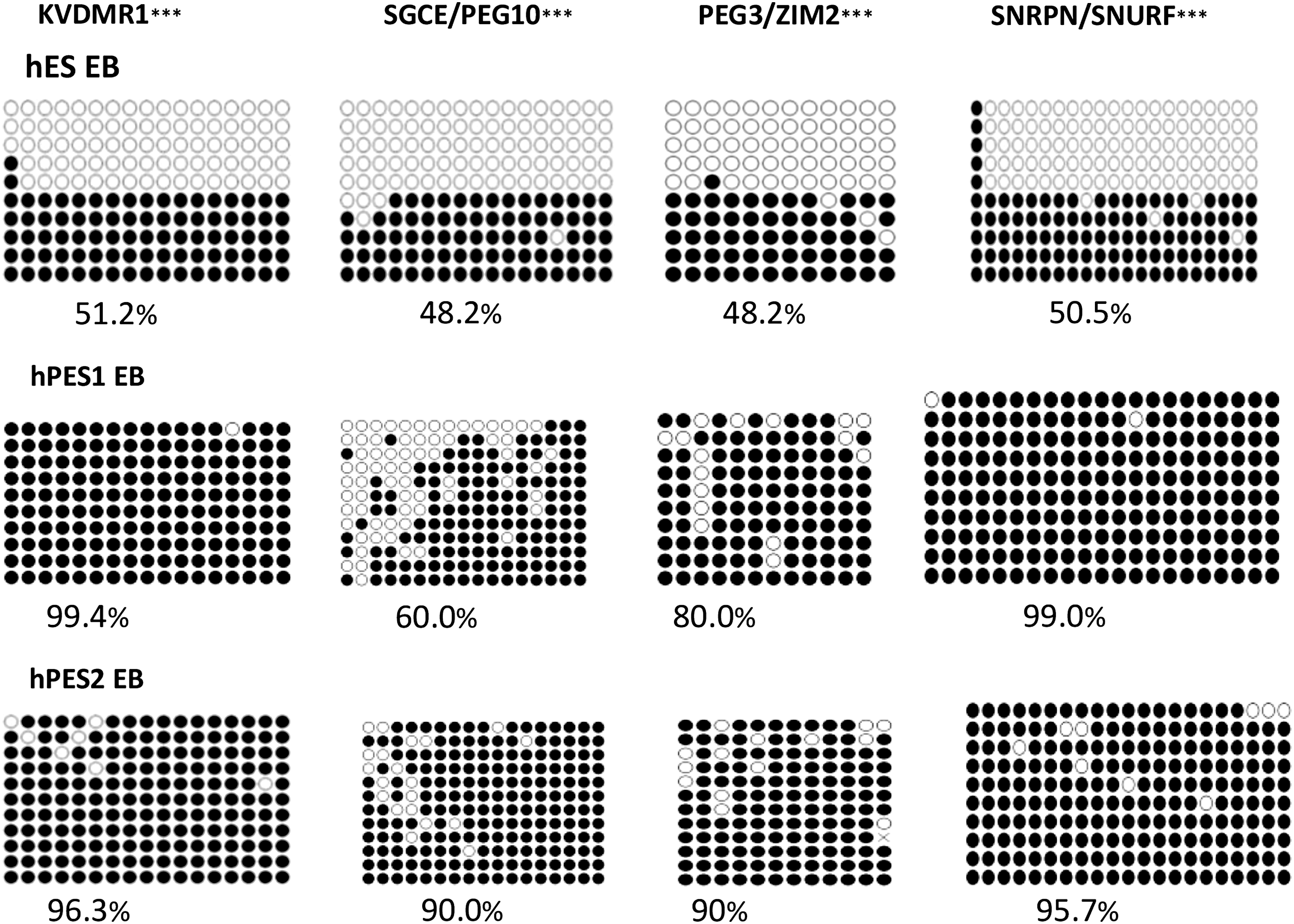
Comparison of the methylation levels of KVDMR1, SGCE/PEG10 DMR, PEG3/ZIM2 DMR, and SNRPN/SNURF in EB of hESC, hPES1 cells, and hPES2 cells. ***P < 0.001.
Discussion
This study quantitatively analyzed the expression patterns of the imprinted genes of hESCs from parthenogenetic embryos within undifferentiated and early differentiated stage. To our knowledge, this is by far the most wide-ranged survey study of the imprinting profile of hPESCs. In addition, the epigenetic status of hPESCs has been evaluated by analyzing the expression of the imprinted genes.
Multiple studies have mentioned that the expression status of the maternally imprinted genes in PESCs and parthenogenetic embryos could be different. Maternally imprinted genes, such as IGF2r, Gtl2, Mirg, and H19, are expressed in a significantly higher level in the mouse parthenogenetic embryos than in their fertilized counterparts [17,18]. In mouse PESCs and/or their derivatives, the expression of H19 and IGF2r has been shown to be similar to that in parthenogenetic embryos [19,20]. The expression level of H19 widely fluctuated between different PESC lines in rhesus monkeys, whereas the transcription level of UBE3A (maternally expressed) was similar in PESCs [15]. However, only a limited number of maternally expressed genes was investigated in these animal experiments.
In contrast to other mammals, such as mice and sheep, preliminary studies on hESCs and hESC-derived EBs have shown that hESCs maintain a substantial degree of epigenetic stability with respect to some imprinted genes [10,11]. In addition, Rugggunn et al. have demonstrated that hESC lines possess an epigenetic stability despite differences in the genetic background and in their derivation and initial propagation conditions [21]. The expression of H19, IGF2, and MEG3 are more variable and may provide sensitive indications of epigenetic status. Our results showed normal expression of H19, IGF2, and MEG3 in the two control hESC lines, which confirmed the normal expression status of the imprinted gene in these two hESC lines.
In this study, a similar expression pattern of the imprinted genes has been found in two separate hPESC lines as compared with the hESC lines. This result suggests that hPESCs also possess an epigenetic stability. Despite the different oocyte origins, the selected 63 imprinted genes have shown a similar expression pattern in two hPESC lines. All analyzed maternally imprinted genes were expressed in both undifferentiated hPESC lines. Twenty-nine (80.6%) of the analyzed paternal genes were expressed in both cell lines, and more than half (hPESC1: 55.6%, hPESC2: 58.3%) of which were expressed at a similar level as compared with the gene expression in the hESC lines. Since these two hPESC lines were maternally derived, the heterologous diploid genome of oocytes might attribute to variant expression levels of SNURF and CDKN1C.
A previous study has demonstrated that the maternally expressed H19 is normally expressed in the mouse PESCs [20]. In line with this data, our result illustrated that the majority of maternally imprinted genes (25/27, 92.6%) showed a similar expression pattern in two hPESC lines, with two exceptions, the ATP10A and ZNF264 showed an increased expression level.
The lack of a paternal genome in hPESCs might lead to disorder of genome imprinting as what happened in parthenogenetic embryos [22]. However, previous animal studies suggest that paternal genes could be expressed in the PESCs. It has been shown that mouse PESCs and/or its EBs have expressed all seven analyzed paternal genes [17]. Although the expression of paternal genes, MAGEL2, MKRN3, SNRPN, and NDN are missing, activated expression of PEG10, DIRAS3, SGCE, and IGF2 were found in PESCs of rhesus monkeys [15]. Our study has for the first time demonstrated that the paternal genes were not totally silent in hPESCs. Instead, 80.6% of analyzed paternal genes were activated and around 55% of the paternal genes maintained normal expression levels.
Our data showed that 2 maternal genes (ATP10A and ZNF264) were highly expressed, whereas 15 paternal genes (PEG10, MAGEL2, NDN, KCNQ1OT1, SNORD109A, SNRPN, SNURF, SNORD107, SNORD64, SNORD108, SNORD109B, ZIM2, PEG3, IPW, and PSIMCT-1) were significantly lowered or silenced in two hPESC lines. Aberrant imprinting status might alter the amount of the corresponding gene products and affects the cell proliferation or commitment and results in severe clinical syndromes and carcinogenesis [23,24]. The role of ZNF264 is not yet clear, but a high expression level of ATP10A often leads to autism spectrum disorder [25]. On the other hand, some of the repressed paternal genes were also correlated to neural disorders.
Inactivation of the large cluster of paternally expressed genes, such as SNURF, SNRPN, SNORD107, SNORD64, SNORD108, SNORD109A, SNORD109B, IPW, NDN, and MAGEL2, in the PWS region results in the neurological and behavioral phenotypes of Prader–Willi Syndrome [26]. Mice lacking the PEG3 increased apoptosis during the development process of nervous system, which affects its exploratory behavior, olfactory sensation, and the neuroendocrine regulation of female sexual behavior [27]. In addition, PEG3 is also a tumor suppressor gene [28], and decreased PEG3 expression has been detected in human ovarian cancer [5] and glioma [29]. SGCE mutations have been implicated in myoclonus dystonia [30].
Although parthenogenetic dopamine neurons from primate ESCs have been documented to restore function in experimental Parkinson's disease [31], it remains to be investigated whether the aberrant transcription of these genes interferes with the directed differentiation of hPESCs (especially neural differentiation) and ultimately affects cell/tissue function or tumorigenesis posttransplantation. Furthermore, deletion of the paternal allele of PEG10 results in early embryo lethality or prenatal growth retardation [32].
In this study, the expression levels of the majority of imprinted genes did not change during long-term culture process, which implies that the long-term in vitro culturing environment does not have a significant effect on the expression status of imprinted genes in hPESCs. To our surprise, some inactive paternally imprinted genes, such as SNRPN, SNURF, SNORD107, SNORD64, SNORD109B, and ZIM2 could be gradually activated during the in vitro culture process; the expression level of SNURF was upregulated as continuously passaged, and SNRPN and PEG10 were expressed in a significantly higher level after p50 of hPESCs. These results indicate that the expression of some repressed paternally expressed genes could be modified to some degree through extended culture.
In our previous study, five maternally imprinted genes (SLC22A2, SLC22A3, COPG2IT1, H19, and CPA4) and one paternally imprinted gene (IGF2) were markedly increased in early differentiated hESCs [16]. In this study, significantly upregulated expression levels of H19, IGF2, SLC22A2, SLC22A3/SLC22A18, and CPA4 were found during the early differentiation of hPESCs. Taken together, these data suggest that the expression changes of imprinted genes, which are accompanied with the cell differentiation in hPESCs, were consistent with that of hESCs. To our knowledge, this is the first time that this phenomenon has been clearly demonstrated.
H19 and IGF2 are related to the growth of embryos and fetus, and overexpression of IGF2 may lead to overgrowth of somatic cells [20]. In metabolic exuberant tissue, SLC22A1, SLC22A3, and SLC22A18 expressed at high levels and these metabolic transporter genes are able to adjust the growth of placenta and fetus [33 –36]6. CPA4 plays an important role in cellular proliferation and differentiation [37]. These genes may act as important regulators during the early differentiation stage of hPESCs and hESCs.
Accumulated evidence demonstrated that the expression of imprinted genes are regulated by certain epigenetic mechanisms, which act on the regulation and coding region of the genome and induce special chromatin structure [38]. These epigenetic mechanisms modify the DNA and chromatin, do not affect the DNA sequence, and are heritable.
The most studied epigenetic mechanism is DNA methylation of CpG dinucleotide [39]. It is also the main regulatory mechanism of parental-specific expression of imprinted genes, usually related to gene silencing. Most of the DNA methylation occurs in DMRs. The DMRs of the imprinting control center are CpG-rich regions, where two alleles from different origins are in different methylation status [21].
As demonstrated in our data, PEG3/ZIM2 DMRs, SNURF/SNRPN DMRs, and KVDMR1 DMRs in the genes of undifferentiated hPESCs and its EBs are highly methylated in early passages (p27), whereas in the DMRs of undifferentiated hESCs and its EBs are half methylated, and clones are almost completely methylated or demethylated. This might be the reason why 12 paternally imprinted genes (SNURF, SNRPN, SNORD107, SNORD64, SNORD108, SNORD109A, SNORD109B, IPW, NDN, MAGEL2, ZIM2, and KCNQ1OT1) are inactivated or downregulated in hPESCs. In contrast, the methylation status of SGCE/PEG10 is inconsistent with decreased PEG10 and PEG3 expression levels.
A possible explanation is that some other mechanisms regulate the expression of PEG10 and PEG3. Significant demethylation of the SGCE DMR occurred in hPESC1, which contributed to the activation and normal expression of SGCE. In hPESC2, SGCE DMR shows mild demethylation and activated SGC maintained a low-level expression. These results suggest that the demethylation status in SGCE DMR correlated with the activation degree of SGCE in hPESCs. This is the first study that has shown that the methylation of PEG3/ZIM2 DMRs, SNURF/SNRPN DMRs, KVDMR1 DMRs, and SGCE DMRs might be the important regulation mechanism for the abnormal paternal gene expression in hPESCs. It suggests a possibility to correct the depressed paternal gene expression in hPESCs by regulating the methylation status of related DMRs. However, this hypothesis requires further researches to verify.
The limitation of this study is that the expression level of the imprinted genes was only detected in undifferentiated and early differentiated hPESCs. The aberrant imprinted gene status was not detected in the derivative of hPESCs. Their influence on hPESCs differentiation and derivate cell function are still unknown. Previous study has shown that mouse pESCs exhibit pluripotency by correlating the expression of imprinting genes and by germline competence in chimera production [40,41]. More investigations are needed to further clarify the influence of imprinted gene status of hPESCs on cell differentiation capability and function or possibility of tumorigenesis after transplantation.
Our study results demonstrated a substantial degree of epigenetic stability in hPESCs with respect to the imprinted genes. Most of detected maternally imprinted genes and over half of detected paternal genes maintain the normal expression level in two undifferentiated hPESC lines. The expression level of imprinted genes is stable in undifferentiated hPESC lines during the long-term culture process. A same expression pattern of imprinted genes was observed in hPESC lines as compared with hESC lines during the early differentiation stage. Moreover, the 15 paternally imprinted genes, which were inactivated or downregulated in two hPESC lines, are correlated with the methylation level of PEG3/ZIM2 DMRs, SNURF/SNRPN DMRs, and KVDMR1 DMRs. Since there is a relation between the aberrant expressions of the 15 paternally imprinted genes and the development of nervous system and oncogenesis, it is necessary to clarify the effect of the imprinted genes upon derivatives of hPESCs. That would be important to the clinical application of hPESCs in cell replacement therapy and regeneration medicine in the future.
Footnotes
Acknowledgments
This work was supported in part by grants from National Natural Science Foundation of China (no.: 81270750), Guangdong Provincial Key Laboratory of Reproductive Medicine (no.: 2012A061400003), Fundamental Research Funds for the Central Universities (no.: 16ykjc10), and Guangzhou Technology Foundation (no.: 155700011/201508020006). The authors thank Pingpin HONG for her help with the passaging of ES cells.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.