
Abstract
Multipotent mesenchymal stem/stromal cells (MSCs) are strongly involved in tissue homeostasis mainly through paracrine regulation. In this study, we examined the influence of simulated microgravity on the angiogenic potential of adipose-derived MSCs (ASCs). The conditioned medium (CM) from random positioning machine (RPM)-exposed ASCs stimulated the formation of vessel network in ovo, endothelial cell (EC) capillary-like network, and nondirected EC migration in vitro. These effects were driven by alteration of both angiogenesis-related gene and protein expression. The elevation of angiogenic regulators Serpin E1, Serpin F1, IGFBP, VEGF, and IL-8 was detected in ASC-CM after 3D-clinorotation. In addition, transcription of genes encoding growth factors with proangiogenic activity were upregulated including VEGF-c and VEGF-a. These data evidenced that besides direct effect on ECs, microgravity could provoke MSC-mediating specific microenvironment for ECs supporting their functions, that is, proliferation and migration via increased production of IL-8 and VEGF as well as other paracrine factors involved in angiogenesis regulation.
Introduction
M
The microenvironmental milieu, including mechanical factors, is playing an important role in the determination of stem cell fate and programming of progenitor cell differentiation. It has been demonstrated that MSCs are sensitive to gravitational changes such as real or simulated microgravity [4 –6]. Morphological and functional rearrangements in cultured MSCs were observed at the earliest stages of exposure to simulated microgravity [7]. As a source of trophic mediators, MSCs secrete a range of growth factors and other chemokines to induce cell functions. It was shown earlier that the production of cytokines were susceptible to real and simulated microgravity [8 –10].
It is now convincingly demonstrated that a subset of MSCs in vivo is located in the perivascular niche in close association with endothelial cells (ECs) being strongly involved in EC regulation [11 –13]. A number of studies are focusing on the impact of microgravity on ECs, but there are no data on MSC angiogenic activity under real or simulated microgravity. Meanwhile, a broad spectrum of angiogenic factors has been detected in the secretome of MSCs, including vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF), fibroblast growth factors (FGFs), angiopoietin-1 (Ang-1), monocyte chemoattractant protein-1 (MCP-1), interleukin-6 (IL-6), interleukin-8 (IL-8), placental growth factor (PLGF), and so on [1,2,14]. For this reason, the goal of present study was to characterize MSC angiogenic potential under simulated microgravity.
Materials and Methods
Isolation and culture of adipose tissue-derived MSCs
Adipose tissue samples were obtained in the frame of Scientific Agreement from multidisciplinary clinic “Soyuz” (Moscow, Russia) after elective liposuction procedures under local anesthesia from healthy patients with written informed consent. Adipose tissue was processed using guidelines specifically approved by Biomedicine Ethics Committee of Institute of Biomedical Problems, Russian Academy of Sciences (Physiology Section of the Russian Bioethics Committee, Russian Federation National Commission for UNESCO, Permit No. 314/MCK/09/03/13). Adipose-derived MSCs (ASCs) were isolated using standard method described by Zuk et al. [3] with modifications of Buravkova et al. [15]. Cells were expanded in α‐MEM (Gibco, Life Technologies) with 50‐U/mL penicillin–streptomycin (PanEco, Russia), and 10% fetal bovine serum (FBS) (HyClone) at standard conditions (5% CO2, 37°C).
ASCs complied with the minimal set of criteria of a joint IFATS and ISCT statement [16,17]. In brief, ASCs were adhered fast and displayed a fibroblast-like morphology. More than 95% of ASCs were positively stained with antibody against stromal markers: CD90, CD73, and CD105, and negatively–against pan-leukocyte CD45 antigen, as was revealed using flow cytometry (Accuri C6; BD Biosciences). ASCs underwent osteo- and adipogenic differentiation in the presence of the appropriate stimuli in the medium. To induce osteogenic differentiation, complete α-MEM was supplemented with 10−8 M dexamethasone, 10 mM glycerol-2-phosphate, and 0.2 mM L-ascorbic acid 2-phosphate (Sigma). Osteogenic differentiation was confirmed with Alizarin red staining of the mineralized matrix components (Millipore). To induce adipogenic differentiation, the medium was supplemented with 0.5 mM isobutyl methylxanthine, 1 μM dexamethasone, 10 μg/mL insulin, and 200 μM indomethacin (Sigma). Adipogenic differentiation was assessed by the evaluation of cytoplasmic Oil-Red-O-stained lipid droplets (Millipore). ASC immunophenotype and differentiation capacity were evaluated after each passage.
Isolation and culture of ECs
Cryopreserved human umbilical vein endothelial cell (HUVEC) samples were provided by the Cryocenter Cord Blood Bank (Moscow, Russia), as part of a Scientific Agreement. The cells were cultured in 199 medium (M199) (Gibco, Life Technologies) supplemented with 10% FBS (HyClone), 200 μg/mL EC growth factor (Sigma-Aldrich), 2 mM glutamine (Gibco, Life Technologies), 1 mM sodium pyruvate (Gibco, Life Technologies), 50 U/mL penicillin, and 50 μg/mL streptomycin under 5% CO2, 37°C in a CO2-incubator (Sanyo, Japan). HUVECs formed a typical confluent monolayer of elongated and polygonal cells positively stained with antibody against CD31–a marker of ECs.
Microgravity simulation
A desktop Random Positioning Machine (RPM) (Dutch Space, Leiden, Netherlands) was used to simulate microgravity effects. Speed (53–65 °/s) and direction of device rotation were randomized by dedicated control software at the computer user interface. The maximum distance between cell monolayer and center of rotation was 7.5 cm. The gravity value averaged 10−4g [18].
Cells were plated into culture flasks 5 days before the beginning of the experiment at a density of 3,000 cells/cm2. After ASCs reached 80%–90% of confluence, culture flasks were completely filled with medium without air bubbles to prevent sloshing of the medium and shear stress. The RPM with flasks (surface area: 25 cm2, volume: 50 mL, CELLSTAR; Greiner Bio-One, Germany) fixed on desktop was placed into thermostat under standard temperature conditions at 37°C. Samples were run on the RPM for 96 h. A part of flasks was placed in routine CO2 incubator (37°C), to serve as static control (Control). Another part of flasks was exposed on orbital shaker as dynamic control (DC) at 37°C in thermostat. DC was applied to estimate the contribution of medium stirring during RPM exposure. Conditioned medium (CM) from all samples was collected for further analysis, cells were washed with PBS, and total RNA was extracted for PCR analysis of gene expression.
Chorioallantoic membrane assay in ovo
ASC angiogenic activity was evaluated in chorioallantoic membrane (CAM) assay using Japanese quail embryos [19]. The fertilized eggs were placed into the incubator and kept at 37°C for 6 days. Then, a window was made in the shell and 30 μL of αMEM or ASC CM were added. The windows were sealed with medicinal glue BF-6 (Vertex, Russia) and the incubation was continued for extra 24 h. Then, the vascularization of CAM was analyzed by morphometric analysis using AngioQuant software (
Capillary-like tube formation
ASCs angiogenic activity in vitro was examined using model of capillary-like tube formation in Matrigel according to the manufacturer's protocol. HUVECs from second to fourth passages were seeded in 96-well plates coated with growth factor reduced Matrigel (Corning) in concentration 2 × 104 cells per well. Conditioning medium was diluted with full M199 1:1. At least two wells were used for each sample of ASC CM. M199 serum-free was utilized as negative control; M199 with 10% FBS and VEGF (25 ng/mL) served as positive control. Plates were placed into CO2-incubator at 37°C and capillary-like structures were examined after 15 h under the phase contrast microscope (Leica, Germany). Total number of tubular complexes was counted with Image Processing Software—Image-Pro Plus (Media Cybernetics).
Nontargeted cell migration assay (“wound healing”)
Nontargeted EC migration was evaluated in cell monolayer at a cell density of 104 cells per cm2 using the in vitro “scratch” assay [20]. ECs were scratched with a sterile pipette tip to create a “wound.” The migration area was calculated as the difference between the initial and final wound area.
Analysis of ASC secreted proteins
To characterize paracrine activity of ASCs, CM was collected, centrifuged at 2,500 g to remove cell debris, and stored at −70°C until the measurements. VEGF-a, TGF-β, IL-6, and IL-8 concentrations in ASC CM were evaluated using Human VEGF ELISA Set (Peprotech), Human TGF-beta 1 DuoSet ELISA (R&D), Human IL-6 ELISA Set (BD), and Human IL-8 ELISA Set (BD) according to the manufacturer's instructions. To detect 55 human angiogenesis-related proteins, CM was analyzed using Proteome Profiler Human Angiogenesis Array Kit (R&D) according to the manufacturer's instructions. The data were analyzed using Image Lab™ Software Version 5.0 (Bio-Rad).
Quantitative PCR analysis
Expression of 84 growth factor genes in the RPM-exposed ASCs compared with static control was analyzed using The Human Growth Factors RT2 Profiler PCR Array (Qiagen). To evaluate gene expression, total RNA was extracted with QIAzol Reagent (Qiagen) and purified by the phenol/chloroform technique. Reverse transcription was performed using a QuantiTect Reverse Transcription Kit (Qiagen) according to the manufacturer's protocol. Resulting cDNA was mixed with RT2 SYBR Green/ROX PCR Master Mix (Qiagen) and added to 96-well plates. Polymerase chain reaction was performed using the M × 3000P system (Stratagene). The data were analyzed using RT2 Profiler PCR Array Data Analysis ver. 3.5 software (
Statistical analysis
At least three independent experiments were performed in three replicates each and the averaged data are expressed as mean ± standard deviation (M ± SD). Analysis of group differences was performed by nonparametric Mann–Whitney test for independent samples using SPSS 14.0 software. Statistical significance was considered at P < 0.05.
Results
Functional assays of ASC angiogenic activity
The effect of simulated microgravity on ASC angiogenic activity was evaluated after 96 h of 3D-clinorotation using RPM. For this purpose, the CM was collected and processed as described above.
To assess angiogenic activity of ASC CM, several experimental models were applied. The effect on the blood vessel growth in ovo was examined in CAM assay using Japanese quail embryo. After 24 h of exposure with CM, a significant increase of vessel number was detected only after addition of CM from RPM-exposed ASCs, compared with both static and DCs (Fig. 1). The formation of capillary-like tubes by HUVECs in 3D basement membrane matrix “Matrigel” was used to assess ASC CM influence ex vivo. The number of tubule complexes after RPM-CM exceeded significantly those in static and DCs (P < 0.01) (Fig. 2). ASC-CM is known to stimulate nontargeted HUVEC migration in “wound healing” assay [11]. Compared to static and DCs, the wound closure in RPM-CM was faster (P < 0.05) (Fig. 3). Thus, the analysis of the ASC-CM effects in functional assays revealed the stimulative impact of 3D-clinorotation on ASC angiogenic potential. In functional assays of ASC angiogenic activity (in ovo and in vitro), there were no differences between DC and Control data.
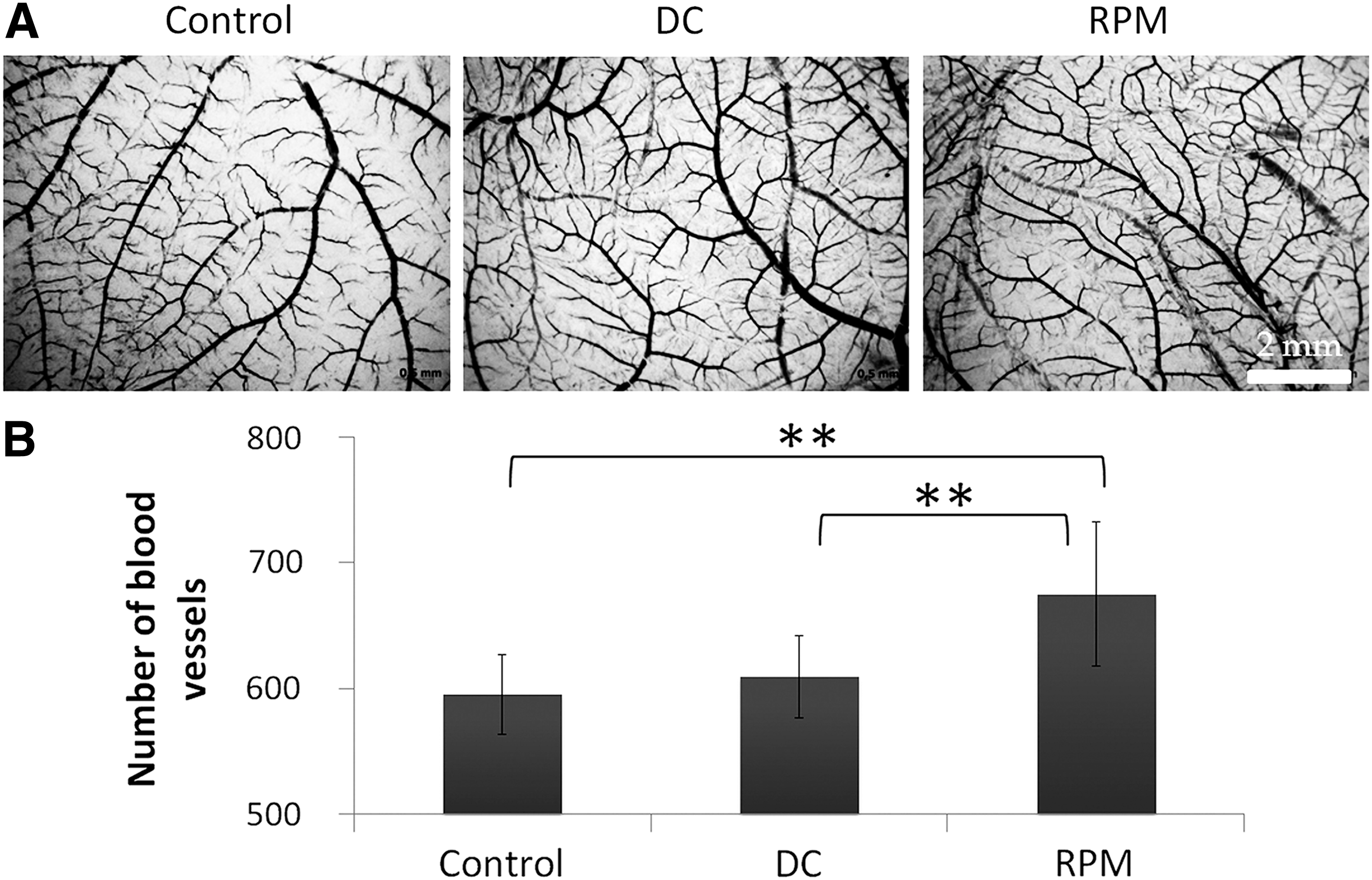
The effect of ASCs on the angiogenesis in ovo:

The effect of ASCs on the capillary-like tube formation in Matrigel:
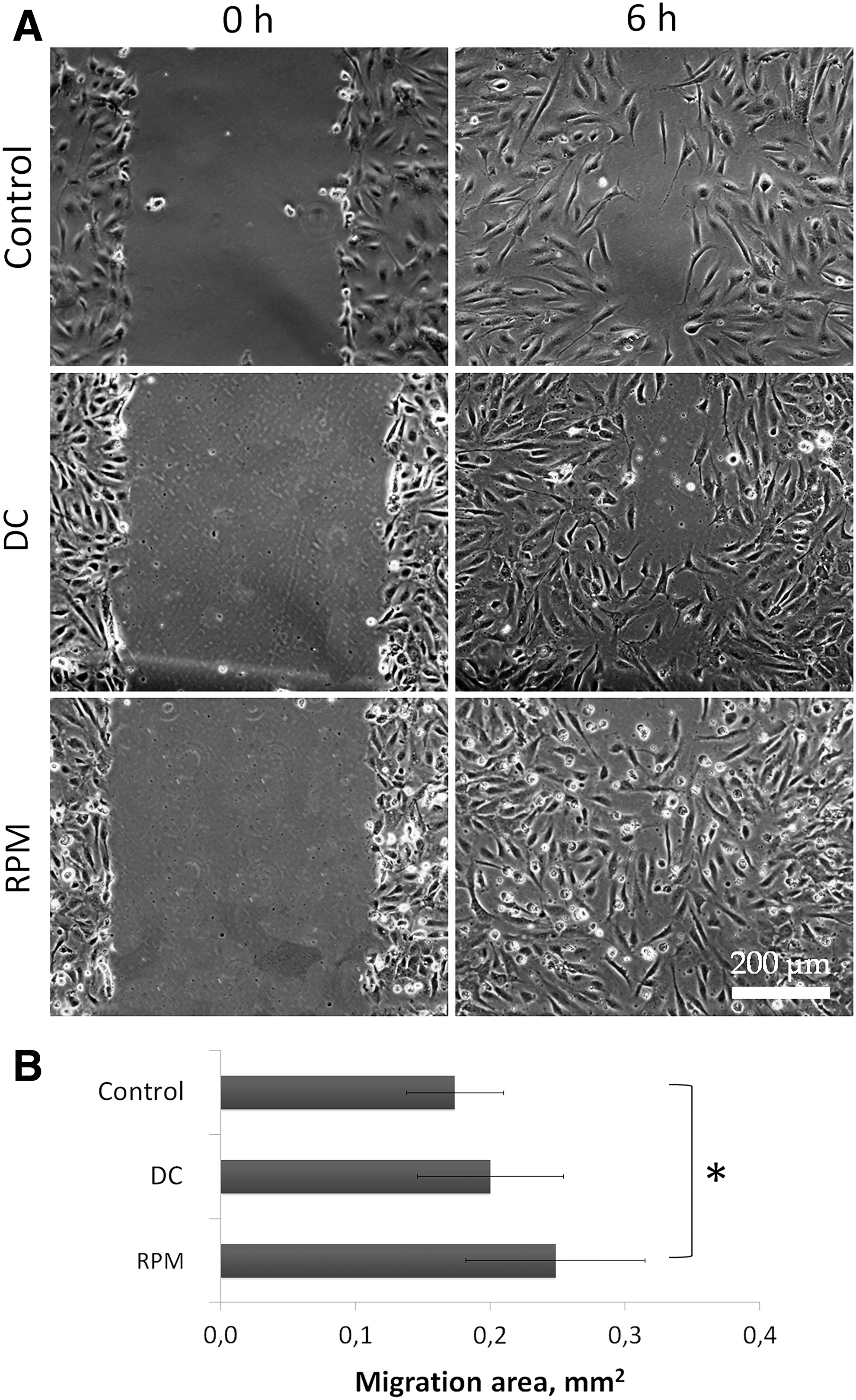
The effect of ASCs on ECs migration in “wound healing” assay:
Characterization of molecules involved in realization of ASC angiogenic effects
To characterize the profile of ASC-CM, we analyzed 55 human angiogenesis-related proteins using Proteome Profiler Human Angiogenesis Array Kit. Only eight proteins were detected both in static control and RPM-CM. The level of Serpin E1, Serpin F1, IGFBP3, IL-8, and VEGF was increased, while the level of TIMP-1 was decreased under simulated microgravity. Concentration of PTX3 and TSP-1 remained essentially unchanged (Fig. 4).
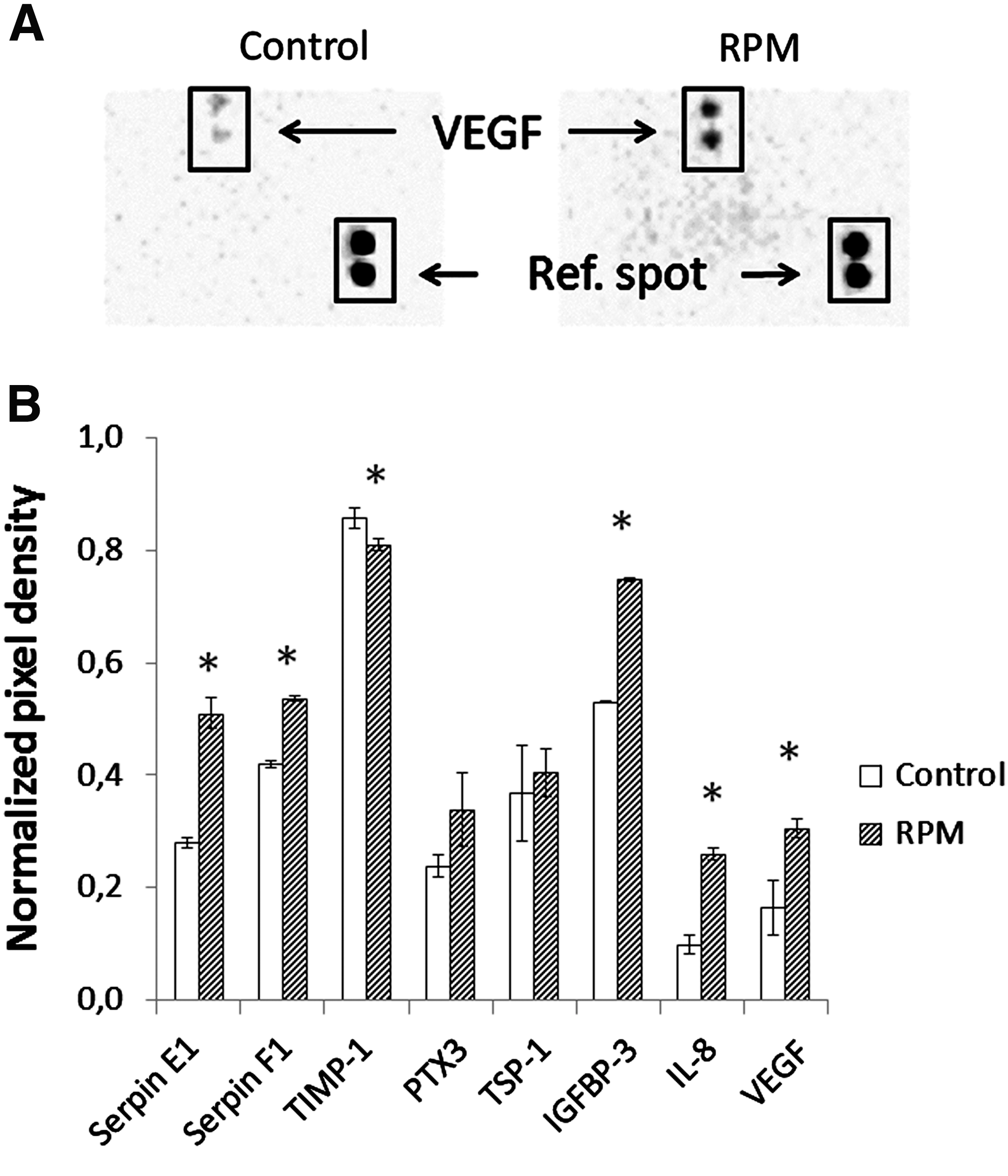
Evaluation of human angiogenesis-related proteins in ASC conditioning medium after 3D-clinorotation:
ELISA was applied to estimate the paracrine mediators with proangiogenic activity. Compared to the static and DCs, the level of VEGF-α and IL-8 was increased whereas TGF-β remained unchanged under simulated microgravity. The concentration of IL-6 was elevated in the DC (Fig. 5). According to the data presented above, IL-6 level was only one which was modificated.
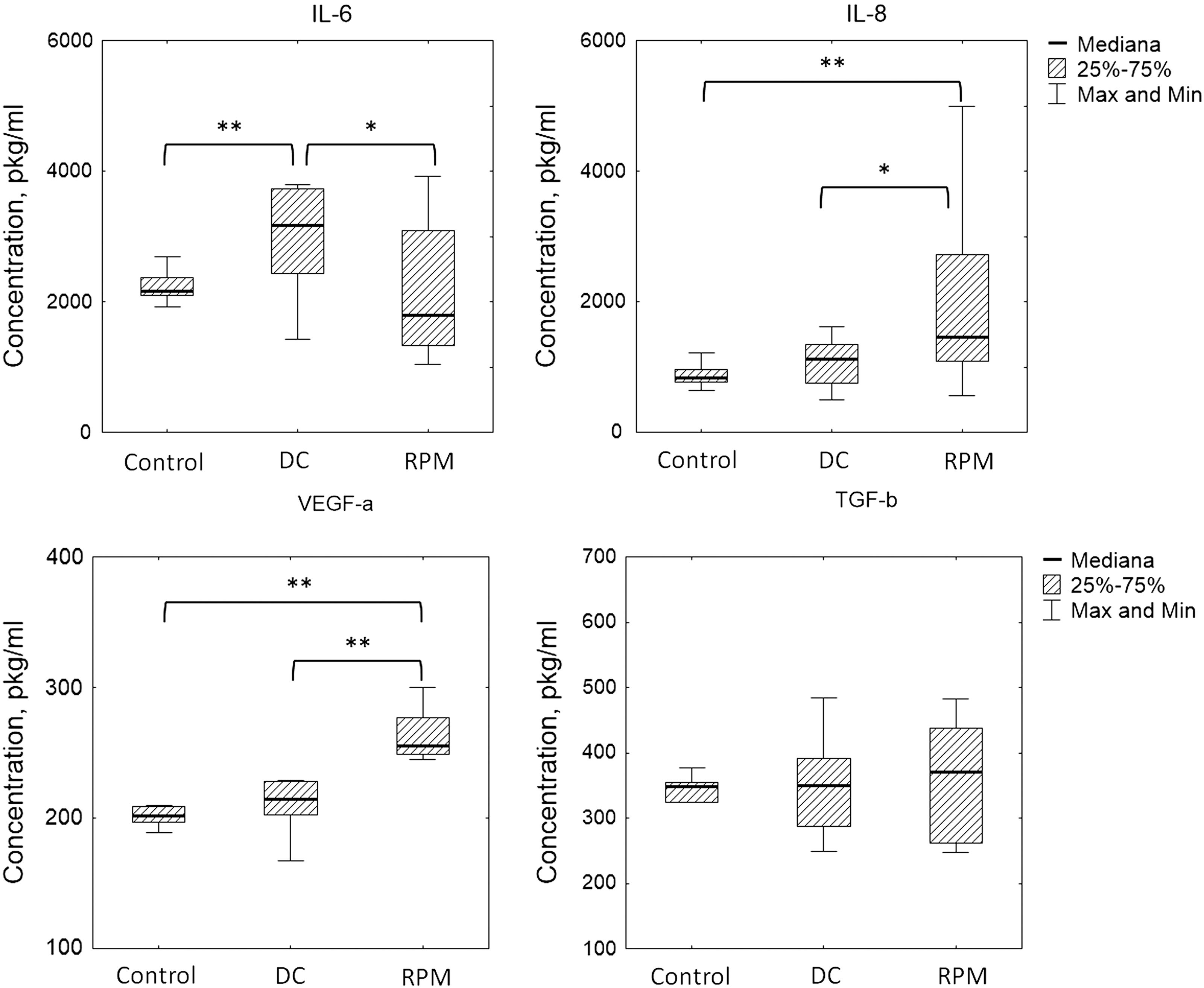
The effect of simulated microgravity on the cytokine production by ASCs. The data are presented as box plots (n ≥ 9, *P < 0.05, **P < 0.01).
Using human Growth Factors RT2 Profiler PCR Array, the transcription of 84 genes of ASCs in control and RPM groups was examined. Under simulated microgravity, seven genes encoded growth factors with proangiogenic activity were more than twofold upregulated, including BDNF, CXCL1, VEGF-c, DKK1, FGF5, GDF10, and VEGF-a (Fig. 6).

The effect of simulated microgravity on the ASC growth factor gene expression. The data are presented as Volcano plot (n = 3).
Discussion
Numerous in vitro studies demonstrated the high sensitivity of ECs to gravitational changes [22–24], although many issues still remain controversial because of the implication of various cell lines and different protocols. Meanwhile, certain generally recognized conclusions already have been done. So far, it is commonly agreed that the proliferative activity of macrovascular ECs, that is, HUVEC and bovine aortic ECs, is increased under simulated microgravity, whereas that of microvascular ECs decreased; endothelial NO synthesis is enhanced, cytoskeleton is rearranged, and gene expression is modified [22 –26]. It is postulated that actin cytoskeleton rearrangement modulated by lowered activity of small RhoA GTPase is one of the major mechanisms responsible for EC gravitational sensitivity [27].
The response of ECs to microgravity strongly depends on their local microenvironment both extracellular and cellular. As perivascular niche is one of principal tissue depots of MSCs, these cells play an important role in EC regulation [12,13]. In this study, we were the first who examined the impact of simulated microgravity-exposed MSCs on ECs in addition to direct microgravity effects on those. Conditioning medium from RPM-exposed ASCs was used as a source of stimulating factors for ECs. The ability to produce bioactive molecules (trophic function) is assumed as a major path of MSC biological activity. They secrete a whole range of angiogenic factors providing EC regulation in vitro and in vivo [2]. Our previous study demonstrated high viability, proliferative activity, and enhanced paracrine secretion of ASCs under simulated microgravity [28]. In particular, it was found that IL-8 production was increased during 10- and 20-day RPM exposures of BM-MSCs and their osteogenic progeny [10].
Using ASCs, we succeeded to demonstrate the angiogenic effects of paracrine cocktail from RPM-exposed ASCs on the formation of vessel network in ovo, EC capillary-like network and nondirected EC migration in vitro. This enhanced angiogenic activity of ASCs was driven by alteration of both angiogenesis-related gene and protein expression. Specifically, the level of proangiogenic VEGF and IL-8 as well as negative angiogenic regulators–serine proteases (Serpin E1, Serpin F1), insulin-like growth factor-binding protein 3 (IGFBP3), were increased in ASCs under simulated microgravity.
These findings were confirmed by ELISA data on elevation of VEGF-a and IL-8. Besides, transcription of genes encoding growth factors with proangiogenic activity were upregulated, including VEGF-c and VEGF-a. VEGF is a key positive regulator of principal phases of angiogenesis, as EC proliferation and migration [29], while IL-8 is an inflammatory chemokine that has an angiogenic potential and can stimulate VEGF production in MSCs via P13K/Akt and MAPK/ERK signaling pathways [30]. It is noteworthy that although we detected the increased level of both positive and negative regulators of angiogenesis in microgravity-exposed ASCs, the functional assays confirmed the stimulating effect of RPM-CM on EC migration and proliferation. From these data one can suppose that the multidirectional regulation could occur in MSCs under microgravity with the predominance of proangiogenic activity.
Recently, it was demonstrated that simulated microgravity could potentiate the direct differentiation of MSCs into endothelial-like cells under certain induction besides elevation of paracrine proangiogenic activity [31]. It was shown that after the microgravity exposure, gene expression and production of Flk-1, an EC marker, were increased. Moreover, Flk-1 acts as a VEGF receptor and is therefore involved in MSC differentiation into ECs [32,33].
The presented data supported the assumption that under simulated microgravity MSCs could provide specific microenvironment for ECs supporting their functions, that is, proliferation and migration via increased production of IL-8 and VEGF as well as other secondary growth factors involved in angiogenesis regulation.
Footnotes
Acknowledgments
This study was supported by the Russian Science Foundation (project no. 16–15-10407).
Author Disclosure Statement
No competing financial interests exist.