
Abstract
Osteoporosis is characterized by deterioration of bone microarchitecture and low bone mass. One of the primary causes of osteoporosis is the decrease in the osteogenic differentiation of mesenchymal stem cells (MSCs). Tissue engineering therapy with genetically modified MSCs has attracted much attention in the study of bone regeneration. In this study, we found that the expression level of miR-450b was upregulated during osteogenic differentiation of human adipose-derived mesenchymal stem cells (hADSCs). To explore the effect of miR-450b on the osteogenesis of hADSCs, we performed a series of gain- and loss-of-function analyses and demonstrated that miR-450b not only promoted the process of hADSC differentiation to osteoblasts in vitro but also enhanced ectopic bone formation in vivo. Bone morphogenetic protein 3 (BMP3), the most abundant BMP member in bone, was identified as a direct target of miR-450b. Downregulation of the endogenous expression of BMP3 could mimic the effect of miR-450b upregulation on the osteogenic differentiation of hADSCs. Overall, our study first demonstrated that a novel microRNA miR-450b was essential for hADSC differentiation, which could promote osteogenic differentiation in vitro and enhance bone formation in vivo by directly suppressing BMP3.
Introduction
O
To increase the rate of commitment of human adipose-derived mesenchymal stem cells (hADSCs) into the osteogenic lineage, a large number of studies have explored the basis of the complex interactive networks [6,7]. Several crucial molecular pathways, such as those of Wnt/β-catenin [8], transforming growth factor beta (TGF-β) [9], bone morphogenetic protein (BMP) [10], and MAPK [11], regulate each other. Osteoblasts and mesenchymal stem cells (MSCs) are the main cells involved in these processes [12]. MSCs can be recruited to bone and then differentiate into osteoblasts, thus promoting bone formation [13]. Previous studies have reported that gene repression before commitment of MSCs is mediated by microRNAs (miRNAs), because gene repression is most prevalent before commitment within a lineage [14]. Therefore, it is important to understand how miRNAs coordinate with intracellular signaling pathways and regulate the osteogenic differentiation of MSCs.
miRNAs are small (∼22 nucleotides) noncoding RNAs that can post-transcriptionally regulate gene expression by either inducing mRNA degradation or by repressing translation through binding to the mRNA 3′-untranslated region (3′-UTR) of complementary sequences of target genes [15]. miRNAs play a critical role in biological processes including stem cell self-renewal, differentiation, proliferation, and the cell cycle [16], and ∼30% of human protein-coding genes are regulated by miRNAs [17]. Elucidating the molecular mechanisms by which miRNAs control osteogenesis and bone formation will provide more effective therapeutic methods for osteoporosis. Several miRNAs involved in regulating osteogenic differentiation have been identified, such as miR-146a [18], miR-214 [19], miR-139-5p [20], miR-1192 [21], and miR-34a [22,23]. Most of them are negative regulators of osteogenic differentiation and bone formation.
Recently, miRNA-based gene therapy has been considered a potential therapeutic method. A number of miRNAs, such as let-7 family [24], miR-200 family [25], miR-10b [26], miR-155 [27], and miR-208 [28], have been used in preclinical models of miRNA therapeutics that have been carried out successively, and some have moved into clinical development. A mimic of the tumor suppressor miRNA, miR-34, has reached phase II clinical trials for melanoma treatment [29]. Anti-miRs targeted at miR-122 have reached phase II trials for treating hepatitis [30]. Similarly, safety, tolerability, and pharmacokinetic studies of miR-201 and miR-106 are also being studied in phase I. The use of therapeutic miRNAs in clinical treatment of osteoporosis is now limited by the lack of cell-specific delivery systems. Researchers have developed polyurethane nanomicelles modified by the acidic peptide Asp8 [19]. This delivery system encapsulates and selectively delivers miRNAs to OSCAR-positive cell surfaces in vivo and are used for osteoporosis therapy in mice.
Here, we identified that miR-450b, as a positive regulator of osteogenic differentiation, can promote osteogenic differentiation in vitro and enhance bone formation in vivo. BMP3 was identified as a direct target of miR-450b. With the miRNA therapeutics becoming a clinical reality, miR-450b may be a candidate in miRNA therapeutics for the prevention and treatment of osteoporosis and other bone metabolism-related diseases.
Materials and Methods
Isolation and culture of hADSCs
All experiments followed the procedures approved by the Ethics Committee at the Chinese Academy of Medical Sciences and Peking Union Medical College. Human adipose tissue was obtained from donors (20–45 years old) undergoing liposuction. hADSCs were isolated from human adipose tissue and were cultured as previously described [31]. Briefly, the adipose tissue was washed thrice with phosphate-buffered saline (PBS) and digested with 0.2% collagen enzyme for 30 min at 37°C, followed by being treated with a 100-μm sieve mesh to remove undigested tissue. The substrates that contained hADSCs were washed twice by PBS, and then resuspended in culture medium, and were lastly seeded at a density of 3 × 106 cells per dish (75 cm2). Cells were cultured at 37°C in a humidified incubator with 5% CO2 and were used in passage 3.
Osteogenic differentiation of hADSCs
For osteogenic differentiation, at 60%–80% confluence, the cells were cultured in high-glucose Dulbecco's modified Eagle's medium containing 10% FBS (Gibco), 0.2 mM ascorbic acid (Sigma), 10 mM β-glycerophosphate (Sigma), and 10 nm dexamethasone (Sigma). The medium was changed every 2 days. The mRNA and protein expression levels of the osteoblast differentiation-related genes were analyzed by real-time PCR (RT-PCR) and western blot, respectively. Early osteogenic differentiation was identified by alkaline phosphatase (ALP) staining and relative ALP activity assays. Alizarin red S staining was used to identify calcium salt deposition in later osteogenesis differentiation.
Relative ALP activity analysis and ALP staining
Cells were lysed with radio immunoprecipitation (RIPA) lysis buffer (Beyotime) with 1 mM phenylmethanesulfonyl fluoride (PMSF) (Beyotime) and were centrifuged at 10,000 rpm 30 min. Then, ∼3–5 μL (∼5–10 μg) of supernatant was added to each hole of a 96-well plate and were incubated with 200 μL of the p-nitrophenylphosphate (pNPP) (Sigma) reagent for 30 min at 37°C, and the photometric value was read at 405 nm using a spectrophotometer. The relative ALP activity was normalized to the total protein of cell lysates.
For the ALP staining, the procedure was performed according to the manufacturer's protocol of ALP staining kit (Institute of Hematology and Blood Diseases Hospital, Chinese Academy of Medical Sciences, Tianjin). After staining, the unbound dye was removed with distilled water and visualized using light microscopy.
Alizarin red S staining and quantification
Cells were washed twice with PBS and fixed with 4% paraformaldehyde for 5 min at room temperature, and then were rinsed with distilled water and stained with 0.5% Alizarin red solution with a pH of 4.2 for 10 min at 37°C. Lastly, unbound dye was removed with distilled water and visualizing using light microscopy and photographing.
For the quantification of Alizarin red S staining, the dye was added with 1 mL 10% (w/v) cetylpyridinium chloride (Sigma) reagent to each hole with shaking for 10 min, then they were transferred to 96-well plates and quantified using a spectrophotometer at 540 nm.
RNA isolation and RT-PCR
Total RNA was extracted with TRIzol reagent (Invitrogen) in accordance with the manufacturer's instruction and RNA was treated with DNase I (Ambion) at 37°C for 30 min. In addition, first-strand cDNA was then synthesized using a high-capacity cDNA reverse transcription kit (Applied Biosystems). The RT-PCRs were performed in ABI Prism 7300 according to manufacturer's procedures (TaKaRa). The amount of 100 ng cDNA was used in each sample. The relative expression level of miRNA or mRNA was evaluated using the 2−ΔΔCt method and were normalized to the expression of U6 or B-ACT, respectively. Primers are shown in Table 1.
Western blot analysis
Protein was extracted in RIPA lysis buffer (Beyotime) with 1 mM PMSF, centrifuged and quantified using a BCA Protein Assay kit (Beyotime), and then denatured in boiling water for 10 min. Supernatant was separated using 10% SDS-PAGE and was transferred to polyvinylidene difluoride membranes (0.22 μm; Millipore). The membranes were blocked with 5% bovine serum albumin for 1 h at room temperature, then were incubated overnight at 4°C with the following antibodies: anti-ALP polyclonal antibody (No. ab108337; Abcam), anti-OPN polyclonal antibody (No. ab69498; Abcam), anti-RUNX2 polyclonal antibody (No. 8486; Cell Signaling Technology), anti-IBSP polyclonal antibody (No. 5468; Cell Signaling Technology), anti-BMPR2 polyclonal antibody (No. 6979; Cell Signaling Technology), anti-OCN polyclonal antibody (No. ab133612; Abcam), anti-BMP3 polyclonal antibody (No. sc-390046; Santa Cruz Biotechnology), and anti-B-ACTIN polyclonal antibody (No. 60008-1-Ig; Proteintech); this incubation was then followed by an incubation with horseradish peroxidase-coupled secondary antibodies for 1 h at room temperature. Protein was detected using a chemiluminescent ECL reagent (Millipore).
Heterotopic bone formation in vivo
Approximately 2 × 106 hADSCs infected with lenti-450b or lenti-negative control (NC) were loaded in hydroxyapatite/tricalcium phosphate (HA/TCP) scaffolds (φ 2–4 × 5 mm, porosity 60% ± 10%, aperture 200–500 μm, National Engineering Research Center for Biomaterials) and were incubated at 37°C overnight, and then implanted subcutaneously into the upper dorsal surface of 6-week-old male NOD/SCID mice (12 mice/group). After 12 weeks, xenografts were harvested and fixed in 4% paraformaldehyde for 24 h, then were decalcified in 10% EDTA for 14 days. The osteoid formation was observed using hematoxylin-eosin (HE) staining, Masson's trichrome staining (Masson) and the Safranin-O/fast green staining (Safranin-O/fast green). All staining procedures were performed according to the manufacturer's protocol from ServiceBio (
Lentiviral vector preparation and infection
An overexpressed miR-450b recombinant lentiviral vector (lenti-450b) was used to upregulate endogenous miR-450b expression and was purchased from GenePharma (
A sponge-450b vector was used to sponge endogenous miR-450b mRNA. The vector was a pLV-CAG-EGFP-2A-mPuro vector and was purchased from SyngenTech (
Plasmid construction
Classic plasmids were constructed by inserting a 90–100 bp synthetic fragment containing the putative miR-450b recognition sequence or its mutated sequence into a psiCHECK-2 vector (Promega), which had Renilla luciferase and firefly luciferase reporter genes.
Another kind of plasmid that was inserted was a 1,000-bp fragment containing the putative miR-450b recognition sequence. The 1,000-bp fragment of 3′-UTR containing the putative miR-450b recognition sequences was generated from hADSCs cDNA using sense and antisense primer PCR. The mutant segments were produced by overlapping PCRs. First, new internal and flanking primers were used to generate overlapping sequences, which were complementary in the intermediate segments. Then, the deleted fragments were obtained by annealing and subsequent PCRs. The wild-type (WT) and mutant fragments were extended to generate new segments, which had XhoI and NotI enzyme sites at both ends, and then were inserted into the psiCHECK-2 plasmid.
Dual luciferase assay
According to the manufacturer's instructions, a total of 5 × 104 293T cells were cotransfected with 50 ng WT or mutant psiCHECK-2 vector and with 100 nM miR-450b mimic or miR-NC by lipofectamine 2000 (Invitrogen). Lysates were harvested at 48 h after the transfection, and the luciferase activity was measured using the Dual Luciferase Reporter Assay System (Promega). The Renilla luciferase activity was normalized to the firefly luciferase activity.
Statistical analysis
In general, Student's t-tests (two-tailed) were performed to analyze the significant differences among the groups. All of the numerical results are presented as the means ± standard deviation from more than three experiments. For multiple-group testing, analysis of variance was conducted followed by a posteriori t-testing. P < 0.05 was considered statistically significant, as indicated by asterisks (*P < 0.05, **P < 0.01, ***P < 0.001).
Results
Expression pattern and conservation analysis of miR-450b
Because hADSCs have the same osteogenic capacity as bone marrow MSCs, and since they are easy to obtain from adipose tissue [8], hADSCs were selected as the seed cells to generate osteoblasts, and they were characterized before being used (Supplementary Fig. S1; Supplementary Data are available online at
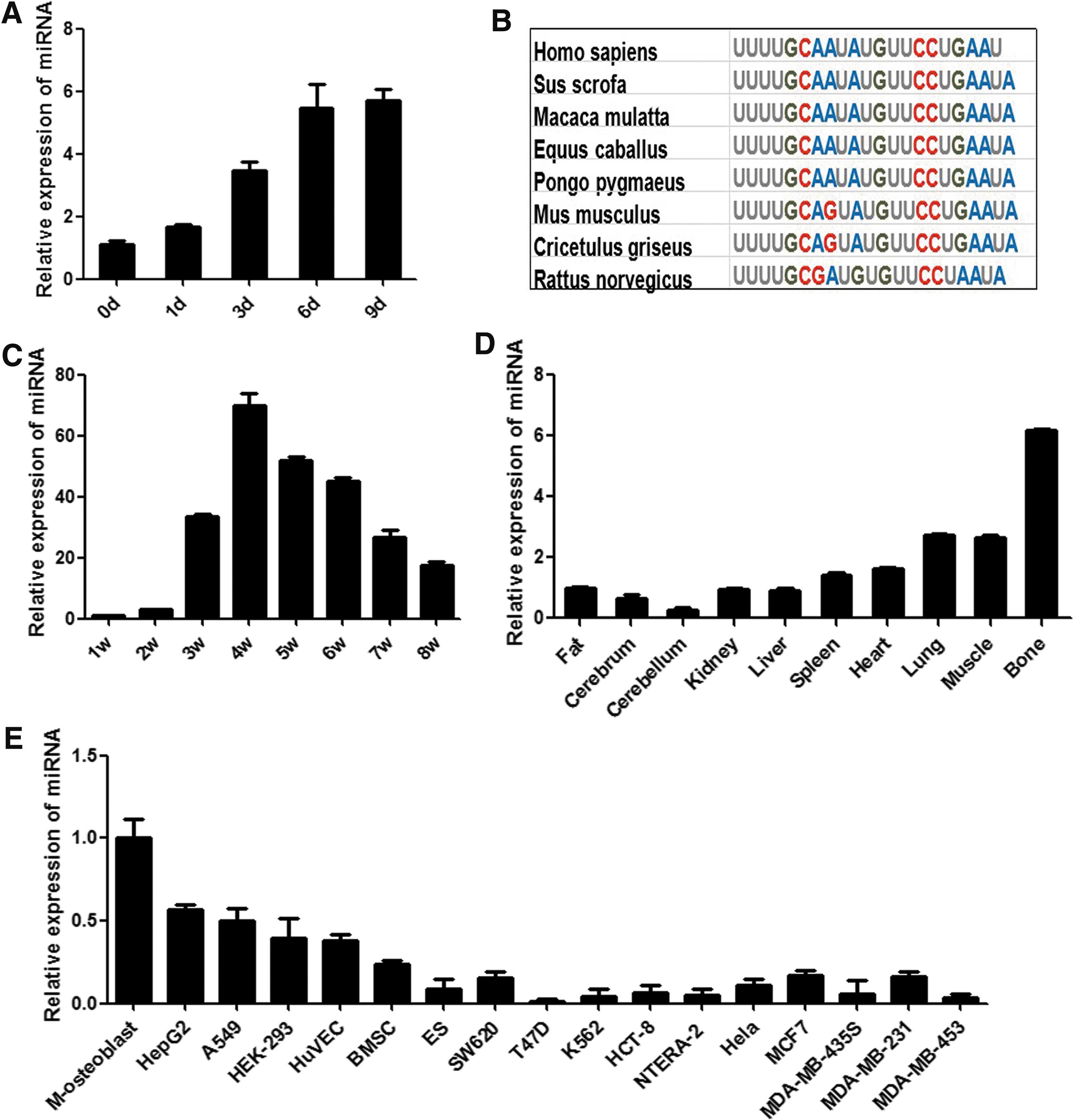
Expression pattern and conservation analysis of miR-450b. The expression level of miR-450b in hADSCs osteogenic differentiation was validated by RT-PCR
Inhibition of endogenous miR-450b delays the osteogenic differentiation of hADSCs
To investigate the effect of miR-450b on the osteogenic differentiation of hADSCs, a specific sponge adsorption of a miR-450b virus vector (sponge-450b) was used to block endogenous miR-450b effects, and the same vector expressing a scrambled sequence (sponge-NC) was used as a control. After sifting by puromycin, GFP-expressing cells were obtained stably. RT-PCR and western blot analysis revealed that osteogenic transcription factors and marker genes RUNX2, OPN, ALP, IBSP, BMPR2, and OCN were significantly decreased in sponge-450b vector-infected cells compared with sponge-NC vector-infected cells (Fig. 2A, B). Accordingly, blocking miR-450b function reduced ALP staining-positive cells and decreased relative ALP activity (Fig. 2C, D). Consistently, Alizarin red S staining on day 12 of osteogenic differentiation showed that the calcium deposition in sponge-450b vector-infected cells was less than that in sponge-NC vector-infected cells, and the Alizarin red S staining quantification also obtained the same result (Fig. 2E, F). Based on these observations, it was suggested that miR-450b may play an important role in the osteogenic differentiation of hADCSs.
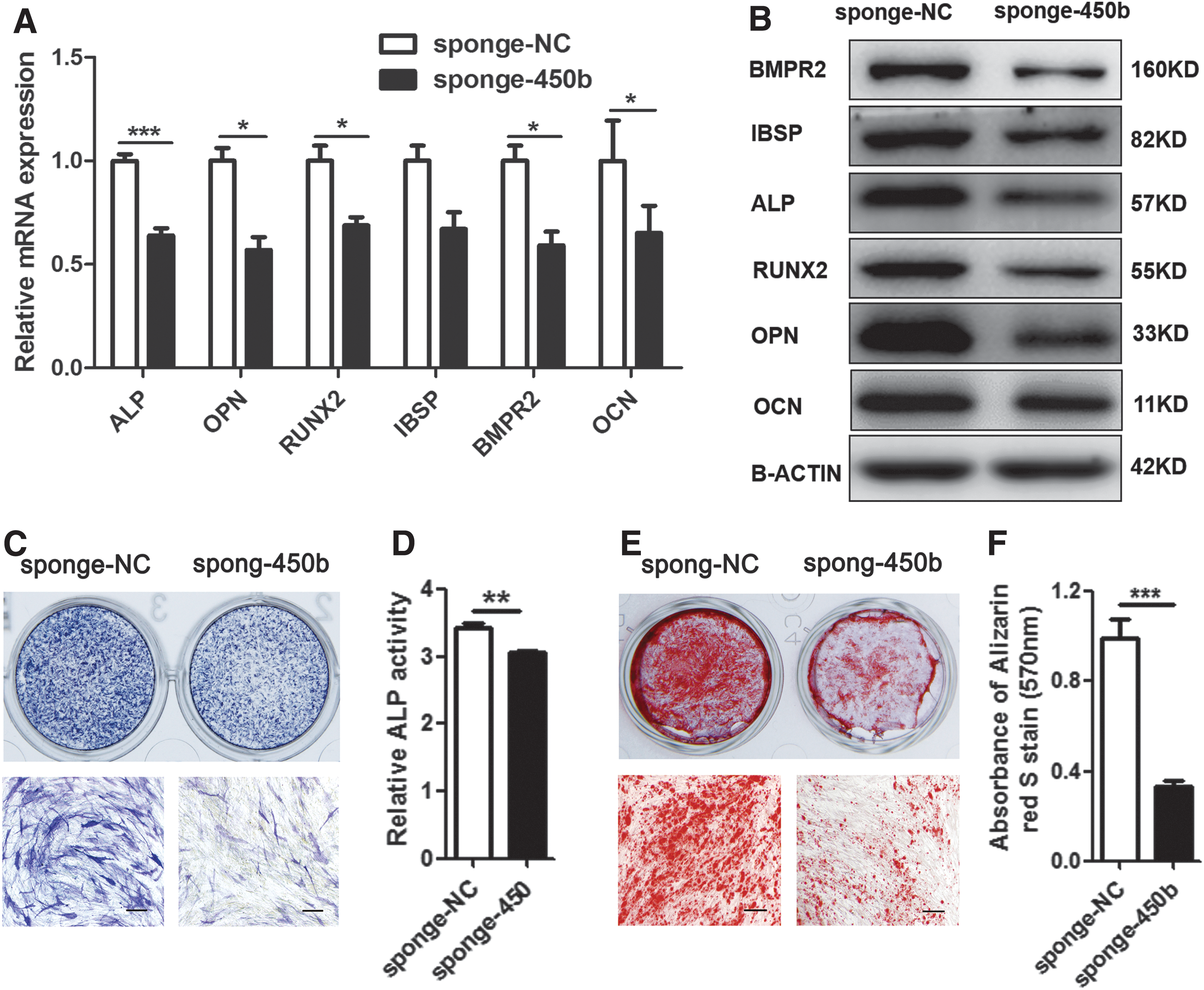
Blocking miR-450b effect suppressed osteogenic differentiation. A specific sponge adsorption of miR-450b (sponge-450b) virus vector was used to block endogeneous miR-450b effects, and the same vector that expressed a scrambled sequence (sponge-NC) was used as a control. RT-PCR and western blot analysis detected the osteogenic transcription factors and marker genes ALP, OPN, RUNX2, IBSP, BMPR2, and OCN expression level on day 6 of osteogenic differentiation
Overexpression of miR-450b promotes osteogenesis of hADSCs in vitro
To further verify the effect of miR-450b on hADSC osteogenic differentiation, we infected hADSCs with miR-450b precursor lentiviral vectors (lenti-450b) or a scrambled sequence control (lenti-NC). Cells were sifted through puromycin until almost all cells were GFP-positive. RT-PCR confirmed that the intracellular miR-450b mRNA levels were significantly increased in lenti-450b-infected cells (Fig. 3A). The osteogenic transcription factors and marker genes RUNX2, OPN, ALP, IBSP, BMPR2, and OCN were significantly upregulated in lenti-450b-infected cells compared with lenti-NC-infected cells, as detected by RT-PCR and western blot analysis (Fig. 3B, C). Moreover, ALP staining on day 6 of osteogenic differentiation increased in lenti-450b-infected cells, and relative ALP activity heightened in lenti-450b-infected cells (Fig. 3D, E). Staining of calcium deposition by Alizarin red S staining on day 12 of osteogenic differentiation and quantification was also increased in lenti-450b-infected cells (Fig. 3F, G).

Upregulation of miR-450b promoted hADSC osteogenic differentiation. An overexpressed miR-450b recombinant lentiviral vector (lenti-450b) was used to upregulate endogenous miR-450b expression, and the same vector that expressed a scrambled sequence (lenti-NC) was used as a control. miRNA-specific RT-PCR detected miR-450b expression level in lenti-450b infected cell and control
Overexpression of miR-450b enhances the heterotopic bone formation of hADSCs in vivo
Our previous research found that the expression of miR-450b also gradually increased with the development of bone in mice, suggesting a positive role of miR-450b in bone formation. To examine whether overexpressing miR-450b could also enhance heterotopic bone formation in vivo, a classic ectopic bone formation model of NOD/SCID mice was used. Cells were infected with lenti-450b or lenti-NC and were induced into osteogenic lineage for 3 days and were then loaded in HA/TCP scaffolds and implanted subcutaneously into the 6-week-old male NOD/SCID mice for 12 weeks (Fig. 4A, B). HE and Safranin-O/fast green staining showed significant increases in the quantity and size of osteoid formation in the xenografts that were incubated with lenti-450b infected cells. Collagen I was an important component of bone formation, which can be stained blue by Masson's trichrome staining (Masson). In the xenografts, which were incubated with lenti-450b-infected cells, the blue dye was also deeper, indicating that more Collagen I emerged (Fig. 4C). We chose sections from another sample and added images as a Supplementary Fig. S2. These data further confirmed that miR-450b is a positive regulator of bone formation.

Upregulation of miR-450b enhanced heterotopic bone formation in vivo. An overexpressed miR-450b recombinant lentiviral vector (lenti-450b) was used to upregulate endogenous miR-450b expression, and the same vector that expressed a scrambled sequence (lenti-NC) was used as a control. Images showed GFP-positive cell proportion in lenti-450b infected cell and control
BMP3, a suppressor of the BMP family, is a direct target of miR-450b
To explore the molecular mechanism of miR-450b promotion of osteogenic differentiation and bone formation, we used various ways to predict potential targets of miR-450b. Bioinformatics analysis using TargetScan, miRBase, and PicTar showed that the osteogenic-related genes ESRRG and BMP3 had miR-450b binding sites in their 3′UTR sequences (Fig. 5A). To further clarify whether miR-450b can combine with the 3′UTR sequences of these genes, we constructed psiCHECK-2 plasmids that had either a WT 3′UTR or a mutant 3′UTR (MUT), with ∼90–100 bp sequences of these genes and cotransfected them with miR-450b mimic (miR-450b) or its NC (miR-NC) into HEK-293T cells. Luciferase assays showed significantly reduced luciferase activity in HEK-293T cells that were cotransfected with the miR-450b mimic and WT ESRRG 3′UTR plasmid or WT BMP3 3′UTR plasmid but not with the MUT 3′UTR plasmid of ESRRG or BMP3 (Fig. 5B).
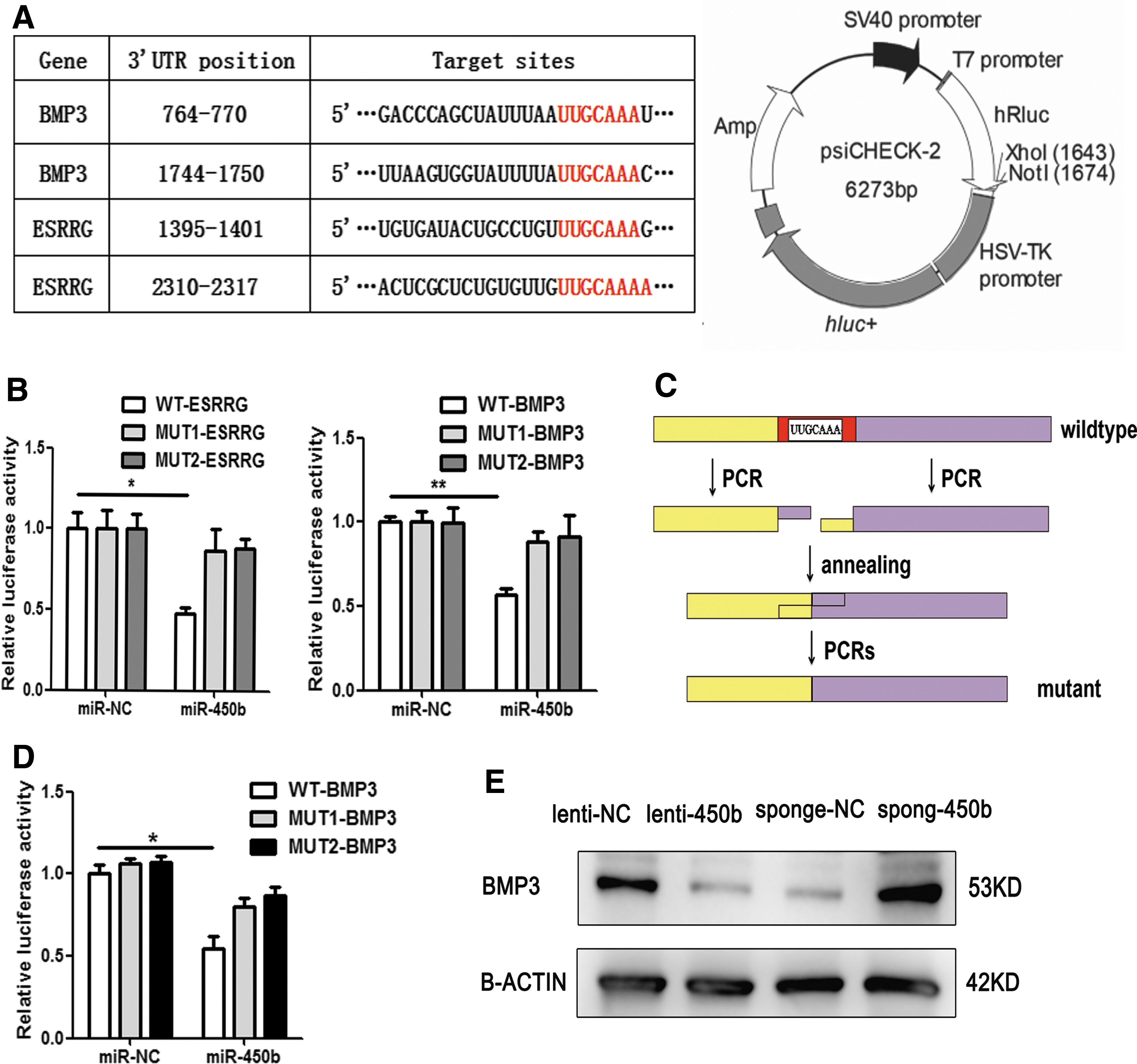
Prediction and validation of the direct targets of miR-450b. Bioinformatic analysis predicted complementary of the miR-450b seed sequences to the 3′UTR of ESRRG and BMP3
Considering the secondary structure of mRNA 3′UTR, we extended the inserted fragments to a length of 1,000 bp length around the binding sites. The WT or mutant fragments were amplified by cDNA PCR and overlapping PCRs (Fig. 5C). In addition, similarly, the 1,000-bp inserted plasmids with miR-450b mimic or NC were cotransfected into HEK-293T cells. Similar results were obtained in the BMP3 3′UTR insertion group but not in the ESRRG 3′UTR insertion group (Fig. 5D). Consistently, the expression of BMP3 protein was markedly downregulated in lenti-450b-infected cells but increased when miR-450b function was blocked (Fig. 5E). Therefore, our findings indicated that BMP3 is a direct target of miR-450b in the regulation of osteogenesis.
Knockdown of endogenous BMP3 can mimic the promoting effect of miR-450b
To further confirm the interaction between miR-450b and BMP3, we chose two short interference RNAs (siRNAs) to inhibit endogenous BMP3 expression in hADSCs and analyzed the effect of BMP3 siRNAs on osteogenic differentiation. RT-PCR verified that both of them sharply decreased mRNA levels of BMP3 compared with the NC (Fig. 6A). As expected, when endogenous BMP3 was suppressed, the osteogenic transcription factors and marker genes RUNX2, OPN, ALP, IBSP, BMPR2, and OCN were significantly upregulated, as indicated by RT-PCR, and western blot analysis (Fig. 6B, C). In addition, the number of ALP-positive cells and the relative ALP activity in BMP3 siRNA-transfected cells were increased, as indicated by ALP staining and an ALP activity assays on day 6 of osteogenic differentiation (Fig. 6D, E). Consistently, the calcium salt precipitation was also enhanced in BMP3-siRNA transfected cells, indicated by Alizarin red S staining on day 12 of osteogenic differentiation (Fig. 6F, G). These results revealed that knockdown of endogenous BMP3 can mimic the promoting effect of miR-450b on osteogenic differentiation of hADSCs, which further confirmed that BMP3 is a direct target of miR-450b in this process.
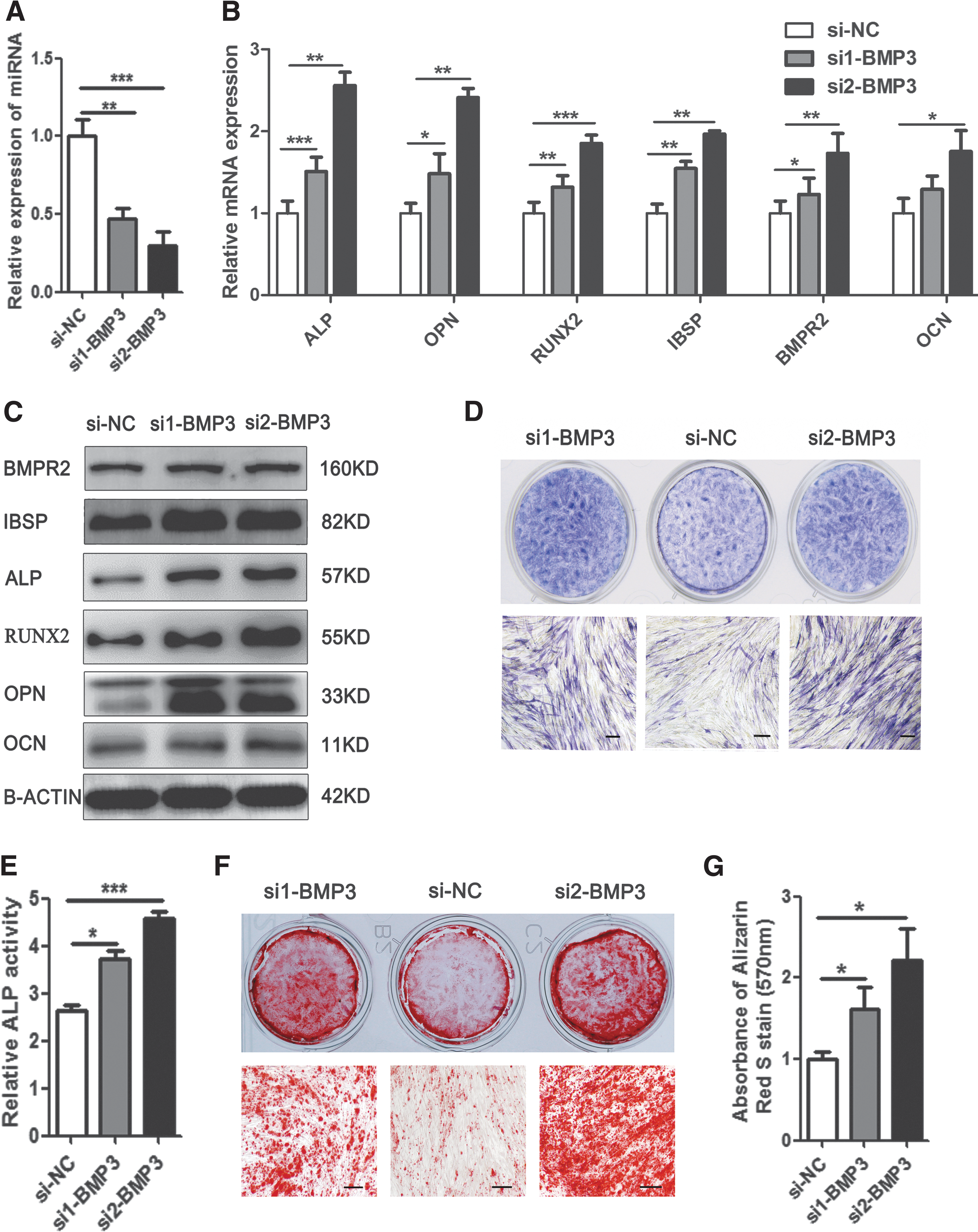
Knockdown of endogeneous BMP3 inhibited osteogenic differentiation of hADSCs. RT-PCR detected the BMP3 mRNA level in siRNAs transfected cell
Discussion
Osteogenesis plays a crucial role in the homeostatic renewal of bone and regenerative fracture healing, while frequently declining in aging organisms, resulting in the loss of bone mass and an increase in fracture incidence [5]. Recently, genetically modified MSC-based tissue engineering therapy has gained more attention in bone tissue regeneration [32,33]. In this study, we found that, as a positive regulator, miR-450b can promote osteogenic differentiation in hADSCs in vitro and can enhance heterotopic bone formation in vivo. Furthermore, we demonstrated that a suppressor of the BMP family, BMP3, is a direct target of miR-450b in the regulation of osteogenesis of hADSCs. Our findings suggested that miR-450b may be a new and attractive osteoinductive candidate for diseases related to bone tissue engineering or osteoblast differentiation.
Bone formation is a progression of osteoblast differentiation and maturation, and osteoblasts and MSCs are the main cells involved in these processes [12,34]. MSCs can be recruited to bone and then differentiate into osteoblasts, thus promoting bone formation. Decreased osteogenic differentiation of MSCs contributes to the development of osteoporosis [35]. For osteoporosis patients who are chronically exposed to glucocorticoids, adipose tissue-derived MSCs show lower ALP activity, leading to a reduced osteogenic differentiation ability [36]. Systemically administered rhBMP-2 could increase MSC numbers and proliferation, thus increasing osteogenic activity and bone formation, and reversing bone and cartilage loss in osteopenic mice [37]. Therefore, looking for more safe and effective ways to improve MSC osteogenesis has attracted much attention in relation to bone regeneration.
miRNA is superior to genome editing in clinical applications because it is not integrated in the genome. miR-450b is a newly identified miRNA that is located in the Xq26 region [38,39]. Currently, there are few reports of its biological functions. Previous studies have demonstrated that miR-450b can inhibit the progression of lung cancer and suppress stemness and the development of chemoresistance by targeting SOX2 in colorectal cancer [39,40]. miR-450b has also been found to inhibit Pax6 expression and to regulate corneal epithelial fate in vitro [38]. In addition to playing an important role in tumor development, miR-450b was found to arrest the growth of rhabdomyosarcomas and to promote their myogenic differentiation by targeting ENOX2 and PAX9 [41]. We found that the expression level of miR-450b was upregulated during osteogenic differentiation of hADSCs. In male C57BL/6J mice with growth and aging, the expression level of miR-450b was dynamic and correlated with bone formation genes in the femur [42], which suggested that miR-450b may be a positive regulator of bone development. To confirm our speculation, we performed a series of gain- and loss-of-function analyses of the process of osteogenic differentiation. We demonstrated that miR-450b not only promoted the process of hADSC differentiation to osteoblasts in vitro but also enhanced ectopic bone formation in vivo. Our findings revealed that miR-450b may become an effective target for specific therapeutic interventions in bone defect repair and fracture healing.
Most miRNAs post-transcriptionally regulate gene expression by either inducing mRNA degradation or by repressing translation through binding to the mRNA 3′-UTR complementary sequences of target genes [15]. Bioinformatic analyses have suggested to us that the osteogenic-related genes SOX2, GSK3B, ESRRG, ACVR2B, BMP3, and JAG1 may be potential targets of miR-450b. Luciferase assays confirmed that miR-450b can bind to the 3′-UTR of BMP3. Furthermore, changes in the expression of miR-450b can regulate the protein level of BMP3 in hADSCs, suggesting that miR-450b can suppress BMP3 translation through binding to the mRNA 3′-UTR complementary sequences.
BMPs are members of the TGF-β superfamily, acting as potent regulators during bone and cartilage formation and embryogenesis [43]. They were first identified as signals that induce ectopic bone formation through effects on skeletal progenitor cells [44]. BMPs, such as BMP2 [37,45,46], BMP4 [47], BMP6 [48], BMP7 [49], and BMP9 [50], have been widely used in bone and cartilage regeneration. The most abundant BMP in bone is BMP3, accounting for >65% of the total BMP stored in bone matrix [51]. BMP3 −/−mice have twice as much trabecular bone as their WT littermates, whereas in transgenic mice, which overexpress BMP3, endochondral bone formation in the developing skeleton is altered and generates spontaneous rib fractures [52]. BMP3 could inhibit BMP2-mediated induction of MSX2 and block BMP2-mediated osteoprogenitor cell differentiation into osteoblasts [53]. In vitro, overexpression of BMP3 suppresses osteoblast differentiation, whereas loss of BMP3 increases osteoblasts and osteocyte colony formation [54]. In this study, we found that miR-450b could target the 3′UTR of BMP3, and miR-450b overexpression could decrease the BMP3 expression at the protein level. As a result of the downregulation of endogenous BMP3 expression in hADSCs, the osteogenic differentiation process was obviously delayed, which was similar to the effect of miR-450b overexpression on osteogenesis.
Collectively, BMP3 is a suppressor member of the BMP family in the regulation of osteogenesis and bone formation. For the first time, we demonstrated that miR-450b can promote osteogenic differentiation in vitro and can enhance bone formation in vivo by directly suppressing BMP3. Previous studies have shown that BMP2 and BMP7 can be used to accelerate bone healing in bone tissue engineering. Using miR-450b to inhibit the expression of endogenous BMP3, thereby promoting bone healing, may be a more effective approach, perhaps at a lower cost, for the treatment of osteoporosis, bone defection, and other bone metabolism-related diseases.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (no. 81700782, 81370466, 81370879, 81473450, 81672313); the National Key Research and Development Program of China (2016YFA0101000, 2016YFA0101003); the key technologies R & D program of Sichuan (2015SZ0020); CAMS Innovation Fund for Medical Sciences (2017-I2 M-3-007, 2017-I2 M-3-006); the Beijing Key Laboratory of New Drug Development and Clinical Trial of Stem Cell Therapy (BZ0381); and undertaken government transfer functions and public service demonstration projects (2017GGFZ001).
Author Disclosure statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.