
Abstract
Voltage-gated sodium and calcium channels as well as transient receptor potential (TRP) channels are expressed during the differentiation of human neural progenitor cells (hNPCs) and are likely to be involved in regulating neurogenesis. However, the molecular composition of these ion channels in proliferating and differentiating hNPCs is largely unknown. In this study, we investigated fetal mesencephalic hNPCs in respect to their sodium, calcium, and TRP channel subunit expression and function. Quantitative real-time polymerase chain reaction indicated a significant upregulation of voltage-gated sodium and calcium channel subunits in hNPCs after differentiation for 3 weeks in vitro. In contrast, the TRP channel expression did not increase significantly during hNPC maturation. Intracellular Ca2+ measurements showed the marked reduction of KCl-induced Ca2+ transients through inhibition of voltage-gated Ca2+ channels by verapamil and mibefradil in differentiated hNPCs. Application of TRP channel agonists induced intracellular Ca2+ peaks already in proliferating hNPCs without affecting their cell division. The coincubation of hNPCs with TRP channel agonists pregnenolone sulfate or RN1747 did not have any significant effect on their proliferation and differentiation. These data indicate that hNPCs derived from fetal midbrain tissue acquire essential voltage-gated sodium and calcium channel properties during neuronal maturation in vitro. An early role of TRP channels in neurogenesis which may be important for regenerative clinical applications or cellular models could not be elucidated using hNPCs.
Introduction
P
Besides the generation of action potentials in neurons, in which the inward current is conducted by voltage-dependent sodium channels, at early stages of neural development (before synapse formation), voltage-gated calcium channels are also of particular importance. Proliferation, migration, ion channel expression, neurotransmitter specification, axon pathfinding, and dendrite outgrowth of human neural progenitor cells (hNPCs) are all, in part, regulated by the expanded repertoire of excitability. Further critical ion channel species are the transient receptor potential (TRP) channels that for example are involved in spontaneous Ca2+ activity during proliferation of hNPCs and generation of growth cone calcium transients [6 –12].
Nine isoforms of the most relevant voltage-gated sodium channel alpha subunits (Nav), Nav1.1 to Nav 1.9, have been identified so far, which differ in their function and tissue presence, for example, action potential forwarding in the CNS or nociception in the peripheral nervous system [13 –15]. In contrast to Nav, voltage-gated calcium channels (Cav) can be detected predominantly in the CNS [16 –18]. To date, the three main families Cav1 (L-type), Cav2 (P/Q-, N-, R-type), and Cav3 (T-type) containing primary alpha1 subunits as well as several auxiliary subunits have been identified [19].
TRP channels are a receptor family with 28 members, spread over several subfamilies: TRPA (ankyrin), TRPC (canonical), TRPM (melastatin), TRPML (mucolipin), TRPN (no mechanoreceptor potential), TRPP (polycystin), and TRPV (vanilloid) [20 –23]. TRP channels have a relatively low selectivity for monovalent and divalent cations and are permeable to Ca2+ in addition to Na+ [24]. TRPC channels have been reported to act as essential cellular sensors in multiple processes during neuronal development, including neural stem cell proliferation and differentiation, neuronal survival, neurite outgrowth and axon path finding, and synaptogenesis [10,25].
Even though ion channel properties of hNPC-derived neurons generated in vitro are of high relevance for preclinical and clinical studies, these investigations are rare. Midbrain-derived hNPCs have the capacity to self-renew and to differentiate into functional (dopaminergic) neurons [27
–31]. Clinical trials will determine if fetal cell-based therapy is a promising treatment for Parkinson's disease (Neural transplantation in the treatment of patients with Parkinson's disease,
Materials and Methods
Cultivation and differentiation of human fetal mesencephalic neural progenitor cells
hNPCs were isolated from aborted fetal midbrain tissue as described previously [26 –30]. The hNPCs were derived from CNS tissue of aborted human fetuses (gestational week 10–16) with mother's consent. Experiments using hNPCs were approved by the Ethics Committees of Hannover Medical School (No. 776) and the University of Leipzig (Reg. No. 066/2002), Germany and are in accordance with all state and federal guidelines.
For subsequent experiments up to six hNPC lines from four different donors were used as indicated. Three independent experiments were mostly performed for each cell line used as stated in the respective figure legends. The cells were cultivated under reduced oxygen (3%) on polyornithine/fibronectin-coated (Sigma-Aldrich) culture flasks in proliferation medium composed of Dulbecco's modified Eagle's medium (DMEM)/Ham's F12, 1xN2, 1xB27, 1% penicillin/streptomycin (all from Life Technologies), 20 ng/mL epidermal growth factor (EGF), and 20 ng/mL fibroblast growth factor 2 (FGF2; both from Peprotech). hNPCs were passaged after reaching confluency using accutase (Life Technologies).
For differentiation, the medium of confluent cultures was replaced by Neurobasal medium containing 2% B27, gentamicin,
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was extracted from proliferating as well as differentiated hNPCs using the RNeasy Mini Kit (Qiagen) and reversely transcribed using oligo-dT primer and Superscript II reverse transcriptase (both from Life Technologies) according to manufacturers' instructions. Quantitative real-time polymerase chain reaction (PCR) experiments were performed as previously described [31].
Complementary DNA (cDNA) from 50 ng total RNA, 1.75 μM forward and reverse primer, and Power SYBR-Green PCR Master Mix (Life Technologies) were used in a StepOnePlus instrument (Applied Biosystems, Darmstadt, Germany) under the following amplification conditions: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, and 60°C for 1 min. The specificity of the PCR products was ensured by melting curve analysis. Equal PCR efficiency of all primer pairs was validated by serial cDNA dilution. Please refer to Stanslowsky et al. [32] for primer sequences of voltage-gated sodium and calcium channel subunits and Table 1 for primer details of TRP channel subunits. Target gene expressions were normalized to the endogenous references β2-microglobulin [Ct (target) − Ct (reference) = ΔCt] and are given as means ± standard errors of the mean (SEM).
F, forward; R, reverse.
Calcium imaging
Measurement of intracellular calcium transients was carried out as described previously [31]. In brief, proliferating or differentiated hNPCs were incubated with Fura 2-AM (Sigma-Aldrich) for 30 min at 37°C. Cytosolic Ca2+ changes upon application of 50 mM KCl were imaged by exciting Fura 2-AM at 340 and 380 nm and monitoring its emission at 510 nm with the Life Acquisition Software (T.I.L.L. Photonics) coupled to an upright microscope (Axioskop 2 FS plus; Zeiss).
To antagonize the KCl-induced Ca2+ influx through voltage-gated calcium channels, the KCl solution was supplemented with the nonselective blockers mibefradil (3 μM) or verapamil (100 μM). TRP channel-mediated Ca2+ flux was initiated by application of 5 μM pregnenolone sulfate (TRPM3 agonist) or 1 μM RN1747 (TRPV4-specific agonist [33]). After background subtraction of individual cells, the 340/380 nm excitation ratio for Fura 2-AM was calculated, which increases as a function of the cytosolic free Ca2+ ([Ca2+]i).
Immunofluorescence
Immunofluorescent stainings were conducted following standard protocols [31]. The applied primary antibodies were mouse anti-Nestin IgG1 (1:500; R&D), rabbit anti-Sox1 IgG (1:500; Millipore), mouse anti-beta III tubulin IgG2 (Tuj1, 1:500; Millipore), rabbit anti-beta III tubulin IgG (1:1,000; Abcam), mouse anti-tyrosine hydroxylase (TH) IgG1 (1:500; Millipore), rabbit polyclonal anti-GABA (1:1,000; Sigma), and rabbit anti-Ki67 IgG (1:1,000; Abcam). Appropriate Alexa Fluor 488 or 555 labeled secondary antibodies (1:500; Life Technologies) were incubated 1 h at room temperature and nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; 10 mg/mL; Life Technologies). Images were taken with an Olympus DP72 camera under an Olympus BX61 fluorescent microscope using the image-analysis software CellF (Olympus).
Coincubation of hNPCs with TRP channel agonists
To investigate the influence of TRP channel agonists on proliferating hNPCs, the undifferentiated cultures were treated with 5 μM pregnenolone sulfate (TRPM3 agonist) or 1 μM RN1747 (TRPV4-specific agonist [33]) for 2 weeks. During differentiation, we treated hNPCs with 5 μM pregnenolone sulfate or 1 μM RN1747 for 3 weeks to analyze their impact on (dopaminergic) neurogenesis.
Appropriate controls without drug treatments were cultured in parallel. The given drug concentrations were chosen because they had no effects on cell viability and still gave a signal in calcium imaging experiments. The drugs were renewed with every media change. The number of viable and dead cells was determined by trypan blue staining during proliferation. The protein content of expanding hNPCs was analyzed using the Pierce™ BCA Protein Assay Kit (Thermo Fisher) according to manufacturer's instructions.
Statistics
Data were analyzed with GraphPad Prism 5 (GraphPad Software). Gene expressions were statistically compared by multiple t-tests (single subunits in undifferentiated vs. differentiated cells) or one-way analysis of variance (ANOVA) with Dunnett's multiple comparison test (all subunits within one group). Calcium transients were statistically analyzed by Wilcoxon test or unpaired t-test as indicated. All data are presented as means ± SEM, and the significance level was set as P < 0.05.
Results
Undifferentiated proliferating as well as differentiated human neural progenitor cells (hNPCs) were investigated by immunofluorescence. Nearly all the proliferating hNPCs stained positive for the neural precursor marker nestin and Sox1. A minor proportion of 8% ± 4% also expressed the early neuronal marker βIII-tubulin (Fig. 1A). hNPCs that had been differentiated for 3 weeks in vitro under standard conditions showed βIII-tubulin expression in 96% ± 3%, whereas only few cells (1.9% ± 0.2%) were positive for the dopaminergic marker TH (Fig. 1B).
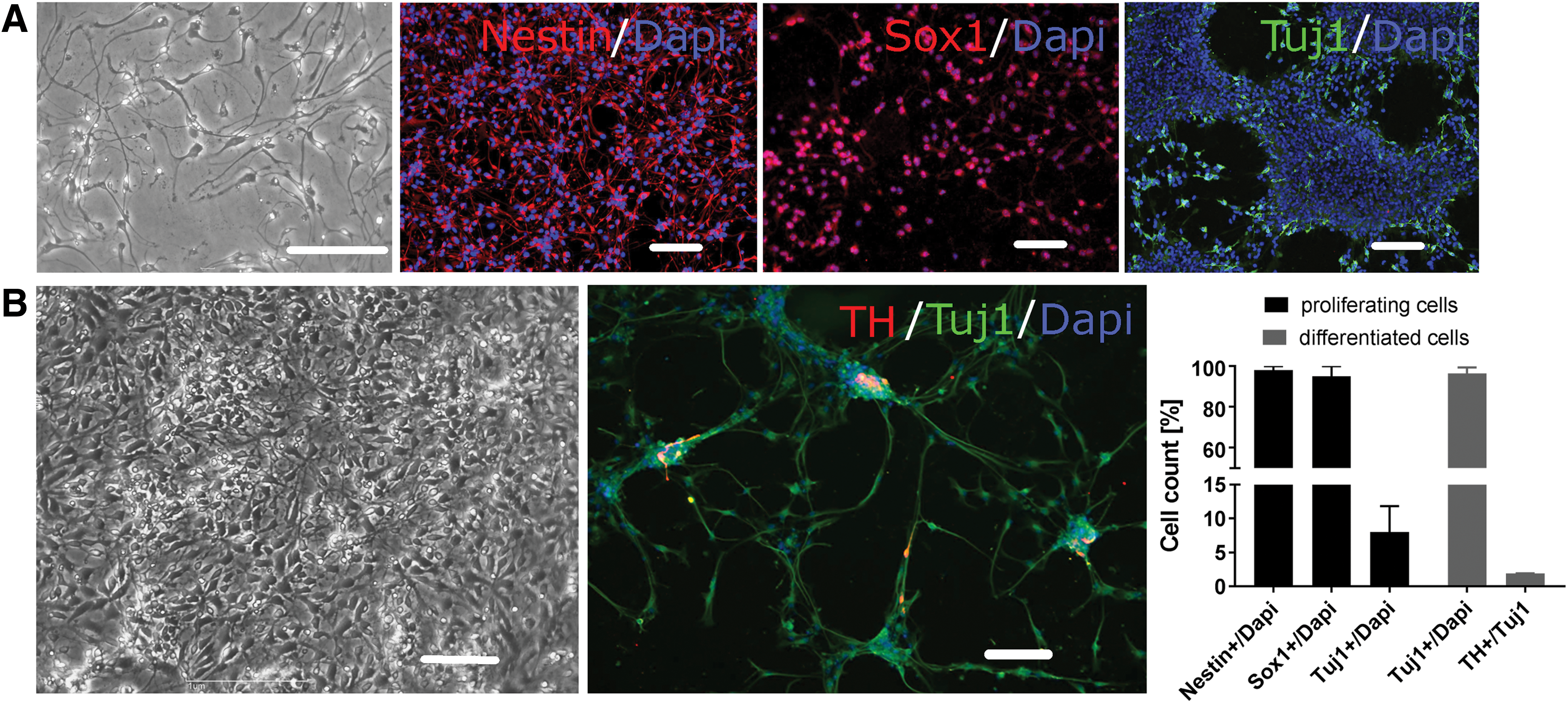
Differentiation of human fetal midbrain-derived neural progenitor cells (hNPCs).
For the functionality of neuronal cells, among others, the expression of voltage-gated sodium and calcium channels is a prerequisite. Therefore, we quantified the expression of sodium and calcium channel subunits in multiple hNPC lines (n = 2–4) by real-time PCR after 3 weeks of differentiation in vitro and compared it to the expression in undifferentiated proliferating hNPCs. Significantly higher 2−ΔCt values in differentiated hNPCs, for nearly all main alpha-subunits and auxiliary subunits investigated, supported the improved maturation status of these hNPCs (Fig. 2A, B, P < 0.05, t-test). In differentiated hNPCs, a predominant expression of single subunits compared to others could not be detected (one-way ANOVA with Dunnett's posttest).
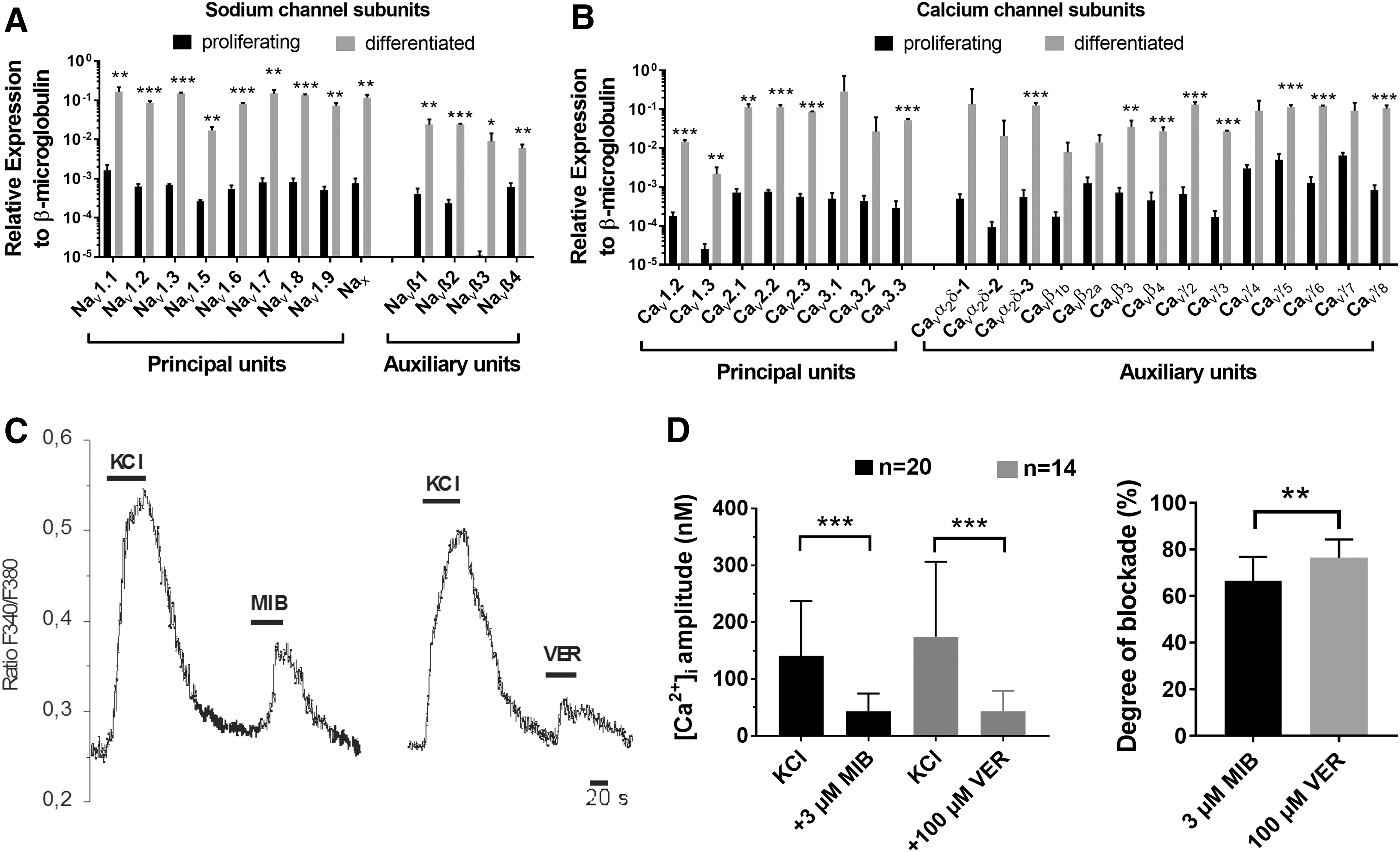
Sodium and calcium channel expression in undifferentiated proliferating hNPCs and after 3 weeks of differentiation.
Next, we determined the functionality of voltage-gated calcium channels by calcium imaging experiments. In differentiated hNPCs, the application of the depolarizing agent KCl (50 mM) induced an increase of intracellular Ca2+ of 155 ± 19 nM (Fig. 2C, D). The KCl-mediated influx was significantly antagonized by simultaneous application of the unspecific voltage-dependent calcium channel blockers mibefradil (3 μM) or verapamil (100 μM), the Ca2+ amplitudes with blockers were only 43 ± 7 nM and 43 ± 10 nM, respectively (P < 0.001, Wilcoxon test). The degree of blockade with verapamil by 77% ± 2% was significantly stronger compared with mibefradil (67% ± 2%) (Fig. 2D, P < 0.01, t-test).
Besides voltage-gated sodium and calcium channels, TRP channels are considered to play a crucial role during neuronal development as well, in part, showing a species-specific activation [34]. Interestingly, real-time PCR experiments revealed no significant differences in subunit expression between proliferating and differentiated hNPC lines (Fig. 3A, n = 5, t-test). Among TRPM channel subunits the expression of TRPM3 was significantly elevated compared with TRPM2 in our proliferating hNPCs (P < 0.05, one-way ANOVA).

TRP channel expression in proliferating hNPCs and after 3 weeks of differentiation.
Ca2+ transients through TRP channels were monitored in our undifferentiated proliferating hNPCs, since they expressed TRP channel subunits to a similar extent as differentiated cells. Upon application of pregnenolone sulfate, among others, an activator of TRPM3 channels and the selective TRPV4 agonist RN1747 [33], the intracellular calcium concentration increased (Fig. 3B). The percentages of cells showing Ca2+ responses caused by application of different agonists were ∼90% (Fig. 3C) (n = 210, t-test). The mean amplitudes represented by the ratio of fluorescence emission at 340 and 380 nm from fura-2-loaded cells were significantly lower after application of pregnenolone sulfate (0.054 ± 0.001) compared to RN1747 (0.074 ± 0.001, P < 0.001, t-test).
Each measurement was terminated by application of the depolarizing agent KCl (50 mM). The lack of KCl-mediated Ca2+ influx supported the notion that none of the proliferating cells functionally expressed voltage-gated sodium or calcium channels (data not shown). Furthermore, ligand-gated ion channels such as NMDA receptors, which could be modulated by pregnenolone sulfate, are not expressed in undifferentiated hNPCs as reported previously [28].
To analyze the effects of TRP channel agonist treatment during proliferation of hNPCs, ∼100,000 cells were plated and coincubated for 2 weeks with the lowest concentrations of pregnenolone sulfate (5 μM) and RN1747 (1 μM) that still evoked signals in hNPCs during Ca2+ imaging experiments. Coincubation, no matter which agonist was used, did not reveal significant changes in proliferation rates, number of living cells or protein content compared with untreated controls (Fig. 4A–C).
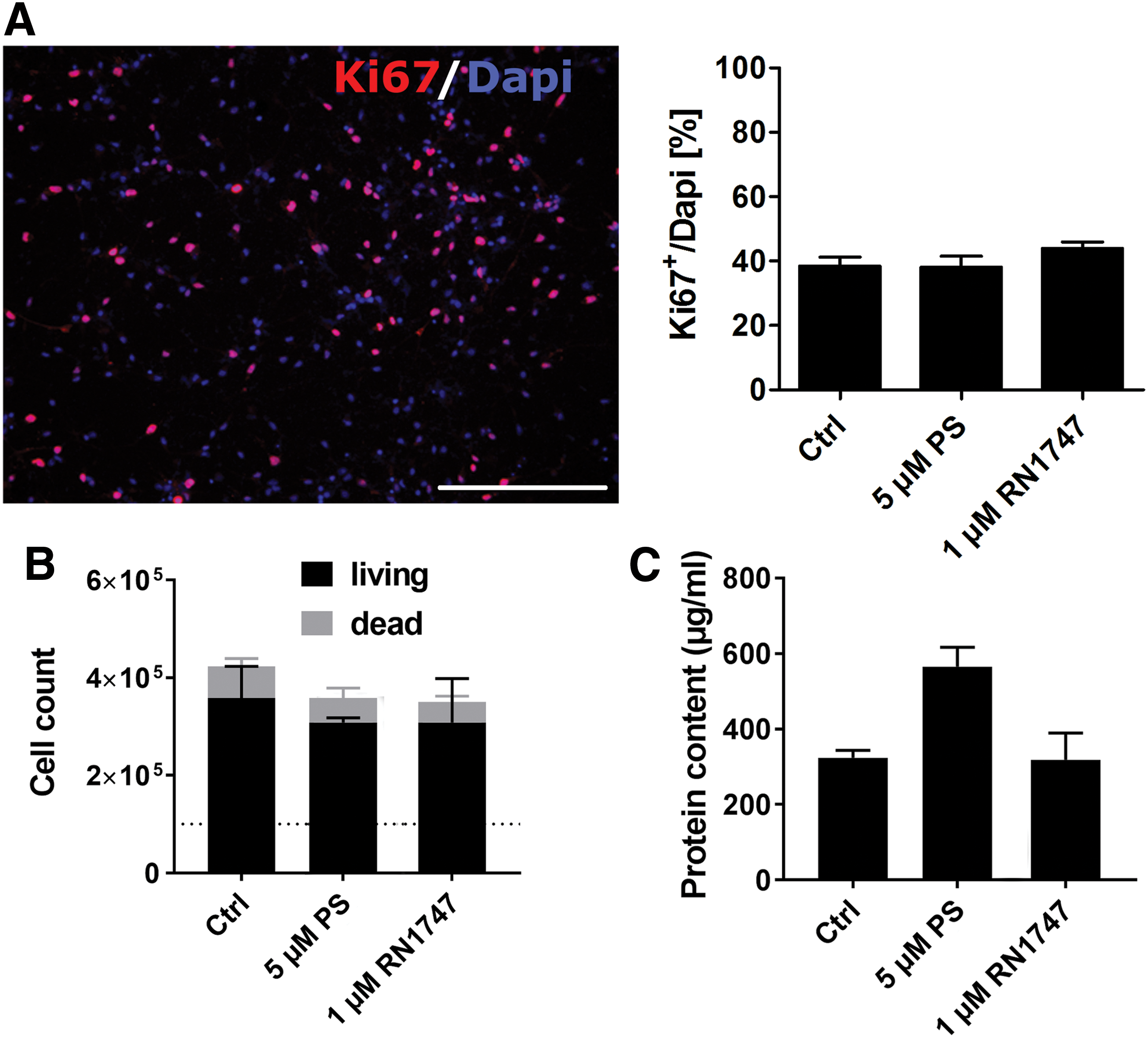
Coincubation of proliferating hNPCs with pregnenolone sulfate or RN1747.
The percentage of cells expressing the proliferation marker Ki67 was around 40% under all conditions tested. The number of cells after 2 weeks of proliferation with or without TRP channel agonists increased threefold. The amount of dead cells (∼16%, n = 3) was not significantly different. Thus, the treatment of undifferentiated fetal mesencephalic hNPCs for 2 weeks during expansion with TRP channel agonists mainly activating TRPM3 (pregnenolone sulfate) and TRPV4 (RN1747) did not enhance cell proliferation or survival rates.
The coincubation of differentiating hNPCs with pregnenolone sulfate (5 μM) or RN1747 (1 μM) for 3 weeks did not have any significant effect on the number of TH- (∼1%/Tuj1) and Tuj1-immunoreactive cells (84%–88%/DAPI) in comparison to untreated controls (Fig. 5A, B). Furthermore, the percentage of cells expressing GABA did not significantly differ between the conditions tested (data not shown). The main neuronal proportion (∼80%) of differentiated NPCs was GABAergic neurons in our human fetal midbrain-derived cultures as shown also in a previous study [27].

Coincubation of differentiating hNPCs with 5 μM PS or 1 μM RN1747 for 3 weeks.
Discussion
In this study, we differentiated human fetal mesencephalic NPCs to neurons, including dopaminergic cells that are mainly affected by neurodegeneration in Parkinson's disease. While the number of βIII-tubulin positive cells in undifferentiated hNPC lines was comparable to a previous study [30], in our differentiated hNPC cultures the amount of βIII-tubulin positive cells was higher and the number of TH immunoreactive dopaminergic neurons lower than reported, suggesting a variability in the maturation potential of fetal midbrain-derived hNPC lines. Other studies using human embryonic stem cells (hESCs) or induced pluripotent stem cells in various neuronal differentiation approaches showed quite varying efficiencies as well, ranging from 20% to almost 90% βIII-tubulin positive cells [31,35 –38] and 3% to 30% dopaminergic neurons [36,39,40].
Human mesencephalic NPCs developed essential neuronal properties such as tetrodotoxin-sensitive sodium channels and action potentials as well as functional voltage-dependent calcium channels after maturation for 3 weeks [27,28]. However, systematic studies on the genomic expression of voltage-gated sodium, calcium, or TRP channels in human stem cell-derived neural precursors are rare. In contrast to our hNPCs, in differentiated hESC-derived neural progenitors, an upregulation of sodium and calcium channel subunits compared to undifferentiated progenitors could not be detected [41]. Another study, focusing on a limited number of ion channels, reported the significant upregulation of Nav1.1, Nav1.2, and Cav2.2 in neuron-like cells generated from hESCs [42].
The functional expression of voltage-gated sodium channels is a sign of neuronal maturation and necessary for action potential firing. In the CNS Nav1.1, Nav1.2, Nav1.3, and Nav1.6 are highly but not exclusively expressed [43], whereas Nav1.7, Nav1.8, and Nav1.9 are more restricted to peripheral neurons [44]. In our hNPCs, nearly all channel subunits were significantly upregulated after differentiation, but no predominance of certain subunits was detected, indicating maturation without strict regional specification.
The only calcium channels that were not significantly upregulated upon neuronal differentiation were the T-type channels Cav3.1 and Cav3.2. This is in line with developmental studies revealing high expression levels of T-type calcium channels in early stages that are exceeded by the expression of N or L-type currents during maturation of the nervous system [45]. These quantitative real-time PCR data of voltage-dependent sodium and calcium channels suggest that human fetal NPCs and neural progenitors derived from other sources may have varying expression profiles of some main subunits.
The KCl-evoked intracellular Ca2+ increases, with amplitudes comparable to a previous study [28], were mainly mediated by functional voltage-gated Ca2+ channels. This is in line with a previous investigation using differentiated hNPCs, where the coapplication of verapamil induced an ∼80% degree of blockade [28]. Despite its potent and nonselective calcium channel antagonism, verapamil is not known to be embryo- or fetotoxic, therefore, this blocker could be used as treatment to achieve a marked inhibition of voltage-gated calcium channels in hNPC, which may become relevant in future cell transplantations and medical therapies in Parkinson's disease patients [46].
Different to the late expression of voltage-gated Na+ and Ca2+ channels during neuronal development, TRP channels are already functionally expressed in neural progenitor cells, a relatively early stage in neurogenesis. Upon differentiation, the expression profile of TRPs did not change significantly. In a study with ESC-derived hNPCs, numerous TRP channels (eg, TRPV4) were even expressed at lower levels after differentiation [41]. Our quantitative PCR data indicated an upregulation of TRPM3 in respect to TRPM2 in proliferating NPCs. While it was reported that activation of TRPM2 can lead to apoptosis and neuronal death and inhibition has a neuroprotective role [47], little is known about the role of TRPM3 in brain physiology. There is evidence that TRPM3 activation facilitates glutamatergic transmission in the developing brain [48].
In line with previous pharmacological characterizations, our data of Ca2+ increase upon application of pregnenolone sulfate or RN1747—respective TRPM3 and TRPV4 agonists—indicated that in proliferating hNPCs, most activity is the result of TRP channels representing the main route of Ca2+ entry into these cells [12]. TRP-mediated increase in calcium entry is even required for the proliferation of NPCs [25,49].
TRP channels promote many developmental processes, including proliferation and survival, which are of particular importance for various stem or progenitor cell sources intended for cell replacement therapies [12]. But there is growing evidence that TRPs can also induce neuronal degeneration in numerous disorders, including Parkinson's disease. TRPC3 for example is associated with abnormalities in GABAergic neuron firing in the substantia nigra by controlling GABA projection neuron activity [50]. Pharmacological modulation of TRP channels in stem cell-derived progenitor cells may reduce unwanted proliferation and tumor formation that still challenge cell replacement therapy [26].
Conclusions
These data indicate that hNPCs derived from fetal midbrain tissue expressed essential voltage-gated calcium and sodium channels during neuronal differentiation. In contrast, the TRP channel expression did not enhance significantly during hNPC maturation. The activation of predominantly expressed TRP channels (TRPM3, TRPV4) by treatment with agonists did not have a functional impact on the proliferation, survival, and differentiation of hNPCs in vitro.
Footnotes
Acknowledgments
The authors thank Mrs. Annett Brandt, Mrs. Ute Römuß, and Mr. Andreas Niesel for excellent technical assistance. This study was supported, in part, by the Petermax-Müller-Stiftung for Parkinson research and the Karlheinz-Hartmann-Stiftung (Hannover, Germany).
Author Disclosure Statement
The authors declare that there is no conflict of interest regarding the publication of this article.